
Morphology and Phylogeny Reveal Vamsapriyaceae fam. nov. (Xylariales, Sordariomycetes) with Two Novel Vamsapriya Species

 ,
,  ,
, 
Abstract
1. Introduction
2. Materials and Methods
2.1. Collection, Examination, Isolation and Conservation
2.2. DNA Extraction, PCR Amplification and Sequencing
2.3. Phylogenetic Analyses
3. Results
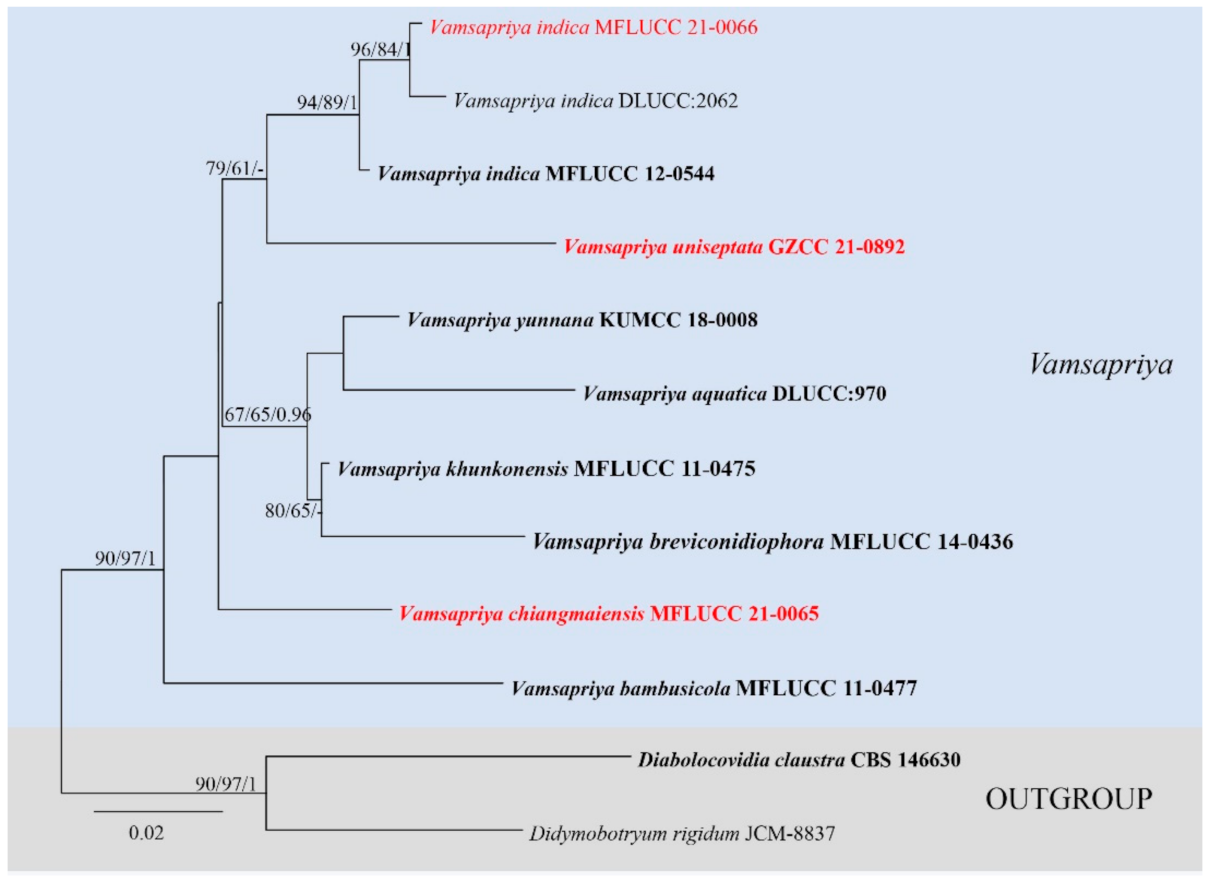
3.1. Phylogenetic Analyses
3.2. Taxonomy
3.3. Other Accepted Genera in Vamsapriyaceae
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Eriksson, O.E.; Winka, K. Supraordinal taxa of Ascomycota. Myconet 1997, 1, 1–16. [Google Scholar]
- Smith, G.J.D.; Liew, E.C.; Hyde, K.D. The Xylariales: A monophyletic order containing 7 families. Fungal Diversity 2003, 13, 185–218. [Google Scholar]
- Stadler, M. Importance of secondary metabolites in the Xylariaceae as parameters for assessment of their taxonomy, phylogeny, and functional biodiversity. Curr. Res. Environ. Appl. Mycol. 2011, 1, 75–133. [Google Scholar] [CrossRef]
- U’Ren, J.M.; Miadlikowska, J.; Zimmerman, N.B.; Lutzoni, F.; Stajich, J.E.; Arnold, A.E. Contributions of North American endophytes to the phylogeny, ecology, and taxonomy of Xylariaceae (Sordariomycetes, Ascomycota). Mol. Phylogenet. Evol. 2016, 98, 210–232. [Google Scholar] [CrossRef] [PubMed]
- Helaly, S.E.; Thongbai, B.; Stadler, M. Diversity of biologically active secondary metabolites from endophytic and saprotrophic fungi of the ascomycete order Xylariales. Nat. Product Rep. 2018, 35, 992–1014. [Google Scholar] [CrossRef]
- Becker, K.; Stadler, M. Recent progress in biodiversity research on the Xylariales and their secondary metabolism. J. Antibiot. 2021, 74, 1–23. [Google Scholar] [CrossRef]
- Nannfeldt, J.A. Studien uber die Morphologie und Systematik der nicht-lichenisierten inoperculaten Discomyceten. Nova Acta Regiae Societatis Scientiarum Upsaliensis Ser. IV 1932, 8, 1–368. [Google Scholar]
- Munk, A. The system of the Pyrenomycetes. Dansk Botanisk Arkin 1953, 15, 1–163. [Google Scholar]
- Müller, E.; von Arx, J.A. Die Gattungen der didymosporen Pyrenomyceten. Beiträge Kryptogamenflora Schweiz 1962, 11, 1–922. [Google Scholar]
- Muller, E. Pyrenomycetes: Meliolales, coronophorales, sphaeriales. In The Fungi, an Advanced Treatise; Academic Press: London, UK, 1973. [Google Scholar]
- Wehmeyer, L.E. The Pyrenomycetous fungi. Mycol. Mem. 1975, 6, 1–250. [Google Scholar]
- Barr, M.E. Prodromus to nonlichenized, pyrenomycetous members of class Hymenoascomycetes. Mycotaxon 1990, 39, 43–184. [Google Scholar]
- Hawksworth, D.L.; Kirk, P.M.; Sutton, B.C.; Pegler, D.N. Ainsworth and Bisby’s Dictionary of the Fungi, 8th ed.; CAB International: Oxon, UK, 1995. [Google Scholar]
- Kang, J.C.; Kong, R.Y.C.; Hyde, K.D. Studies on the Amphisphaeriales 1. Amhisphaeriaceae (sensu stricto) and its phylogenetic relationships inferred from 5.8S rDNA and ITS2 sequences. Fungal Divers. 1998, 1, 147–157. [Google Scholar]
- Kang, J.C.; Kong, R.Y.C.; Hyde, K.D. Phylogeny of Amhisphaeriaceae (sensu stricto) and related taxa revisited based on nrDNA sequences. Mycotaxon 2002, 81, 321–330. [Google Scholar]
- Eriksson, O.E.; Baral, H.O.; Currah, R.S.; Hansen, K.; Kurtzman, C.P.; Rambold, G.; Laessoe, T. Outline of Ascomycota—2003. Myconet 2003, 9, 1–89. [Google Scholar]
- Lumbsch, H.T.; Huhndorf, S.M. Myconet volume 14 part one. Outline of ascomycota—2009. Fieldiana Life Earth Sci. 2010, 1, 1–922. [Google Scholar] [CrossRef]
- Senanayake, I.C.; Maharachchikumbura, S.S.N.; Hyde, K.D.; Bhat, J.D.; Jones, E.B.G.; McKenzie, E.H.C.; Dai, D.Q.; Daranagama, D.A.; Dayarathne, M.C.; Goonasekara, I.D.; et al. Towards unraveling relationships in Xylariomycetidae (Sordariomycetes). Fungal Divers. 2015, 73, 73–144. [Google Scholar] [CrossRef]
- Hyde, K.D.; Norphanphoun, C.; Maharachchikumbura, S.S.N.; Bhat, D.J.; Jones, E.B.G.; Bundhun, D.; Chen, Y.J.; Bao, D.F.; Boonmee, S.; Calabon, M.S.; et al. Refined families of Sordariomycetes. Mycosphere 2020, 11, 305–1059. [Google Scholar] [CrossRef]
- Hyde, K.D.; Dong, Y.; Phookamsak, R.; Jeewon, R.; Bhat, D.J.; Jones, E.B.G.; Liu, N.G.; Abeywickrama, P.D.; Mapook, A.; Wei, D.P.; et al. Fungal diversity notes 1151–1276: Taxonomic and phylogenetic contributions on genera and species of fungal taxa. Fungal Divers. 2020, 100, 5–277. [Google Scholar] [CrossRef]
- Maharachchikumbura, S.S.N.; Hyde, K.D.; Jones, E.B.G.; McKenzie, E.H.C.; Bhat, J.D.; Dayarathne, M.C.; Huang, S.K.; Norphanphoun, C.; Senanayake, I.C.; Perera, R.H.; et al. Families of Sordariomycetes. Fungal Divers. 2016, 79, 1–317. [Google Scholar] [CrossRef]
- Fröhlich, J.; Hyde, K.D. Palm Microfungi. Fungal Divers. Res. Ser. 2000, 3, 1–393. [Google Scholar]
- Daranagama, D.A.; Camporesi, E.; Liu, X.Z.; Bhat, D.J.; Chamyuang, S.; Bahkali, A.H.; Stadler, M.; Hyde, K.D. Tristratiperidium microsporum gen. et sp. nov. (Xylariales) on dead leaves of Arundo plinii. Mycol. Prog. 2015, 15, 1–8. [Google Scholar] [CrossRef]
- Crous, P.W.; Wingfield, M.J.; Burgess, T.I.; Hardy, G.E.S.J.; Crane, C.; Barrett, S.; Cano-Lira, J.F.; le Roux, J.J.; Thangavel, R.; Guarro, J.; et al. Fungal Planet description sheetS: 469–557. Persoonia 2016, 37, 218–403. [Google Scholar] [CrossRef]
- Fiuza, P.O.; Silva, C.; Santos, T.A.B.; Raja, H.; Castañeda-Ruiz, R.F.; Gusmão, L.F.P. Roselymyces, a new asexual genus of the Xylariales (Ascomycota) from Brazil. Sydowia 2018, 70, 59–65. [Google Scholar]
- Gawas, P.; Bhat, D.J. Vamsapriya indica gen. et sp. nov., a bambusicolous, synnematous fungus from India. Mycotaxon 2005, 94, 149–154. [Google Scholar]
- Dai, D.Q.; Bahkali, A.H.; Li, Q.R.; Bhat, D.J.; Wijayawardene, N.N.; Li, W.J.; Chukeatirote, E.; Zhao, R.L.; Xu, J.C.; Hyde, K.D. Vamsapriya (Xylariaceae) redescribed, with two new species and molecular sequence data. Cryptogamie Mycologie 2014, 35, 339–357. [Google Scholar] [CrossRef]
- Dai, D.Q.; Phookamsak, R.; Wijayawardene, N.N.; Li, W.J.; Bhat, D.J.; Xu, J.C.; Taylor, J.E.; Hyde, K.D.; Chukeatirote, E. Bambusicolous fungi. Fungal Divers. 2017, 82, 1–105. [Google Scholar] [CrossRef]
- Castañeda-Ruiz, R.F.; Zhang, X.G.; Li, D.W.; Gusmão, L.F.P.; Pérez-Martínez, S.; Sosa, D. Notes on Vamsapriya and V. camagueyensis comb. nov. Mycotaxon 2017, 132, 553–557. [Google Scholar] [CrossRef]
- Hyde, K.D.; Norphanphoun, C.; Abreu, V.P.; Bazzicalupo, A.; Thilini, C.K.W.; Clericuzio, M.; Dayarathne, M.C.; Dissanayake, A.J.; Ekanayaka, A.H.; He, M.Q.; et al. Fungal diversity notes 603–708: Taxonomic and phylogenetic notes on genera and species. Fungal Divers. 2017, 87, 1–235. [Google Scholar] [CrossRef]
- Jiang, H.B.; Phookamsak, R.; Bhat, D.J.; Khan, S.; Bahkali, A.; Elgorban, A.M.; Hyde, K.D. Vamsapriya yunnana, a new species of Vamsapriya (Xylariaceae; Xylariales) associated with bamboo from Yunnan; China. Phytotaxa 2018, 356, 61–70. [Google Scholar] [CrossRef]
- Bao, D.F.; Hyde, K.D.; McKenzie, E.H.; Jeewon, R.; Su, H.Y.; Nalumpang, S.; Luo, Z.L. Biodiversity of Lignicolous Freshwater Hyphomycetes from China and Thailand and Description of Sixteen Species. J. Fungi 2021, 7, 669. [Google Scholar] [CrossRef]
- Ling, Y.; Li, H.H.; Xia, J.W.; Zhang, X.G.; Li, Z. Vamsapriya jinniuensis sp. nov.; and a first record of Garnaudia elegans from southern China. Mycotaxon 2018, 133, 367–372. [Google Scholar] [CrossRef]
- Samarakoon, M.C.; Thongbai, B.; Hyde, K.D.; Brönstrup, M.; Beutling, U.; Lambert, C.; Miller, A.N.; Liu, J.K.; Promputtha, I.; Stadler, M. Elucidation of the life cycle of the endophytic genus Muscodor and its transfer to Induratia in Induratiaceae fam. nov.; based on a polyphasic taxonomic approach. Fungal Divers. 2020, 101, 177–210. [Google Scholar] [CrossRef]
- Senanayake, I.; Rathnayake, A.; Marasinghe, D.; Calabon, M.; Gentekaki, E.; Lee, H.B.; Hurdeal, V.G.; Pem, D.; Dissanayake, L.S.; Wijesinghe, S.N.; et al. Morphological approaches in studying fungi: Collection; Examination; Isolation; Sporulation and Preservation. Mycosphere 2020, 11, 2678–2754. [Google Scholar] [CrossRef]
- Jayasiri, S.C.; Hyde, K.D.; Ariyawansa, H.A.; Bha, T.J.; Buyck, B.; Cai, L.; Dai, Y.C.; Abd-Elsalam, K.A.; Ertz, D.; Hidayat, I.; et al. The Faces of Fungi database: Fungal names linked with morphology; Phylogeny and human impacts. Fungal Divers. 2015, 74, 3–18. [Google Scholar] [CrossRef]
- Index Fungorum. Available online: http://www.indexfungorum.org/Names/Names.asp (accessed on 6 September 2021).
- Vilgalys, R.; Hester, M. Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several Cryptococcus species. J. Bacteriol. 1990, 172, 4238–4246. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. PCR Protoc. Guide Methods Appl. 1990, 18, 315–322. [Google Scholar]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef]
- Larsson, A. AliView: A fast and lightweight alignment viewer and editor for large datasets. Bioinformatics 2014, 30, 3276–3278. [Google Scholar] [CrossRef]
- Vaidya, G.; Lohman, D.J.; Meier, R. SequenceMatrix: Concatenation software for the fast assembly of multi-gene datasets with character set and codon information. Cladistics 2011, 27, 171–180. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef]
- Nylander, J. MrModeltest (Version 2.2); Evolutionary Biology Centre, Uppsala University: Uppsala, Sweden, 2004. [Google Scholar]
- Trifinopoulos, J.; Nguyen, L.-T.; von Haeseler, A.; Minh, B.Q. W-IQ-TREE: A fast online phylogenetic tool for maximum likelihood analysis. Nucleic Acids Res. 2016, 44, W232–W235. [Google Scholar] [CrossRef]
- Swofford, D.L. PAUP*: Phylogenetic Analysis Using Parsimony (*and Other Methods); Version 4.0 b10; Sinauer Associates: Sunderland, MA, USA, 2002. [Google Scholar]
- Hillis, D.M.; Bull, J.J. An empirical test of bootstrapping as a method for assessing confidence in phylogenetic analysis. Syst. Biol. 1993, 42, 182–192. [Google Scholar] [CrossRef]
- Rambaut, A. FigTree. Version 1.4.2; University of Edinburgh: Edinburgh, UK, 2014. [Google Scholar]
- Crous, P.W.; Wingfield, M.J.; Chooi, Y.H.; Gilchrist, C.L.; Lacey, E.; Pitt, J.I.; Roets, F.; Swart, W.J.; Cano-Lira, J.F.; Valenzuela-Lopez, N. Fungal Planet description sheets:1042–1111. Pers. Mol. Phylogeny Evol. Fungi 2020, 44, 301. [Google Scholar] [CrossRef]
- Seifert, K. Contributions toward a mycobiota of Indonesia: Synnematous hyphomycetes. Mem. N. Y. Bot. Gard 1990, 59, 109–154. [Google Scholar]
- Zhang, Y.D.; Ma, J.; Ma, L.G.; Zhang, X.G. A new species of Podosporium and a new record from southern China. Mycotaxon 2010, 114, 401–405. [Google Scholar] [CrossRef]
- Wang, J.Y.; Zhang, K.; Yang, C.L.; Xia, J.W.; Ma, Y.R.; Gao, J.M.; Li, X.Y.; Zhang, X.G.; Cai, Y.M. Podosporium bacilliforme sp. nov. and a first record of Phaeoblastophora peckii from southern China. Mycotaxon 2016, 131, 841–846. [Google Scholar] [CrossRef]
- Seifert, K.; Morgan-Jones, G.; Gams, W.; Kendrick, B. The Genera of Hyphomycetes; CBS-KNA W Fungal Biodiversity Centre: Utrecht, The Netherlands, 2011. [Google Scholar]
- Jeewon, R.; Hyde, K.D. Establishing species boundaries and new taxa among fungi: Recommendations to resolve taxonomic ambiguities. Mycosphere 2016, 7, 1669–1677. [Google Scholar] [CrossRef]
- Saccardo, P.A. Sylloge Hyphomycetum. Sylloge Fungorum 1886, 4, 807. [Google Scholar]
- Berkeley, M.J.; Broome, C.E. Enumeration of the fungi of Ceylon. Part II; containing the remainder of the hymenomycetes; with the remaining established tribes of fungi. J. Linn. Soc. Bot. 1873, 14, 29–140. [Google Scholar] [CrossRef]
- Ellis, M.B. Dematiaceous Hyphomycetes; Commonwealth Mycological Institute: Kew, UK, 1971. [Google Scholar]
- Zhuang, W.Y. Higher Fungi of Tropical China; Mycotaxon Ltd.: Ithaca, NY, USA, 2001. [Google Scholar]
- Wang, X.M.; Liu, G.N.; Chen, S.S.; Du, X.F. Records of Beltrania rhombica and Didymobotryum rigidum from China. Mycotaxon 2017, 132, 525–529. [Google Scholar] [CrossRef]
- Von Schweinitz, L.D. Synopsis fungorum in America boreali media degentium. Trans. Am. Philos. Soc. 1832, 4, 141–316. [Google Scholar] [CrossRef]
- Ciferri, R. Observations on meliolicolous hyphales from Santo Domingo. Sydowia 1955, 9, 302. [Google Scholar]
- Mercado, S. Nuevos e interesantes hifomicetes enterblásticos de Cuba. Acta Bot. Cuba 1983, 16, 1–8. [Google Scholar]
- Crous, P.W.; Wingfield, M.J.; Kendrick, W.B. Foliicolous dematiaceous hyphomycetes from Syzygium cordatum. Can. J. Bot. 1995, 73, 224–234. [Google Scholar] [CrossRef][Green Version]
- Mycobank. Available online: https://www.mycobank.org/ (accessed on 6 September 2021).
- Yang, H.; Dong, W.; Yu, X.D.; Bhat, D.J.; Boonmee, S.; Zhang, H. Four freshwater dematiaceous hyphomycetes in Sordariomycetes with two new species of Parafuscosporella. Phytotaxa 2020, 44, 19–34. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Locus | Primers | PCR Procedure | Reference |
|---|---|---|---|
| LSU | LR0R | 94 °C 3 min; 35 cycles of 94 °C 30 s, 52 °C 30 s, 72 °C 1 min; 72 °C 8 min; 4 °C on hold | [38,39] |
| LR5 | |||
| ITS | ITS5 | ||
| ITS4 |
| Taxa | Strain Numbers | ITS | LSU | rpb2 | tub2 |
|---|---|---|---|---|---|
| Amphirosellinia fushanensis | HAST 91111209 | GU339496 | N/A | GQ848339 | GQ495950 |
| Amphirosellinia nigrospora | HAST 91092308 | GU322457 | N/A | GQ848340 | GQ495951 |
| Amphisphaeria sorbi | MFLUCC 13-0721 | NR_153531 | KP744475 | N/A | N/A |
| Amphisphaeria thailandica | MFLU 18-0794 | NR_168783 | NG_068588 | MK033640 | MK033639 |
| Anthostomella formosa | MFLUCC 14-0170 | KP297403 | KP340544 | KP340531 | KP406614 |
| Anthostomella obesa | MFLUCC 14-0171 | KP297405 | KP340546 | KP340533 | KP406616 |
| Anthostomelloides krabiensis | MFLUCC 15-0678 | KX305927 | KX305928 | KX305929 | N/A |
| Barrmaelia rhamnicola | CBS 142772 | NR_153497 | N/A | MF488999 | MF489018 |
| Barrmaelia rappazii | CBS 142771 | NR_153496 | N/A | MF488998 | MF489017 |
| Barrmaelia macrospora | CBS 142768 | NR_167684 | N/A | MF488995 | MF489014 |
| Biscogniauxia arima | WSP 122 | NR_167683 | N/A | GQ304736 | AY951672 |
| Biscogniauxia mangiferae | MFLU 18-0827 | MN337232 | MN336236 | MN366247 | MN509782 |
| Biscogniauxia nummularia | MUCL 51395 | NR_153649 | NG_066378 | KY624236 | KX271241 |
| Biscogniauxia repanda | ATCC 62606 | KY610383 | KY610428 | N/A | KX271242 |
| Brunneiperidium gracilentum | MFLUCC 14-0011 | KP297400 | KP340542 | KP340528 | KP406611 |
| Brunneiperidium involucratum | MFLUCC 14-0009 | KP297399 | KP340541 | KP340527 | KP406610 |
| Cainia anthoxanthis | MFLUCC 15-0539 | NR_138407 | NG_070382 | N/A | N/A |
| Cainia graminis | MFLUCC 15-0540 | KR092793 | KR092781 | N/A | N/A |
| Cainia desmazieri | CBS 137.62 | MH858124 | MH869702 | N/A | N/A |
| Clypeosphaeria mamillana | CBS 140735 | N/A | NG_067338 | MF489001 | MH704637 |
| Collodiscula bambusae | GZUH 0102 | KP054279 | KP054280 | KP276675 | N/A |
| Collodiscula japonica | CBS 124266 | JF440974 | JF440974 | KY624273 | KY624316 |
| Collodiscula fangjingshanensis | GZUH0109 | KR002590 | KR002591 | KR002592 | KR002589 |
| Collodiscula leigongshanensis | GZUH0107 | KP054281 | KP054282 | KR002588 | KR002587 |
| Coniocessia anandra | CBS 125766 | MH863747 | MH875215 | N/A | N/A |
| Coniocessia maxima | CBS 593.74 | NR_137751 | NG_070051 | N/A | N/A |
| Coniocessia cruciformis | CBS 126674 | MH864206 | MH875663 | N/A | N/A |
| Coniolariella limonispora | CBS 283.64 | KF719198 | KF719210 | N/A | N/A |
| Coniolariella gamsii | CBS 114379 | GU553325 | GU553329 | N/A | N/A |
| Daldinia macaronesica | CBS 113040 | JX658504 | KY610477 | KY624294 | KX271266 |
| Daldinia loculatoides | CBS 113279 | MH862918 | MH874491 | KY624247 | KX271246 |
| Diabolocovidia claustri | CBS 146630 | MT373367 | MT373350 | N/A | N/A |
| Diatrypella heveae | MFLUCC 17-0368 | NR_154046 | NG_069531 | N/A | MG334557 |
| Diatrypella tectonae | MFLUCC 12-0172 | NR_154029 | NG_069423 | N/A | KY421043 |
| Didymobotryum rigidum | JCM 8837 | LC228650 | LC228707 | N/A | N/A |
| Entosordaria quercina | CBS 142774 | NR_153499 | N/A | MF489004 | MF489022 |
| Entosordaria perfidiosa | CBS 142773 | NR_153498 | N/A | MF489003 | MF489021 |
| Eutypa linearis | MFLUCC 11-0503 | KU940150 | KU863138 | N/A | N/A |
| Fasciatispora arengae | MFLUCC 15-0326b | MK120301 | MK120276 | N/A | N/A |
| Fasciatispora arengae | MFLUCC 15-0326c | MK120302 | MK120277 | N/A | N/A |
| Graphostroma platystomum | CBS 270.87 | JX658535 | N/A | N/A | HG934108 |
| Hansfordia pulvinata | CBS 254.59 | KF893288 | MH869394 | N/A | N/A |
| Hansfordia pulvinata | CBS 144422 | MK442587 | MK442527 | N/A | N/A |
| Hansfordia pruni | CBS 125767 | MH863748 | MH875216 | N/A | N/A |
| Hansfordia pruni | CBS 276.51 | MH856854 | MH868374 | N/A | N/A |
| Hypoxylon fragiforme | MUCL 51264 | KC477229 | NG_066364 | N/A | KX271282 |
| Hypoxylon neosublenormandii | MFLUCC 11-0618 | NR_155174 | NG_066168 | N/A | N/A |
| Induratia sp. | SMH1255 | MN250031 | AY780069 | N/A | AY780119 |
| Induratia fengyangensis | CBS 126601 | HM034852 | HM034858 | HM034847 | HM034839 |
| Induratia ziziphi | MFLUCC 17-2662 | MK762705 | MK762712 | MK791281 | MK776958 |
| Induratia thailandica | MFLUCC 17-2669 | MK762707 | MK762714 | MK791283 | MK776960 |
| Lopadostoma dryophilum | CBS 133213 | NR_132028 | N/A | KC774526 | MF489023 |
| Lopadostoma americanum | CBS 133211 | NR_132027 | N/A | N/A | N/A |
| Lopadostoma fagi | CBS 133206 | NR_132029 | N/A | KC774531 | N/A |
| Lopadostoma turgidum | CBS 133207 | NR_132036 | N/A | KC774562 | MF489024 |
| Microdochium lycopodinum | CBS 125585 | NR_145223 | KP858952 | KP859125 | KP859080 |
| Microdochium phragmitis | CBS 285.71 | NR_132916 | NG_058147 | KP859122 | KP859077 |
| Nemania abortiva | WSP 71221 | GU292816 | N/A | GQ844768 | GQ470219 |
| Nemania beaumontii | HAST 405 | GU292819 | N/A | GQ844772 | GQ470222 |
| Nemania bipapillata | HAST 90080610 | GU292818 | N/A | GQ844771 | GQ470221 |
| Nemania primolutea | HAST 91102001 | EF026121 | N/A | GQ844767 | EF025607 |
| Podosordaria mexicana | WSP 176 | GU324762 | N/A | GQ853039 | GQ844840 |
| Podosordaria muli | WSP 167 | GU324761 | N/A | GQ853038 | GQ844839 |
| Poronia pileiformis | WSP 88113001 | GU324760 | GQ853037 | GQ502720 | |
| Poronia punctata | CBS 656.78 | KT281904 | KY610496 | KY624278 | KX271281 |
| Requienella fraxini | CBS 140475 | NR_138415 | N/A | N/A | N/A |
| Requienella seminuda | CBS 140502 | NR_154630 | MH878683 | MK523300 | N/A |
| Rosellinia buxi | JDR-99 | GU300070 | N/A | GQ844780 | GQ470228 |
| Rosellinia necatrix | HAST 89062904 | EF026117 | KF719204 | GQ844779 | EF025603 |
| Vamsapriya aquatica | DLUCC:970 | MZ420740 | N/A | N/A | N/A |
| Vamsapriya bambusicola | MFLUCC 11-0477 | KM462835 | KM462836 | KM462834 | KM462833 |
| Vamsapriya breviconidiophora | MFLUCC 14-0436 | MF621584 | MF621588 | N/A | N/A |
| Vamsapriya chiangmaiensis | MFLUCC 21-0065 | MZ613171 | MZ613168 | N/A | N/A |
| Vamsapriya indica | MFLUCC 12-0544 | KM462839 | KM462840 | KM462841 | KM462838 |
| Vamsapriya indica | DLUCC: 2062 | MZ420747 | MZ420762 | MZ442699 | N/A |
| Vamsapriya indica | MFLUCC 21-0066 | MZ613172 | MZ613169 | OK560921 | N/A |
| Vamsapriya khunkonensis | MFLUCC 13-0497 | KM462830 | KM462831 | KM462829 | KM462828 |
| Vamsapriya uniseptata | GZCC 21-0892 | MZ613173 | MZ613170 | N/A | N/A |
| Vamsapriya yunnana | KUMCC 18-0008 | MG833874 | MG833873 | MG833875 | N/A |
| Xylaria arbuscula | CBS 126415 | KY610394 | KY610463 | KY624287 | KX271257 |
| Xylaria bambusicola | MFLUCC 11-0606 | KU940160 | KU863148 | KU940183 | N/A |
| Xylaria hypoxylon | CBS122620 | AM993141 | KM186301 | KM186302 | KX271279 |
| Zygosporium oscheoides | MFLUCC 14-0402 | MF621585 | MF621589 | N/A | N/A |
| Zygosporium minus | HKAS99625 | MF621586 | MF621590 | N/A | N/A |
| Taxon | Host | Habitat/Location | Synnemata | Conidiogenous Cells | Conidia |
|---|---|---|---|---|---|
| V. indica (M 393674) | Bamboo | Terrestrial/India | 700–1100 μm long, 60–160 μm wide at the base, 30–60 μm wide in the middle, 30–80 μm wide at the apical fertile region | Monotretic, clavate, dark brown, 4–9 × 2.5–4.5 μm | Catenate, acrogenous, brown, cylindrical, vermiform, 10–80 × 4–6 μm, 2–12-septate |
| V. indica (MFLU 13-0370) | Bamboo | Terrestrial/Thailand | 700–1100 μm long, 60–160 μm wide at the base, 30–60 μm wide in the middle, 30–80 μm wide at the apical fertile region | Monotretic, ellipsoidal, brown to dark brown, 4–9 × 2.5–4.5 μm ( = 6.5 × 3.7 μm, n = 20) | Catenate, cylindrical, pale brown to dark brown 35–290 × 4–6.5 μm ( = 66.6 × 5.6 μm, n = 20), 1–3-septate when young, more than 20–septate at maturity |
| V. indica (HKAS 115803) | Bamboo | Freshwater/China | 1145–1475 μm long, 105–235 μm wide at the base, 50–80 μm wide in the middle, 70–155 μm wide at the apical fertile region | Monotretic, clavate, dark brown 5–9 × 3–5 μm ( = 20 × 5 μm, n = 30) | Catenate, brown to dark brown, cylindrical to obclavate, 15–30 × 4–6.5 μm ( = 20 × 5 μm, n = 30), 1–4-septate |
| V. indica (MFLU 21-0088) | Bamboo | Terrestrial/Thailand | 1300–1900 μm long, 80–150 μm wide at the base, 30–40 μm wide in the middle, 60–140 μm wide at the apical fertile region | Cylindrical to clavate, brown, 4.5–8.5 × 3–4.5 μm ( = 6.5 × 4 μm, n = 30) | Catenate, olivaceous brown to brown, cylindrical, 20–48 × 4.5–6.5 μm ( = 32 × 5.5 μm, n = 20), 2–8-septate |
| Species | ITS | ||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 42 | 52 | 73 | 74 | 82 | 106 | 127 | 167 | 171 | 173 | 194 | 196 | 203 | 206 | 207 | 208 | 209 | 213 | 214 | |
| V. bambusicola (MFLUCC 11-0477) | C | T | C | G | T | T | A | A | C | C | C | G | C | C | T | C | T | A | A |
| V. chiangmaiensis (MFLUCC 21-0065) | T | C | G | T | C | C | G | C | T | T | G | A | T | A | C | T | C | T | T |
| 216 | 221 | 229 | 232 | 233 | 235 | 239 | 421 | 432 | 442 | 446 | 447 | 448 | 451 | 461 | 465 | 557 | 558 | ||
| V. bambusicola (MFLUCC 11-0477) | T | A | A | C | T | T | G | T | C | T | C | T | C | C | T | G | T | T | |
| V. chiangmaiensis (MFLUCC 21-0065) | A | G | T | T | C | C | A | C | T | C | T | G | T | T | C | A | C | A | |
| Family | Genus | Asexual Morph | ||
|---|---|---|---|---|
| Synnemata | Conidiogenous Cells | Conidia | ||
| Vamsapriyaceae | Diabolocovidia | Absent | Monoblastic, subcylindrical to clavate, pale brown | Catenate, acrogenous, brown, ellipsoid to obovoid, unseptate |
| Didymobotryum | Present | Monotretic, integrated, terminal, clavate to cylindrical, pale brown to brown | Catenate, olivaceous brown to brown, cylindrical | |
| Podosporium | Present | Mono- or polytretic, subulate or cylindrical, darkly pigmented | Solitary, obclavate or bacilliform, multi-septate, brown to dark brown | |
| Tretophragmia | Present | Monotretic, subulate or cylindrical, darkly pigmented | Solitary, obclavate to fusiform or irregular in shape, multi-septate, dark brown | |
| Vamsapriya | Present | Monotretic, clavate to cylindrical | Catenate or solitary, acrogenous, cylindrical, oblong, fusiform or obclavate, brown to dark brown, septate | |
| Induratiaceae | Emarcea | Absent | Integrated, terminal, pale brown, forming a rachis with numerous small, pimple-like denticles | Hyaline, smooth, falcate, granular, apex subobtuse, base truncate |
| Induratia | Absent | Terminal, solitary or sometimes two celled at the ends of branches, cylindrical, pale brown, bearing inconspicuous denticles | Narrowly ellipsoidal to subglobose, hyaline | |
| Family | Genus | Sexual Morph | |
|---|---|---|---|
| Asci | Ascospores | ||
| Vamsapriyaceae | Vamsapriya | 8-spored, unitunicate, cylindrical, short pedicellate, with J+ apical ring | Apiosporous, fusiform to broad fusiform, hyaline, with sheath |
| Induratiaceae | Emarcea | 8-spored, unitunicate, cylindrical, pedicellate, with J+ ring | Overlapping uniseriate, long fusiform, hyaline, 2-celled |
| Induratia | 8-spored, unitunicate, cylindrical, short pedicellate, with a J+ apical ring | Uniseriate, naviculate to ellipsoidal, mostly hyaline, constricted apiosporous | |
| Clypeosphaeriaceae | Aquasphaeria | 8-spored, unitunicate, cylindrical, with J- apical ring | Biseriate, cylindrical and ovoid, hyaline |
| Apioclypea | 8-spored, pedunculate, cylindrical, fissitunicate | Biseriate, fusiform, hyaline, with sheath | |
| Brunneiapiospora | 8-spored, unitunicate, cylindrical, pedicellate with J+ or J− ascal ring | Hyaline to light brown apiospores with a mucilaginous sheath | |
| Clypeosphaeria | 8-spored, unitunicate, cylindrical to broadly cylindrical, pedicellate, with J+ or J− ascal ring | Ellipsoidal to fusiform, unicellular to septate, hyaline to dark brown ascospores, sometimes with sheaths or appendages | |
| Crassoascus | 8-spored, unitunicate, cylindrical, short pedicellate, with J+ ring | Bright brown to dark brown, multiseptate, fusiform ascospores, with hyaline refractive cap-like appendages at each end | |
| Palmaria | Cylindric to clavate, with a J− subapical ring | Apiosporous, hyaline, 1-septate, obclavate, with a mucilaginous sheath | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, Y.-R.; Liu, N.-G.; Samarakoon, M.C.; Jayawardena, R.S.; Hyde, K.D.; Wang, Y. Morphology and Phylogeny Reveal Vamsapriyaceae fam. nov. (Xylariales, Sordariomycetes) with Two Novel Vamsapriya Species. J. Fungi 2021, 7, 891. https://doi.org/10.3390/jof7110891
Sun Y-R, Liu N-G, Samarakoon MC, Jayawardena RS, Hyde KD, Wang Y. Morphology and Phylogeny Reveal Vamsapriyaceae fam. nov. (Xylariales, Sordariomycetes) with Two Novel Vamsapriya Species. Journal of Fungi. 2021; 7(11):891. https://doi.org/10.3390/jof7110891
Chicago/Turabian StyleSun, Ya-Ru, Ning-Guo Liu, Milan C. Samarakoon, Ruvishika S. Jayawardena, Kevin D. Hyde, and Yong Wang. 2021. "Morphology and Phylogeny Reveal Vamsapriyaceae fam. nov. (Xylariales, Sordariomycetes) with Two Novel Vamsapriya Species" Journal of Fungi 7, no. 11: 891. https://doi.org/10.3390/jof7110891
APA StyleSun, Y.-R., Liu, N.-G., Samarakoon, M. C., Jayawardena, R. S., Hyde, K. D., & Wang, Y. (2021). Morphology and Phylogeny Reveal Vamsapriyaceae fam. nov. (Xylariales, Sordariomycetes) with Two Novel Vamsapriya Species. Journal of Fungi, 7(11), 891. https://doi.org/10.3390/jof7110891