Updates on the Taxonomy of Mucorales with an Emphasis on Clinically Important Taxa

Abstract
1. Introduction
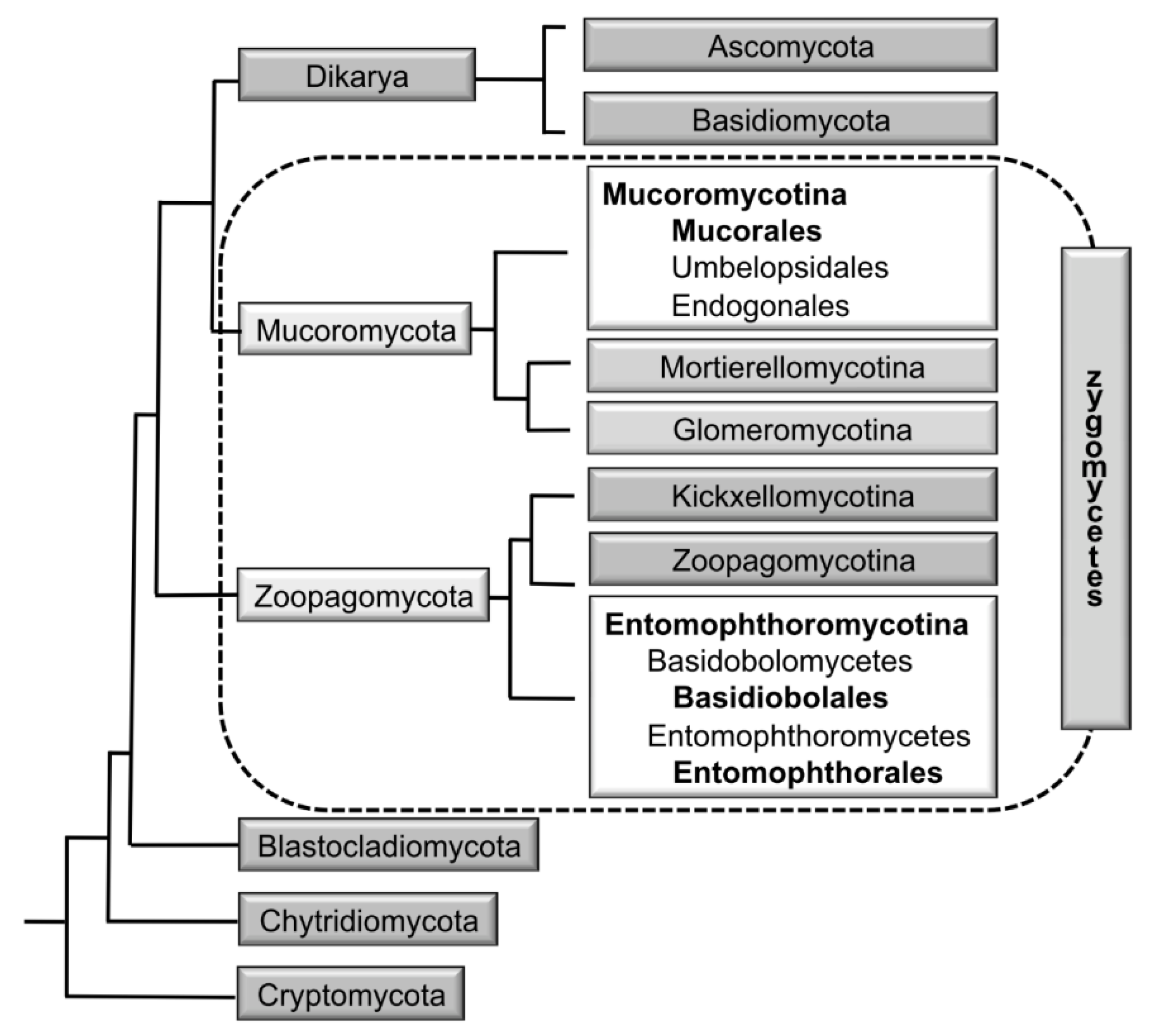
2. Higher-Level Classification and Delimitation of the Mucorales
3. Lower-Level Classification: Molecular Taxonomy and Re-Evaluated Criteria for Genus and Species Delimitation
4. Mucoralean Genera and Species with Changed Taxonomic Concepts
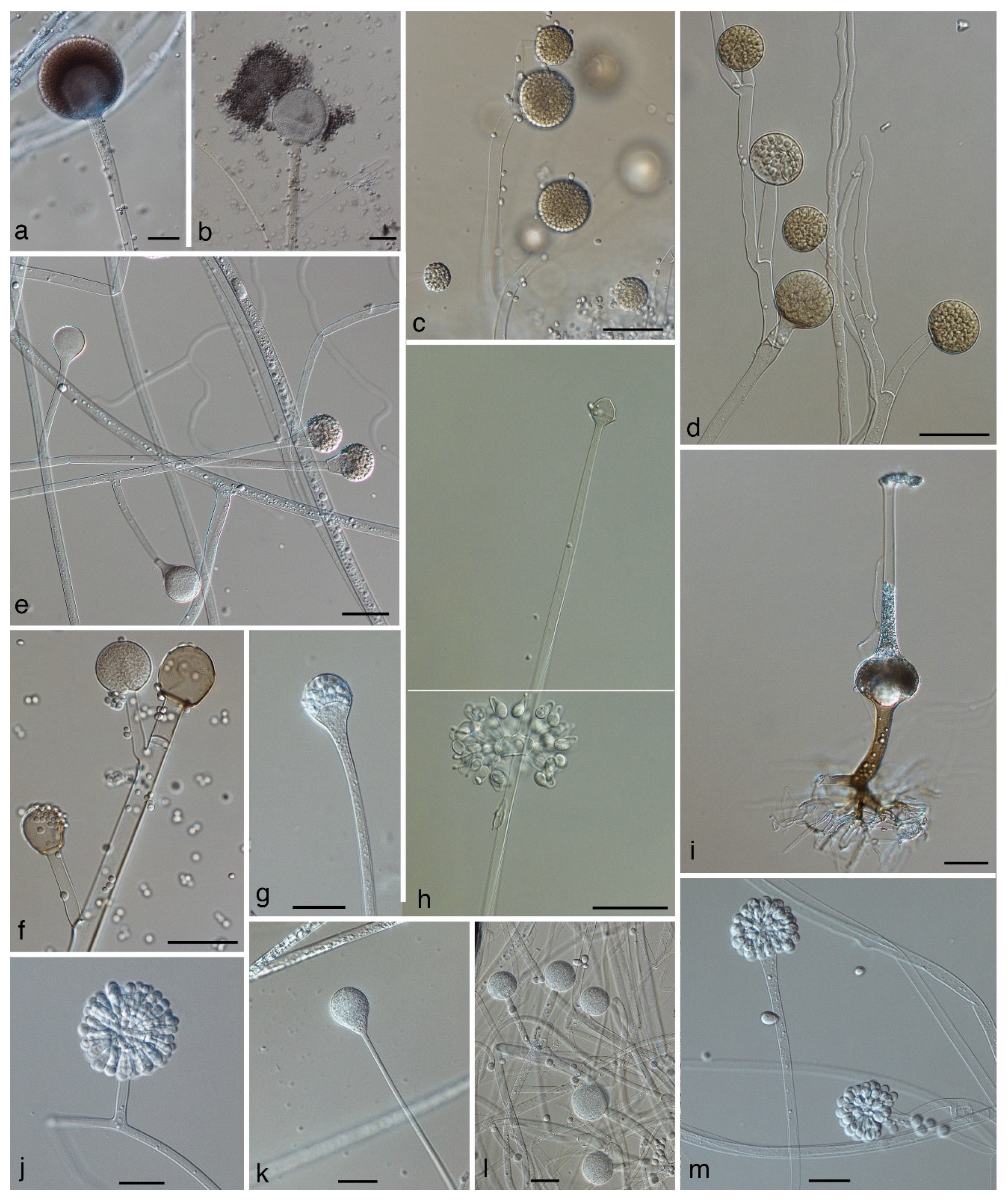
4.1. Absidia, Lentamyces, and Lichtheimia (Figure 3e)
4.2. Actinomucor elegans (Figure 3l)
4.3. Backusella (Figure 1d–f)
4.4. Circinella (Figure 1j)
4.5. Mucor (Including Zygorhynchus) (Figure 3c,d)
4.6. Rhizomucor (Figure 3f)
4.7. Rhizopus (Figure 3a,b)
4.7.1. Rhizopus microsporus
4.7.2. Rhizopus arrhizus (syn. R. oryzae) (Figure 3a,b)
5. Identification and Detection
6. Ecology and Geographic Distribution
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Benny, G.L.; Humber, R.A.; Voigt, K. Zygomycetous Fungi: Phylum Entomophthoromycota and Subphyla Kickxellomycotina, Mortierellomycotina, Mucoromycotina, and Zoopagomycotina. In The Mycota (A Comprehensive Treatise on Fungi as Experimental Systems for Basic and Applied Research); Springer: Berlin/Heidelberg, Germany, 2014; Volume 7A, pp. 209–250. [Google Scholar]
- Morin-Sardin, S.; Nodet, P.; Coton, E.; Jany, J.-L. Mucor: A Janus-faced fungal genus with human health impact and industrial applications. Fungal Biol. Rev. 2017, 31, 12–32. [Google Scholar] [CrossRef]
- Roden, M.M.; Zaoutis, T.E.; Buchanan, W.L.; Knudsen, T.A.; Sarkisova, T.A.; Schaufele, R.L.; Sein, M.; Sein, T.; Chiou, C.C.; Chu, J.H.; et al. Epidemiology and outcome of zygomycosis: A review of 929 reported cases. Clin. Infect. Dis. 2005, 41, 634–653. [Google Scholar] [CrossRef] [PubMed]
- De Hoog, G.S.; Guarro, J.; Gené, J.; Figueras, M.J. Atlas of Clinical Fungi; Webmaster Atlas: Utrecht, The Netherlands, 2014. [Google Scholar]
- Nout, M.J.R.; Aidoo, K.E. Asian fungal fermented food. In Industrial Applications. The Mycota (A Comprehensive Treatise on Fungi as Experimental Systems for Basic and Applied Research); Hofrichter, M., Ed.; Springer: Berlin, Germary, 2010; Volume 10. [Google Scholar]
- Hüttel, W.; Hoffmeister, D. Fungal biotransformations in pharmaceutical sciences. In Industrial Applications. The Mycota (A Comprehensive Treatise on Fungi as Experimental Systems for Basic and Applied Research); Hofrichter, M., Ed.; Springer: Berlin/Heidelberg, Germanry, 2011; Volume 10. [Google Scholar]
- Benny, G.L.; Humber, R.A.; Morton, J.B. Zygomycota: Zygomycetes. In Systematics and Evolution; McLaughlin, D.J., McLaughlin, E.G., Lemke, P.A., Eds.; Springer: Berlin/Heidelberg, Germary, 2001. [Google Scholar] [CrossRef]
- Hibbett, D.S.; Binder, M.; Bischoff, J.F.; Blackwell, M.; Cannon, P.F.; Eriksson, O.E.; Huhndorf, S.; James, T.; Kirk, P.M.; Lucking, R.; et al. A higher-level phylogenetic classification of the fungi. Mycol. Res. 2007, 111, 509–547. [Google Scholar] [CrossRef] [PubMed]
- Kwon-Chung, K.J. Taxonomy of fungi causing mucormycosis and entomophthoramycosis (zygomycosis) and nomenclature of the disease: Molecular mycologic perspectives. Clin. Infect. Dis. 2012, 54, S8–S15. [Google Scholar] [CrossRef]
- Kumar, C.; Jain, P.; Wadhwa, N.; Diwaker, P.; Nirupma Panikar, K. Nosocomial jejunal mucormycosis-an unusual cause of perforation peritonitis. Iran. J. Pathol. 2017, 12, 295–300. [Google Scholar] [PubMed]
- Kokkayil, P.; Pandey, M.; Agarwal, R.; Kale, P.; Singh, G.; Xess, I. Rhizopus homothallicus causing invasive infections: Series of three cases from a single centre in North India. Mycopathologia 2017, 182, 921–926. [Google Scholar] [CrossRef] [PubMed]
- Spatafora, J.W.; Chang, Y.; Benny, G.L.; Lazarus, K.; Smith, M.E.; Berbee, M.L.; Bonito, G.; Corradi, N.; Grigoriev, I.; Gryganskyi, A.; et al. A phylum-level phylogenetic classification of zygomycete fungi based on genome-scale data. Mycologia 2016, 108, 1028–1046. [Google Scholar] [CrossRef]
- Summerbell, R.C. Root endophyte and mycorrhizosphere fungi of black spruce, Picea mariana, in a boreal forest habitat: Influence of site factors on fungal distributions. Stud. Mycol. 2005, 53, 121–145. [Google Scholar] [CrossRef]
- O’Donnell, K.; Lutzoni, F.; Ward, T.J.; Benny, G. Evolutionary relationships among mucoralean fungi (Zygomycota): Evidence for family polyphyly on a large scale. Mycologia 2001, 93, 286–297. [Google Scholar] [CrossRef]
- Voigt, K.; Wöstemeyer, J. Phylogeny and origin of 82 zygomycetes from all 54 genera of the Mucorales and Mortierellales based on combined analysis of actin and translation elongation factor EF-1α genes. Gene 2001, 270, 113–120. [Google Scholar] [CrossRef]
- Kirk, P.M.; Cannon, P.; David, J.; Stalpers, J. Ainsworth & Bisby’s Dictionary of the Fungi; CSIRO Publishing: Collingwood, Astralia, 2008. [Google Scholar]
- Hoffmann, K.; Pawlowska, J.; Walther, G.; Wrzosek, M.; de Hoog, G.S.; Benny, G.L.; Kirk, P.M.; Voigt, K. The family structure of the Mucorales: A synoptic revision based on comprehensive multigene-genealogies. Persoonia 2013, 30, 57–76. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.J.; Ibrahim, A.S.; Skory, C.; Grabherr, M.G.; Burger, G.; Butler, M.; Elias, M.; Idnurm, A.; Lang, B.F.; Sone, T.; et al. Genomic analysis of the basal lineage fungus Rhizopus oryzae reveals a whole-genome duplication. PLoS Genet. 2009, 5, e1000549. [Google Scholar] [CrossRef] [PubMed]
- Corrochano, L.M.; Kuo, A.; Marcet-Houben, M.; Polaino, S.; Salamov, A.; Villalobos-Escobedo, J.M.; Grimwood, J.; Álvarez, M.I.; Avalos, J.; Bauer, D.; et al. Expansion of signal transduction pathways in fungi by extensive genome duplication. Curr. Biol. 2016, 26, 1577–1584. [Google Scholar] [CrossRef] [PubMed]
- Schwartze, V.U.; Winter, S.; Shelest, E.; Marcet-Houben, M.; Horn, F.; Wehner, S.; Linde, J.; Valiante, V.; Sammeth, M.; Riege, K.; et al. Gene expansion shapes genome architecture in the human pathogen Lichtheimia corymbifera: An evolutionary genomics analysis in the ancient terrestrial Mucorales (Mucoromycotina). PLoS Genet. 2014, 10, e1004496. [Google Scholar] [CrossRef]
- De Souza, J.I.; Pires-Zottarelli, C.L.; Dos Santos, J.F.; Costa, J.P.; Harakava, R. Isomucor (Mucoromycotina): A new genus from a Cerrado reserve in state of Sao Paulo, Brazil. Mycologia 2012, 104, 232–241. [Google Scholar] [CrossRef]
- Walther, G.; Pawlowska, J.; Alastruey-Izquierdo, A.; Wrzosek, M.; Rodriguez-Tudela, J.L.; Dolatabadi, S.; Chakrabarti, A.; de Hoog, G.S. DNA barcoding in Mucorales: An inventory of biodiversity. Persoonia 2013, 30, 11–47. [Google Scholar] [CrossRef]
- Wagner, L.; Stielow, J.B.; de Hoog, S.; Bensch, K.; Schwartze, V.; Voigt, K.; Alastruey-Izquierdo, A.; Kurzai, O.; Walther, G. A new species concept for the clinically relevant Mucor circinelloides complex. Persoonia 2019. [Google Scholar] [CrossRef]
- Taylor, J.W.; Jacobson, D.J.; Kroken, S.; Kasuga, T.; Geiser, D.M.; Hibbett, D.S.; Fisher, M.C. Phylogenetic species recognition and species concepts in fungi. Fungal. Genet. Biol. 2000, 31, 21–32. [Google Scholar] [CrossRef]
- Alastruey-Izquierdo, A.; Hoffmann, K.; de Hoog, G.S.; Rodriguez-Tudela, J.L.; Voigt, K.; Bibashi, E.; Walther, G. Species recognition and clinical relevance of the zygomycetous genus Lichtheimia (syn. Absidia pro parte, Mycocladus). J. Clin. Microbiol. 2010, 48, 2154–2170. [Google Scholar] [CrossRef]
- Dolatabadi, S.; de Hoog, G.S.; Meis, J.F.; Walther, G. Species boundaries and nomenclature of Rhizopus arrhizus (syn. R. oryzae). Mycoses 2014, 57, 108–127. [Google Scholar] [CrossRef]
- Ariyawansa, H.A.; Hyde, K.D.; Jayasiri, S.C.; Buyck, B.; Chethana, K.W.T.; Dai, D.Q.; Dai, Y.C.; Daranagama, D.A.; Jayawardena, R.S.; Lücking, R.; et al. Fungal diversity notes 111–252—taxonomic and phylogenetic contributions to fungal taxa. Fungal Divers. 2015, 75, 27–274. [Google Scholar] [CrossRef]
- Bonifaz, A.; Stchigel, A.M.; Guarro, J.; Guevara, E.; Pintos, L.; Sanchis, M.; Cano-Lira, J.F. Primary cutaneous mucormycosis produced by the new species Apophysomyces mexicanus. J. Clin. Microbiol. 2014, 52, 4428–4431. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Álvarez, E.; Stchigel, A.M.; Cano, J.; Sutton, D.A.; Fothergill, A.W.; Chander, J.; Salas, V.; Rinaldi, M.G.; Guarro, J. Molecular phylogenetic diversity of the emerging mucoralean fungus Apophysomyces: Proposal of three new species. Rev. Iberoam. Micol. 2010, 27, 80–89. [Google Scholar] [CrossRef]
- Khuna, S.; Suwannarach, N.; Kumla, J.; Meerak, J.; Nuangmek, W.; Kiatsiriroat, T.; Lumyong, S. Apophysomyces thailandensis (Mucorales, Mucoromycota), a new species isolated from soil in northern Thailand and its solubilization of non-soluble minerals. MycoKeys 2019, 45, 75–92. [Google Scholar] [CrossRef] [PubMed]
- Prakash, H.; Rudramurthy, S.M.; Gandham, P.S.; Ghosh, A.K.; Kumar, M.M.; Badapanda, C.; Chakrabarti, A. Apophysomyces variabilis: Draft genome sequence and comparison of predictive virulence determinants with other medically important Mucorales. BMC Genom. 2017, 18, 736. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Walther, G.; Van Diepeningen, A.D.; Gerrits Van Den Ende, A.H.G.; Li, R.Y.; Moussa, T.A.; Almaghrabi, O.A.; de Hoog, G.S. DNA barcoding of clinically relevant Cunninghamella species. Med. Mycol. 2015, 53, 99–106. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Hermoso, D.; Hoinard, D.; Gantier, J.C.; Grenouillet, F.; Dromer, F.; Dannaoui, E. Molecular and phenotypic evaluation of Lichtheimia corymbifera (formerly Absidia corymbifera) complex isolates associated with human mucormycosis: Rehabilitation of L. ramosa. J. Clin. Microbiol. 2009, 47, 3862–3870. [Google Scholar] [CrossRef]
- Hermet, A.; Meheust, D.; Mounier, J.; Barbier, G.; Jany, J.L. Molecular systematics in the genus Mucor with special regards to species encountered in cheese. Fungal Biol. 2012, 116, 692–705. [Google Scholar] [CrossRef]
- Lima, D.X.; Souza-Motta, C.M.; Wagner, L.; Voigt, K.; De Souza, C.A.F.; De Oliveira, R.J.V.; Da Silva, G.A.; Santiago, A.L.D.A.; Walther, G. Circinella simplex—A misapplied name of Mucor circinatus sp. nov. Phytotaxa 2017, 329. [Google Scholar] [CrossRef]
- Wagner, L.; de Hoog, S.; Alastruey-Izquierdo, A.; Voigt, K.; Kurzai, O.; Walther, G. A revised species concept for opportunistic Mucor species reveals species-specific antifungal susceptibility profiles. Antimicrob. Agents Chemother. 2019, 63, e00653-19. [Google Scholar] [CrossRef]
- Lu, X.L.; Najafzadeh, M.J.; Dolatabadi, S.; Ran, Y.P.; Gerrits van den Ende, A.H.; Shen, Y.N.; Li, C.Y.; Xi, L.Y.; Hao, F.; Zhang, Q.Q.; et al. Taxonomy and epidemiology of Mucor irregularis, agent of chronic cutaneous mucormycosis. Persoonia 2013, 30, 48–56. [Google Scholar] [CrossRef] [PubMed]
- Li, G.J.; Hyde, K.D.; Zhao, R.L.; Hongsanan, S.; Abdel-Aziz, F.A.; Abdel-Wahab, M.A.; Alvarado, P.; Alves-Silva, G.; Ammirati, J.F.; Ariyawansa, H.A.; et al. Fungal diversity notes 253–366: Taxonomic and phylogenetic contributions to fungal taxa. Fungal Divers. 2016, 78, 1–237. [Google Scholar] [CrossRef]
- Liu, X.Y.; Wang, Y.N.; Zheng, R.Y. Molecular phylogeny of Pilaira (Mucorales, Zygomycetes) inferred from ITS rDNA and pyrG sequences. Sydowia 2012, 64, 55–66. [Google Scholar]
- Urquhart, A.S.; Coulon, P.M.L.; Idnurm, A. Pilaira australis sp. nov. (Mucorales, Mucoromycota) isolated from emu faeces in Australia. Phytotaxa 2017, 329. [Google Scholar] [CrossRef]
- Abe, A.; Oda, Y.; Asano, K.; Sone, T. Rhizopus delemar is the proper name for Rhizopus oryzae fumaric-malic acid producers. Mycologia 2007, 99, 714–722. [Google Scholar] [CrossRef]
- Liu, X.; Huang, H.; Zheng, R. Molecular phylogenetic relationships within Rhizopus based on combined analyses of ITS rDNA and pyrG gene sequences. Sydowia 2007, 59, 235–254. [Google Scholar]
- Gryganskyi, A.P.; Lee, S.C.; Litvintseva, A.P.; Smith, M.E.; Bonito, G.; Porter, T.M.; Anishchenko, I.M.; Heitman, J.; Vilgalys, R. Structure, function, and phylogeny of the mating locus in the Rhizopus oryzae complex. PLoS ONE 2010, 5, e15273. [Google Scholar] [CrossRef]
- Dolatabadi, S.; Walther, G.; Gerrits van den Ende, A.H.G.; de Hoog, G.S. Diversity and delimitation of Rhizopus microsporus. Fungal Divers. 2013, 64, 145–163. [Google Scholar] [CrossRef]
- Liou, G.Y.; Chen, S.R.; Wei, Y.H.; Lee, F.L.; Fu, H.M.; Yuan, G.F.; Stalpers, J.A. Polyphasic approach to the taxonomy of the Rhizopus stolonifer group. Mycol. Res. 2007, 111, 196–203. [Google Scholar] [CrossRef]
- Álvarez, E.; Garcia-Hermoso, D.; Sutton, D.A.; Cano, J.F.; Stchigel, A.M.; Hoinard, D.; Fothergill, A.W.; Rinaldi, M.G.; Dromer, F.; Guarro, J. Molecular phylogeny and proposal of two new species of the emerging pathogenic fungus Saksenaea. J. Clin. Microbiol. 2010, 48, 4410–4416. [Google Scholar] [CrossRef]
- Crous, P.W.; Wingfield, M.J.; Burgess, T.I.; Hardy, G.E.; Crane, C.; Barrett, S.; Cano-Lira, J.F.; Le Roux, J.J.; Thangavel, R.; Guarro, J.; et al. Fungal Planet description sheets: 469-557. Persoonia 2016, 37, 218–403. [Google Scholar] [CrossRef] [PubMed]
- Crous, P.W.; Wingfield, M.J.; Burgess, T.I.; Hardy, G.; Barber, P.A.; Alvarado, P.; Barnes, C.W.; Buchanan, P.K.; Heykoop, M.; Moreno, G.; et al. Fungal Planet description sheets: 558-624. Persoonia 2017, 38, 240–384. [Google Scholar] [CrossRef] [PubMed]
- Adamčík, S.; Cai, L.; Chakraborty, D.; Chen, X.-H.; Cotter, H.V.T.; Dai, D.-Q.; Dai, Y.-C.; Das, K.; Deng, C.; Ghobad-Nejhad, M.; et al. Fungal Biodiversity Profiles 1–10. Cryptogam. Mycol. 2015, 36, 121–166. [Google Scholar] [CrossRef]
- De Souza, J.I.; Marano, A.V.; Pires-Zottarelli, C.L.A.; Chambergo, F.S.; Harakava, R. A new species of Backusella (Mucorales) from a Cerrado reserve in Southeast Brazil. Mycol. Prog. 2014, 13. [Google Scholar] [CrossRef]
- Lima, D.X.; Voigt, K.; De Souza, C.A.F.; De Oliveira, R.J.V.; Souza-Motta, C.M.; Santiago, A.L.C.M.D.E.A. Description of Backusella constricta sp. nov. (Mucorales, ex Zygomycota) from the Brazilian Atlantic Rainforest, including a key to species of Backusella. Phytotaxa 2016, 289. [Google Scholar] [CrossRef]
- Tibpromma, S.; Hyde, K.D.; Jeewon, R.; Maharachchikumbura, S.S.N.; Liu, J.-K.; Bhat, D.J.; Jones, E.B.G.; McKenzie, E.H.C.; Camporesi, E.; Bulgakov, T.S.; et al. Fungal diversity notes 491–602: Taxonomic and phylogenetic contributions to fungal taxa. Fungal Divers. 2017, 83, 1–261. [Google Scholar] [CrossRef]
- Nagalakhsmi, S.; Venkat Jothi, R.; Subrahmanyam, A. Zygambella phosphophila gen. & sp. nov. Curr. Trends Biotechnol. Pharm. 2008, 2, 198–200. [Google Scholar]
- Zheng, R.-Y.; Liu, X.-Y. Taxa of Pilaira (Mucorales, Zygomycota) from China. Nova Hedwig. 2009, 88, 255–267. [Google Scholar] [CrossRef]
- Zheng, R.-Y.; Liu, X.-Y.; Wang, Y.-N. Circinella (Mucorales, Mucoromycotina) from China. Mycotaxon 2017, 132, 43–62. [Google Scholar] [CrossRef]
- Loh, L.S.; Nawawi, A.; Kuthubutheen, A.J. Mucoraceous Fungi from Malaysia (Kuala Lumpur); Institute of Biological Sciences, University of Malaya: Kuala Lumpur, Malaysia, 2001. [Google Scholar]
- Ellis, J.J.; Hesseltine, C.W. Two new members of the Mucorales. Mycologia 1969, 61, 863–872. [Google Scholar] [CrossRef]
- Vuillemin, P. Importance taxonomique de l’appareil zygosporé des Mucorinées. Bull. Soc. Mycol. France 1903, 19, 106–118. [Google Scholar]
- Schipper, M.A.A. A study on variability in Mucor hiemalis and related species. Stud. Mycol. 1973, 4, 1–40. [Google Scholar]
- Schipper, M.A.A. On Mucor mucedo, Mucor flavus and related species. Stud. Mycol. 1975, 10, 1–33. [Google Scholar]
- Schipper, M.A.A. On certain species of Mucor with a key to all accepted species. Stud. Mycol. 1978, 17, 1–48. [Google Scholar]
- O’Donnell, K.L.; Ellis, J.J.; Hesseltine, C.W.; Hooper, G.R. Morphogenesis of azygospores induced in Gilbertella persicaria (+) by imperfect hybridization with Rhizopus stolonifer (−). Can. J. Bot. 1977, 55, 2721–2727. [Google Scholar] [CrossRef]
- Schell, W.A.; O’Donnell, K.; Alspaugh, J.A. Heterothallic mating in Mucor irregularis and first isolate of the species outside of Asia. Med. Mycol. 2011, 49, 714–723. [Google Scholar] [CrossRef][Green Version]
- Benny, G.L.; Smith, M.E.; Kirk, P.M.; Tretter, E.D.; White, M.M. Challenges and future perspectives in the systematics of Kickxellomycotina, Mortierellomycotina, Mucoromycotina, and Zoopagomycotina. In Biology of Microfungi; Springer: Berlin, Germany, 2016; pp. 65–126. [Google Scholar] [CrossRef]
- Wijayawardene, N.N.; Pawłowska, J.; Letcher, P.M.; Kirk, P.M.; Humber, R.A.; Schüßler, A.; Wrzosek, M.; Muszewska, A.; Okrasińska, A.; Istel, Ł.; et al. Notes for genera: Basal clades of Fungi (including Aphelidiomycota, Basidiobolomycota, Blastocladiomycota, Calcarisporiellomycota, Caulochytriomycota, Chytridiomycota, Entomophthoromycota, Glomeromycota, Kickxellomycota, Monoblepharomycota, Mortierellomycota, Mucoromycota, Neocallimastigomycota, Olpidiomycota, Rozellomycota and Zoopagomycota). Fungal Divers. 2018, 92, 43–129. [Google Scholar] [CrossRef]
- Mahmud, A.; Lee, R.; Munfus-McCray, D.; Kwiatkowski, N.; Subramanian, A.; Neofytos, D.; Carroll, K.; Zhang, S.X. Actinomucor elegans as an emerging cause of Mucormycosis. J. Clin. Microbiol. 2012, 50, 1092–1095. [Google Scholar] [CrossRef]
- Ryan, L.J.; Ferrieri, P.; Powell, R.D., Jr.; Paddock, C.D.; Zaki, S.R.; Pambuccian, S.E. Fatal Cokeromyces recurvatus pneumonia: Report of a case highlighting the potential for histopathologic misdiagnosis as Coccidioides. Int. J. Surg. Pathol. 2011, 19, 373–376. [Google Scholar] [CrossRef]
- Navanukroh, O.; Jitmuang, A.; Chayakulkeeree, M.; Ngamskulrungroj, P. Disseminated Cunninghamella bertholletiae infection with spinal epidural abscess in a kidney transplant patient: Case report and literature review. Transpl. Infect. Dis. 2014, 16, 658–665. [Google Scholar] [CrossRef]
- García-Rodríguez, J.; Quiles-Melero, I.; Humala-Barbier, K.; Monzon, A.; Cuenca-Estrella, M. Isolation of Cunninghamella blakesleeana in an immunodepressed patient. Mycoses 2012, 55, 463–465. [Google Scholar] [CrossRef] [PubMed]
- Álvarez, E.; Sutton, D.A.; Cano, J.; Fothergill, A.W.; Stchigel, A.; Rinaldi, M.G.; Guarro, J. Spectrum of zygomycete species identified in clinically significant specimens in the United States. J. Clin. Microbiol. 2009, 47, 1650–1656. [Google Scholar] [CrossRef] [PubMed]
- Álvarez, E.; Cano, J.; Stchigel, A.M.; Sutton, D.A.; Fothergill, A.W.; Salas, V.; Rinaldi, M.G.; Guarro, J. Two new species of Mucor from clinical samples. Med. Mycol. 2011, 49, 62–72. [Google Scholar] [CrossRef] [PubMed]
- De Repentigny, L.; St-Germain, G.; Charest, H.; Kokta, V.; Vobecky, S. Fatal zygomycosis caused by Mucor indicus in a child with an implantable left ventricular assist device. Pediatr. Infect. Dis. J. 2008, 27, 355–357. [Google Scholar] [CrossRef]
- Hesseltine, C.W.; Ellis, J.J. An interesting species of Mucor, M. ramosissimus. Sabouraudia 1964, 3, 151–154. [Google Scholar] [CrossRef] [PubMed]
- Iwen, P.C.; Freifeld, A.G.; Sigler, L.; Tarantolo, S.R. Molecular identification of Rhizomucor pusillus as a cause of sinus-orbital zygomycosis in a patient with acute myelogenous leukemia. J. Clin. Microbiol. 2005, 43, 5819–5821. [Google Scholar] [CrossRef] [PubMed]
- Chakrabarti, A.; Marak, R.S.; Shivaprakash, M.R.; Gupta, S.; Garg, R.; Sakhuja, V.; Singhal, S.; Baghela, A.; Dixit, A.; Garg, M.K.; et al. Cavitary pulmonary zygomycosis caused by Rhizopus homothallicus. J. Clin. Microbiol. 2010, 48, 1965–1969. [Google Scholar] [CrossRef]
- Weitzman, I.; McGough, D.A.; Rinaldi, M.G.; Della-Latta, P. Rhizopus schipperae, sp. nov., a new agent of zygomycosis. Mycotaxon 1996, 59, 217–225. [Google Scholar]
- Schlebusch, S.; Looke, D.F. Intraabdominal zygomycosis caused by Syncephalastrum racemosum infection successfully treated with partial surgical debridement and high-dose amphotericin B lipid complex. J. Clin. Microbiol. 2005, 43, 5825–5827. [Google Scholar] [CrossRef]
- Xess, I.; Mohapatra, S.; Shivaprakash, M.R.; Chakrabarti, A.; Benny, G.L.; O’Donnell, K.; Padhye, A.A. Evidence implicating Thamnostylum lucknowense as an etiological agent of rhino-orbital mucormycosis. J. Clin. Microbiol. 2012, 50, 1491–1494. [Google Scholar] [CrossRef]
- Hoffmann, K.; Discher, S.; Voigt, K. Revision of the genus Absidia (Mucorales, Zygomycetes) based on physiological, phylogenetic, and morphological characters; thermotolerant Absidia spp. form a coherent group, Mycocladiaceae fam. nov. Mycol. Res. 2007, 111, 1169–1183. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, K.; Walther, G.; Voigt, K. Mycocladus vs. Lichtheimia, a correction (Lichtheimiaceae fam. nov., Mucorales, Mucoromycotina). Mycol. Res. 2009, 113, 277–278. [Google Scholar]
- De A Santiago, A.L.C.M.; Hoffmann, K.; Lima, D.X.; de Oliveira, R.J.V.; Vieira, H.E.E.; Malosso, E.; Maia, L.C.; da Silva, G.A. A new species of Lichtheimia (Mucoromycotina, Mucorales) isolated from Brazilian soil. Mycol. Prog. 2013, 13, 343–352. [Google Scholar] [CrossRef]
- Schwartze, V.U.; Santiago, A.L.; Jacobsen, I.D.; Voigt, K. The pathogenic potential of the Lichtheimia genus revisited: Lichtheimia brasiliensis is a novel, non-pathogenic species. Mycoses 2014, 57, 128–131. [Google Scholar] [CrossRef]
- Jeong, W.; Keighley, C.; Wolfe, R.; Lee, W.; Slavin, M.A.; Kong, D.C.M.; Chen, C.-A. The epidemiology and clinical manifestations of mucormycosis: A systematic review and meta-analysis of case reports. Clin. Microbiol. Inf. 2019, 25, 26–34. [Google Scholar] [CrossRef]
- Nottebrock, H.; Scholer, H.J.; Wall, M. Taxonomy and identification of mucormycosis-causing fungi I. Synonymity of Absidia ramosa with A. corymbifera. Sabouraudia 1974, 12, 64–74. [Google Scholar] [CrossRef]
- Gomes, M.Z.R.; Lewis, R.E.; Kontoyiannis, D.P. Mucormycosis caused by unusual Mucormycetes, non-Rhizopus, -Mucor, and -Lichtheimia species. Clin. Microbiol. Rev. 2011, 24, 411–445. [Google Scholar] [CrossRef]
- Ganesan, A.; Wells, J.; Shaikh, F.; Peterson, P.; Bradley, W.; Carson, L.; Petfield, J.L.; Klassen-Fischer, M.; Akers, K.S.; Downing, K.; et al. Molecular detection of filamentous fungi in formalin-fixed paraffin-embedded specimens in invasive fungal wound infections is feasible with high specificity. J. Clin. Microbiol. 2019, JCM.01259-19. [Google Scholar] [CrossRef]
- Zheng, R.-Y.; Liu, X.-Y. Actinomucor elegans var. meitauzae, the correct name for A. taiwanensis and Mucor meitauzae (Mucorales, Zygomycota). Nova Hedwig. 2005, 80, 419–432. [Google Scholar] [CrossRef]
- Khan, Z.U.; Ahmad, S.; Mokaddas, E.; Chandy, R.; Cano, J.; Guarro, J. Actinomucor elegans var. kuwaitiensis isolated from the wound of a diabetic patient. Antonie Van Leeuwenhoek 2008, 94, 343–352. [Google Scholar] [CrossRef]
- Jong, S.C.; Yuan, G.F. Actinomucor taiwanensis sp. nov., for manufacture of fermented soybean food. Mycotaxon 1985, 23, 261–264. [Google Scholar]
- Hesseltine, C.W.; Fennell, D.I. The Genus Circinella. Mycologia 1955, 47, 193–212. [Google Scholar] [CrossRef]
- Van Tieghem, P. Nouvelles recherches sur les Mucorinées. Ann. Scienc. Nat. Bot. 1875, Sér. 6, 1, 5–175. [Google Scholar]
- Schipper, M.A.A. On Mucor circinelloides, Mucor racemosus and related species. Stud. Mycol. 1976, 12, 1–40. [Google Scholar]
- Lu, X.L.; Liu, Z.H.; Shen, Y.N.; She, X.D.; Lu, G.X.; Zhan, P.; Fu, M.H.; Zhang, X.L.; Ge, Y.P.; Liu, W.D. Primary cutaneous zygomycosis caused by Rhizomucor variabilis: A new endemic zygomycosis? A case report and review of 6 cases reported from China. Clin. Infect. Dis. 2009, 49, e39–e43. [Google Scholar] [CrossRef]
- Schipper, M.A.A.; Stalpers, J.A.A. Revision of the genus Rhizopus. Stud. Mycol. 1984, 25, 1–34. [Google Scholar]
- Zheng, R.Y.; Chen, G.Q.; Huang, H.; Liu, X.Y. A monograph of Rhizopus. Sydowia 2007, 59, 273–372. [Google Scholar]
- Abe, A.; Asano, K.; Sone, T. A molecular phylogeny-based taxonomy of the genus Rhizopus. Biosci. Biotechnol. Biochem. 2010, 74, 1325–1331. [Google Scholar] [CrossRef]
- Vagvolgyi, C.; Heinrich, H.; Acs, K.; Papp, T. Genetic variability in the species Rhizopus stolonifer, assessed by random amplified polymorphic DNA analysis. Antonie Van Leeuwenhoek 2004, 86, 181–188. [Google Scholar] [CrossRef]
- Zheng, R.Y.; Chen, G.Q. Rhizopus microsporus var. tuberosus var. nov. Mycotaxon 1998, 69, 181–186. [Google Scholar]
- Gryganskyi, A.P.; Golan, J.; Dolatabadi, S.; Mondo, S.; Robb, S.; Idnurm, A.; Muszewska, A.; Steczkiewicz, K.; Masonjones, S.; Liao, H.L.; et al. Phylogenetic and phylogenomic definition of Rhizopus species. G3 (Bethesda) 2018, 8, 2007–2018. [Google Scholar] [CrossRef] [PubMed]
- Fischer, A.; Hauck, F.; Limpricht, G.; Luerssen, C.; Richter, P.; Winter, G. Dr. L. Rabenhorst’s Kryptogamen-Flora von Deutschland, Österreich und der Schweiz-Pilze-Phycomycetes, 1892; 1, 1–505.
- Went, F.A.F.C.; Prinsen Geerligs, H.C. Beobachtungen über die Hefearten und zuckerbildenden Pilze der Arackfabrikation. Verh. Kon. Ned. Akad. Wet Sect. 2 1895, 4, 3–31. [Google Scholar]
- Ellis, J.J. Species and varieties in the Rhizopus arrhizus-Rhizopus oryzae group as indicated by their DNA complementarity. Mycologia 1985, 77, 243–247. [Google Scholar] [CrossRef]
- Saito, K.; Saito, A.; Ohnishi, M.; Oda, Y. Genetic diversity in Rhizopus oryzae strains as revealed by the sequence of lactate dehydrogenase genes. Arch. Microbiol. 2004, 182, 30–36. [Google Scholar] [CrossRef]
- Min, B.; Lee, T.; Choi, Y. Chromosomal studies on the genus of Rhizopus. Korean J. Microbiol. Rev. 1982, 20, 134–146. [Google Scholar]
- Chibucos, M.C.; Soliman, S.; Gebremariam, T.; Lee, H.; Daugherty, S.; Orvis, J.; Shetty, A.C.; Crabtree, J.; Hazen, T.H.; Etienne, K.A.; et al. An integrated genomic and transcriptomic survey of mucormycosis-causing fungi. Nat. Commun. 2016, 7, 12218. [Google Scholar] [CrossRef]
- Kito, H.; Abe, A.; Sujaya, I.N.; Oda, Y.; Asano, K.; Sone, T. Molecular characterization of the relationships among Amylomyces rouxii, Rhizopus oryzae, and Rhizopus delemar. Biosci. Biotechnol. Biochem. 2009, 73, 861–864. [Google Scholar] [CrossRef]
- Vitale, R.G.; de Hoog, G.S.; Schwarz, P.; Dannaoui, E.; Deng, S.; Machouart, M.; Voigt, K.; van de Sande, W.W.; Dolatabadi, S.; Meis, J.F.; et al. Antifungal susceptibility and phylogeny of opportunistic members of the order Mucorales. J. Clin. Microbiol. 2012, 50, 66–75. [Google Scholar] [CrossRef]
- Patel, R. A moldy application of MALDI: MALDI-ToF Mass Spectrometry for fungal identification. J. Fungi 2019, 5, 4. [Google Scholar] [CrossRef]
- Dolatabadi, S.; Kolecka, A.; Versteeg, M.; de Hoog, S.G.; Boekhout, T. Differentiation of clinically relevant Mucorales Rhizopus microsporus and R. arrhizus by matrix-assisted laser desorption ionization time-of-flight mass spectrometry (MALDI-TOF MS). J. Med. Microbiol. 2015, 64, 694–701. [Google Scholar] [CrossRef] [PubMed]
- Schrödl, W.; Heydel, T.; Schwartze, V.U.; Hoffmann, K.; Grosse-Herrenthey, A.; Walther, G.; Alastruey-Izquierdo, A.; Rodriguez-Tudela, J.L.; Olias, P.; Jacobsen, I.D.; et al. Direct analysis and identification of pathogenic Lichtheimia species by matrix-assisted laser desorption ionization-time of flight analyzer-mediated mass spectrometry. J. Clin. Microbiol. 2012, 50, 419–427. [Google Scholar] [CrossRef] [PubMed]
- Rychert, J.; Slechta, E.S.; Barker, A.P.; Miranda, E.; Babady, N.E.; Tang, Y.W.; Gibas, C.; Wiederhold, N.; Sutton, D.; Hanson, K.E. Multicenter evaluation of the Vitek MS v3.0 System for the identification of filamentous fungi. J. Clin. Microbiol. 2018, 56. [Google Scholar] [CrossRef] [PubMed]
- Shao, J.; Wan, Z.; Li, R.; Yu, J. Species identification and delineation of pathogenic Mucorales by Matrix-Assisted Laser Desorption Ionization-Time of Flight Mass Spectrometry. J. Clin. Microbiol. 2018, 56. [Google Scholar] [CrossRef] [PubMed]
- Machouart, M.; Larche, J.; Burton, K.; Collomb, J.; Maurer, P.; Cintrat, A.; Biava, M.F.; Greciano, S.; Kuijpers, A.F.; Contet-Audonneau, N.; et al. Genetic identification of the main opportunistic Mucorales by PCR-restriction fragment length polymorphism. J. Clin. Microbiol. 2006, 44, 805–810. [Google Scholar] [CrossRef]
- Bialek, R.; Konrad, F.; Kern, J.; Aepinus, C.; Cecenas, L.; Gonzalez, G.M.; Just-Nubling, G.; Willinger, B.; Presterl, E.; Lass-Flörl, C.; et al. PCR based identification and discrimination of agents of mucormycosis and aspergillosis in paraffin wax embedded tissue. J. Clin. Pathol. 2005, 58, 1180–1184. [Google Scholar] [CrossRef]
- Hammond, S.P.; Bialek, R.; Milner, D.A.; Petschnigg, E.M.; Baden, L.R.; Marty, F.M. Molecular methods to improve diagnosis and identification of mucormycosis. J. Clin. Microbiol. 2011, 49, 2151–2153. [Google Scholar] [CrossRef]
- Rickerts, V.; Just-Nubling, G.; Konrad, F.; Kern, J.; Lambrecht, E.; Bohme, A.; Jacobi, V.; Bialek, R. Diagnosis of invasive aspergillosis and mucormycosis in immunocompromised patients by seminested PCR assay of tissue samples. Eur. J. Clin. Microbiol. Infect. Dis. 2006, 25, 8–13. [Google Scholar] [CrossRef]
- Bernal-Martinez, L.; Buitrago, M.J.; Castelli, M.V.; Rodriguez-Tudela, J.L.; Cuenca-Estrella, M. Development of a single tube multiplex real-time PCR to detect the most clinically relevant Mucormycetes species. Clin. Microbiol. Infect. 2013, 19, E1–E7. [Google Scholar] [CrossRef]
- Hata, D.J.; Buckwalter, S.P.; Pritt, B.S.; Roberts, G.D.; Wengenack, N.L. Real-time PCR method for detection of zygomycetes. J. Clin. Microbiol. 2008, 46, 2353–2358. [Google Scholar] [CrossRef]
- Hrncirova, K.; Lengerova, M.; Kocmanova, I.; Racil, Z.; Volfova, P.; Palousova, D.; Moulis, M.; Weinbergerova, B.; Winterova, J.; Toskova, M.; et al. Rapid detection and identification of mucormycetes from culture and tissue samples by use of high-resolution melt analysis. J. Clin. Microbiol. 2010, 48, 3392–3394. [Google Scholar] [CrossRef] [PubMed]
- Kasai, M.; Harrington, S.M.; Francesconi, A.; Petraitis, V.; Petraitiene, R.; Beveridge, M.G.; Knudsen, T.; Milanovich, J.; Cotton, M.P.; Hughes, J.; et al. Detection of a molecular biomarker for zygomycetes by quantitative PCR assays of plasma, bronchoalveolar lavage, and lung tissue in a rabbit model of experimental pulmonary zygomycosis. J. Clin. Microbiol. 2008, 46, 3690–3702. [Google Scholar] [CrossRef] [PubMed]
- Legrand, M.; Gits-Muselli, M.; Boutin, L.; Garcia-Hermoso, D.; Maurel, V.; Soussi, S.; Benyamina, M.; Ferry, A.; Chaussard, M.; Hamane, S.; et al. Detection of circulating Mucorales DNA in critically ill burn patients: Preliminary report of a screening strategy for early diagnosis and treatment. Clin. Infect. Dis. 2016, 63, 1312–1317. [Google Scholar] [CrossRef] [PubMed]
- Lengerova, M.; Racil, Z.; Hrncirova, K.; Kocmanova, I.; Volfova, P.; Ricna, D.; Bejdak, P.; Moulis, M.; Pavlovsky, Z.; Weinbergerova, B.; et al. Rapid detection and identification of mucormycetes in bronchoalveolar lavage samples from immunocompromised patients with pulmonary infiltrates by use of high-resolution melt analysis. J. Clin. Microbiol. 2014, 52, 2824–2828. [Google Scholar] [CrossRef]
- Millon, L.; Herbrecht, R.; Grenouillet, F.; Morio, F.; Alanio, A.; Letscher-Bru, V.; Cassaing, S.; Chouaki, T.; Kauffmann-Lacroix, C.; Poirier, P.; et al. Early diagnosis and monitoring of mucormycosis by detection of circulating DNA in serum: Retrospective analysis of 44 cases collected through the French Surveillance Network of Invasive Fungal Infections (RESSIF). Clin. Microbiol. Infect. 2016, 22, 810.e1–810.e8. [Google Scholar] [CrossRef]
- Millon, L.; Larosa, F.; Lepiller, Q.; Legrand, F.; Rocchi, S.; Daguindau, E.; Scherer, E.; Bellanger, A.P.; Leroy, J.; Grenouillet, F. Quantitative polymerase chain reaction detection of circulating DNA in serum for early diagnosis of mucormycosis in immunocompromised patients. Clin. Infect. Dis. 2013, 56, e95–e101. [Google Scholar] [CrossRef]
- Scherer, E.; Iriart, X.; Bellanger, A.P.; Dupont, D.; Guitard, J.; Gabriel, F.; Cassaing, S.; Charpentier, E.; Guenounou, S.; Cornet, M.; et al. Quantitative PCR (qPCR) detection of Mucorales DNA in bronchoalveolar lavage fluid to diagnose pulmonary mucormycosis. J. Clin. Microbiol. 2018, 56. [Google Scholar] [CrossRef]
- Springer, J.; Lackner, M.; Ensinger, C.; Risslegger, B.; Morton, C.O.; Nachbaur, D.; Lass-Flörl, C.; Einsele, H.; Heinz, W.J.; Löffler, J. Clinical evaluation of a Mucorales-specific real-time PCR assay in tissue and serum samples. J. Med. Microbiol. 2016, 65, 1414–1421. [Google Scholar] [CrossRef]
- Alanio, A.; Garcia-Hermoso, D.; Mercier-Delarue, S.; Lanternier, F.; Gits-Muselli, M.; Menotti, J.; Denis, B.; Bergeron, A.; Legrand, M.; Lortholary, O.; et al. Molecular identification of Mucorales in human tissues: Contribution of PCR electrospray-ionization mass spectrometry. Clin. Microbiol. Infect. 2015, 21, 594.e1–594.e5. [Google Scholar] [CrossRef]
- Baldin, C.; Soliman, S.S.M.; Jeon, H.H.; Alkhazraji, S.; Gebremariam, T.; Gu, Y.; Bruno, V.M.; Cornely, O.A.; Leather, H.L.; Sugrue, M.W.; et al. PCR-based approach targeting Mucorales-specific gene family for diagnosis of mucormycosis. J. Clin. Microbiol. 2018, 56. [Google Scholar] [CrossRef]
- Zhao, Z.; Li, L.; Wan, Z.; Chen, W.; Liu, H.; Li, R. Simultaneous detection and identification of Aspergillus and Mucorales species in tissues collected from patients with fungal rhinosinusitis. J. Clin. Microbiol. 2011, 49, 1501–1507. [Google Scholar] [CrossRef] [PubMed]
- Potenza, L.; Vallerini, D.; Barozzi, P.; Riva, G.; Gilioli, A.; Forghieri, F.; Candoni, A.; Cesaro, S.; Quadrelli, C.; Maertens, J.; et al. Mucorales-specific T cells in patients with hematologic malignancies. PLoS ONE 2016, 11, e0149108. [Google Scholar] [CrossRef] [PubMed]
- Domsch, K.H.; Gams, W.; Anderson, T. Compendium of soil fungi; Academic Press: London, UK, 1980. [Google Scholar]
- Karimi, K.; Arzanlou, M.; Ahari, A.B.; Ghazi, M.M. Phenotypic and molecular characterization of the causal agent of chafer beetle mortality in the wheat fields of the Kurdistan province, Iran. J. Plant Prot. Res. 2015, 55. [Google Scholar] [CrossRef]
- Voglmayr, H.; Clémençon, H. Identification and taxonomic position of two mucoralean endoparasites of Hysterangium (Basidiomycota) based on molecular and morphological data. Mycol. Prog. 2015, 15. [Google Scholar] [CrossRef]
- Zheng, R.; Jiang, H. Rhizomucor endophyticus sp. nov., an endophytic zygomycetes from higher plants. Mycotaxon 1995, 56, 455–466. [Google Scholar]
- Zycha, H.; Siepmann, R. Mucorales. Eine Beschreibung aller Gattungen und Arten dieser Pilzgruppe I; Lubrecht & Cramer Ltd.: Lehre, Germany, 1969. [Google Scholar]
- de Souza, C.A.; Lima, D.X.; Gurgel, L.M.; Santiago, A.L. Coprophilous Mucorales (ex Zygomycota) from three areas in the semi-arid of Pernambuco, Brazil. Braz. J. Microbiol. 2017, 48, 79–86. [Google Scholar] [CrossRef][Green Version]
- de Azevedo Santiago, A.L.; Dos Santos, P.J.; Maia, L.C. Mucorales from the semiarid of Pernambuco, Brazil. Braz. J. Microbiol. 2013, 44, 299–305. [Google Scholar] [CrossRef]
- Mousavi, B.; Botterel, F.; Costa, J.M.; Arne, P.; Guillot, J.; Dannaoui, E. Occurrence and species diversity of human-pathogenic Mucorales in commercial food-stuffs purchased in Paris area. Med. Mycol. 2018. [Google Scholar] [CrossRef]
- Mousavi, B.; Costa, J.M.; Arne, P.; Guillot, J.; Chermette, R.; Botterel, F.; Dannaoui, E. Occurrence and species distribution of pathogenic Mucorales in unselected soil samples from France. Med. Mycol. 2018, 56, 315–321. [Google Scholar] [CrossRef]
- Richardson, M. The ecology of the zygomycetes and its impact on environmental exposure. Clin. Microbiol. Infect. 2009, 15, 2–9. [Google Scholar] [CrossRef]
- Benny, G.L.; Benjamin, R.K. Observations on Thamnidiaceae (Mucorales). New taxa, new combinations, and notes on selected species. Aliso 1975, 8, 301–351. [Google Scholar] [CrossRef][Green Version]
- Benny, G.L.; Benjamin, R.K. The Radiomycetaceae (Mucorales; Zygomycetes). III. A New species of Radiomyces, and cladistic analysis and taxonomy of the family; with a discussion of evolutionary ordinal relationships in Zygomycotina. Mycologia 1991, 83, 713–735. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Studied Taxon | rDNA | actin | tef | rpb1 | ldhB | rpb2 | tpt | RNA Helicase | tsr1 | mcm7 | cfs | h3 | pyrG | Reference |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Mucorales | LSU/SSU | x | O’Donnell et al. 2001 [14] | |||||||||||
| Mucorales | x | x | Voigt et al. 2001 [15] | |||||||||||
| Mucorales | LSU/SSU | x | x | Hoffmann et al. 2013 [17] | ||||||||||
| Absidia koreana | ITS/LSU/SSU | x | x | Ariyawansa et al. 2015 [27] | ||||||||||
| Apophysomyces | ITS/LSU | x | Bonifaz et al. 2014 [28] | |||||||||||
| Apophysomyces | ITS/LSU | x | Álvarez et al. 2010 [29] | |||||||||||
| Apophysomyces | ITS/LSU | x | Khuna et al. 2019 [30] | |||||||||||
| Apophysomyces | x (+ 13 other) | Prakash et al. 2017 [31] | ||||||||||||
| Cunninghamella | ITS | x | Yu et al. [32] | |||||||||||
| Gongronella koreana | ITS/LSU/SSU | x | x | Ariyawansa et al. 2015 [27] | ||||||||||
| Lichtheimia | ITS/LSU | x | Garcia-Hermosa et al. 2009 [33] | |||||||||||
| Lichtheimia | ITS/LSU | x * | Alastruey-Izquierdo et al. 2010 [25] | |||||||||||
| Mucor | ITS/LSU/SSU | x | x | x | Hermet et al. 2012 [34] | |||||||||
| Mucor / Isomucor | ITS/LSU | x * | x | de Souza et al. [21] | ||||||||||
| Mucor circinatus and relatives | ITS/LSU | x | Lima et al. 2017 [35] | |||||||||||
| Mucor circinelloides complex | ITS | x | x | x | x | Wagner et al. 2019 [36] | ||||||||
| Mucor irregularis | LSU | x | x | x | x | Lu et al. 2013 [37] | ||||||||
| Mucor koreanus | ITS/LSU/SSU | x | x | Li et al. 2016 [38] | ||||||||||
| Pilaira | ITS | x | Liu et al. 2012 [39] | |||||||||||
| Pilaira australis | ITS | x | Urquhart et al. 2017 [40] | |||||||||||
| Rhizopus | ITS | x | x | x | Abe et al. 2007 [41] | |||||||||
| Rhizopus | ITS | x | Liu 2007 [42] | |||||||||||
| Rhizopus arrhizus | ITS | x | x | x | x | Gryganskyi 2010 [43] | ||||||||
| Rhizopus arrhizus | ITS | x | x * | x | Dolatabadi et al. 2014 [26] | |||||||||
| Rhizopus | IGS | Liu et al. 2007 [42] | ||||||||||||
| Rhizopus microsporus | ITS | x | x | Dolatabadi et al. 2013 [44] | ||||||||||
| Rhizopus stolonifer group | LSU | Liou et al. 2007 [45] | ||||||||||||
| Saksenaea | ITS/LSU | x | Álvarez et al. 2010 [46] | |||||||||||
| Saksenaea | ITS/LSU | x | Crous et al. 2016 [47] | |||||||||||
| Saksenaea | ITS/LSU | x | Crous et al. 2017 [48] |
| Genus | Family According to Hoffmann et al. 2013 | Sequences Available | Accepted Species | Clinically Relevant Species | Species Described after the Year 2000 |
|---|---|---|---|---|---|
| Absidia | Cunninghamellaceae | yes | 20 | 0 | 7 |
| Actinomucor | Mucoraceae | yes | 1 | 1 | 0 |
| Ambomucor | Mucoraceae (?) | no | 3 | 0 | 3 |
| Apophysomyces | Saksenaeaceae | yes | 6 | 4 | 5 |
| Backusella | Backusellaceae | yes | 14 | 0 | 4 |
| Benjaminiella | Mucoraceae | yes | 3 | 0 | 0 |
| Blakeslea | Choanephoraceae | yes | 2 | 0 | 0 |
| Chaetocladium | Mucoraceae | yes | 2 | 0 | 0 |
| Chlamydoabsidia | Cunninghamellaceae | yes | 1 | 0 | 0 |
| Choanephora | Choanephoraceae | yes | 2 | 0 | 0 |
| Circinella | Lichtheimiaceae | yes | 8 | 0 | 2 |
| Cokeromyces | Mucoraceae | yes | 1 | 1 | 0 |
| Cunninghamella | Cunninghamellaceae | yes | 14 | 4 | 4 |
| Dichotomocladium | Lichtheimiaceae | yes | 5 | 0 | 0 |
| Dicranophora | Mucoraceae | yes | 1 | 0 | 0 |
| Ellisomyces | Mucoraceae | yes | 1 | 0 | 0 |
| Fennellomyces | Lichtheimiaceae | yes | 4 | 0 | 0 |
| Gilbertella | Choanephoraceae | yes | 1 | 0 | 0 |
| Gongronella | Cunninghamellaceae | yes | 6 | 0 | 4 |
| Halteromyces | Cunninghamellaceae | yes | 1 | 0 | 0 |
| Helicostylum | Mucoraceae | yes | 2 | 0 | 0 |
| Hesseltinella | Cunninghamellaceae | yes | 1 | 0 | 0 |
| Hyphomucor | Mucoraceae | yes | 1 | 0 | 0 |
| Isomucor | Mucoraceae | yes | 1 | 0 | 1 |
| Kirkiana | Mucoraceae (?) | no | 1 | 0 | 1 |
| Kirkomyces | Mucoraceae | yes | 1 | 0 | 0 |
| Lentamyces | Lentamycetaceae | yes | 4 | 0 | 0 |
| Lichtheimia | Lichtheimiaceae | yes | 6 | 3 | 2 |
| Mucor | Mucoraceae | yes | 76 | 13 | 20 |
| Mycotypha | Mycotyphaceae | yes | 3 | 1 | 0 |
| Nawawiella | Mucoraceae (?) | no | 1 | 0 | 1 |
| Parasitella | Mucoraceae | yes | 1 | 0 | 0 |
| Phascolomyces | Lichtheimiaceae | yes | 1 | 0 | 1 |
| Phycomyces | Phycomycetaceae | yes | 3 | 0 | 0 |
| Pilaira | Mucoraceae | yes | 8 | 0 | 3 |
| Pilobolus | Pilobolaceae | yes | 7 | 0 | 0 |
| Pirella | Mucoraceae | yes | 2 | 0 | 0 |
| Poitrasia | Choanephoraceae | yes | 1 | 0 | 0 |
| Protomycocladus | Syncephalastraceae | yes | 1 | 0 | 0 |
| Radiomyces | Radiomycetaceae | yes | 3 | 0 | 0 |
| Rhizomucor | Lichtheimiaceae | yes | 2 | 2 | 0 |
| Rhizopodopsis | Rhizopodacea (?) | no | 1 | 0 | 0 |
| Rhizopus | Rhizopodaceae | yes | 10 | 4 | 1 |
| Saksenaea | Saksenaeaceae | yes | 5 | 4 | 4 |
| Spinellus | Phycomycetaceae | yes | 5 | 0 | 0 |
| Sporodiniella | Rhizopodaceae | yes | 1 | 0 | 0 |
| Syncephalastrum | Syncephalastraceae | yes | 2 | 1 | 0 |
| Syzygites | Rhizopodaceae | yes | 1 | 0 | 0 |
| Thamnidium | Mucoraceae | yes | 1 | 0 | 0 |
| Thamnostylum | Lichtheimiaceae | yes | 4 | 1 | 0 |
| Thermomucor | Lichtheimiaceae | yes | 1 | 0 | 0 |
| Tortumyces | Mucoraceae (?) | no | 2 | 0 | 2 |
| Utharomyces | Pilobolaceae | yes | 1 | 0 | 0 |
| Zychaea | Lichtheimiaceae | yes | 1 | 0 | 0 |
| Zygambella | Rhizopodaceae (?) | no | 1 | 0 | 1 |
| Mucorales total | 260 | 39 | 67 |
| Species | Previous Names/Important Synonyms | Reference |
|---|---|---|
| Actinomucor elegans | Mahmud et al. 2011 [66] | |
| (Apophysomyces elegans) * | Alvarez et al. 2010 [29] | |
| Apophysomyces mexicanus | Bonifaz et al. 2014 [28] | |
| Apophysomyces ossiformis | Álvarez et al. 2010 [29] | |
| Apophysomyces trapeziformis | Álvarez et al. 2010 [29] | |
| Apophysomyces variabilis | Álvarez et al. 2010 [29] | |
| Cokeromyces recurvatus | Ryan et al. 2011 [67] | |
| Cunninghamella bertholletiae | Navanukroh et al. 2014 [68] | |
| Cunninghamella blakesleeana | GarcíaRodríguez et al. 2012 [69] | |
| Cunninghamella echinulata | Álvarez et al. 2009 [70] | |
| Cunninghamella elegans | Yu et al. 2015 [32] | |
| Lichtheimia corymbifera | Absidia corymbifera, Mycocladus corymbifer | Alastruey-Izquierdo et al. 2010 [25] |
| Lichtheimia ornata | Absidia ornata | Alastruey-Izquierdo et al. 2010 [25] |
| Lichtheimia ramosa | Absidia ramosa, Mycocladus ramosus | Alastruey-Izquierdo et al. 2010 [25] |
| Mucor amphibiorum | Walther et al. 2013 [22] | |
| Mucor ardhlaengiktus | Mucor ellipsoideus | Álvarez et al. 2011 [71] |
| Mucor circinelloides | Mucor circinelloides f. circinelloides, Rhizomucor regularior, Rhizomucor variabilis var. regularior | Wagner et al. 2019 [36] |
| Mucor griseocyanus | Mucor circinelloides f. griseocyanus | Wagner et al. 2019 [36] |
| Mucor indicus | de Repentigny et al. 2008 [72] | |
| Mucor irregularis | Rhizomucor variabilis | Lu et al. 2013 [37] |
| Mucor janssenii | Mucor circinelloides f. janssenii | Walther et al. 2013 [22] |
| Mucor lusitanicus | Mucor circinelloides f. lusitanicus | Álvarez et al. 2011 [71] |
| Mucor plumbeus | Walther et al. 2013 [22] | |
| Mucor racemosus | Walther et al. 2013 [22] | |
| Mucor ramosissimus | Hesseltine & Ellis 1964 [73] | |
| Mucor variicolumellatus | Álvarez et al. 2011 (as M. fragilis) [71] | |
| Mucor velutinosus | Álvarez et al. 2011 [71] | |
| Rhizomucor miehei | Walther et al. 2013 [22] | |
| Rhizomucor pusillus | Iwen et al. 2005 [74] | |
| Rhizopus arrhizus (incl. var. delemar) | Rhizopus oryzae | Dolatabadi et al. 2014 [26] |
| Rhizopus homothallicus | Chakrabarti et al. 2010 [75] | |
| Rhizopus microsporus | Rhizopus microsporus var. azygosporus, var. chinensis, var. oligosporus, var. rhizopodiformis, var. tuberosus | Dolatabadi et al. 2013 [44] |
| Rhizopus schipperae | Weizmann et al. 1996 [76] | |
| Saksenaea erythrospora | Álvarez et al. 2010 [46] | |
| Saksenaea loutrophoriformis | Crous et al. 2017 [48] | |
| Saksenaea trapezispora | Álvarez et al. 2010 [46] | |
| Saksenaea vasiformis | Álvarez et al. 2010 [46] | |
| Syncephalastrum racemosum | Schlebusch et al. 2005 [77] | |
| Thamnostylum lucknowense | Xess et al. 2012 [78] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Walther, G.; Wagner, L.; Kurzai, O. Updates on the Taxonomy of Mucorales with an Emphasis on Clinically Important Taxa. J. Fungi 2019, 5, 106. https://doi.org/10.3390/jof5040106
Walther G, Wagner L, Kurzai O. Updates on the Taxonomy of Mucorales with an Emphasis on Clinically Important Taxa. Journal of Fungi. 2019; 5(4):106. https://doi.org/10.3390/jof5040106
Chicago/Turabian StyleWalther, Grit, Lysett Wagner, and Oliver Kurzai. 2019. "Updates on the Taxonomy of Mucorales with an Emphasis on Clinically Important Taxa" Journal of Fungi 5, no. 4: 106. https://doi.org/10.3390/jof5040106
APA StyleWalther, G., Wagner, L., & Kurzai, O. (2019). Updates on the Taxonomy of Mucorales with an Emphasis on Clinically Important Taxa. Journal of Fungi, 5(4), 106. https://doi.org/10.3390/jof5040106