A Solvent-Free Approach for Converting Cellulose Waste into Volatile Organic Compounds with Endophytic Fungi

 , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
3. Results and Discussion
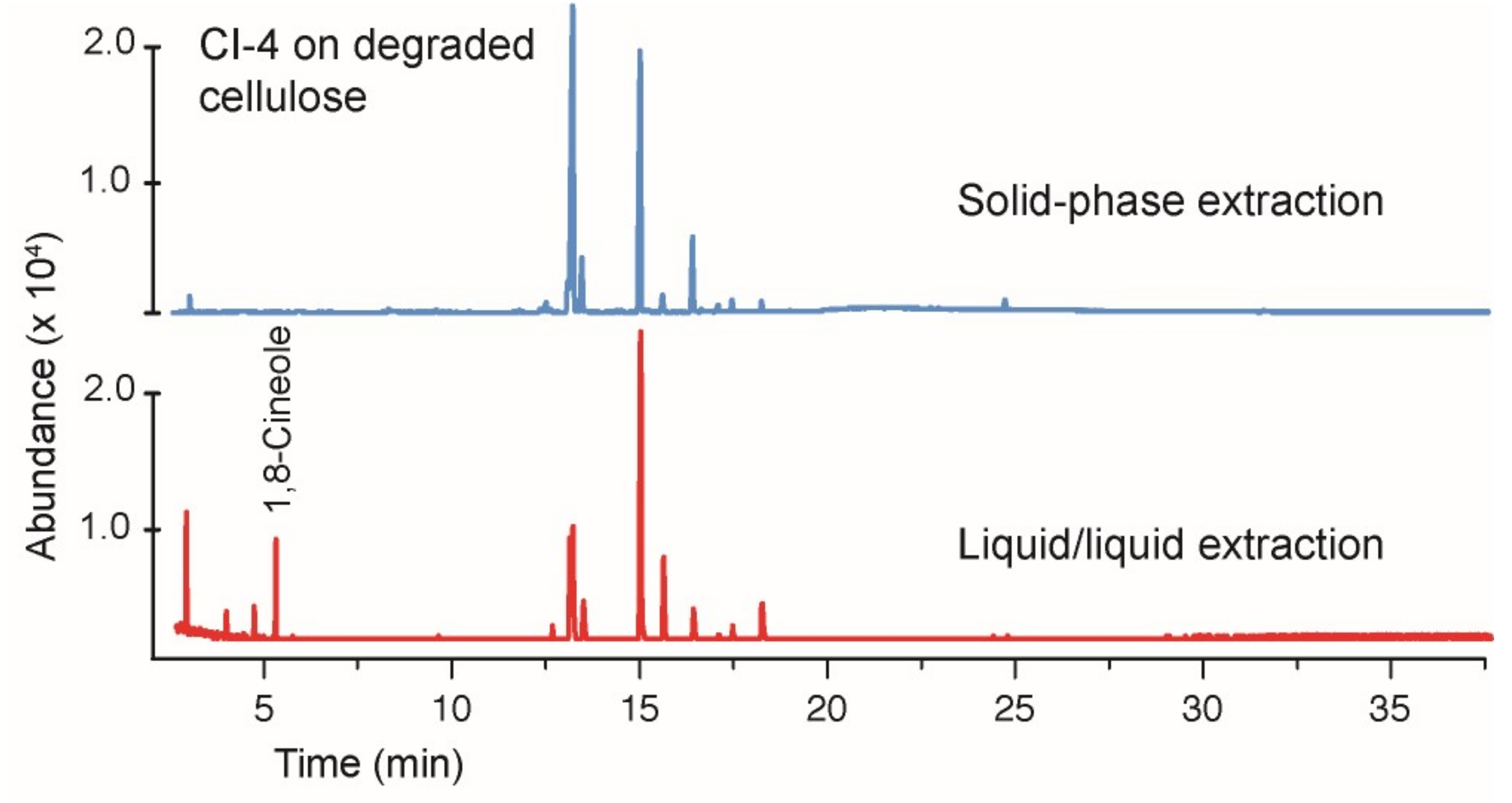
3.1. Growth of the Hypoxylon CI-4 on Degraded Cellulose
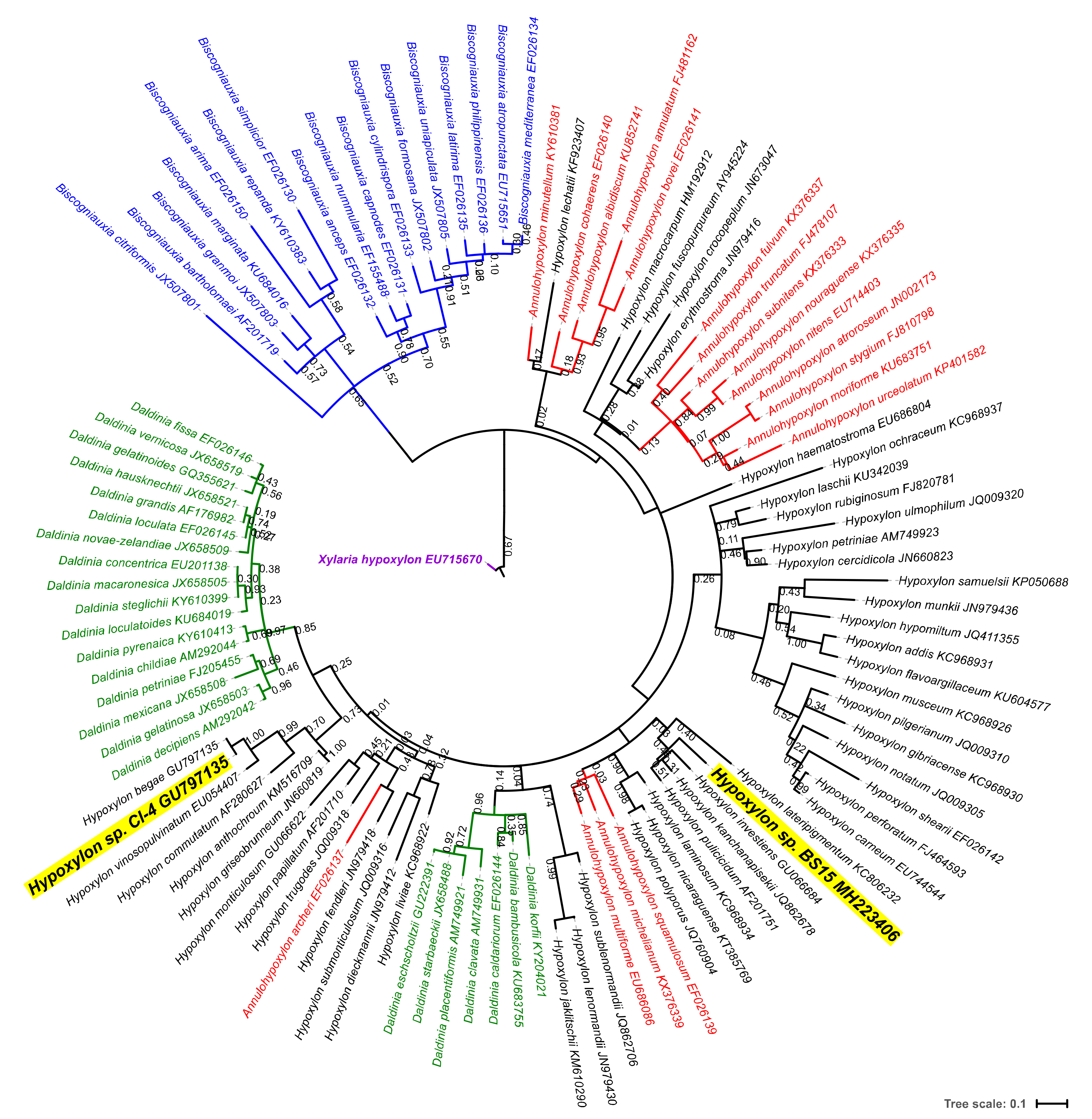
3.2. Phylogenetic Characterization of a New Hypoxylon sp., BS15
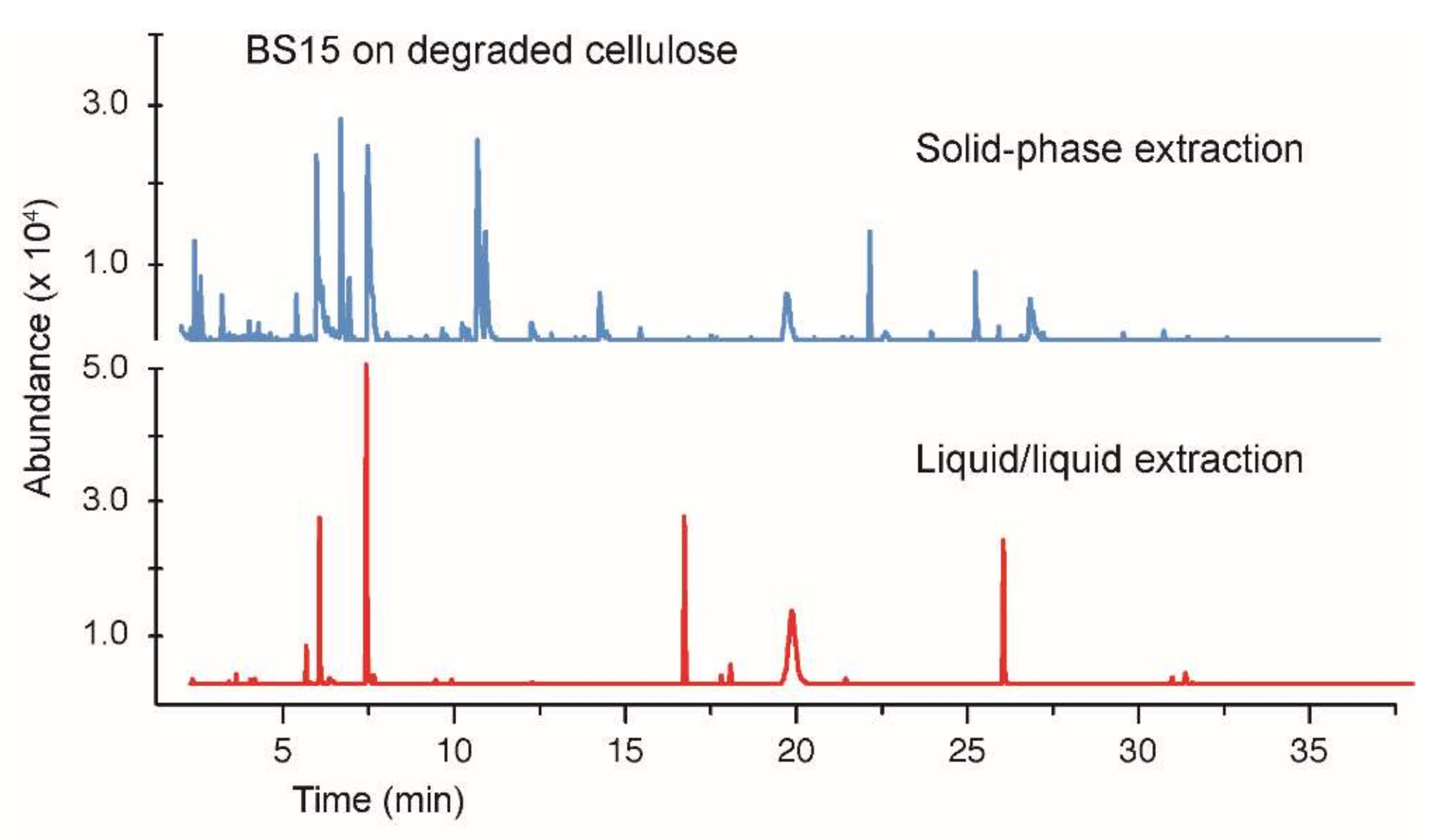
3.3. Growth of BS15 on Degraded Cellulose and Analysis of Volatile Hydrocarbons
3.4. Comparing Solid-Phase and Liquid–Liquid Extraction Methods
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Stone, J.K.; Bacon, C.W.; White, J.F., Jr. An overview of endophytic microbes: Endophytism defined. In Microbial Endophytes; Bacon, C.W., White, J.F., Jr., Eds.; Marcel Dekker: New York, NY, USA, 2000; pp. 29–33. ISBN 0.8247-8831-1. [Google Scholar]
- Strobel, G.A.; Daisy, B.; Castillo, U.C.; Harper, J.K. Natural products from endophytic microorganisms. J. Nat. Prod. 2004, 67, 257–268. [Google Scholar] [CrossRef] [PubMed]
- Strobel, G.A.; Knighton, B.; Kluck, K.; Ren, Y.; Livinghouse, T.; Griffin, M.; Spakowicz, D.; Sears, J. The production of myco-diesel hydrocarbons and their derivatives by the endophytic fungus Gliocladium roseum (NRRL 50072). Microbiology 2008, 154, 3319–3328. [Google Scholar] [CrossRef] [PubMed]
- Strobel, G.A. Bioprospecting—Fuels from fungi. Biotechnol. Lett. 2015, 37, 973–982. [Google Scholar] [CrossRef] [PubMed]
- Strobel, G. Muscodor species-endophytes with biological promise. Phytochem. Rev. 2011, 10, 165–172. [Google Scholar] [CrossRef]
- Banerjee, D.; Strobel, G.A.; Booth, E.; Geary, B.; Sears, J.; Spakowicz, D.; Busse, S. An endophytic Myrothecium inundatum producing volatile organic compounds. Mycosphere 2010, 1, 229–240. [Google Scholar]
- Ul-Hassan, S.R.; Strobel, G.A.; Booth, E.; Knighton, B.; Floerchinger, C.; Sears, J. Modulation of volatile organic compound formation in the Mycodiesel-producing endophyte Hypoxylon sp. CI-4. Microbiology 2012, 158, 465–473. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.K.; Strobel, G.A.; Knighton, B.; Geary, B.; Sears, J.; Ezra, D. An Endophytic Phomopsis sp. possessing bioactivity and fuel potential with its volatile organic compounds. Microb. Ecol. 2011, 61, 729–739. [Google Scholar] [CrossRef] [PubMed]
- Griffin, M.A.; Spakowicz, D.J.; Gianoulis, T.A.; Strobel, S.A. Volatile organic compound production by organisms in the genus Ascocoryne and a re-evaluation of myco-diesel production by NRRL 50072. Microbiology 2010, 156, 3814–3829. [Google Scholar] [CrossRef] [PubMed]
- Strobel, G.; Singh, S.K.; Riyaz-Ul-Hassan, S.; Mitchell, A.M.; Geary, B.; Sears, J. An endophytic/pathogenic Phoma sp. from creosote bush producing biologically active volatile compounds having fuel potential. FEMS Microbiol. Lett. 2011, 320, 87–94. [Google Scholar] [CrossRef] [PubMed]
- Zhao, G.; Yin, G.; Inamdar, A.A.; Luo, J.; Zhang, N.; Yang, I.; Buckley, B.; Bennett, J.W. Volatile organic compounds emitted by filamentous fungi isolated from flooded homes after Hurricane Sandy show toxicity in a Drosophila bioassay. Indoor Air 2017, 27, 518–528. [Google Scholar] [CrossRef] [PubMed]
- Siddiquee, S.; Azad, S.A.; Abu Bakar, F.; Naher, L.; Vijay Kumar, S. Separation and identification of hydrocarbons and other volatile compounds from cultures of Aspergillus niger by GC–MS using two different capillary columns and solvents. J. Saudi Chem. Soc. 2015, 19, 243–256. [Google Scholar] [CrossRef]
- Fiers, M.; Lognay, G.; Fauconnier, M.-L.; Jijakli, M.H. Volatile Compound-Mediated Interactions between Barley and Pathogenic Fungi in the Soil. PLoS ONE 2013, 8, e66805. [Google Scholar] [CrossRef] [PubMed]
- Savelieva, E.I.; Gustyleva, L.K.; Kessenikh, E.D.; Khlebnikova, N.S.; Leffingwell, J.; Gavrilova, O.P.; Gagkaeva, T.Y. Study of the vapor phase over Fusarium fungi cultured on various substrates. Chem. Biodivers. 2016, 13, 891–903. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, A.M.; Strobel, G.A.; Moore, E.; Robison, R.; Sears, J. Volatile antimicrobials from Muscodor crispans, a novel endophytic fungus. Microbiology 2010, 156, 270–277. [Google Scholar] [CrossRef] [PubMed]
- Meshram, V.; Kapoor, N.; Saxena, S. Muscodor kashayum sp. nov.—A new volatile anti-microbial producing endophytic fungus. Mycology 2013, 4, 196–204. [Google Scholar] [CrossRef] [PubMed]
- Meshram, V.; Saxena, S.; Kapoor, N. Muscodor strobelii, a new endophytic species from South India. Mycotaxon 2014, 128, 93–104. [Google Scholar] [CrossRef]
- Saxena, S.; Meshram, V.; Kapoor, N. Muscodor darjeelingensis, a new endophytic fungus of Cinnamomum camphora collected from northeastern Himalayas. Sydowia 2014, 66, 55–67. [Google Scholar]
- Saxena, S.; Meshram, V.; Kapoor, N. Muscodor tigerii sp. nov.-Volatile antibiotic producing endophytic fungus from the Northeastern Himalayas. Ann. Microbiol. 2015, 65, 47–57. [Google Scholar] [CrossRef]
- Suwannarach, N.; Kumla, J.; Bussaban, B.; Hyde, K.D.; Matsui, K.; Lumyong, S. Molecular and morphological evidence support four new species in the genus Muscodor from northern Thailand. Ann. Microbiol. 2013, 63, 1341–1351. [Google Scholar] [CrossRef]
- Kudalkar, P.; Strobel, G.; Riyaz-Ul-Hassan, S.; Geary, B.; Sears, J. Muscodor sutura, a novel endophytic fungus with volatile antibiotic activities. Mycoscience 2012, 53, 319–325. [Google Scholar] [CrossRef]
- Zhang, C.L.; Wang, G.P.; Mao, L.J.; Komon-Zelazowska, M.; Yuan, Z.L.; Lin, F.C.; Druzhinina, I.S.; Kubicek, C.P. Muscodor fengyangensis sp. nov. from southeast China: Morphology, physiology and production of volatile compounds. Fungal Biol. 2010, 114, 797–808. [Google Scholar] [CrossRef] [PubMed]
- Samaga, P.V.; Rai, V.R.; Rai, K.M.L. Bionectria ochroleuca NOTL33-an endophytic fungus from Nothapodytes foetida producing antimicrobial and free radical scavenging metabolites. Ann. Microbiol. 2014, 64, 275–285. [Google Scholar] [CrossRef]
- Naznin, H.A.; Kiyohara, D.; Kimura, M.; Miyazawa, M.; Shimizu, M.; Hyakumachi, M. Systemic Resistance Induced by Volatile Organic Compounds Emitted by Plant Growth-Promoting Fungi in Arabidopsis thaliana. PLoS ONE 2014, 9, e86882. [Google Scholar] [CrossRef] [PubMed]
- Crespo, R.; Pedrini, N.; Juárez, M.P.; Dal Bello, G.M. Volatile organic compounds released by the entomopathogenic fungus Beauveria bassiana. Microbiol. Res. 2008, 163, 148–151. [Google Scholar] [CrossRef] [PubMed]
- Shurin, J.B.; Burkart, M.D.; Mayfield, S.P.; Smith, V.H. Recent progress and future challenges in algal biofuel production. F1000Research 2016, 5, 2434. [Google Scholar] [CrossRef] [PubMed]
- Nass, L.L.; Pereira, P.A.A.; Ellis, D. Biofuels in Brazil: An overview. Crop Sci. 2007, 47, 2228–2237. [Google Scholar] [CrossRef]
- Hicks, S.M.; Griebel, C.; Restrepo, D.T.; Truitt, J.H.; Baker, E.J.; Bylda, C.; Blair, R.G. Mechanocatalysis for biomass-derived chemicals and fuels. Green Chem. 2010, 12, 468–474. [Google Scholar] [CrossRef]
- Strobel, G.A.; Dirkse, E.; Sears, J.; Markworth, C. Volatile antimicrobials from Muscodor albus, a novel endophytic fungus. Microbiology 2001, 147, 2943–2950. [Google Scholar] [CrossRef] [PubMed]
- Riyaz-Ul-Hassan, S.; Strobel, G.A.; Geary, B.; Sears, J. An endophytic Nodulisporium sp. from Central America producing volatile organic compounds with both biological and fuel potential. J. Microbiol. Biotechnol. 2013, 23, 29–35. [Google Scholar] [CrossRef] [PubMed]
- Boom, R.; Sol, C.J.A.; Salimans, M.M.M.; Jansen, C.L.; Wertheim-van Dillen, P.M.E.; van der Noordaa, J. Rapid and simple method for purification of nucleic acids. J. Clin. Microbiol. 1990, 28, 495–503. [Google Scholar] [PubMed]
- Schoch, C.L.; Seifert, K.A.; Huhndorf, S.; Robert, V.; Spouge, J.L.; Levesque, C.; André; Chen, W. Nuclear ribosomal internal transcribed spacer (ITS) region as a universal DNA barcode marker for Fungi. Proc. Natl. Acad. Sci. USA 2012, 109, 6241–6246. [Google Scholar] [CrossRef] [PubMed]
- Saiki, R.K.; Scharf, S.; Faloona, F.; Mullis, K.B.; Horn, G.T.; Erlich, H.A.; Arnheim, N. Enzymatic amplification of beta-globin genomic sequences and restriction site analysis for diagnosis of sickle cell anemia. Science 1985, 230, 1350–1354. [Google Scholar] [CrossRef] [PubMed]
- Toju, H.; Tanabe, A.S.; Yamamoto, S.; Sato, H. High-Coverage ITS Primers for the DNA-Based Identification of Ascomycetes and Basidiomycetes in Environmental Samples. PLoS ONE 2012, 7, e40863. [Google Scholar] [CrossRef] [PubMed]
- Carbone, I.; Kohn, L.M. A method for designing primer sets for speciation studies in filamentous ascomycetes. Mycologia 1999, 91, 553–556. [Google Scholar] [CrossRef]
- O’Donnell, K.; Cigelnik, E. Two divergent intragenomic rDNA ITS2 types within a monophyletic lineage of the fungus Fusarium are nonorthologous. Mol. Phylogenet Evol. 1997, 7, 103–116. [Google Scholar] [CrossRef] [PubMed]
- Cock, P.J.A.; Antao, T.; Chang, J.T.; Chapman, B.A.; Cox, C.J.; Dalke, A.; Friedberg, I.; Hamelryck, T.; Kauff, F.; Wilczynski, B.; et al. Biopython: Freely available Python tools for computational molecular biology and bioinformatics. Bioinformatics 2009, 25, 1422–1423. [Google Scholar] [CrossRef] [PubMed]
- Bengtsson-Palme, J.; Ryberg, M.; Hartmann, M.; Branco, S.; Wang, Z.; Godhe, A.; De Wit, P.; Sánchez-García, M.; Ebersberger, I.; de Sousa, F.; et al. ITSx: Improved software detection and extraction of ITS1 and ITS2 from ribosomal ITS sequences of fungi and other eukaryotes for use in environmental sequencing. Methods Ecol. Evol. 2013, 4, 914–919. [Google Scholar] [CrossRef]
- Kõljalg, U.; Larsson, K.-H.; Abarenkov, K.; Nilsson, R.H.; Alexander, I.J.; Eberhardt, U.; Erland, S.; Høiland, K.; Kjøller, R.; Larsson, E.; et al. UNITE: A database providing web-based methods for the molecular identification of ectomycorrhizal fungi. New Phytol. 2005, 166, 1063–1068. [Google Scholar] [CrossRef]
- Boratyn, G.M.; Camacho, C.; Cooper, P.S.; Coulouris, G.; Fong, A.; Ma, N.; Madden, T.L.; Matten, W.T.; McGinnis, S.D.; Merezhuk, Y.; et al. BLAST: A more efficient report with usability improvements. Nucleic Acids Res. 2013, W29–W33. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, H.-M.; Ju, Y.-M.; Rogers, J.D. Molecular Phylogeny of Hypoxylon and Closely Related Genera. Mycologia 2005, 97, 844–865. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [PubMed]
- Holder, M.; Lewis, P.O. Phylogeny estimation: Traditional and Bayesian approaches. Nat. Rev. Genet. 2003, 4, 275–284. [Google Scholar] [CrossRef] [PubMed]
- Letunic, I.; Bork, P. Interactive tree of life (iTOL) v3: An online tool for the display and annotation of phylogenetic and other trees. Nucleic Acids Res. 2016, 44, W242–W245. [Google Scholar] [CrossRef] [PubMed]
- Tomsheck, A.R.; Strobel, G.A.; Booth, E.; Geary, B.; Spakowicz, D.; Knighton, B.; Floerchinger, C.; Sears, J.; Liarzi, O.; Ezra, D. Hypoxylon sp., an endophyte of Persea indica, producing 1,8-cineole and other bioactive volatiles with fuel potential. Microb. Ecol. 2010, 60, 903–914. [Google Scholar] [CrossRef] [PubMed]
- Shaw, J.J.; Berbasova, T.; Sasaki, T.; Jefferson-George, K.; Spakowicz, D.J.; Dunican, B.F.; Portero, C.E.; Narváez-Trujillo, A.; Strobel, S.A. Identification of a fungal 1,8-cineole synthase from Hypoxylon sp. with common specificity determinants to the plant synthases. J. Biol. Chem. 2015, 290, 8511–8526. [Google Scholar] [CrossRef] [PubMed]
- Warnow, T. Standard maximum likelihood analyses of alignments with gaps can be statistically inconsistent. PLoS Curr. 2012, 4, RRN1308. [Google Scholar] [CrossRef] [PubMed]
- Wendt, L.; Sir, E.B.; Kuhnert, E.; Heitkämper, S.; Lambert, C.; Hladki, A.I.; Romero, A.I.; Luangsa-ard, J.J.; Srikitikulchai, P.; Peršoh, D.; et al. Resurrection and emendation of the Hypoxylaceae, recognised from a multigene phylogeny of the Xylariales. Mycol. Prog. 2018, 17, 115–154. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Feedstock | Percent Hydrolyzed a |
|---|---|
| Cherry pit | 95.7 |
| Flint corn kernals | 93.4 |
| St. Augustine grass | 92.5 |
| Oat | 90.3 |
| Orange peel | 85.0 |
| Corn cobs | 81.5 |
| Bamboo | 75.1 |
| Cedar | 74.0 |
| Red Oak | 72.4 |
| Maple | 72.0 |
| Douglas Fir | 71.1 |
| Nannochloropsis | 69.2 |
| Aspen | 68.0 |
| Poplar | 66.9 |
| Yellow pine | 65.3 |
| Wheat | 65.0 |
| Miscanthus grass | 64.7 |
| White pine | 64.4 |
| Mixed yard waste | 58.1 |
| Switch grass | 57.9 |
| Hickory | 55.9 |
| Paper, newsprint | 54.7 |
| Flint corn stover | 52.1 |
| Banana leaf | 52.0 |
| Big blue stem grass | 50.1 |
| Little blue stem grass | 48.9 |
| Coffee grounds | 45.2 |
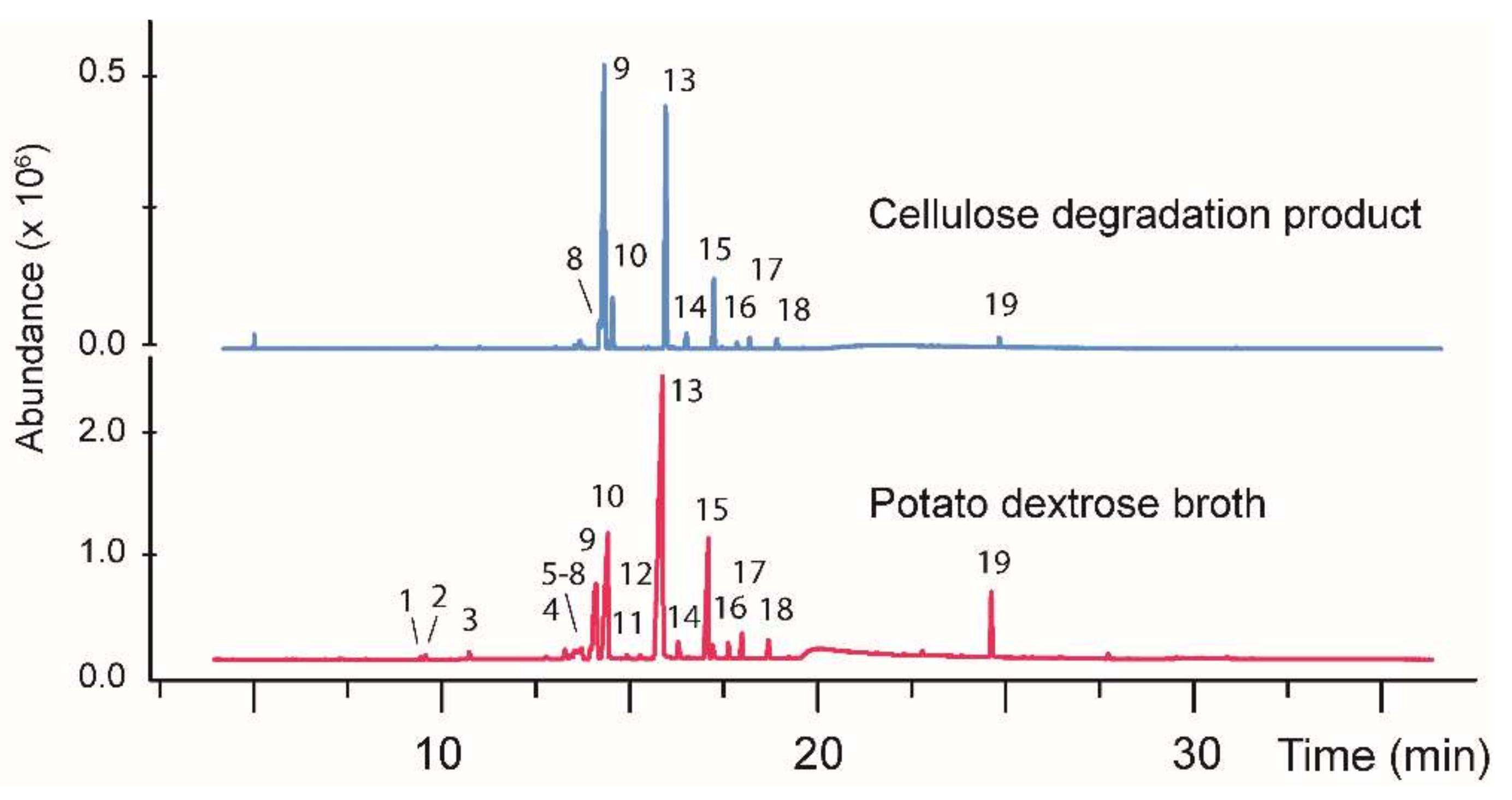
| Fungus | Peak # a | R.T. (min) | Area (%) PD, DC b | Tentative Identity c | Mol. mass (Da) | Qual. d |
|---|---|---|---|---|---|---|
| CI-4 | 1 | 9.58 | 0.1, - | Unknown | 126 | - |
| CI-4 | 2 | 9.72 | 0.3, - | Unknown | 138 | - |
| CI-4 | 3 | 10.88 | 0.5, - | Unknown | 124 | - |
| CI-4 | 4 | 12.93 | 0.2, - | Unknown | 152 | - |
| CI-4 | 5 | 13.43 | 0.8, - | Unknown | 122 | - |
| CI-4 | 6 | 13.57 | 0.3, - | Unknown | 154 | - |
| CI-4 | 7 | 13.68 | 1.4, - | 3-Ethenyl-2-methylene cyclopentanecarboxylic acid | 152 | 50 |
| CI-4 | 8 | 14.11 | 0.6, 2.8 | Unknown | 152 | - |
| CI-4 | 9 | 14.25 | 10.6, 45.7 | Unknown | 150 | - |
| CI-4 | 10 | 14.57 | 15.5, 6.0 | Unknown | 154 | - |
| CI-4 | 11 | 15.08 | 0.3, - | Unknown | 154 | - |
| CI-4 | 12 | 15.44 | 0.3, - | Unknown | 152 | - |
| CI-4 | 13 | 16.02 | 43.8, 2.0 | Unknown | 168 | - |
| CI-4 | 14 | 16.45 | 1.1, 2.0 | 1-Acetyl-2-(1-hydroxyethyl)-cyclohexene | 168 | 50 |
| CI-4 | 15 | 17.24 | 10.4, 7.6 | Unknown | 150 | - |
| CI-4 | 16 | 17.79 | 1.0, 0.8 | Unknown | 170 | - |
| CI-4 | 17 | 18.14 | 1.5, 1.2 | Unknown | 170 | - |
| CI-4 | 18 | 18.84 | 1.1, 1.1 | 3-Isopropoxy 5-methyl-phenol | 166 | 61 |
| CI-4 | 19 | 24.78 | 4.0, 1.0 | 2,3-Dimethoxy-naphthalene | 188 | 85 |
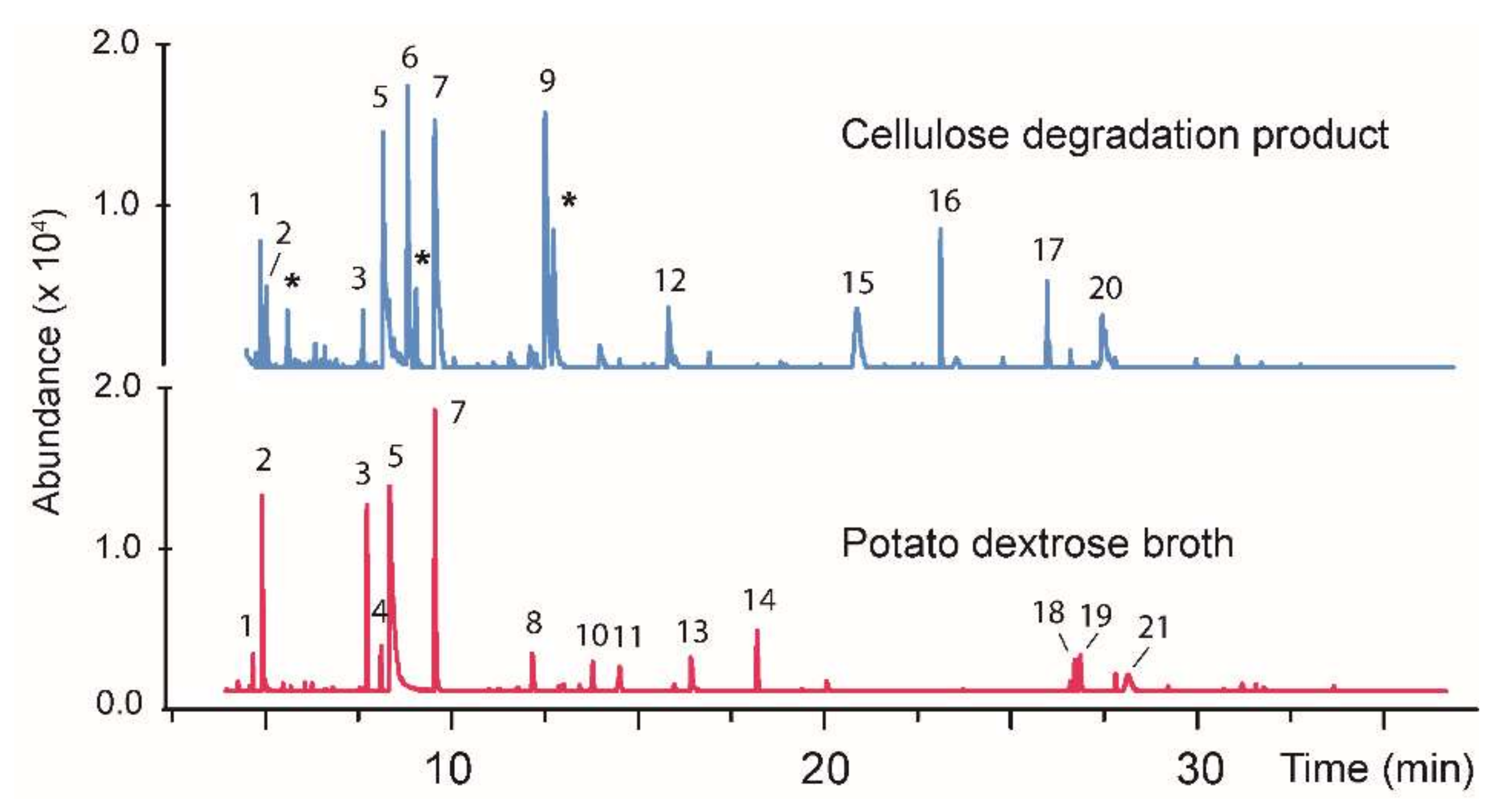
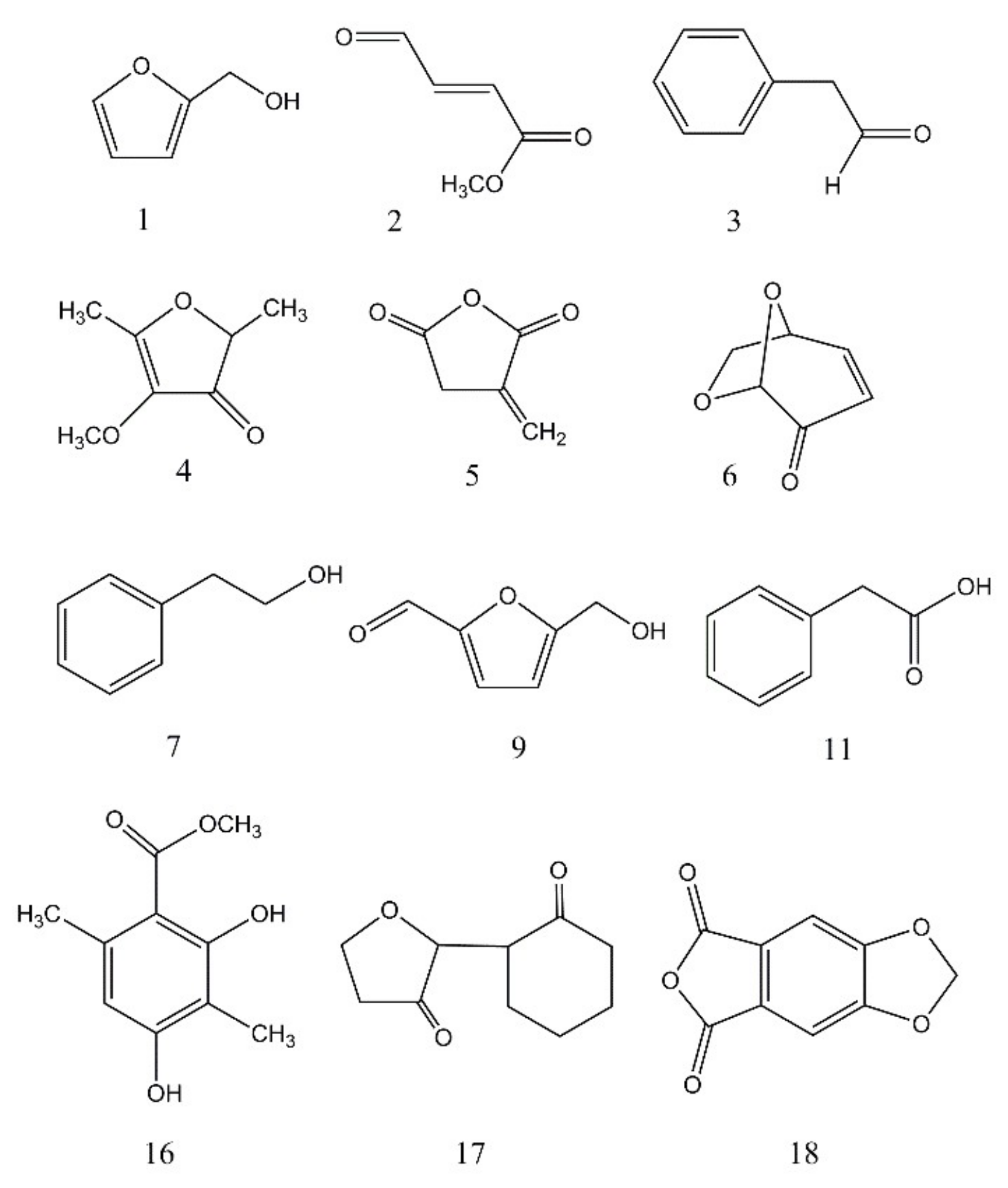
| BS15 | 1 | 4.82 | 1.1, 2.7 | Furfuryl alcohol | 98 | 72 |
| BS15 | 2 | 5.07 | 9.9, 2.4 | Methyl 4-oxo-2-butenoate | 114 | 94 |
| BS15 | 3 | 7.85 | 9.3, 1.7 | Benzeneacetaldehyde | 120 | 70 |
| BS15 | 4 | 8.22 | 1.6, - | 4-methoxy-2,5-dimethyl-3 (2H)-furanone | 142 | 77 |
| BS15 | 5 | 8.45 | 36.7, 26.1 | 2,5-furandione dihydro-3-methylene | 112 | 55 |
| BS15 | 6 | 8.65 | -, 21.5 | Levoglucosenone | 126 | 78 |
| BS15 | 7 | 9.65 | 20.9, 12.8 | 2-Phenyethanol | 122 | 86 |
| BS15 | 8 | 12.24 | 3.0, - | Unknown | 158 | - |
| BS15 | 9 | 12.34 | -, 20.3 | 5-(Hydroxymethyl)furfural | 126 | 91 |
| BS15 | 10 | 13.84 | 17.8 | Unknown | 86 | – |
| BS15 | 11 | 14.56 | 2.5, - | Phenylacetic acid | 136 | 75 |
| BS15 | 12 | 16.23 | 3.0, - | Unknown | 138 | - |
| BS15 | 13 | 16.45 | -, 3.2 | Unknown | 142 | - |
| BS15 | 14 | 18.20 | 3.7, – | Unknown | 154 | - |
| BS15 | 15 | 20.47 | -, 8.1 | Unknown | 162 | - |
| BS15 | 16 | 22.73 | -, 3.2 | 2,4-dihydroxy-3,6-dimethyl Benzoic acid, methyl ester | 196 | 72 |
| BS15 | 17 | 25.58 | -, 2.1 | Dihydro-5-(2-oxocyclohexylidene) 2(3H)-furanone | 180 | 70 |
| BS15 | 18 | 26.63 | 1.7, - | Furo [3, 4-f][1,3] benzodioxole-5,7-dione | 192 | 65 |
| BS15 | 19 | 26.78 | 2.9, - | Unknown | 97 | - |
| BS15 | 20 | 27.06 | 8.7, - | Unknown | 127 | - |
| BS15 | 21 | 28.04 | 1.8, - | Unknown | 127 | - |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maxwell, T.; Blair, R.G.; Wang, Y.; Kettring, A.H.; Moore, S.D.; Rex, M.; Harper, J.K. A Solvent-Free Approach for Converting Cellulose Waste into Volatile Organic Compounds with Endophytic Fungi. J. Fungi 2018, 4, 102. https://doi.org/10.3390/jof4030102
Maxwell T, Blair RG, Wang Y, Kettring AH, Moore SD, Rex M, Harper JK. A Solvent-Free Approach for Converting Cellulose Waste into Volatile Organic Compounds with Endophytic Fungi. Journal of Fungi. 2018; 4(3):102. https://doi.org/10.3390/jof4030102
Chicago/Turabian StyleMaxwell, Tyler, Richard G. Blair, Yuemin Wang, Andrew H. Kettring, Sean D. Moore, Matthew Rex, and James K. Harper. 2018. "A Solvent-Free Approach for Converting Cellulose Waste into Volatile Organic Compounds with Endophytic Fungi" Journal of Fungi 4, no. 3: 102. https://doi.org/10.3390/jof4030102
APA StyleMaxwell, T., Blair, R. G., Wang, Y., Kettring, A. H., Moore, S. D., Rex, M., & Harper, J. K. (2018). A Solvent-Free Approach for Converting Cellulose Waste into Volatile Organic Compounds with Endophytic Fungi. Journal of Fungi, 4(3), 102. https://doi.org/10.3390/jof4030102