
Regulation of Yeast Cytokinesis by Calcium


Abstract
1. Introduction
2. Molecular Mechanism of Cytokinesis
3. Ca2+ Homeostasis of Eukaryotic Cells
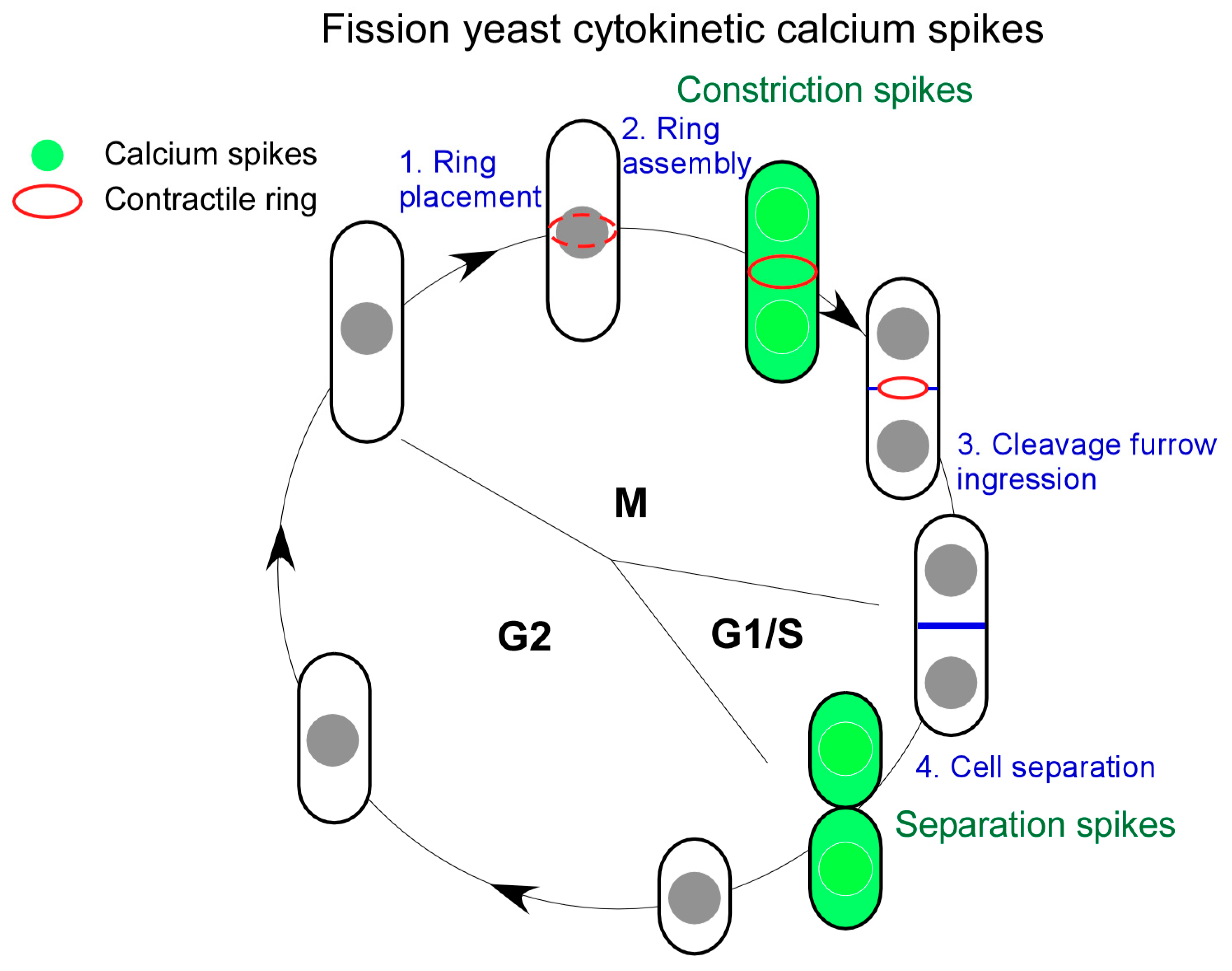
4. Cytokinetic Ca2+ Spikes in Yeast
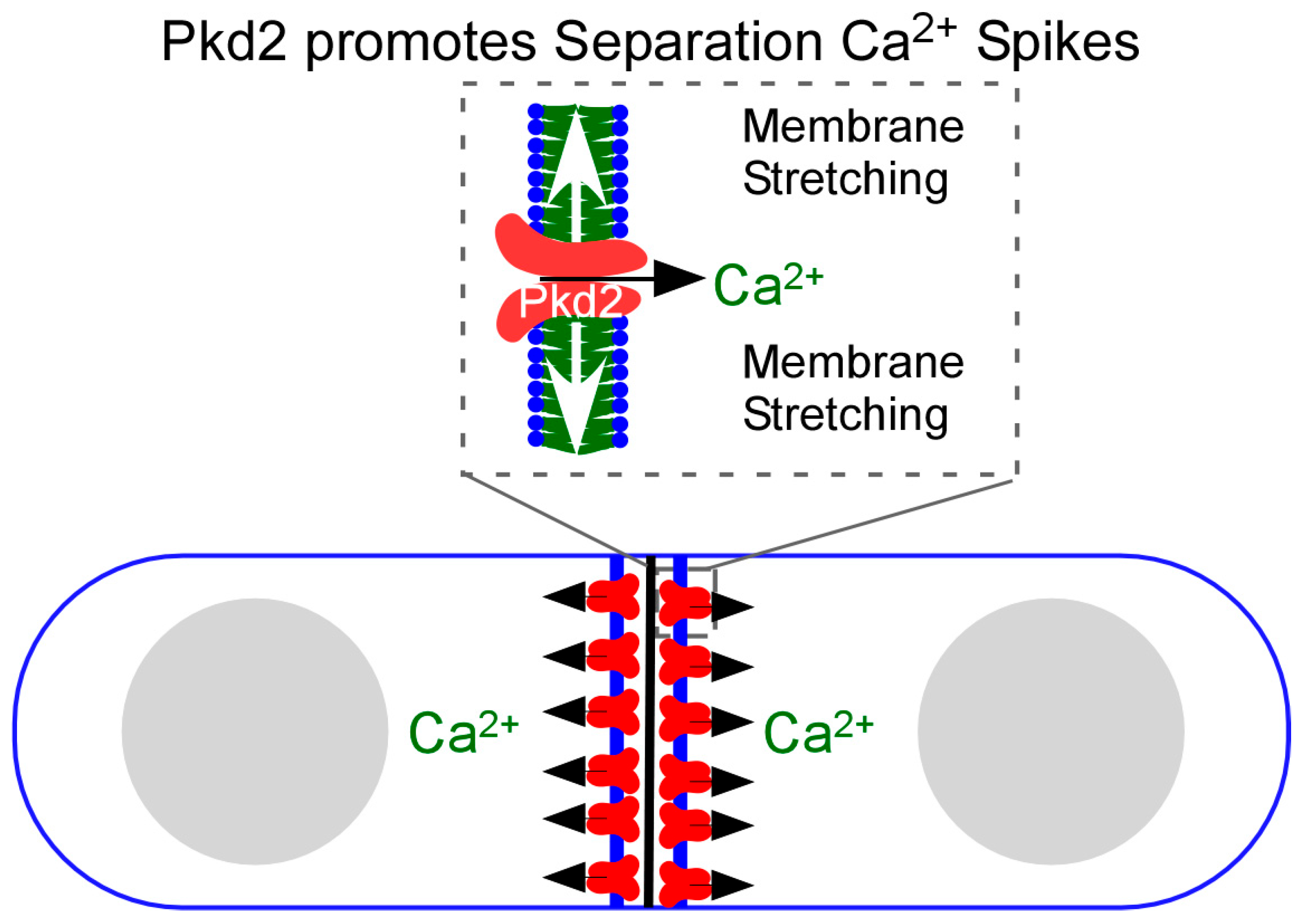
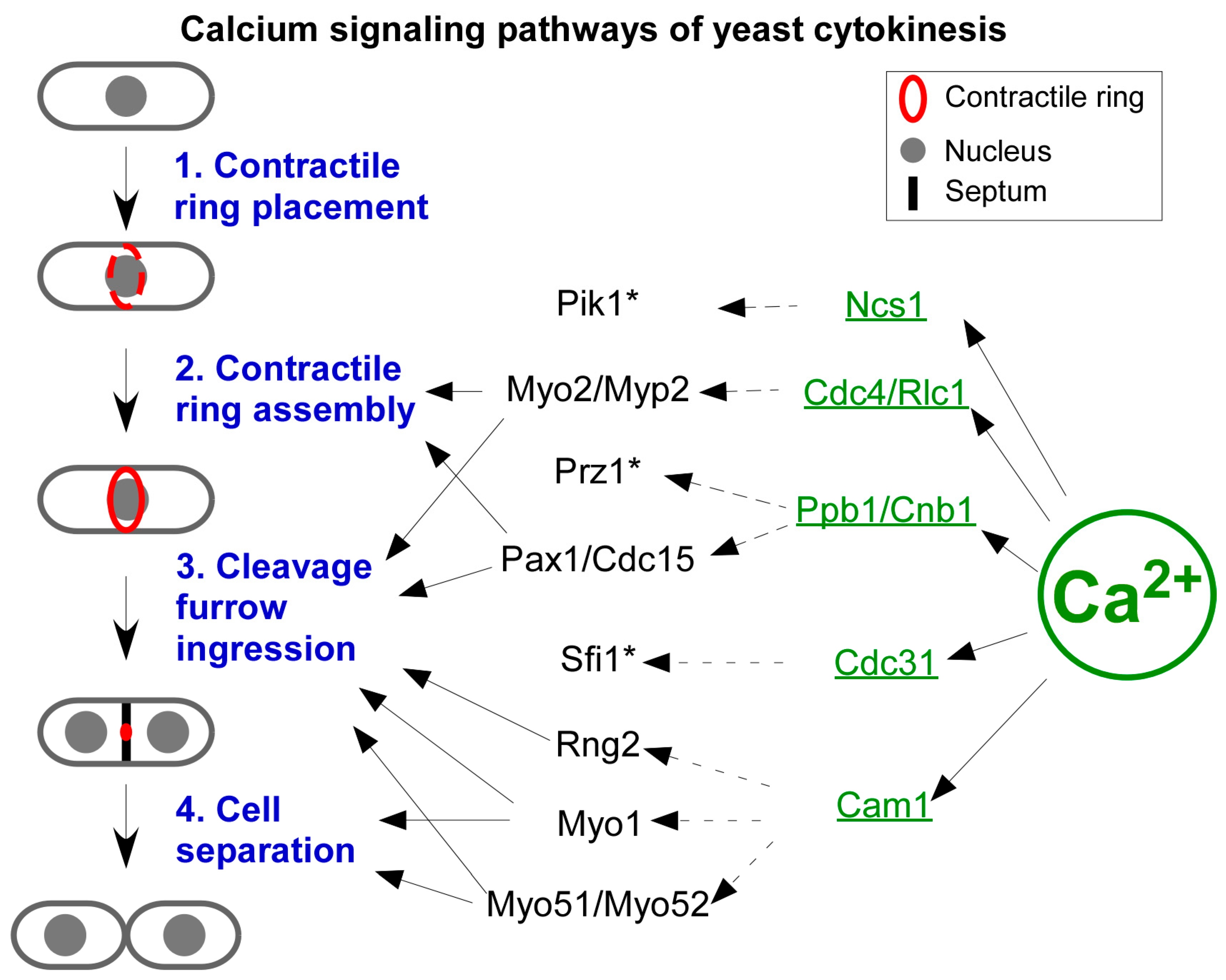
5. Regulatory Mechanism of Cytokinetic Ca2+ Spikes
6. The Effectors of Cytokinetic Ca2+ Spikes
7. Comparison of Ca2+ Homeostasis and Signaling Between Two Yeasts

{kind=link}
{kind=link}
{kind=link}
| S. cerevisiae | S. pombe | ||
|---|---|---|---|
| Ca2+ Channels | Voltage-gated | Cch1+Mid1+Ecm7 [24,95] | Cch1+Yam8 [50,96] |
| TRP family | Yvc1 [87] | None identified | |
| Mechanosensitive Pkd2 family | Flc1, Flc2, Flc3, and YOR365C [83,97] | Pkd2, Trp1322, and Trp663 [44] | |
| Mechanosensitive MscS-like | None identified | Msy1 and Msy2 [47] | |
| Others | Csg2 [91] | None identified | |
| Ca2+-ATPases | ER | Spf1 [98] | Pmr1 and Cta4 [27,28] |
| Golgi | Pmr1 [21] | None identified | |
| Vacuolar | Pmc1 [99] | Pmc1 [27] | |
| Ca2+-binding EF-hand proteins | Calmodulin | Cmd1 [100] | Cam1 [57] |
| Calcineurin | Cna1/Cna2+Cnb1 [101] | Ppb1+Cnb1 [67,102] | |
| Centrin | Cdc31 [103] | Cdc31 [72] | |
| Myosin light chains | Mlc1 and Mlc2 [104,105] | Cdc4 and Rlc1 [73,74] | |
| Neuronal Ca2+ sensor | Frq1 [77] | Ncs1 [75] |
8. Comparison Between Regulation of Cytokinesis by Ca2+ in Yeast and Animal Cells
| Animal | Yeast | |
|---|---|---|
| Cytokinetic Ca2+ transients | Furrowing wave Zipping wave [1] | Constriction spike Separation spike [35] |
| Efflux channel promoting Ca2+ transients | IP3 receptor [37] | None identified |
| Influx channel promoting Ca2+ transients | ORAI [108] | Pkd2 [46] |
| Potential targets of Ca2+ transients | Calmodulin [115] | Cam1 [58] |
| Calcineurin [116] | Ppb1+Cnb1 [67,117] | |
| MLCK [118] | None identified | |
| CaMKII | Cmk1 and Cmk2 | |
| Annexin [119] | None identified | |
| VAMP2 [120] | None identified |
9. Conclusions and Open Questions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fluck, R.A.; Miller, A.L.; Jaffe, L.F. Slow calcium waves accompany cytokinesis in medaka fish eggs. J. Cell Biol. 1991, 115, 1259–1265. [Google Scholar] [PubMed]
- Arnold, J.M. Effect of calcium in cytokinesis as demonstrated with ionophore A231ß7. Cytobiologie 1975, 11, 1–9. [Google Scholar]
- Snow, P.; Nuccitelli, R. Calcium buffer injections delay cleavage in Xenopus laevis blastomeres. J. Cell Biol. 1993, 122, 387–394. [Google Scholar] [CrossRef]
- Chang, D.C.; Meng, C. A localized elevation of cytosolic free calcium is associated with cytokinesis in the zebrafish embryo. J. Cell Biol. 1995, 131, 1539–1545. [Google Scholar]
- Webb, S.E.; Miller, A.L. Ca2+ Signalling and Membrane Dynamics During Cytokinesis in Animal Cells. Adv. Exp. Med. Biol. 2017, 981, 389–412. [Google Scholar] [CrossRef]
- Miller, A.L.; Fluck, R.A.; McLaughlin, J.A.; Jaffe, L.F. Calcium buffer injections inhibit cytokinesis in Xenopus eggs. J. Cell Sci. 1993, 106 Pt 2, 523–534. [Google Scholar]
- Noguchi, T.; Mabuchi, I. Localized calcium signals along the cleavage furrow of the Xenopus egg are not involved in cytokinesis. Mol. Biol. Cell 2002, 13, 1263–1273. [Google Scholar] [CrossRef]
- Fujiwara, T.; Bandi, M.; Nitta, M.; Ivanova, E.V.; Bronson, R.T.; Pellman, D. Cytokinesis failure generating tetraploids promotes tumorigenesis in p53-null cells. Nature 2005, 437, 1043–1047. [Google Scholar] [CrossRef]
- Pollard, T.D.; O’Shaughnessy, B. Molecular Mechanism of Cytokinesis. Annu. Rev. Biochem. 2019, 88, 661–689. [Google Scholar] [CrossRef]
- Pollard, T.D.; Wu, J.Q. Understanding cytokinesis: Lessons from fission yeast. Nat. Rev. 2010, 11, 149–155. [Google Scholar]
- Daga, R.R.; Chang, F. Dynamic positioning of the fission yeast cell division plane. Proc. Natl. Acad. Sci. USA 2005, 102, 8228–8232. [Google Scholar] [CrossRef] [PubMed]
- Kitayama, C.; Sugimoto, A.; Yamamoto, M. Type II myosin heavy chain encoded by the myo2 gene composes the contractile ring during cytokinesis in Schizosaccharomyces pombe. J. Cell Biol. 1997, 137, 1309–1319. [Google Scholar] [CrossRef] [PubMed]
- Chang, F.; Drubin, D.; Nurse, P. cdc12p, a protein required for cytokinesis in fission yeast, is a component of the cell division ring and interacts with profilin. J. Cell Biol. 1997, 137, 169–182. [Google Scholar] [PubMed]
- Chen, Q.; Pollard, T.D. Actin filament severing by cofilin is more important for assembly than constriction of the cytokinetic contractile ring. J. Cell Biol. 2011, 195, 485–498. [Google Scholar] [CrossRef]
- Proctor, S.A.; Minc, N.; Boudaoud, A.; Chang, F. Contributions of turgor pressure, the contractile ring, and septum assembly to forces in cytokinesis in fission yeast. Curr. Biol. 2012, 22, 1601–1608. [Google Scholar] [CrossRef]
- Johnson, A.E.; McCollum, D.; Gould, K.L. Polar opposites: Fine-tuning cytokinesis through SIN asymmetry. Cytoskeleton 2012, 69, 686–699. [Google Scholar] [CrossRef]
- Wu, J.Q.; Kuhn, J.R.; Kovar, D.R.; Pollard, T.D. Spatial and temporal pathway for assembly and constriction of the contractile ring in fission yeast cytokinesis. Dev. Cell 2003, 5, 723–734. [Google Scholar] [CrossRef]
- Courtemanche, N.; Pollard, T.D.; Chen, Q. Avoiding artefacts when counting polymerized actin in live cells with LifeAct fused to fluorescent proteins. Nat. Cell Biol. 2016, 18, 676–683. [Google Scholar] [CrossRef]
- Pollard, T.D. Nine unanswered questions about cytokinesis. J. Cell Biol. 2017, 216, 3007–3016. [Google Scholar] [CrossRef]
- Dunn, T.; Gable, K.; Beeler, T. Regulation of cellular Ca2+ by yeast vacuoles. J. Biol. Chem. 1994, 269, 7273–7278. [Google Scholar] [CrossRef]
- Strayle, J.; Pozzan, T.; Rudolph, H.K. Steady-state free Ca2+ in the yeast endoplasmic reticulum reaches only 10 microM and is mainly controlled by the secretory pathway pump pmr1. EMBO J. 1999, 18, 4733–4743. [Google Scholar] [CrossRef] [PubMed]
- Clapham, D.E. Calcium signaling. Cell 2007, 131, 1047–1058. [Google Scholar] [CrossRef] [PubMed]
- Catterall, W.A.; Lenaeus, M.J.; Gamal El-Din, T.M. Structure and Pharmacology of Voltage-Gated Sodium and Calcium Channels. Annu. Rev. Pharmacol. Toxicol. 2020, 60, 133–154. [Google Scholar] [CrossRef]
- Paidhungat, M.; Garrett, S. A homolog of mammalian, voltage-gated calcium channels mediates yeast pheromone-stimulated Ca2+ uptake and exacerbates the cdc1(Ts) growth defect. Mol. Cell Biol. 1997, 17, 6339–6347. [Google Scholar] [CrossRef]
- Carnero, E.; Ribas, J.C.; Garcia, B.; Duran, A.; Sanchez, Y. Schizosaccharomyces pombe ehs1p is involved in maintaining cell wall integrity and in calcium uptake. Mol. Gen. Genet. 2000, 264, 173–183. [Google Scholar] [CrossRef]
- Lee, G.M.; Weng, F.; Cranley, J.; Rajasekhar, A.; Stoeckel, M.; Kane, T.; Tisi, R.; Wang, Y. The Ycx1 protein encoded by the yeast YDL206W gene plays a role in calcium and calcineurin signaling. J. Biol. Chem. 2023, 299, 104647. [Google Scholar] [CrossRef]
- Cortes, J.C.; Katoh-Fukui, R.; Moto, K.; Ribas, J.C.; Ishiguro, J. Schizosaccharomyces pombe Pmr1p is essential for cell wall integrity and is required for polarized cell growth and cytokinesis. Eukaryot. Cell 2004, 3, 1124–1135. [Google Scholar] [CrossRef]
- Facanha, A.L.; Appelgren, H.; Tabish, M.; Okorokov, L.; Ekwall, K. The endoplasmic reticulum cation P-type ATPase Cta4p is required for control of cell shape and microtubule dynamics. J. Cell Biol. 2002, 157, 1029–1039. [Google Scholar] [CrossRef]
- Furune, T.; Hashimoto, K.; Ishiguro, J. Characterization of a fission yeast P(5)-type ATPase homologue that is essential for Ca2+/Mn2+ homeostasis in the absence of P(2)-type ATPases. Genes. Genet. Syst. 2008, 83, 373–381. [Google Scholar] [CrossRef]
- Duffus, J.H.; Patterson, L.J. Control of cell division in yeast using the ionophore, A23187 with calcium and magnesium. Nature 1974, 251, 626–627. [Google Scholar] [CrossRef]
- Williams, D.A.; Fogarty, K.E.; Tsien, R.Y.; Fay, F.S. Calcium gradients in single smooth muscle cells revealed by the digital imaging microscope using Fura-2. Nature 1985, 318, 558–561. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.W.; Wardill, T.J.; Sun, Y.; Pulver, S.R.; Renninger, S.L.; Baohan, A.; Schreiter, E.R.; Kerr, R.A.; Orger, M.B.; Jayaraman, V.; et al. Ultrasensitive fluorescent proteins for imaging neuronal activity. Nature 2013, 499, 295–300. [Google Scholar] [CrossRef] [PubMed]
- Tian, L.; Hires, S.A.; Mao, T.; Huber, D.; Chiappe, M.E.; Chalasani, S.H.; Petreanu, L.; Akerboom, J.; McKinney, S.A.; Schreiter, E.R.; et al. Imaging neural activity in worms, flies and mice with improved GCaMP calcium indicators. Nat. Methods 2009, 6, 875–881. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Rozsa, M.; Liang, Y.; Bushey, D.; Wei, Z.; Zheng, J.; Reep, D.; Broussard, G.J.; Tsang, A.; Tsegaye, G.; et al. Fast and sensitive GCaMP calcium indicators for imaging neural populations. Nature 2023, 615, 884–891. [Google Scholar] [CrossRef]
- Poddar, A.; Sidibe, O.; Ray, A.; Chen, Q. Calcium spikes accompany cleavage furrow ingression and cell separation during fission yeast cytokinesis. Mol. Biol. Cell 2021, 32, 15–27. [Google Scholar] [CrossRef]
- Cerutti, L.; Simanis, V. Asymmetry of the spindle pole bodies and spg1p GAP segregation during mitosis in fission yeast. J. Cell Sci. 1999, 112 Pt 14, 2313–2321. [Google Scholar] [CrossRef]
- Lee, K.W.; Webb, S.E.; Miller, A.L. Ca2+ released via IP3 receptors is required for furrow deepening during cytokinesis in zebrafish embryos. Int. J. Dev. Biol. 2003, 47, 411–421. [Google Scholar]
- Palmer, C.P.; Aydar, E.; Djamgoz, M.B. A microbial TRP-like polycystic-kidney-disease-related ion channel gene. Biochem. J. 2005, 387, 211–219. [Google Scholar] [CrossRef]
- Hu, J.; Harris, P.C. Regulation of polycystin expression, maturation and trafficking. Cell. Signal. 2020, 72, 109630. [Google Scholar] [CrossRef]
- Chapin, H.C.; Caplan, M.J. The cell biology of polycystic kidney disease. J. Cell Biol. 2010, 191, 701–710. [Google Scholar] [CrossRef]
- Morris, Z.; Sinha, D.; Poddar, A.; Morris, B.; Chen, Q. Fission yeast TRP channel Pkd2p localizes to the cleavage furrow and regulates cell separation during cytokinesis. Mol. Biol. Cell 2019, 30, 1791–1804. [Google Scholar] [CrossRef] [PubMed]
- Aydar, E.; Palmer, C.P. Polycystic kidney disease channel and synaptotagmin homologues play roles in schizosaccharomyces pombe cell wall synthesis/repair and membrane protein trafficking. J. Membr. Biol. 2009, 229, 141–152. [Google Scholar] [CrossRef] [PubMed]
- Koyano, T.; Fujimoto, T.; Onishi, K.; Matsuyama, M.; Fukushima, M.; Kume, K. Pkd2, mutations linking to autosomal dominant polycystic kidney disease, localizes to the endoplasmic reticulum and regulates calcium signaling in fission yeast. Genes Cells 2023, 28, 811–820. [Google Scholar] [CrossRef] [PubMed]
- Malla, M.; Sinha, D.; Chowdhury, P.; Bisesi, B.T.; Chen, Q. The cytoplasmic tail of the mechanosensitive channel Pkd2 regulates its internalization and clustering in eisosomes. J. Cell Sci. 2023, 136, jcs.260598. [Google Scholar] [CrossRef]
- Sinha, D.; Ivan, D.; Gibbs, E.; Chetluru, M.; Goss, J.; Chen, Q. Fission yeast polycystin Pkd2p promotes cell size expansion and antagonizes the Hippo-related SIN pathway. J. Cell Sci. 2022, 135, jcs.259046. [Google Scholar] [CrossRef]
- Poddar, A.; Hsu, Y.Y.; Zhang, F.; Shamma, A.; Kreais, Z.; Muller, C.; Malla, M.; Ray, A.; Liu, A.; Chen, Q. Membrane stretching activates calcium-permeability of a putative channel Pkd2 during fission yeast cytokinesis. Mol. Biol. Cell 2022, 33, ar134. [Google Scholar] [CrossRef]
- Nakayama, Y.; Yoshimura, K.; Iida, H. Organellar mechanosensitive channels in fission yeast regulate the hypo-osmotic shock response. Nat. Commun. 2012, 3, 1020. [Google Scholar] [CrossRef]
- Nakayama, Y.; Hirata, A.; Iida, H. Mechanosensitive channels Msy1 and Msy2 are required for maintaining organelle integrity upon hypoosmotic shock in Schizosaccharomyces pombe. FEMS Yeast Res. 2014, 14, 992–994. [Google Scholar] [CrossRef]
- Mu, B.; Rutkowski, D.M.; Grenci, G.; Vavylonis, D.; Zhang, D. Ca2+-dependent vesicular and non-vesicular lipid transfer controls hypoosmotic plasma membrane expansion. bioRxiv 2024. [Google Scholar] [CrossRef]
- Ma, Y.; Sugiura, R.; Koike, A.; Ebina, H.; Sio, S.O.; Kuno, T. Transient receptor potential (TRP) and Cch1-Yam8 channels play key roles in the regulation of cytoplasmic Ca2+ in fission yeast. PLoS ONE 2011, 6, e22421. [Google Scholar] [CrossRef]
- Fluck, R.A.; Abraham, V.C.; Miller, A.L.; Jaffe, L.F. Calcium Buffer Injections Block Ooplasmic Segregation in Oryzias latipes (Medaka) Eggs. Biol. Bull. 1992, 183, 371–372. [Google Scholar] [CrossRef] [PubMed]
- Kuo, I.Y.; Ehrlich, B.E. Signaling in muscle contraction. Cold Spring Harb. Perspect. Biol. 2015, 7, a006023. [Google Scholar] [CrossRef]
- Wu, J.Q.; Pollard, T.D. Counting cytokinesis proteins globally and locally in fission yeast. Science 2005, 310, 310–314. [Google Scholar] [PubMed]
- Vavylonis, D.; Kovar, D.R.; O’Shaughnessy, B.; Pollard, T.D. Model of formin-associated actin filament elongation. Mol. Cell 2006, 21, 455–466. [Google Scholar]
- Kim, D.U.; Hayles, J.; Kim, D.; Wood, V.; Park, H.O.; Won, M.; Yoo, H.S.; Duhig, T.; Nam, M.; Palmer, G.; et al. Analysis of a genome-wide set of gene deletions in the fission yeast Schizosaccharomyces pombe. Nat. Biotechnol. 2010, 28, 617–623. [Google Scholar] [CrossRef]
- Cyert, M.S. Genetic analysis of calmodulin and its targets in Saccharomyces cerevisiae. Annu. Rev. Genet. 2001, 35, 647–672. [Google Scholar] [CrossRef]
- Takeda, T.; Yamamoto, M. Analysis and in vivo disruption of the gene coding for calmodulin in Schizosaccharomyces pombe. Proc. Natl. Acad. Sci. USA 1987, 84, 3580–3584. [Google Scholar] [CrossRef]
- Moser, M.J.; Lee, S.Y.; Klevit, R.E.; Davis, T.N. Ca2+ binding to calmodulin and its role in Schizosaccharomyces pombe as revealed by mutagenesis and NMR spectroscopy. J. Biol. Chem. 1995, 270, 20643–20652. [Google Scholar] [CrossRef]
- Moser, M.J.; Flory, M.R.; Davis, T.N. Calmodulin localizes to the spindle pole body of Schizosaccharomyces pombe and performs an essential function in chromosome segregation. J. Cell Sci. 1997, 110 Pt 15, 1805–1812. [Google Scholar]
- Flory, M.R.; Morphew, M.; Joseph, J.D.; Means, A.R.; Davis, T.N. Pcp1p, an Spc110p-related calmodulin target at the centrosome of the fission yeast Schizosaccharomyces pombe. Cell Growth Differ. 2002, 13, 47–58. [Google Scholar]
- Fong, C.S.; Sato, M.; Toda, T. Fission yeast Pcp1 links polo kinase-mediated mitotic entry to gamma-tubulin-dependent spindle formation. EMBO J. 2010, 29, 120–130. [Google Scholar] [CrossRef] [PubMed]
- Lee, W.L.; Bezanilla, M.; Pollard, T.D. Fission yeast myosin-I, Myo1p, stimulates actin assembly by Arp2/3 complex and shares functions with WASp. J. Cell Biol. 2000, 151, 789–800. [Google Scholar] [CrossRef] [PubMed]
- Win, T.Z.; Gachet, Y.; Mulvihill, D.P.; May, K.M.; Hyams, J.S. Two type V myosins with non-overlapping functions in the fission yeast Schizosaccharomyces pombe: Myo52 is concerned with growth polarity and cytokinesis, Myo51 is a component of the cytokinetic actin ring. J. Cell Sci. 2001, 114, 69–79. [Google Scholar] [PubMed]
- Tang, Q.; Billington, N.; Krementsova, E.B.; Bookwalter, C.S.; Lord, M.; Trybus, K.M. A single-headed fission yeast myosin V transports actin in a tropomyosin-dependent manner. J. Cell Biol. 2016, 214, 167–179. [Google Scholar] [CrossRef]
- Itadani, A.; Nakamura, T.; Shimoda, C. Localization of type I myosin and F-actin to the leading edge region of the forespore membrane in Schizosaccharomyces pombe. Cell Struct. Funct. 2006, 31, 181–195. [Google Scholar] [CrossRef]
- Li, H.; Rao, A.; Hogan, P.G. Interaction of calcineurin with substrates and targeting proteins. Trends Cell Biol. 2011, 21, 91–103. [Google Scholar] [CrossRef]
- Yoshida, T.; Toda, T.; Yanagida, M. A calcineurin-like gene ppb1+ in fission yeast: Mutant defects in cytokinesis, cell polarity, mating and spindle pole body positioning. J. Cell Sci. 1994, 107 Pt 7, 1725–1735. [Google Scholar] [CrossRef]
- Martin-Garcia, R.; Arribas, V.; Coll, P.M.; Pinar, M.; Viana, R.A.; Rincon, S.A.; Correa-Bordes, J.; Ribas, J.C.; Perez, P. Paxillin-Mediated Recruitment of Calcineurin to the Contractile Ring Is Required for the Correct Progression of Cytokinesis in Fission Yeast. Cell Rep. 2018, 25, 772–783.e774. [Google Scholar] [CrossRef]
- Mangione, M.C.; Snider, C.E.; Gould, K.L. The intrinsically disordered region of the cytokinetic F-BAR protein Cdc15 performs a unique essential function in maintenance of cytokinetic ring integrity. Mol. Biol. Cell 2019, 30, 2790–2801. [Google Scholar] [CrossRef]
- Hirayama, S.; Sugiura, R.; Lu, Y.; Maeda, T.; Kawagishi, K.; Yokoyama, M.; Tohda, H.; Giga-Hama, Y.; Shuntoh, H.; Kuno, T. Zinc finger protein Prz1 regulates Ca2+ but not Cl- homeostasis in fission yeast. Identification of distinct branches of calcineurin signaling pathway in fission yeast. J. Biol. Chem. 2003, 278, 18078–18084. [Google Scholar] [CrossRef]
- Matsuo, Y.; Kawamukai, M. cAMP-dependent protein kinase involves calcium tolerance through the regulation of Prz1 in Schizosaccharomyces pombe. Biosci. Biotechnol. Biochem. 2017, 81, 231–241. [Google Scholar] [CrossRef] [PubMed]
- Paoletti, A.; Bordes, N.; Haddad, R.; Schwartz, C.L.; Chang, F.; Bornens, M. Fission yeast cdc31p is a component of the half-bridge and controls SPB duplication. Mol. Biol. Cell 2003, 14, 2793–2808. [Google Scholar] [CrossRef] [PubMed]
- McCollum, D.; Balasubramanian, M.K.; Pelcher, L.E.; Hemmingsen, S.M.; Gould, K.L. Schizosaccharomyces pombe cdc4+ gene encodes a novel EF-hand protein essential for cytokinesis. J. Cell Biol. 1995, 130, 651–660. [Google Scholar]
- Le Goff, X.; Motegi, F.; Salimova, E.; Mabuchi, I.; Simanis, V. The S. pombe rlc1 gene encodes a putative myosin regulatory light chain that binds the type II myosins myo3p and myo2p. J. Cell Sci. 2000, 113 Pt 23, 4157–4163. [Google Scholar]
- Hamasaki-Katagiri, N.; Molchanova, T.; Takeda, K.; Ames, J.B. Fission yeast homolog of neuronal calcium sensor-1 (Ncs1p) regulates sporulation and confers calcium tolerance. J. Biol. Chem. 2004, 279, 12744–12754. [Google Scholar] [CrossRef]
- Willet, A.H.; Turner, L.A.; Park, J.S.; Ren, L.; Snider, C.E.; Gould, K.L. Characterization of Pik1 function in fission yeast reveals its conserved role in lipid synthesis and not cytokinesis. J. Cell Sci. 2023, 136, jcs261415. [Google Scholar] [CrossRef]
- Hendricks, K.B.; Wang, B.Q.; Schnieders, E.A.; Thorner, J. Yeast homologue of neuronal frequenin is a regulator of phosphatidylinositol-4-OH kinase. Nat. Cell Biol. 1999, 1, 234–241. [Google Scholar] [CrossRef]
- Ohya, Y.; Kawasaki, H.; Suzuki, K.; Londesborough, J.; Anraku, Y. Two yeast genes encoding calmodulin-dependent protein kinases. Isolation, sequencing and bacterial expressions of CMK1 and CMK2. J. Biol. Chem. 1991, 266, 12784–12794. [Google Scholar]
- Rasmussen, C.D. Cloning of a calmodulin kinase I homologue from Schizosaccharomyces pombe. J. Biol. Chem. 2000, 275, 685–690. [Google Scholar] [CrossRef]
- Cisneros-Barroso, E.; Yance-Chavez, T.; Kito, A.; Sugiura, R.; Gomez-Hierro, A.; Gimenez-Zaragoza, D.; Aligue, R. Negative feedback regulation of calcineurin-dependent Prz1 transcription factor by the CaMKK-CaMK1 axis in fission yeast. Nucleic Acids Res. 2014, 42, 9573–9587. [Google Scholar] [CrossRef]
- Sanchez-Piris, M.; Posas, F.; Alemany, V.; Winge, I.; Hidalgo, E.; Bachs, O.; Aligue, R. The serine/threonine kinase Cmk2 is required for oxidative stress response in fission yeast. J. Biol. Chem. 2002, 277, 17722–17727. [Google Scholar] [CrossRef] [PubMed]
- Rhind, N.; Chen, Z.; Yassour, M.; Thompson, D.A.; Haas, B.J.; Habib, N.; Wapinski, I.; Roy, S.; Lin, M.F.; Heiman, D.I.; et al. Comparative functional genomics of the fission yeasts. Science 2011, 332, 930–936. [Google Scholar] [CrossRef] [PubMed]
- Rigamonti, M.; Groppi, S.; Belotti, F.; Ambrosini, R.; Filippi, G.; Martegani, E.; Tisi, R. Hypotonic stress-induced calcium signaling in Saccharomyces cerevisiae involves TRP-like transporters on the endoplasmic reticulum membrane. Cell Calcium 2015, 57, 57–68. [Google Scholar] [CrossRef] [PubMed]
- Geiser, J.R.; Sundberg, H.A.; Chang, B.H.; Muller, E.G.; Davis, T.N. The essential mitotic target of calmodulin is the 110-kilodalton component of the spindle pole body in Saccharomyces cerevisiae. Mol. Cell Biol. 1993, 13, 7913–7924. [Google Scholar] [CrossRef]
- Stathopoulos, A.M.; Cyert, M.S. Calcineurin acts through the CRZ1/TCN1-encoded transcription factor to regulate gene expression in yeast. Genes Dev. 1997, 11, 3432–3444. [Google Scholar] [CrossRef]
- Geiser, J.R.; van Tuinen, D.; Brockerhoff, S.E.; Neff, M.M.; Davis, T.N. Can calmodulin function without binding calcium? Cell 1991, 65, 949–959. [Google Scholar]
- Palmer, C.P.; Zhou, X.L.; Lin, J.; Loukin, S.H.; Kung, C.; Saimi, Y. A TRP homolog in Saccharomyces cerevisiae forms an intracellular Ca2+-permeable channel in the yeast vacuolar membrane. Proc. Natl. Acad. Sci. USA 2001, 98, 7801–7805. [Google Scholar] [CrossRef]
- Denis, V.; Cyert, M.S. Internal Ca2+ release in yeast is triggered by hypertonic shock and mediated by a TRP channel homologue. J. Cell Biol. 2002, 156, 29–34. [Google Scholar] [CrossRef]
- Fischer, M.; Schnell, N.; Chattaway, J.; Davies, P.; Dixon, G.; Sanders, D. The Saccharomyces cerevisiae CCH1 gene is involved in calcium influx and mating. FEBS Lett. 1997, 419, 259–262. [Google Scholar] [CrossRef]
- Iida, H.; Nakamura, H.; Ono, T.; Okumura, M.S.; Anraku, Y. MID1, a novel Saccharomyces cerevisiae gene encoding a plasma membrane protein, is required for Ca2+ influx and mating. Mol. Cell Biol. 1994, 14, 8259–8271. [Google Scholar] [CrossRef]
- Liu, S.; Chen, M.; Wang, Y.; Lei, Y.; Huang, T.; Zhang, Y.; Lam, S.M.; Li, H.; Qi, S.; Geng, J.; et al. The ER calcium channel Csg2 integrates sphingolipid metabolism with autophagy. Nat. Commun. 2023, 14, 3725. [Google Scholar] [CrossRef] [PubMed]
- D’Hooge, P.; Coun, C.; Van Eyck, V.; Faes, L.; Ghillebert, R.; Marien, L.; Winderickx, J.; Callewaert, G. Ca2+ homeostasis in the budding yeast Saccharomyces cerevisiae: Impact of ER/Golgi Ca2+ storage. Cell Calcium 2015, 58, 226–235. [Google Scholar] [CrossRef] [PubMed]
- Carbo, N.; Tarkowski, N.; Ipina, E.P.; Dawson, S.P.; Aguilar, P.S. Sexual pheromone modulates the frequency of cytosolic Ca2+ bursts in Saccharomyces cerevisiae. Mol. Biol. Cell 2017, 28, 501–510. [Google Scholar] [CrossRef] [PubMed]
- Spolaor, S.; Rovetta, M.; Nobile, M.S.; Cazzaniga, P.; Tisi, R.; Besozzi, D. Modeling Calcium Signaling in S. cerevisiae Highlights the Role and Regulation of the Calmodulin-Calcineurin Pathway in Response to Hypotonic Shock. Front. Mol. Biosci. 2022, 9, 856030. [Google Scholar] [CrossRef]
- Martin, D.C.; Kim, H.; Mackin, N.A.; Maldonado-Baez, L.; Evangelista, C.C., Jr.; Beaudry, V.G.; Dudgeon, D.D.; Naiman, D.Q.; Erdman, S.E.; Cunningham, K.W. New regulators of a high affinity Ca2+ influx system revealed through a genome-wide screen in yeast. J. Biol. Chem. 2011, 286, 10744–10754. [Google Scholar] [CrossRef]
- Deng, L.; Sugiura, R.; Takeuchi, M.; Suzuki, M.; Ebina, H.; Takami, T.; Koike, A.; Iba, S.; Kuno, T. Real-time monitoring of calcineurin activity in living cells: Evidence for two distinct Ca2+-dependent pathways in fission yeast. Mol. Biol. Cell 2006, 17, 4790–4800. [Google Scholar] [CrossRef]
- Protchenko, O.; Rodriguez-Suarez, R.; Androphy, R.; Bussey, H.; Philpott, C.C. A screen for genes of heme uptake identifies the FLC family required for import of FAD into the endoplasmic reticulum. J. Biol. Chem. 2006, 281, 21445–21457. [Google Scholar] [CrossRef]
- Suzuki, C.; Shimma, Y.I. P-type ATPase spf1 mutants show a novel resistance mechanism for the killer toxin SMKT. Mol. Microbiol. 1999, 32, 813–823. [Google Scholar] [CrossRef]
- Cunningham, K.W.; Fink, G.R. Calcineurin-dependent growth control in Saccharomyces cerevisiae mutants lacking PMC1, a homolog of plasma membrane Ca2+ ATPases. J. Cell Biol. 1994, 124, 351–363. [Google Scholar] [CrossRef]
- Ohya, Y.; Anraku, Y. A galactose-dependent cmd1 mutant of Saccharomyces cerevisiae: Involvement of calmodulin in nuclear division. Curr. Genet. 1989, 15, 113–120. [Google Scholar] [CrossRef]
- Cyert, M.S.; Kunisawa, R.; Kaim, D.; Thorner, J. Yeast has homologs (CNA1 and CNA2 gene products) of mammalian calcineurin, a calmodulin-regulated phosphoprotein phosphatase. Proc. Natl. Acad. Sci. USA 1991, 88, 7376–7380. [Google Scholar] [CrossRef] [PubMed]
- Sio, S.O.; Suehiro, T.; Sugiura, R.; Takeuchi, M.; Mukai, H.; Kuno, T. The role of the regulatory subunit of fission yeast calcineurin for in vivo activity and its relevance to FK506 sensitivity. J. Biol. Chem. 2005, 280, 12231–12238. [Google Scholar] [CrossRef] [PubMed]
- Spang, A.; Courtney, I.; Fackler, U.; Matzner, M.; Schiebel, E. The calcium-binding protein cell division cycle 31 of Saccharomyces cerevisiae is a component of the half bridge of the spindle pole body. J. Cell Biol. 1993, 123, 405–416. [Google Scholar] [CrossRef]
- Stevens, R.C.; Davis, T.N. Mlc1p is a light chain for the unconventional myosin Myo2p in Saccharomyces cerevisiae. J. Cell Biol. 1998, 142, 711–722. [Google Scholar] [CrossRef]
- Luo, J.; Vallen, E.A.; Dravis, C.; Tcheperegine, S.E.; Drees, B.; Bi, E. Identification and functional analysis of the essential and regulatory light chains of the only type II myosin Myo1p in Saccharomyces cerevisiae. J. Cell Biol. 2004, 165, 843–855. [Google Scholar] [CrossRef]
- Liu, X.; Wang, P.; Fu, J.; Lv, D.; Chen, D.; Li, Y.; Ma, W. Two-photon fluorescence real-time imaging on the development of early mouse embryo by stages. J. Microsc. 2011, 241, 212–218. [Google Scholar] [CrossRef]
- Jaffe, L.F.; Creton, R. On the conservation of calcium wave speeds. Cell Calcium 1998, 24, 1–8. [Google Scholar] [CrossRef]
- Chan, C.M.; Chen, Y.; Hung, T.S.; Miller, A.L.; Shipley, A.M.; Webb, S.E. Inhibition of SOCE disrupts cytokinesis in zebrafish embryos via inhibition of cleavage furrow deepening. Int. J. Dev. Biol. 2015, 59, 289–301. [Google Scholar] [CrossRef]
- Trebak, M.; Putney, J.W., Jr. ORAI Calcium Channels. Physiology 2017, 32, 332–342. [Google Scholar] [CrossRef]
- Carrasco, S.; Meyer, T. STIM proteins and the endoplasmic reticulum-plasma membrane junctions. Annu. Rev. Biochem. 2011, 80, 973–1000. [Google Scholar] [CrossRef]
- Collins, S.R.; Meyer, T. Evolutionary origins of STIM1 and STIM2 within ancient Ca2+ signaling systems. Trends Cell Biol. 2011, 21, 202–211. [Google Scholar] [CrossRef]
- Nauli, S.M.; Alenghat, F.J.; Luo, Y.; Williams, E.; Vassilev, P.; Li, X.; Elia, A.E.; Lu, W.; Brown, E.M.; Quinn, S.J.; et al. Polycystins 1 and 2 mediate mechanosensation in the primary cilium of kidney cells. Nat. Genet. 2003, 33, 129–137. [Google Scholar] [CrossRef]
- Djenoune, L.; Mahamdeh, M.; Truong, T.V.; Nguyen, C.T.; Fraser, S.E.; Brueckner, M.; Howard, J.; Yuan, S. Cilia function as calcium-mediated mechanosensors that instruct left-right asymmetry. Science 2023, 379, 71–78. [Google Scholar] [CrossRef] [PubMed]
- Webb, S.E.; Lee, K.W.; Karplus, E.; Miller, A.L. Localized calcium transients accompany furrow positioning, propagation, and deepening during the early cleavage period of zebrafish embryos. Dev. Biol. 1997, 192, 78–92. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.Y.; Chen, Y.; Dai, G.; Chen, J.; Sun, X.M.; Wen, C.J.; Zhao, D.H.; Chang, D.C.; Li, C.J. The association of calmodulin with central spindle regulates the initiation of cytokinesis in HeLa cells. Int. J. Biochem. Cell Biol. 2004, 36, 1562–1572. [Google Scholar] [CrossRef] [PubMed]
- Chircop, M.; Malladi, C.S.; Lian, A.T.; Page, S.L.; Zavortink, M.; Gordon, C.P.; McCluskey, A.; Robinson, P.J. Calcineurin activity is required for the completion of cytokinesis. Cell. Mol. Life Sci. CMLS 2010, 67, 3725–3737. [Google Scholar] [CrossRef]
- Fan, J.Q.; Deng, X.L.; Feng, B.W.; Wang, J.F.; Yu, Y.; Lv, H. Cnb1 involved in cytokinesis in Schizosaccharomyces pombe. Yi Chuan Hered. 2013, 35, 1030–1039. [Google Scholar] [CrossRef]
- Smith, J.L.; Silveira, L.A.; Spudich, J.A. Myosin light chain kinase (MLCK) gene disruption in Dictyostelium: A role for MLCK-A in cytokinesis and evidence for multiple MLCKs. Proc. Natl. Acad. Sci. USA 1996, 93, 12321–12326. [Google Scholar] [CrossRef]
- Benaud, C.; Le Dez, G.; Mironov, S.; Galli, F.; Reboutier, D.; Prigent, C. Annexin A2 is required for the early steps of cytokinesis. EMBO Rep. 2015, 16, 481–489. [Google Scholar] [CrossRef]
- Li, W.M.; Webb, S.E.; Lee, K.W.; Miller, A.L. Recruitment and SNARE-mediated fusion of vesicles in furrow membrane remodeling during cytokinesis in zebrafish embryos. Exp. Cell Res. 2006, 312, 3260–3275. [Google Scholar] [CrossRef]
- Li, C.J.; Heim, R.; Lu, P.; Pu, Y.; Tsien, R.Y.; Chang, D.C. Dynamic redistribution of calmodulin in HeLa cells during cell division as revealed by a GFP-calmodulin fusion protein technique. J. Cell Sci. 1999, 112 Pt 10, 1567–1577. [Google Scholar] [CrossRef] [PubMed]
- Li, W.M.; Webb, S.E.; Chan, C.M.; Miller, A.L. Multiple roles of the furrow deepening Ca2+ transient during cytokinesis in zebrafish embryos. Dev. Biol. 2008, 316, 228–248. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Vien, T.; Duan, J.; Sheu, S.H.; DeCaen, P.G.; Clapham, D.E. Polycystin-2 is an essential ion channel subunit in the primary cilium of the renal collecting duct epithelium. eLife 2018, 7, e33183. [Google Scholar] [CrossRef] [PubMed]
- Su, Q.; Hu, F.; Ge, X.; Lei, J.; Yu, S.; Wang, T.; Zhou, Q.; Mei, C.; Shi, Y. Structure of the human PKD1-PKD2 complex. Science 2018, 361, eaat9819. [Google Scholar] [CrossRef]
- Shen, P.S.; Yang, X.; DeCaen, P.G.; Liu, X.; Bulkley, D.; Clapham, D.E.; Cao, E. The Structure of the Polycystic Kidney Disease Channel PKD2 in Lipid Nanodiscs. Cell 2016, 167, 763–773.e711. [Google Scholar] [CrossRef]
- Chowdhury, P.; Sinha, D.; Poddar, A.; Chetluru, M.; Chen, Q. The Mechanosensitive Pkd2 Channel Modulates the Recruitment of Myosin II and Actin to the Cytokinetic Contractile Ring. J. Fungi 2024, 10, 455. [Google Scholar] [CrossRef]
- Levin, M. Molecular bioelectricity: How endogenous voltage potentials control cell behavior and instruct pattern regulation in vivo. Mol. Biol. Cell 2014, 25, 3835–3850. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, Q. Regulation of Yeast Cytokinesis by Calcium. J. Fungi 2025, 11, 278. https://doi.org/10.3390/jof11040278
Chen Q. Regulation of Yeast Cytokinesis by Calcium. Journal of Fungi. 2025; 11(4):278. https://doi.org/10.3390/jof11040278
Chicago/Turabian StyleChen, Qian. 2025. "Regulation of Yeast Cytokinesis by Calcium" Journal of Fungi 11, no. 4: 278. https://doi.org/10.3390/jof11040278
APA StyleChen, Q. (2025). Regulation of Yeast Cytokinesis by Calcium. Journal of Fungi, 11(4), 278. https://doi.org/10.3390/jof11040278