
The Dissemination of a Single Staphylococcusaureus Strain, Spa-t2873, as the Predominant Cause of Bovine Mastitis in Israeli Dairy Farms


Abstract
:Simple Summary
Abstract
1. Introduction
2. Methods
2.1. Study Design
2.2. Microbiological Methods and Spa Typing
2.3. Next-Generation Sequencing and Analysis
2.4. Epidemiological Study of the t2873 Strain Transmission in Farm ‘A’
3. Results
3.1. S. aureus Prevalence in Israeli Dairy Farms
3.2. Molecular and Microbiological Features of S. aureus Isolates
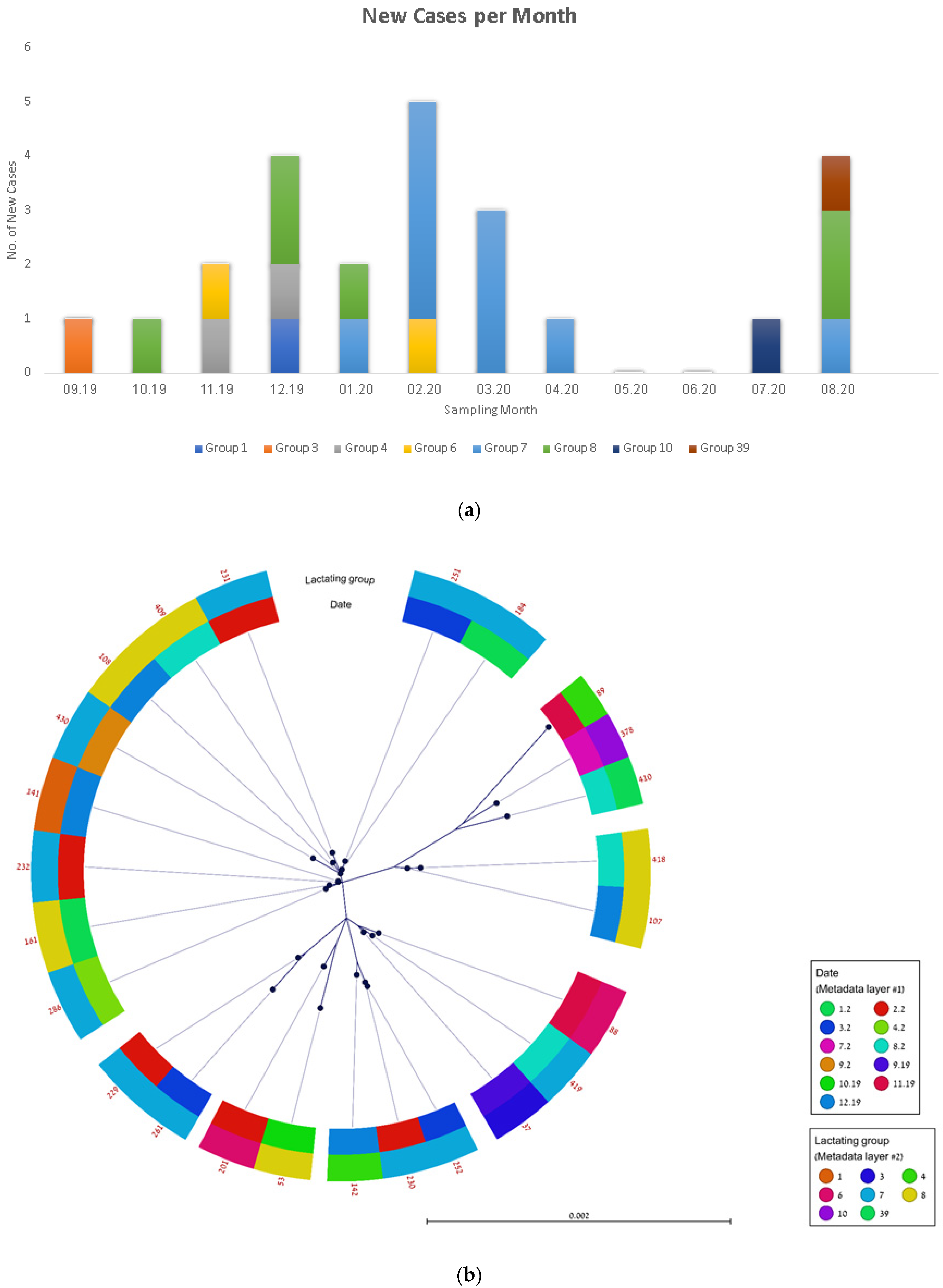
3.3. Molecular Epidemiological Transmission Analysis of Spa t2873 in Farm ‘A’
3.4. Molecular Features of the Spa t2873 Strain
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Petersson-Wolfe, C.; Mullarky, I.K.; Jones, G. Staphylococcus Aureus Mastitis: Cause, Detection, and Control. Va. Coop. Ext. 2010, 404, 7. Available online: http://hdl.handle.net/10919/48390 (accessed on 2 June 2022).
- Asadollahi, P.; Farahani, N.N.; Mirzaii, M.; Khoramrooz, S.S.; Van Belkum, A.; Asadollahi, K.; Dadashi, M.; Darban-Sarokhalil, D. Distribution of the Most Prevalent Spa Types among Clinical Isolates of Methicillin-Resistant and -Susceptible Staphylococcus aureus around the World: A Review. Front. Microbiol. 2018, 9, 163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- International Dairy Federation; Falk, R.; Blum, S.E.; Shwimmer, A.; Friedman, S. IDF Animal Health Report; International Dairy Federation: Schaerbeek, Belgium, 2018. [Google Scholar]
- Wassenberg, M.; Bootsma, M.; Troelstra, A.; Kluytmans, J.; Bonten, M. Transmissibility of livestock-associated methicillin-resistant Staphylococcus aureus (ST398) in Dutch hospitals. Clin. Microbiol. Infect. 2011, 17, 316–319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sheet, O.H.; Grabowski, N.T.; Klein, G.; Reich, F.; Abdulmawjood, A. Characterisation of mecA gene negative Staphylococcus aureus isolated from bovine mastitis milk from Northern Germany. Folia Microbiol. 2019, 64, 845–855. [Google Scholar] [CrossRef] [PubMed]
- Naushad, S.; Nobrega, D.B.; Naqvi, S.A.; Barkema, H.W.; De Buck, J. Genomic Analysis of Bovine Staphylococcus aureus Isolates from Milk to Elucidate Diversity and Determine the Distributions of Antimicrobial and Virulence Genes and Their Association with Mastitis. mSystems 2020, 5, e00063-20. [Google Scholar] [CrossRef] [PubMed]
- Cvetnić, L.; Samardžija, M.; Duvnjak, S.; Habrun, B.; Cvetnić, M.; Tkalec, V.J.; Đuričić, D.; Benić, M. Multi Locus Sequence Typing and spa Typing of Staphylococcus aureus Isolated from the Milk of Cows with Subclinical Mastitis in Croatia. Microorganisms 2021, 9, 725. [Google Scholar] [CrossRef] [PubMed]
- Staphylococci, L. Laboratory Handbook on Bovine Mastitis; National Mastitis Council: New Prague, MN, USA, 2017; pp. 43–52. [Google Scholar]
- Adler, A.; Chmelnitsky, I.; Shitrit, P.; Sprecher, H.; Navon-Venezia, S.; Embon, A.; Khabra, E.; Paitan, Y.; Keren, L.; Halperin, E.; et al. Molecular Epidemiology of Methicillin-Resistant Staphylococcus aureus in Israel: Dissemination of Global Clones and Unique Features. J. Clin. Microbiol. 2012, 50, 134–137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schechner, V.; Levytskyi, K.; Shalom, O.; Yalek, A.; Adler, A. A hospital-wide outbreak of IMI-17-producing Enterobacter ludwigii in an Israeli hospital. Antimicrob. Resist. Infect. Control 2021, 10, 166. [Google Scholar] [CrossRef] [PubMed]
- Kaas, R.S.; Leekitcharoenphon, P.; Aarestrup, F.M.; Lund, O. Solving the Problem of Comparing Whole Bacterial Genomes across Different Sequencing Platforms. PLoS ONE 2014, 9, e104984. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miltgen, G.; Bonnin, R.A.; Avril, C.; Benoit-Cattin, T.; Martak, D.; Leclaire, A.; Traversier, N.; Roquebert, B.; Jaffar-Bandjee, M.-C.; Lugagne, N.; et al. Outbreak of IMI-1 carbapenemase-producing colistin-resistant Enterobacter cloacae on the French island of Mayotte (Indian Ocean). Int. J. Antimicrob. Agents 2018, 52, 416–420. [Google Scholar] [CrossRef] [PubMed]
- Riekerink, R.O.; Barkema, H.; Kelton, D.; Scholl, D. Incidence Rate of Clinical Mastitis on Canadian Dairy Farms. J. Dairy Sci. 2008, 91, 1366–1377. [Google Scholar] [CrossRef] [PubMed]
- Abebe, R.; Hatiya, H.; Abera, M.; Megersa, B.; Asmare, K. Bovine mastitis: Prevalence, risk factors and isolation of Staphylococcus aureus in dairy herds at Hawassa milk shed, South Ethiopia. BMC Vet. Res. 2016, 12, 270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cortimiglia, C.; Luini, M.; Bianchini, V.; Marzagalli, L.; Vezzoli, F.; Avisani, D.; Bertoletti, M.; Ianzano, A.; Franco, A.; Battisti, A. Short Report Prevalence of Staphylococcus aureus and of methicillin-resistant S. aureus clonal complexes in bulk tank milk from dairy cattle herds in Lombardy Region (Northern Italy). Epidemiol. Infect. 2016, 144, 3046–3051. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hasman, H.; Moodley, A.; Guardabassi, L.; Stegger, M.; Skov, R.; Aarestrup, F. spa type distribution in Staphylococcus aureus originating from pigs, cattle and poultry. Vet. Microbiol. 2010, 141, 326–331. [Google Scholar] [CrossRef] [PubMed]
- Bar-Gal, G.K.; Blum, S.; Hadas, L.; Ehricht, R.; Monecke, S.; Leitner, G. Host-specificity of Staphylococcus aureus causing intramammary infections in dairy animals assessed by genotyping and virulence genes. Vet. Microbiol. 2015, 176, 143–154. [Google Scholar] [CrossRef] [PubMed]
- Hoekstra, J.; Zomer, A.L.; Rutten, V.P.M.G.; Benedictus, L.; Stegeman, A.; Spaninks, M.P.; Bennedsgaard, T.W.; Biggs, A.; De Vliegher, S.; Mateo, D.H.; et al. Genomic analysis of European bovine Staphylococcus aureus from clinical versus subclinical mastitis. Sci. Rep. 2020, 10, 18172. [Google Scholar] [CrossRef] [PubMed]
- Unnerstad, H.E.; Mieziewska, K.; Börjesson, S.; Hedbäck, H.; Strand, K.; Hallgren, T.; Landin, H.; Skarin, J.; Bengtsson, B. Suspected transmission and subsequent spread of MRSA from farmer to dairy cows. Vet. Microbiol. 2018, 225, 114–119. [Google Scholar] [CrossRef] [PubMed]
- Lienen, T.; Schnitt, A.; Hammerl, J.A.; Maurischat, S.; Tenhagen, B.-A. Genomic Distinctions of LA-MRSA ST398 on Dairy Farms from Different German Federal States with a Low Risk of Severe Human Infections. Front. Microbiol. 2021, 11, 575321. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
| Spa Type | Farms (n) | Isolates (n) | Sample Type 1 (n) |
|---|---|---|---|
| 11 | 3 | 11 | I (10), T (1) |
| 34 | 6 | 9 | I (7), T (2) |
| 127 | 2 | 2 | I (1), T (1) |
| 355 | 2 | 3 | I (3) |
| 529 | 22 | 46 | I (36), T (10) |
| 543 | 1 | 9 | I (9) |
| 937 | 2 | 2 | I (2) |
| 1250 | 1 | 13 | I (13) |
| 2873 | 112 | 284 | I (259), T (25) |
| 2970 | 1 | 5 | I (4), T (1) |
| 9303 | 12 | 34 | I (32), T (2) |
| 11253 | 1 | 7 | I (7) |
| 16822 | 2 | 2 | I (1), T (1) |
| 19690 | 2 | 2 | I (2) |
| 19693 | 1 | 2 | I (2) |
| Cow Number | Isolation of S. aureus | Presumptive Source | ||||
|---|---|---|---|---|---|---|
| Date | Location | Cluster | Order | Location | Cow Number | |
| 37 | 22.09.19 | 3 | A | 1 | 39 | - |
| 88 | 24.11.19 | 6 | 2 | 39 | 37 | |
| 419 | 17.08.20 | 7 | 3 | 7 | 88 | |
| 53 | 23.10.19 | 8 | B | 1 | 6 | - |
| 201 | 04.02.20 | 6 | 2 | 6 | 53 | |
| 89 | 24.11.19 | 4 | C | 1 | Unknown | |
| 378 | 13.07.20 | 10 | 2 | Unknown | ||
| 410 | 04.08.20 | 39 | 3 | Unknown | ||
| 107 | 10.12.19 | 8 | D | 1 | 8 | - |
| 418 | 17.08.20 | 8 | 2 | 8 | 107 | |
| 108 | 10.12.19 | 8 | E | 1 | 1 | 141 |
| 141 | 30.12.19 | 1 | 2 | 4 | 161 | |
| 161 | 14.01.20 | 8 | 3 | 4 | 141 | |
| 8 | 108 | |||||
| 231 | 19.02.20 | 7 | 4 | 7 | 141/232 | |
| 232 | 19.02.20 | 7 | 5 | 7 | 141/231 | |
| 286 | 14.04.20 | 7 | 6 | 1 | 141 | |
| 7 | 231/232 | |||||
| 409 | 04.08.20 | 8 | 7 | 8 | 161 | |
| 430 | 06.09.20 | 7 | 8 | 7 | 286 | |
| 142 | 30.12.19 | 4 | F | 1 | 4 | - |
| 230 | 19.02.20 | 7 | 2 | 3 | 142 | |
| 252 | 08.03.20 | 7 | 3 | 7 | 230 | |
| 184 | 26.01.20 | 7 | G | 1 | 2/7 | - |
| 251 | 08.03.20 | 7 | 2 | 2/7 | 184 | |
| 229 | 19.02.20 | 7 | H | 1 | 1/7 | - |
| 261 | 12.03.20 | 7 | 2 | 1/7 | 261 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shwimmer, A.; Falk, R.; De-Levie, T.K.; Lasnoy, M.; Levytskyi, K.; Adler, A. The Dissemination of a Single Staphylococcusaureus Strain, Spa-t2873, as the Predominant Cause of Bovine Mastitis in Israeli Dairy Farms. Vet. Sci. 2022, 9, 371. https://doi.org/10.3390/vetsci9070371
Shwimmer A, Falk R, De-Levie TK, Lasnoy M, Levytskyi K, Adler A. The Dissemination of a Single Staphylococcusaureus Strain, Spa-t2873, as the Predominant Cause of Bovine Mastitis in Israeli Dairy Farms. Veterinary Sciences. 2022; 9(7):371. https://doi.org/10.3390/vetsci9070371
Chicago/Turabian StyleShwimmer, Adin, Rama Falk, Tamar K. De-Levie, Michal Lasnoy, Katia Levytskyi, and Amos Adler. 2022. "The Dissemination of a Single Staphylococcusaureus Strain, Spa-t2873, as the Predominant Cause of Bovine Mastitis in Israeli Dairy Farms" Veterinary Sciences 9, no. 7: 371. https://doi.org/10.3390/vetsci9070371
APA StyleShwimmer, A., Falk, R., De-Levie, T. K., Lasnoy, M., Levytskyi, K., & Adler, A. (2022). The Dissemination of a Single Staphylococcusaureus Strain, Spa-t2873, as the Predominant Cause of Bovine Mastitis in Israeli Dairy Farms. Veterinary Sciences, 9(7), 371. https://doi.org/10.3390/vetsci9070371