
Effect of Copper, Zinc, and Selenium on the Migration of Bovine Neutrophils


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals, Blood Collection and Neutrophil Isolation
2.2. Detection of Neutrophil Survival
2.3. Cell Culture and Establishment of Epithelial Layer
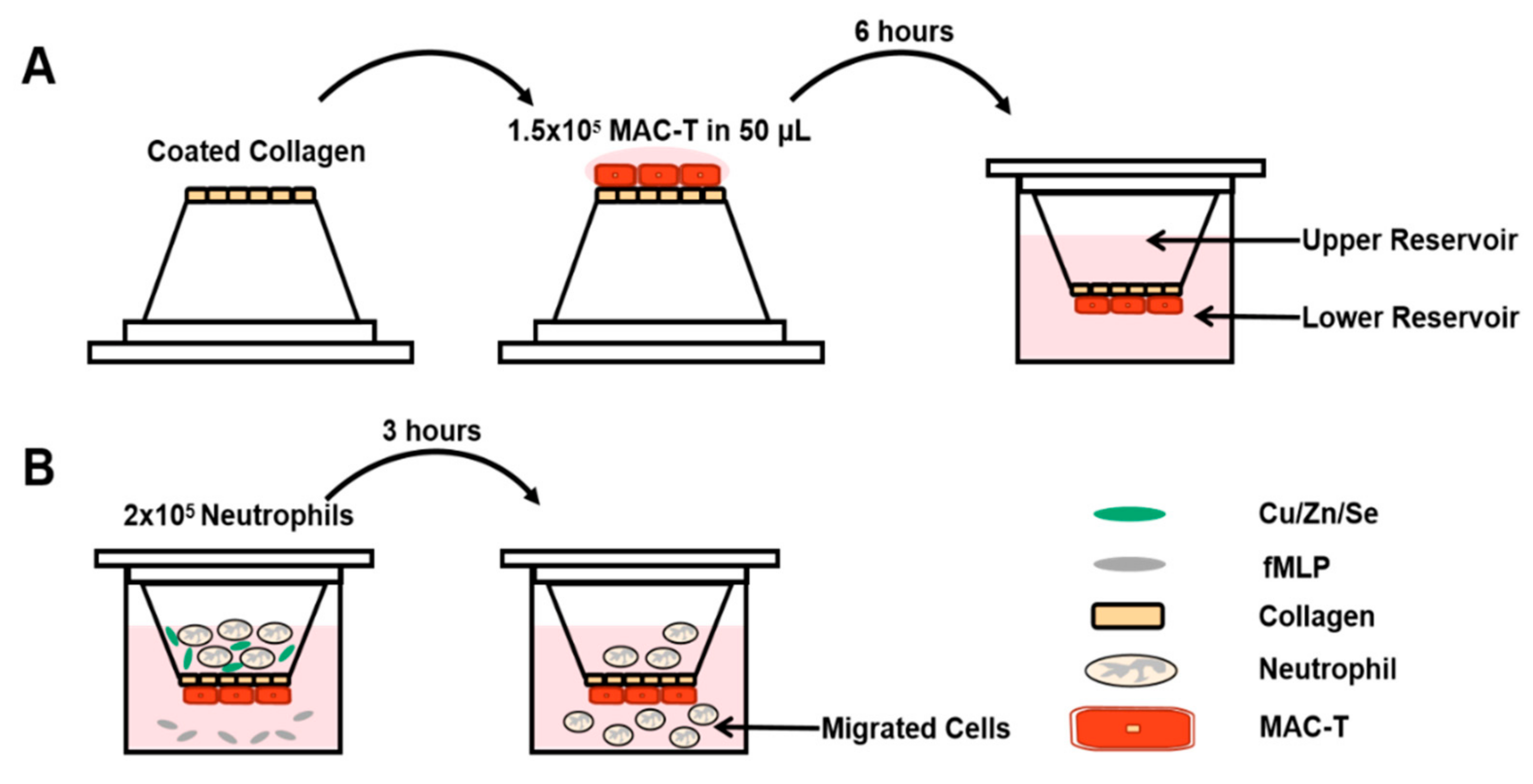
2.4. Neutrophil Migration Assay
2.5. Statistical Analysis
3. Results
3.1. Neutrophil Isolation
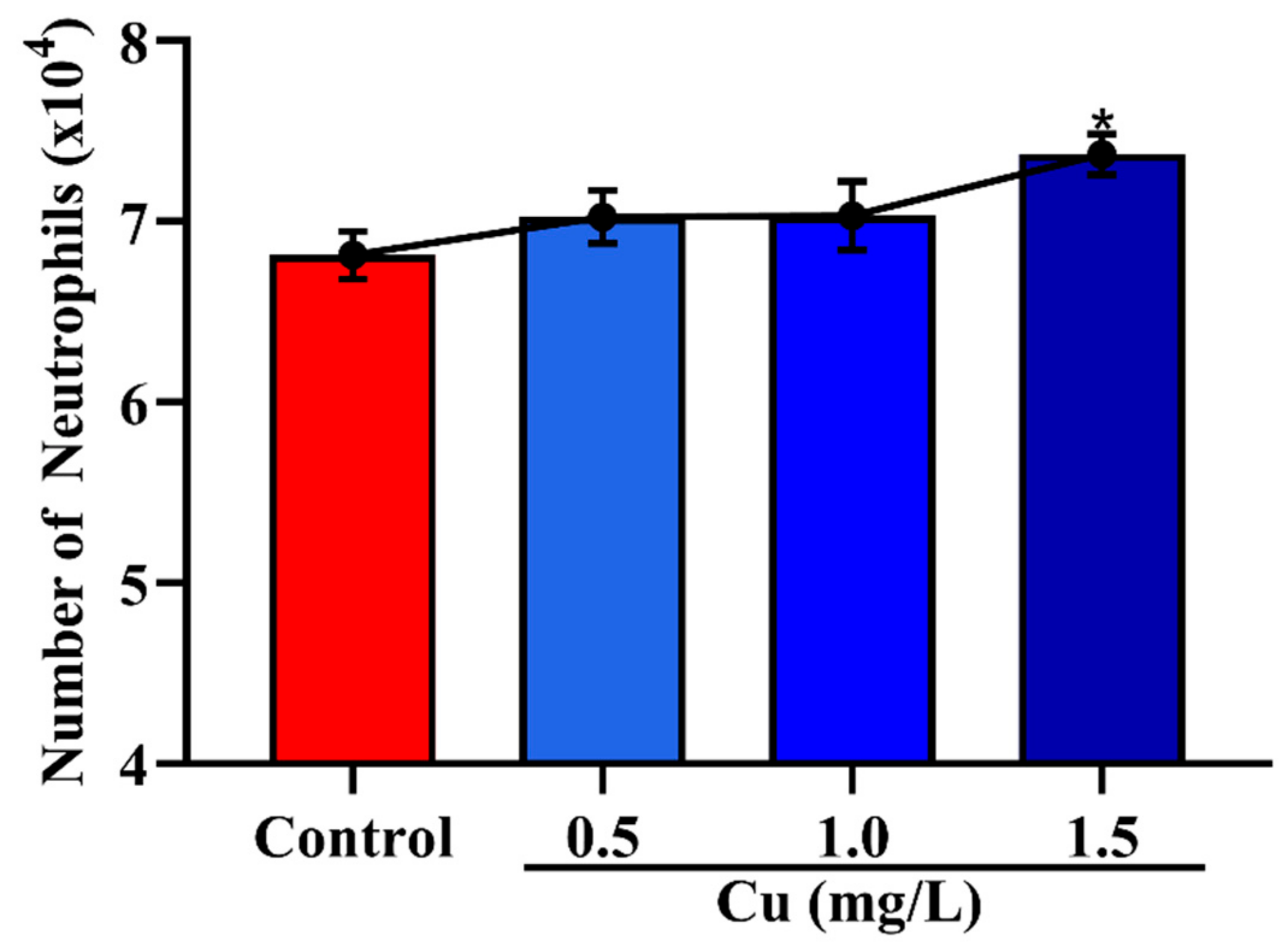
3.2. Effects of Cu on the Migration of Neutrophils across Mammary Epithelial Cells
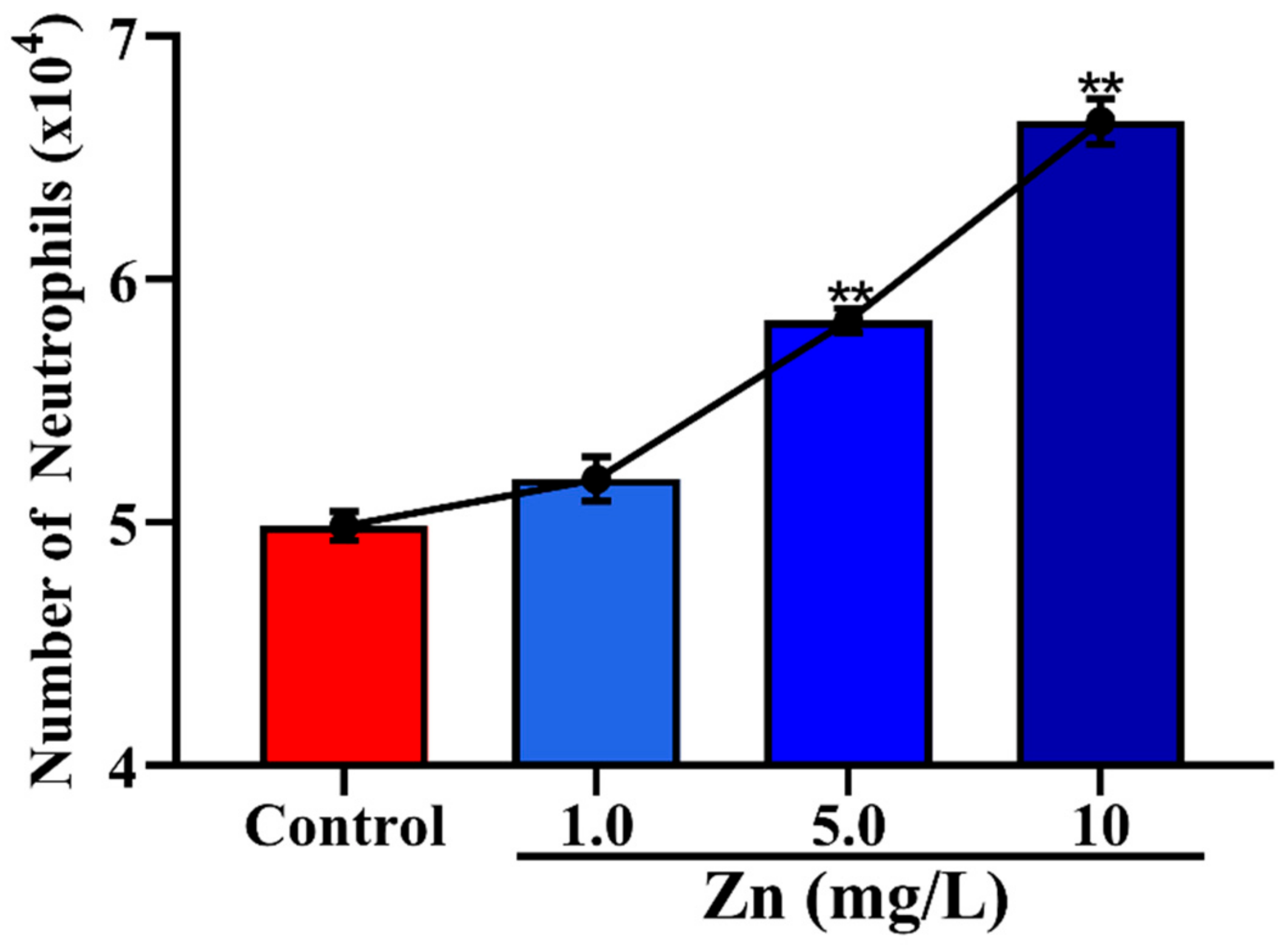
3.3. Effects of Zn on the Migration of Neutrophils across Mammary Epithelial Cells
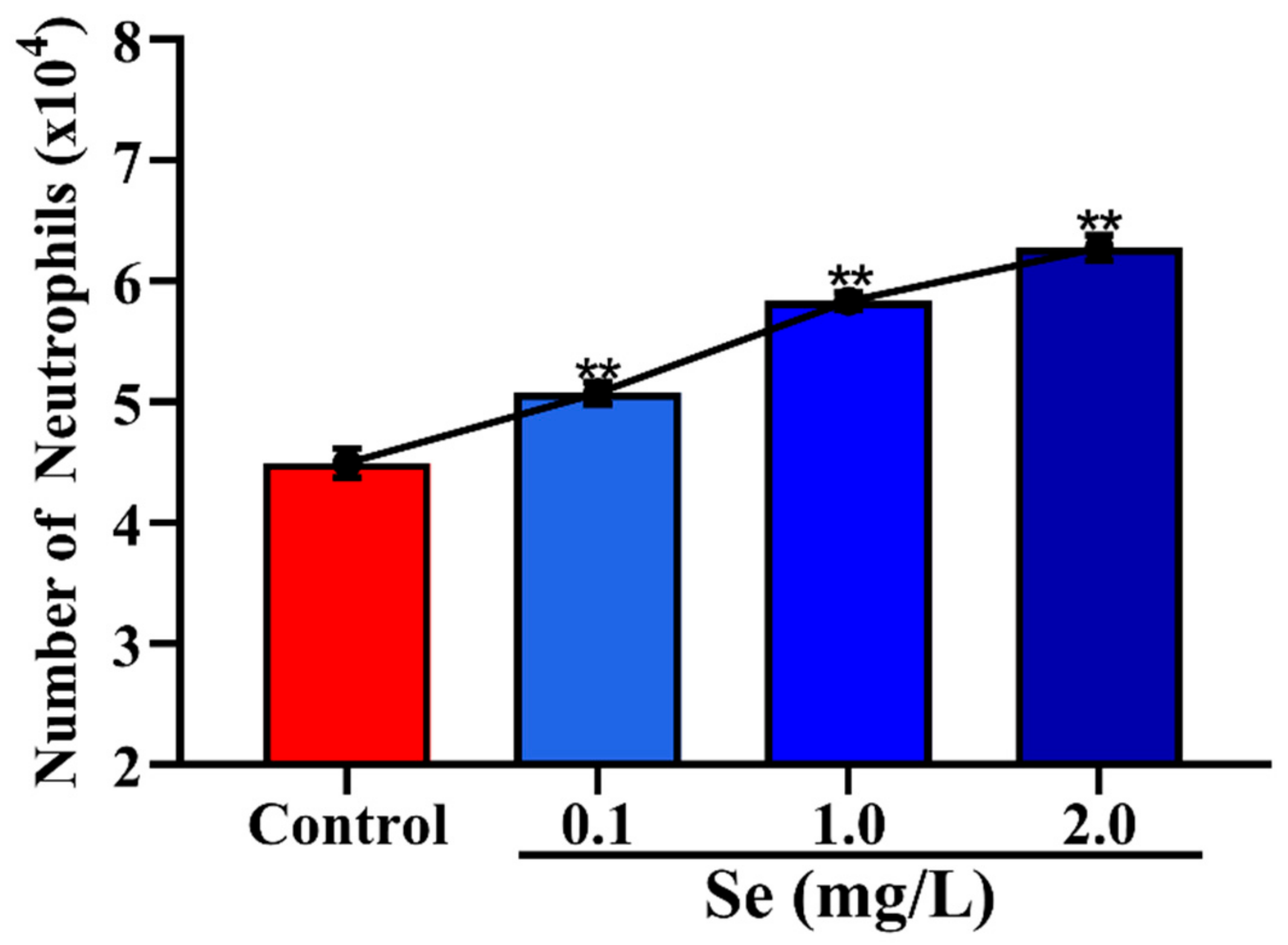
3.4. Effects of Se on the Migration of Neutrophils across Mammary Epithelial Cells
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nathan, C. Neutrophils and immunity: Challenges and opportunities. Nat. Rev. Immunol. 2006, 6, 173–182. [Google Scholar] [CrossRef]
- Segal, A.W. How neutrophils kill microbes. Annu. Rev. Immunol. 2005, 23, 197–223. [Google Scholar] [CrossRef] [Green Version]
- Amulic, B.; Cazalet, C.; Hayes, G.L.; Metzler, K.D.; Zychlinsky, A. Neutrophil function: From mechanisms to disease. Annu. Rev. Immunol. 2012, 30, 459–489. [Google Scholar] [CrossRef]
- Brinkmann, V.; Reichard, U.; Goosmann, C.; Fauler, B.; Uhlemann, Y.; Weiss, D.S.; Weinrauch, Y.; Zychlinsky, A. Neutrophil extracellular traps kill bacteria. Science 2004, 303, 1532–1535. [Google Scholar] [CrossRef] [PubMed]
- Filippi, M.D. Neutrophil transendothelial migration: Updates and new perspectives. Blood 2019, 133, 2149–2158. [Google Scholar] [CrossRef] [PubMed]
- Vestweber, D. How leukocytes cross the vascular endothelium. Nat. Rev. Immunol. 2015, 15, 692–704. [Google Scholar] [CrossRef] [PubMed]
- Kaplan, M.J.; Radic, M. Neutrophil extracellular traps: Double-edged swords of innate immunity. J. Immunol. 2012, 189, 2689–2695. [Google Scholar] [CrossRef] [Green Version]
- Jaganjac, M.; Cipak, A.; Schaur, R.J.; Zarkovic, N. Pathophysiology of neutrophil-mediated extracellular redox reactions. Front. Biosci. Landmark Ed. 2016, 21, 839–855. [Google Scholar] [CrossRef] [Green Version]
- Rainard, P.; Riollet, C. Mobilization of neutrophils and defense of the bovine mammary gland. Reprod. Nutr. Dev. 2003, 43, 439–457. [Google Scholar] [CrossRef] [Green Version]
- Herbelin, C.; Poutrel, B.; Gilbert, F.B.; Rainard, P. Immune recruitment and bactericidal activity of neutrophils in milk of cows vaccinated with staphylococcal alpha-toxin. J. Dairy Sci. 1997, 80, 2025–2034. [Google Scholar] [CrossRef]
- Colditz, I.G.; Watson, D.L. Effect of immunisation on the early influx of neutrophils during staphylococcal mastitis in ewes. Res. Vet. Sci. 1982, 33, 146–151. [Google Scholar] [CrossRef]
- Suttle, N.F.; Jones, D.G. Recent developments in trace element metabolism and function: Trace elements, disease resistance and immune responsiveness in ruminants. J. Nutr. 1989, 119, 1055–1061. [Google Scholar] [CrossRef] [PubMed]
- Wintergerst, E.S.; Maggini, S.; Hornig, D.H. Contribution of selected vitamins and trace elements to immune function. Ann. Nutr. Metab. 2007, 51, 301–323. [Google Scholar] [CrossRef] [Green Version]
- Bicalho, M.L.; Lima, F.S.; Ganda, E.K.; Foditsch, C.; Meira, E.J.; Machado, V.S.; Teixeira, A.G.; Oikonomou, G.; Gilbert, R.O.; Bicalho, R.C. Effect of trace mineral supplementation on selected minerals, energy metabolites, oxidative stress, and immune parameters and its association with uterine diseases in dairy cattle. J. Dairy Sci. 2014, 97, 4281–4295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valko, M.; Rhodes, C.J.; Moncol, J.; Izakovic, M.; Mazur, M. Free radicals, metals and antioxidants in oxidative stress-induced cancer. Chem. Biol. Interact. 2006, 160, 1–40. [Google Scholar] [CrossRef] [PubMed]
- Wessels, I.; Maywald, M.; Rink, L. Zinc as a Gatekeeper of Immune Function. Nutrients 2017, 9, 1286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Avery, J.C.; Hoffmann, P.R. Selenium, Selenoproteins, and Immunity. Nutrients 2018, 10, 1203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boyne, R.; Arthur, J.R. Effects of selenium and copper deficiency on neutrophil function in cattle. J. Comp. Pathol. 1981, 91, 271–276. [Google Scholar] [CrossRef]
- Jones, D.G.; Suttle, N.F. Some effects of copper deficiency on leucocyte function in sheep and cattle. Res. Vet. Sci. 1981, 31, 151–156. [Google Scholar] [CrossRef]
- Olkowski, A.A.; Gooneratne, S.R.; Christensen, D.A. Effects of diets of high sulphur content and varied concentrations of copper, molybdenum and thiamine on in vitro phagocytic and candidacidal activity of neutrophils in sheep. Res. Vet. Sci. 1990, 48, 82–86. [Google Scholar] [CrossRef]
- Percival, S.S. Neutropenia caused by copper deficiency: Possible mechanisms of action. Nutr. Rev. 1995, 53, 59–66. [Google Scholar] [CrossRef]
- Vruwink, K.G.; Keen, C.L.; Gershwin, M.E.; Mareschi, J.P.; Hurley, L.S. The Effect of Experimental Zinc Deficiency on Development of the Immune System. In Nutrient Modulation of the Immune Response; CRC Press: New York, NY, USA, 2020; pp. 263–280. [Google Scholar]
- Bednarek, D.; Kondracki, M.; Krasucki, J. The effect of zinc on mineral, hematological and immunological indices in calves. Pol. Arch. Weter 1991, 31, 129–140. [Google Scholar] [PubMed]
- Rizzo, A.; Pantaleo, M.; Mutinati, M.; Minoia, G.; Trisolini, C.; Ceci, E.; Sciorsci, R.L. Blood and milk oxidative status after administration of different antioxidants during early postpartum in dairy cows. Res. Vet. Sci. 2013, 95, 1171–1174. [Google Scholar] [CrossRef]
- Mehdi, Y.; Dufrasne, I. Selenium in Cattle: A Review. Molecules 2016, 21, 545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ndiweni, N.; Finch, J.M. Effects of in vitro supplementation with alpha-tocopherol and selenium on bovine neutrophil functions: Implications for resistance to mastitis. Vet. Immunol. Immunopathol. 1996, 51, 67–78. [Google Scholar] [CrossRef]
- Williams, M.R.; Azcutia, V.; Newton, G.; Alcaide, P.; Luscinskas, F.W. Emerging mechanisms of neutrophil recruitment across endothelium. Trends Immunol. 2011, 32, 461–469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gordon, S.A.; Lominadze, D.; Saari, J.T.; Lentsch, A.B.; Schuschke, D.A. Impaired deformability of copper-deficient neutrophils. Exp. Biol. Med. 2005, 230, 543–548. [Google Scholar] [CrossRef] [PubMed]
- Lominadze, D.; Saari, J.T.; Percival, S.S.; Schuschke, D.A. Proinflammatory effects of copper deficiency on neutrophils and lung endothelial cells. Immunol. Cell Biol. 2004, 82, 231–238. [Google Scholar] [CrossRef] [Green Version]
- Schuschke, D.A.; Percival, S.S.; Lominadze, D.; Saari, J.T.; Lentsch, A.B. Tissue-specific ICAM-1 expression and neutrophil transmigration in the copper-deficient rat. Inflammation 2002, 26, 297–303. [Google Scholar] [CrossRef] [PubMed]
- Haase, H.; Rink, L. Zinc signals and immune function. Biofactors 2014, 40, 27–40. [Google Scholar] [CrossRef]
- Puls, R. Mineral Levels in Animal Health: Diagnostic Data, 2nd ed.; Sherpa International: Clearbrook, BC, Canada, 1994. [Google Scholar]
- Hasan, R.; Rink, L.; Haase, H. Chelation of Free Zn2+ Impairs Chemotaxis, Phagocytosis, Oxidative Burst, Degranulation, and Cytokine Production by Neutrophil Granulocytes. Biol. Trace Elem. Res. 2016, 171, 79–88. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.; Rose, A.H.; Hoffmann, P.R. The role of selenium in inflammation and immunity: From molecular mechanisms to therapeutic opportunities. Antioxid. Redox Signal. 2012, 16, 705–743. [Google Scholar] [CrossRef] [PubMed] [Green Version]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, H.; Lv, G.; Lian, S.; Wang, J.; Wu, R. Effect of Copper, Zinc, and Selenium on the Migration of Bovine Neutrophils. Vet. Sci. 2021, 8, 281. https://doi.org/10.3390/vetsci8110281
Wang H, Lv G, Lian S, Wang J, Wu R. Effect of Copper, Zinc, and Selenium on the Migration of Bovine Neutrophils. Veterinary Sciences. 2021; 8(11):281. https://doi.org/10.3390/vetsci8110281
Chicago/Turabian StyleWang, Hai, Guanxin Lv, Shuai Lian, Jianfa Wang, and Rui Wu. 2021. "Effect of Copper, Zinc, and Selenium on the Migration of Bovine Neutrophils" Veterinary Sciences 8, no. 11: 281. https://doi.org/10.3390/vetsci8110281
APA StyleWang, H., Lv, G., Lian, S., Wang, J., & Wu, R. (2021). Effect of Copper, Zinc, and Selenium on the Migration of Bovine Neutrophils. Veterinary Sciences, 8(11), 281. https://doi.org/10.3390/vetsci8110281