
The Role of Outer Membrane Protein 16 in Brucella Pathogenesis, Vaccine Development, and Diagnostic Applications

Simple Summary
Abstract
1. Introduction
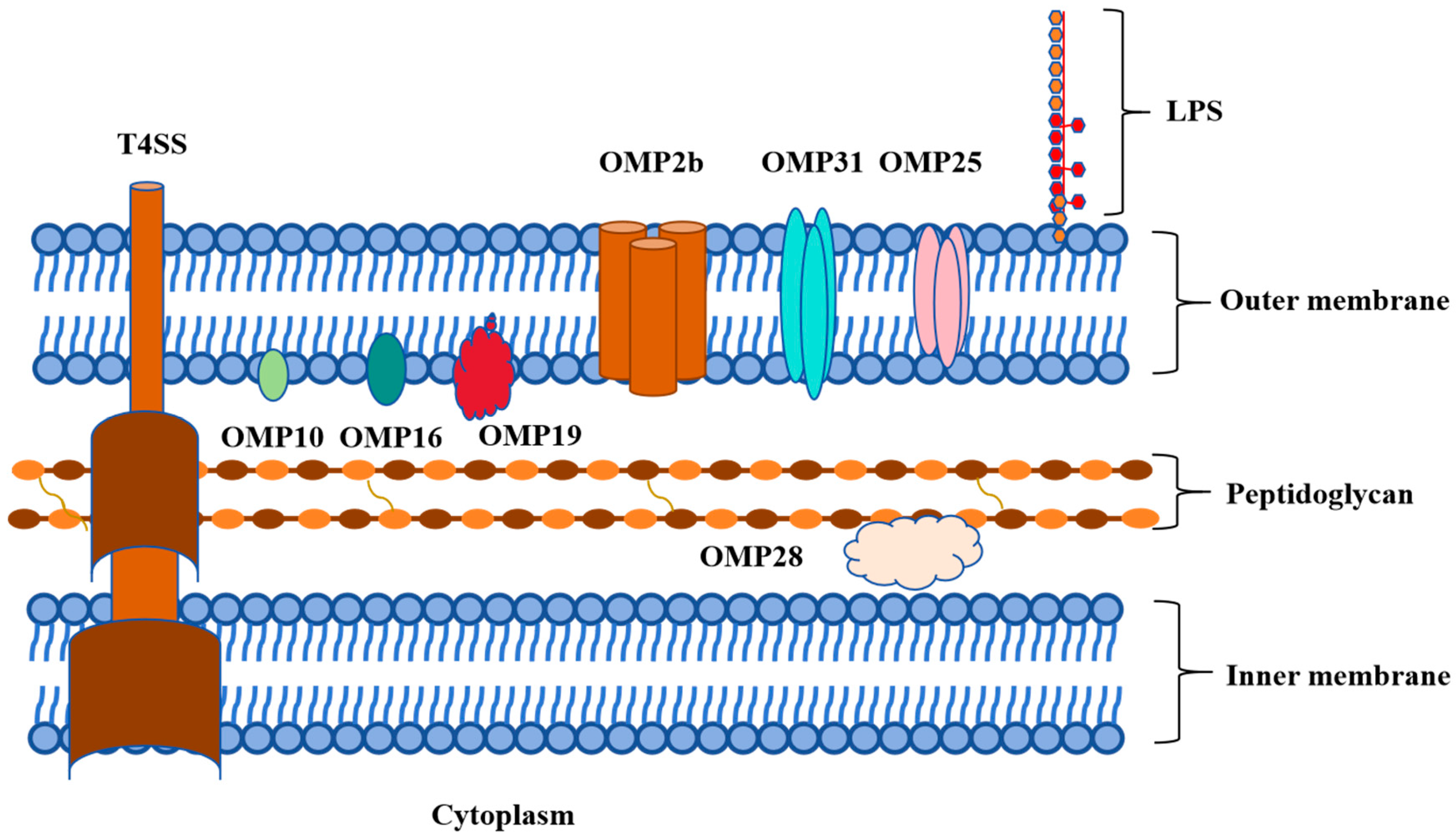
2. Structural and Functional Characteristics of Omp16
3. Omp16 in Vaccine Development
4. Diagnostic Applications of Omp16 in Brucellosis Detection
5. Future Directions and Conclusions
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gonzalez-Espinoza, G.; Arce-Gorvel, V.; Memet, S.; Gorvel, J.P. Brucella: Reservoirs and Niches in Animals and Humans. Pathogens 2021, 10, 186. [Google Scholar] [CrossRef] [PubMed]
- Jiao, H.; Zhou, Z.; Li, B.; Xiao, Y.; Li, M.; Zeng, H.; Guo, X.; Gu, G. The Mechanism of Facultative Intracellular Parasitism of Brucella. Int. J. Mol. Sci. 2021, 22, 3673. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Yang, X.; Lindahl, J.F.; Yang, G.; Gilbert, J.; Unger, F. Participatory and Transdisciplinary Studies of Brucella Infection in Humans and Animals in Yunnan Province, China-Lessons Learned. Trop. Med. Infect. Dis. 2021, 6, 134. [Google Scholar] [CrossRef] [PubMed]
- Ghanbari, M.K.; Gorji, H.A.; Behzadifar, M.; Sanee, N.; Mehedi, N.; Bragazzi, N.L. One health approach to tackle brucellosis: A systematic review. Trop. Med. Health 2020, 48, 86. [Google Scholar] [CrossRef]
- Byndloss, M.X.; Tsolis, R.M. Brucella spp. Virulence Factors and Immunity. Annu. Rev. Anim. Biosci. 2016, 4, 111–127. [Google Scholar] [CrossRef]
- Moreno, E.; Middlebrook, E.A.; Altamirano-Silva, P.; Al Dahouk, S.; Araj, G.F.; Arce-Gorvel, V.; Arenas-Gamboa, A.; Ariza, J.; Barquero-Calvo, E.; Battelli, G.; et al. If You’re Not Confused, You’re Not Paying Attention: Ochrobactrum Is Not Brucella. J. Clin. Microbiol. 2023, 61, e0043823. [Google Scholar] [CrossRef]
- Hordt, A.; Lopez, M.G.; Meier-Kolthoff, J.P.; Schleuning, M.; Weinhold, L.M.; Tindall, B.J.; Gronow, S.; Kyrpides, N.C.; Woyke, T.; Goker, M. Analysis of 1000+ Type-Strain Genomes Substantially Improves Taxonomic Classification of Alphaproteobacteria. Front. Microbiol. 2020, 11, 468. [Google Scholar] [CrossRef]
- L’Hote, L.; Light, I.; Mattiangeli, V.; Teasdale, M.D.; Halpin, A.; Gourichon, L.; Key, F.M.; Daly, K.G. An 8000 years old genome reveals the Neolithic origin of the zoonosis Brucella melitensis. Nat. Commun. 2024, 15, 6132. [Google Scholar] [CrossRef]
- Qureshi, K.A.; Parvez, A.; Fahmy, N.A.; Abdel Hady, B.H.; Kumar, S.; Ganguly, A.; Atiya, A.; Elhassan, G.O.; Alfadly, S.O.; Parkkila, S.; et al. Brucellosis: Epidemiology, pathogenesis, diagnosis and treatment-a comprehensive review. Ann. Med. 2023, 55, 2295398. [Google Scholar] [CrossRef]
- Guo, X.; Zeng, H.; Li, M.; Xiao, Y.; Gu, G.; Song, Z.; Shuai, X.; Guo, J.; Huang, Q.; Zhou, B.; et al. The mechanism of chronic intracellular infection with Brucella spp. Front. Cell. Infect. Microbiol. 2023, 13, 1129172. [Google Scholar]
- Ali, A.; Waris, A.; Khan, M.A.; Asim, M.; Khan, A.U.; Khan, S.; Zeb, J. Recent advancement, immune responses, and mechanism of action of various vaccines against intracellular bacterial infections. Life Sci. 2023, 314, 121332. [Google Scholar] [CrossRef] [PubMed]
- Godessart, P.; Lannoy, A.; Dieu, M.; Van der Verren, S.E.; Soumillion, P.; Collet, J.F.; Remaut, H.; Renard, P.; De Bolle, X. beta-Barrels covalently link peptidoglycan and the outer membrane in the alpha-proteobacterium Brucella abortus. Nat. Microbiol. 2021, 6, 27–33. [Google Scholar] [CrossRef] [PubMed]
- Roop, R.M., 2nd; Barton, I.S.; Hopersberger, D.; Martin, D.W. Uncovering the Hidden Credentials of Brucella Virulence. Microbiol. Mol. Biol. Rev. MMBR 2021, 85, e00021-19. [Google Scholar] [CrossRef] [PubMed]
- Cloeckaert, A.; Verger, J.M.; Grayon, M.; Vizcaino, N. Molecular and immunological characterization of the major outer membrane proteins of Brucella. FEMS Microbiol. Lett. 1996, 145, 1–8. [Google Scholar] [CrossRef]
- Zhi, F.J.; Zhou, D.; Li, J.M.; Tian, L.L.; Zhang, G.D.; Jin, Y.P.; Wang, A.H. Omp16, a conserved peptidoglycan-associated lipoprotein, is involved in virulence. J. Microbiol. 2020, 58, 793–804. [Google Scholar] [CrossRef]
- Sidhu-Munoz, R.S.; Sancho, P.; Vizcaino, N. Brucella ovis PA mutants for outer membrane proteins Omp10, Omp19, SP41, and BepC are not altered in their virulence and outer membrane properties. Vet. Microbiol. 2016, 186, 59–66. [Google Scholar] [CrossRef]
- Pasquevich, K.A.; Garcia Samartino, C.; Coria, L.M.; Estein, S.M.; Zwerdling, A.; Ibanez, A.E.; Barrionuevo, P.; Oliveira, F.S.; Carvalho, N.B.; Borkowski, J.; et al. The protein moiety of Brucella abortus outer membrane protein 16 is a new bacterial pathogen-associated molecular pattern that activates dendritic cells in vivo, induces a Th1 immune response, and is a promising self-adjuvanting vaccine against systemic and oral acquired brucellosis. J. Immunol. 2010, 184, 5200–5212. [Google Scholar]
- Tibor, A.; Weynants, V.; Denoel, P.; Lichtfouse, B.; De Bolle, X.; Saman, E.; Limet, J.N.; Letesson, J.J. Molecular cloning, nucleotide sequence, and occurrence of a 16.5-kilodalton outer membrane protein of Brucella abortus with similarity to pal lipoproteins. Infect. Immun. 1994, 62, 3633–3639. [Google Scholar] [CrossRef]
- Giambartolomei, G.H.; Zwerdling, A.; Cassataro, J.; Bruno, L.; Fossati, C.A.; Philipp, M.T. Lipoproteins, not lipopolysaccharide, are the key mediators of the proinflammatory response elicited by heat-killed Brucella abortus. J. Immunol. 2004, 173, 4635–4642. [Google Scholar] [CrossRef]
- Avila-Calderon, E.D.; Flores-Romo, L.; Sharon, W.; Donis-Maturano, L.; Becerril-Garcia, M.A.; Arreola, M.G.A.; Reynoso, B.A.; Guemes, F.S.; Contreras-Rodriguez, A. Dendritic cells and Brucella spp. interaction: The sentinel host and the stealthy pathogen. Folia Microbiol. 2020, 65, 1–16. [Google Scholar] [CrossRef]
- Avila-Calderon, E.D.; Lopez-Merino, A.; Jain, N.; Peralta, H.; Lopez-Villegas, E.O.; Sriranganathan, N.; Boyle, S.M.; Witonsky, S.; Contreras-Rodriguez, A. Characterization of outer membrane vesicles from Brucella melitensis and protection induced in mice. Clin. Dev. Immunol. 2012, 2012, 352493. [Google Scholar] [CrossRef] [PubMed]
- Avila-Calderon, E.D.; Medina-Chavez, O.; Flores-Romo, L.; Hernandez-Hernandez, J.M.; Donis-Maturano, L.; Lopez-Merino, A.; Arellano-Reynoso, B.; Aguilera-Arreola, M.G.; Ruiz, E.A.; Gomez-Lunar, Z.; et al. Outer Membrane Vesicles From Brucella melitensis Modulate Immune Response and Induce Cytoskeleton Rearrangement in Peripheral Blood Mononuclear Cells. Front. Microbiol. 2020, 11, 556795. [Google Scholar] [CrossRef] [PubMed]
- Pasquevich, K.A.; Estein, S.M.; Garcia Samartino, C.; Zwerdling, A.; Coria, L.M.; Barrionuevo, P.; Fossati, C.A.; Giambartolomei, G.H.; Cassataro, J. Immunization with recombinant Brucella species outer membrane protein Omp16 or Omp19 in adjuvant induces specific CD4+ and CD8+ T cells as well as systemic and oral protection against Brucella abortus infection. Infect. Immun. 2009, 77, 436–445. [Google Scholar] [CrossRef] [PubMed]
- Smaldini, P.L.; Ibanez, A.E.; Fossati, C.A.; Cassataro, J.; Docena, G.H. Oral delivery of Brucella spp. recombinant protein U-Omp16 abrogates the IgE-mediated milk allergy. Hum. Vaccines Immunother. 2014, 10, 2015–2023. [Google Scholar] [CrossRef]
- Alizadeh, H.; Dezfulian, M.; Rahnema, M.; Fallah, J.; Esmaeili, D. Protection of BALB/c mice against pathogenic Brucella abortus and Brucella melitensis by vaccination with recombinant Omp16. Iran. J. Basic Med. Sci. 2019, 22, 1302–1307. [Google Scholar]
- Araiza-Villanueva, M.; Avila-Calderón, E.D.; Flores-Romo, L.; Calderón-Amador, J.; Sriranganathan, N.; Al Qublan, H.; Witonsky, S.; Aguilera-Arreola, M.G.; Ruiz-Palma, M.D.; Ruiz, E.A.; et al. Proteomic Analysis of Membrane Blebs of 2308 and RB51 and Their Evaluation as an Acellular Vaccine. Front. Microbiol. 2019, 10, 2714. [Google Scholar] [CrossRef]
- Li, X.; Zhang, B.; Zhang, D.; Liu, S.; Ren, J. The construction of recombinant Lactobacillus casei vaccine of PEDV and its immune responses in mice. BMC Vet. Res. 2021, 17, 184. [Google Scholar] [CrossRef]
- Ibanez, A.E.; Smaldini, P.; Coria, L.M.; Delpino, M.V.; Pacifico, L.G.; Oliveira, S.C.; Risso, G.S.; Pasquevich, K.A.; Fossati, C.A.; Giambartolomei, G.H.; et al. Unlipidated outer membrane protein Omp16 (U-Omp16) from Brucella spp. as nasal adjuvant induces a Th1 immune response and modulates the Th2 allergic response to cow’s milk proteins. PLoS ONE 2013, 8, e69438. [Google Scholar] [CrossRef]
- Orsini Delgado, M.L.; Rizzo, G.P.; Fossati, C.A.; Pasquevich, K.A.; Cassataro, J.; Smaldini, P.L.; Docena, G.H. Sublingual Omp16-driven redirection of the allergic intestinal response in a pre-clinical model of food allergy. Clin. Exp. Allergy J. Br. Soc. Allergy Clin. Immunol. 2020, 50, 954–963. [Google Scholar] [CrossRef]
- Alkhalaf, S.A.S.; Mohamad, B.T.; Nicoletti, P. Control of Brucellosis in Kuwait by Vaccination of Cattle, Sheep and Goats with Brucella-Abortus Strain-19 or Brucella-Melitensis Strain Rev-1. Trop. Anim. Health Prod. 1992, 24, 45–49. [Google Scholar] [CrossRef]
- Morgan, W.J.; Littlejohn, A.I.; Mackinnon, D.J.; Lawson, J.R. The degree of protection given by living vaccines against experimental infection with Brucella melitensis in goats. Bull. World Health Organ. 1966, 34, 33–40. [Google Scholar] [PubMed]
- Bosseray, N.; Plommet, M. Brucella suis S2, Brucella melitensis Rev. 1 and Brucella abortus S19 living vaccines: Residual virulence and immunity induced against three Brucella species challenge strains in mice. Vaccine 1990, 8, 462–468. [Google Scholar] [CrossRef] [PubMed]
- Schurig, G.G.; Roop, R.M., 2nd; Bagchi, T.; Boyle, S.; Buhrman, D.; Sriranganathan, N. Biological properties of RB51; a stable rough strain of Brucella abortus. Vet. Microbiol. 1991, 28, 171–188. [Google Scholar] [CrossRef] [PubMed]
- Edwards, S.; De Ropp, R.; Mcleod, D. A study of the immunological properties and infectivity of Brucella abortus strain 45/20, McEwen, in cattle. Vet. Rec. 1945, 57, 259–264. [Google Scholar]
- Li, Z.; Zhang, H.; Tang, L.; Chen, C.; Wang, Y. Construction and identification of virB2 deletion mutants of Brucella vaccine strain M5-90. Wei Sheng Wu Xue Bao 2010, 50, 1677–1680. [Google Scholar]
- Yi, X.; Ye, F.; Yao, G.; Gu, W.; Ma, X.; Wu, D.; Zhong, Q. Construction of Brucella abortus A19-delta VirB12 mutant and evaluation of its protective efficacy against 2308 strain challenge in BALB/c mice. Wei Sheng Wu Xue Bao 2013, 53, 1213–1220. [Google Scholar]
- Tabynov, K.; Yespembetov, B.; Sansyzbay, A. Novel vector vaccine against Brucella abortus based on influenza A viruses expressing Brucella L7/L12 or Omp16 proteins: Evaluation of protection in pregnant heifers. Vaccine 2014, 32, 5889–5892. [Google Scholar] [CrossRef]
- Tabynov, K.; Kydyrbayev, Z.; Ryskeldinova, S.; Yespembetov, B.; Zinina, N.; Assanzhanova, N.; Kozhamkulov, Y.; Inkarbekov, D.; Gotskina, T.; Sansyzbay, A. Novel influenza virus vectors expressing Brucella L7/L12 or Omp16 proteins in cattle induced a strong T-cell immune response, as well as high protectiveness against B. abortus infection. Vaccine 2014, 32, 2034–2041. [Google Scholar] [CrossRef]
- Ryskeldinova, S.; Zinina, N.; Kydyrbayev, Z.; Yespembetov, B.; Kozhamkulov, Y.; Inkarbekov, D.; Assanzhanova, N.; Mailybayeva, A.; Bugybayeva, D.; Sarmykova, M.; et al. Registered Influenza Viral Vector Based Brucella abortus Vaccine for Cattle in Kazakhstan: Age-Wise Safety and Efficacy Studies. Front. Cell. Infect. Microbiol. 2021, 11, 669196. [Google Scholar] [CrossRef]
- Mailybayeva, A.; Ryskeldinova, S.; Zinina, N.; Zhou, E.M.; Renukaradhya, G.J.; Tabynov, K. Evaluation of Duration of Immunogenicity and Protective Efficacy of Improved Influenza Viral Vector-Based Brucella abortus Vaccine Against Brucella melitensis Infection in Sheep and Goats. Front. Vet. Sci. 2020, 7, 58. [Google Scholar] [CrossRef]
- Tabynov, K.; Ryskeldinova, S.; Sansyzbay, A. An influenza viral vector Brucella abortus vaccine induces good cross-protection against Brucella melitensis infection in pregnant heifers. Vaccine 2015, 33, 3619–3623. [Google Scholar] [CrossRef]
- Mailybayeva, A.; Yespembetov, B.; Ryskeldinova, S.; Zinina, N.; Sansyzbay, A.; Renukaradhya, G.J.; Petrovsky, N.; Tabynov, K. Improved influenza viral vector based Brucella abortus vaccine induces robust B and T-cell responses and protection against Brucella melitensis infection in pregnant sheep and goats. PLoS ONE 2017, 12, e0186484. [Google Scholar] [CrossRef] [PubMed]
- Tabynov, K.; Yespembetov, B.; Matikhan, N.; Ryskeldinova, S.; Zinina, N.; Kydyrbayev, Z.; Assanzhanova, N.; Tabynov, K.; Renukaradhya, G.J.; Mukhitdinova, G.; et al. First evaluation of an influenza viral vector based Brucella abortus vaccine in sheep and goats: Assessment of safety, immunogenicity and protective efficacy against Brucella melitensis infection. Vet. Microbiol. 2016, 197, 15–20. [Google Scholar] [CrossRef] [PubMed]
- Luo, D.; Ni, B.; Li, P.; Shi, W.; Zhang, S.; Han, Y.; Mao, L.; He, Y.; Wu, Y.; Wang, X. Protective immunity elicited by a divalent DNA vaccine encoding both the L7/L12 and Omp16 genes of Brucella abortus in BALB/c mice. Infect. Immun. 2006, 74, 2734–2741. [Google Scholar] [CrossRef] [PubMed]
- Huy, T.X.N.; Nguyen, T.T.; Reyes, A.W.B.; Vu, S.H.; Min, W.; Lee, H.J.; Lee, J.H.; Kim, S. Immunization with a Combination of Four Recombinant Brucella abortus Proteins Omp16, Omp19, Omp28, and L7/L12 Induces T Helper 1 Immune Response Against Virulent B. abortus 544 Infection in BALB/c Mice. Front. Vet. Sci. 2020, 7, 577026. [Google Scholar] [CrossRef]
- Tabynov, K.; Sansyzbay, A.; Kydyrbayev, Z.; Yespembetov, B.; Ryskeldinova, S.; Zinina, N.; Assanzhanova, N.; Sultankulova, K.; Sandybayev, N.; Khairullin, B.; et al. Influenza viral vectors expressing the Brucella OMP16 or L7/L12 proteins as vaccines against B. abortus infection. Virol. J. 2014, 11, 69. [Google Scholar] [CrossRef]
- Bugybayeva, D.; Ryskeldinova, S.; Zinina, N.; Sarmykova, M.; Assanzhanova, N.; Kydyrbayev, Z.; Tabynov, K. Development of Human Vectored Brucellosis Vaccine Formulation: Assessment of Safety and Protectiveness of Influenza Viral Vectors Expressing Brucella Immunodominant Proteins in Mice and Guinea Pigs. BioMed Res. Int. 2020, 2020, 1438928. [Google Scholar] [CrossRef]
- Tabynov, K.; Yespembetov, B.; Ryskeldinova, S.; Zinina, N.; Kydyrbayev, Z.; Kozhamkulov, Y.; Inkarbekov, D.; Sansyzbay, A. Prime-booster vaccination of cattle with an influenza viral vector Brucella abortus vaccine induces a long-term protective immune response against Brucella abortus infection. Vaccine 2016, 34, 438–444. [Google Scholar] [CrossRef]
- Sadikaliyeva, S.O.; Sultankulova, K.T.; Shorayeva, K.A.; Strochkov, V.M.; Chervyakova, O.V.; Zaitsev, V.L.; Tabynov, K.K.; Sandybayev, N.T.; Sansyzbay, A.R.; Egorov, A.Y. Genetic stability of the HA, NA, and NS genes of the recombinant vector virus Flu-NS1-124-Omp16 (H5N1) expressing the Brucellar gene. Vopr. Virusol. 2015, 60, 18–23. [Google Scholar]
- Rezaei, M.; Rabbani-Khorasgani, M.; Zarkesh-Esfahani, S.H.; Emamzadeh, R.; Abtahi, H. Lactococcus-based vaccine against brucellosis: IgG immune response in mice with rOmp16-IL2 fusion protein. Arch. Microbiol. 2021, 203, 2591–2596. [Google Scholar] [CrossRef]
- Rezaei, M.; Rabbani Khorasgani, M.; Zarkesh Esfahani, S.H.; Emamzadeh, R.; Abtahi, H. Production of Brucella melitensis Omp16 protein fused to the human interleukin 2 in Lactococcus lactis MG1363 toward developing a Lactococcus-based vaccine against brucellosis. Can. J. Microbiol. 2020, 66, 39–45. [Google Scholar] [CrossRef] [PubMed]
- Zhi, F.; Fang, J.; Zheng, W.; Li, J.; Zhang, G.; Zhou, D.; Jin, Y.; Wang, A. A Brucella Omp16 Conditional Deletion Strain Is Attenuated in BALB/c Mice. J. Microbiol. Biotechnol. 2022, 32, 6–14. [Google Scholar] [CrossRef] [PubMed]
- Golchin, M.; Mollayi, S.; Mohammadi, E.; Eskandarzade, N. Development of a diagnostic indirect ELISA test for detection of Brucella antibody using recombinant outer membrane protein 16 kDa (rOMP16). Vet. Res. Forum 2022, 13, 387–391. [Google Scholar] [PubMed]
- Bai, Q.; Li, H.; Wu, X.; Shao, J.; Sun, M.; Yin, D. Comparative analysis of the main outer membrane proteins of Brucella in the diagnosis of brucellosis. Biochem. Biophys. Res. Commun. 2021, 560, 126–131. [Google Scholar] [CrossRef]
- Yao, M.; Guo, X.; Wu, X.; Bai, Q.; Sun, M.; Yin, D. Evaluation of the Combined Use of Major Outer Membrane Proteins in the Serodiagnosis of Brucellosis. Infect. Drug Resist. 2022, 15, 4093–4100. [Google Scholar] [CrossRef]
- Tian, M.X.; Song, M.Y.; Yin, Y.; Lian, Z.M.; Li, Z.C.; Hu, H.; Guan, X.; Cai, Y.M.; Ding, C.; Wang, S.H.; et al. Characterization of the main immunogenic proteins in infection for their application in diagnosis of brucellosis. Comp. Immunol. Microbiol. 2020, 70, 101462. [Google Scholar] [CrossRef]
- Yin, D.; Bai, Q.; Zhang, J.; Xu, K.; Li, J. A novel recombinant multiepitope protein candidate for the diagnosis of brucellosis: A pilot study. J. Microbiol. Methods 2020, 174, 105964. [Google Scholar] [CrossRef]
- Verger, J.M.; Grayon, M.; Tibor, A.; Wansard, V.; Letesson, J.J.; Cloeckaert, A. Differentiation of Brucella melitensis, B. ovis and B. suis biovar 2 strains by use of membrane protein- or cytoplasmic protein-specific gene probes. Res. Microbiol. 1998, 149, 509–517. [Google Scholar] [CrossRef]
- Eoh, H.; Jeon, B.Y.; Kim, Z.; Kim, S.C.; Cho, S.N. Expression and validation of D-erythrulose 1-phosphate dehydrogenase from Brucella abortus: A diagnostic reagent for bovine brucellosis. J. Vet. Diagn. Investig. 2010, 22, 524–530. [Google Scholar] [CrossRef]

{kind=link}
| Vaccine Type | Antigen Composition | Animal Model | Immune Response | Protective Efficacy | Notes | Reference |
|---|---|---|---|---|---|---|
| Influenza viral vector | Omp16 or L7/L12 | Pregnant heifers | Humoral + T-cell | Comparable to S19/RB51 | Safe in pregnancy | [37] |
| Influenza vector + adjuvant | Omp16 + L7/L12 + Montanide/Chitosan | Cattle | Strong T-cell (CD4+/CD8+), IFN-γ | Superior to S19+adjuvant | Bivalent formulation | [38] |
| Influenza viral vector | Omp16 + L7/L12 | Cattle (various ages) | Strong IgG response | 60–75% in adults | Safe in pregnant cows | [39] |
| Influenza vector (Flu-BA_Omp19-SOD) | Omp16 + Omp19 + SOD | Sheep & goats | Th1-biased response | Partial (0–40%) | Lower in sheep | [40] |
| Influenza viral vector | Omp16 + L7/L12 | Cattle | T-cell + long-term IFN-γ | Up to 71% protection | 12 months post-boost | [41] |
| Flu-BA improved | Omp16 + L7/L12 + Omp19 + SOD | Pregnant sheep/goats | IgG2a, IFN-γ | 55–66% in dams, 73–90% in offspring | Safe in pregnancy | [42] |
| Flu-BA | Omp16 + L7/L12 | Sheep/goats | Strong T-cell (IFN-γ) | 50–57% protection | No antibody detected | [43] |
| Recombinant protein (subunit) | Omp16 (U-/L-form) | Mice | CD4+, CD8+ T cells, IFN-γ | Comparable to S19 | Oral/systemic routes tested | [23] |
| Self-adjuvanting | Unlipidated Omp16 | Mice | Th1, dendritic activation | Comparable to S19 | Oral immunization possible | [17] |
| DNA vaccine | Omp16 + L7/L12 | Mice | IgG2a, T-cell, IFN-γ | Higher than monovalent DNA | Divalent DNA construct | [44] |
| Recombinant protein | rOmp16 | Mice | IgG1/2a, IFN-γ, IL-4 | Significant protection | Against B. abortus & B. melitensis | [25] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, L.; Bai, J.; Li, L.; Jia, Y.; Qiu, X.; Luo, Y.; Zhou, D.; Zhang, Z. The Role of Outer Membrane Protein 16 in Brucella Pathogenesis, Vaccine Development, and Diagnostic Applications. Vet. Sci. 2025, 12, 605. https://doi.org/10.3390/vetsci12070605
Zhang L, Bai J, Li L, Jia Y, Qiu X, Luo Y, Zhou D, Zhang Z. The Role of Outer Membrane Protein 16 in Brucella Pathogenesis, Vaccine Development, and Diagnostic Applications. Veterinary Sciences. 2025; 12(7):605. https://doi.org/10.3390/vetsci12070605
Chicago/Turabian StyleZhang, Lu, Jun Bai, Long Li, Yanqing Jia, Xinxin Qiu, Yan Luo, Dong Zhou, and Zhencang Zhang. 2025. "The Role of Outer Membrane Protein 16 in Brucella Pathogenesis, Vaccine Development, and Diagnostic Applications" Veterinary Sciences 12, no. 7: 605. https://doi.org/10.3390/vetsci12070605
APA StyleZhang, L., Bai, J., Li, L., Jia, Y., Qiu, X., Luo, Y., Zhou, D., & Zhang, Z. (2025). The Role of Outer Membrane Protein 16 in Brucella Pathogenesis, Vaccine Development, and Diagnostic Applications. Veterinary Sciences, 12(7), 605. https://doi.org/10.3390/vetsci12070605