
Difference Analysis on Virulence Genes, Biofilms and Antimicrobial Susceptibility of Escherichia coli from Clinical and Subclinical Bovine Mastitis

 ,
,  ,
,
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
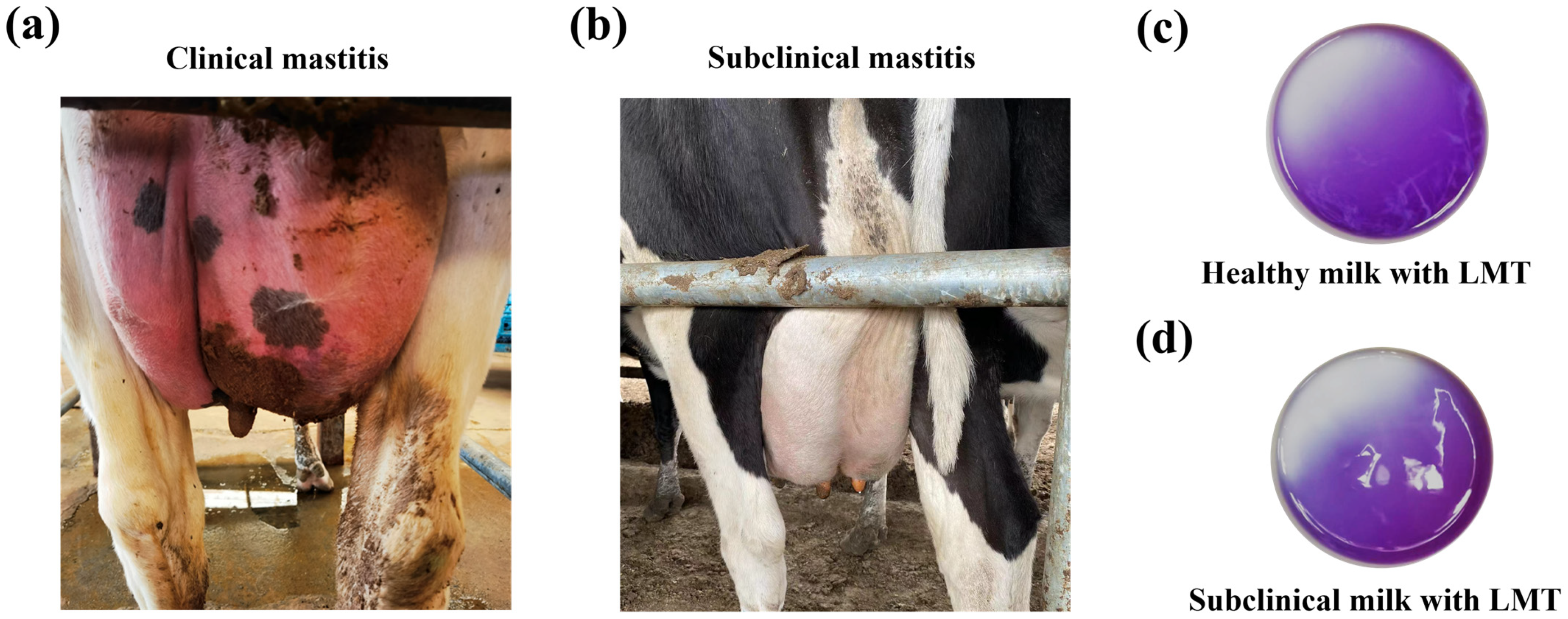
2.1. Bovine Mastitis-Related E. coli
2.2. Extraction of Bacterial Genomic DNA
2.3. Phylogenetic Analysis, Lipopolysaccharide (LPS) Core Types and Multilocus Sequence Typing (MLST) of Mastitis-Related E. coli
2.4. Distribution of Virulence Genes in Mastitis-Related E. coli
2.5. Biofilm Formation
2.6. Antimicrobial Susceptibility
2.7. Distribution of Common Antibiotic-Resistant Genes
2.8. Phylogenetic Analysis Tree
2.9. Statistical Analysis
3. Results
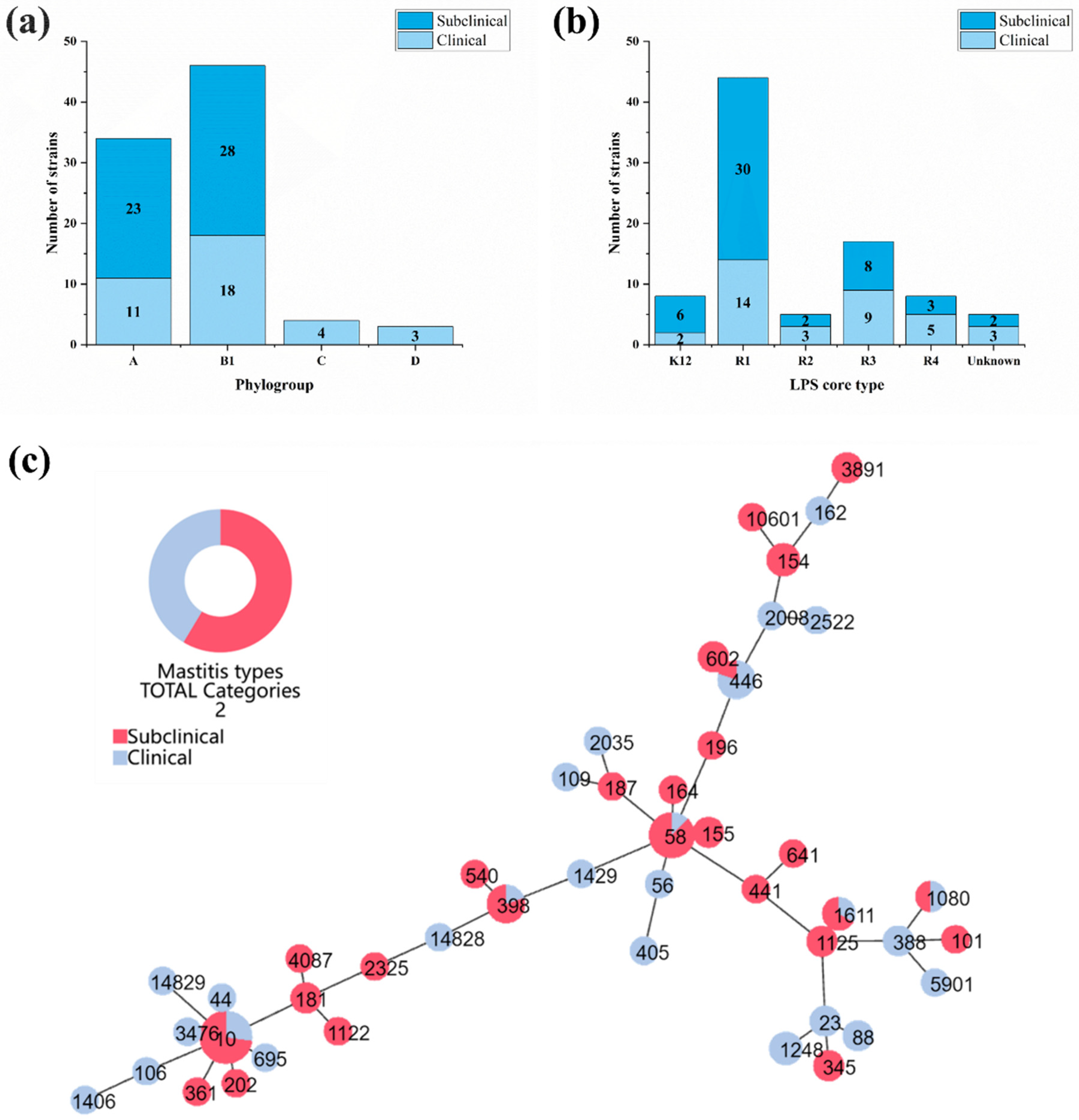
3.1. Molecular Subtyping of Clinical and Subclinical Mastitis-Related E. coli Strains
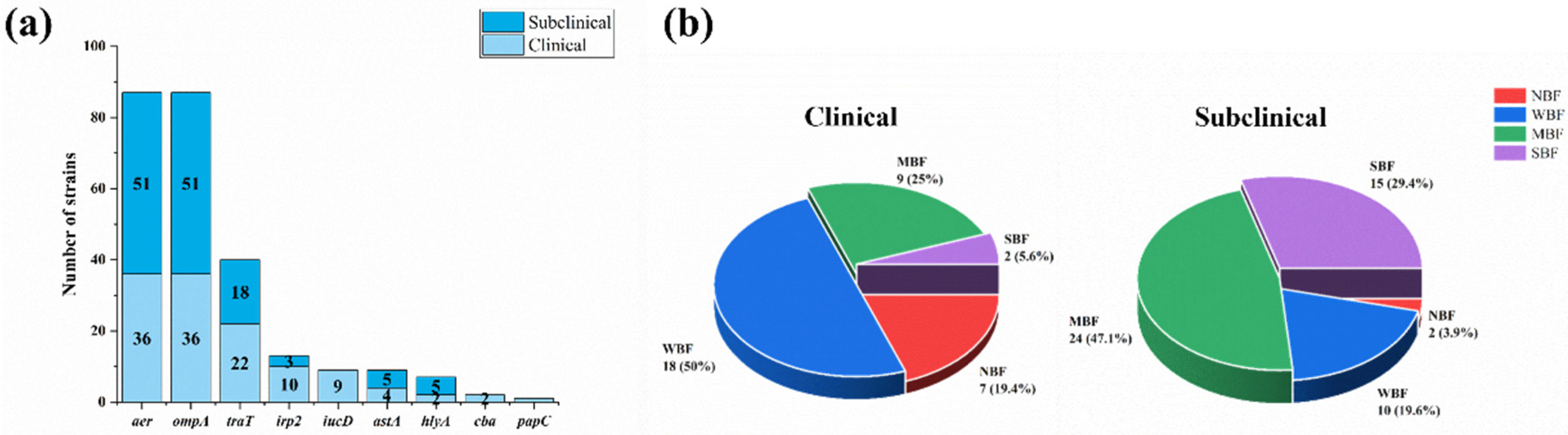
3.2. Profiles of Virulence Genes
3.3. Analysis of Biofilm Formation Abilities of Mastitis-Related E. coli
3.4. Antimicrobial Susceptibility of Mastitis-Related E. coli
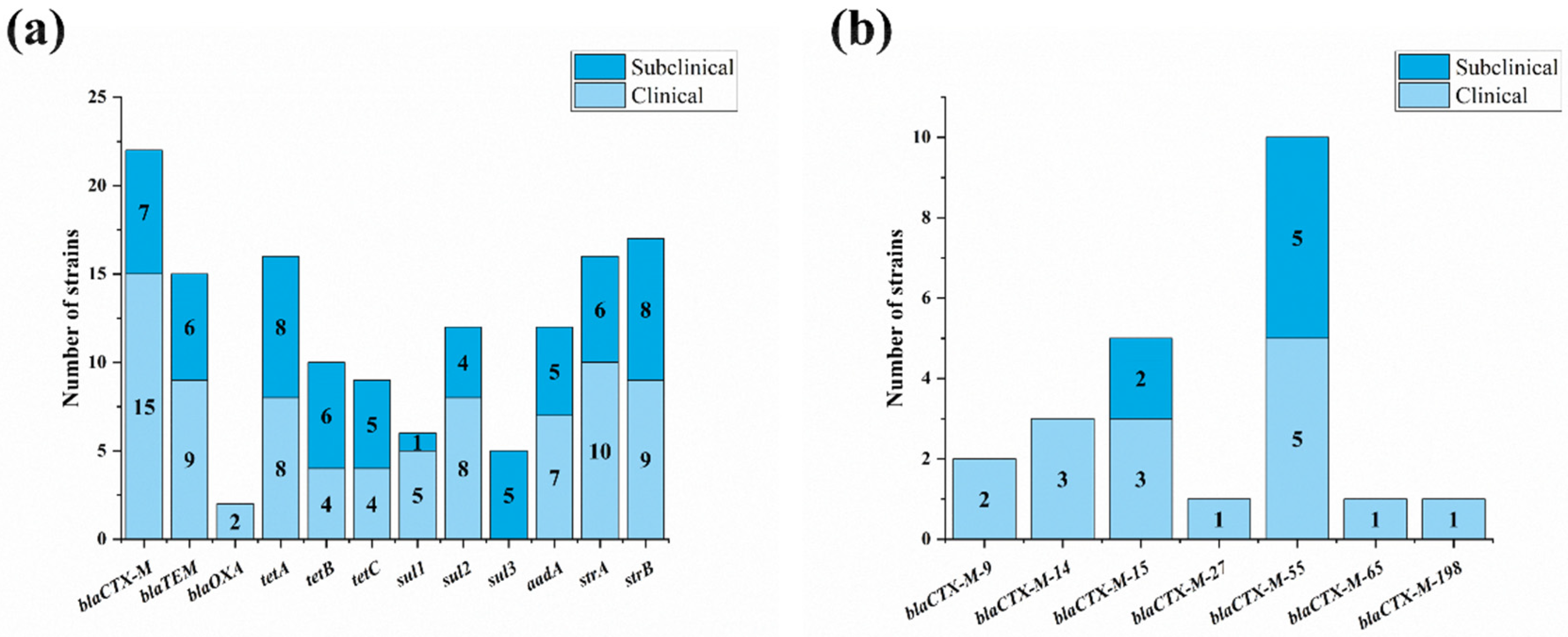
3.5. Resistance Genes
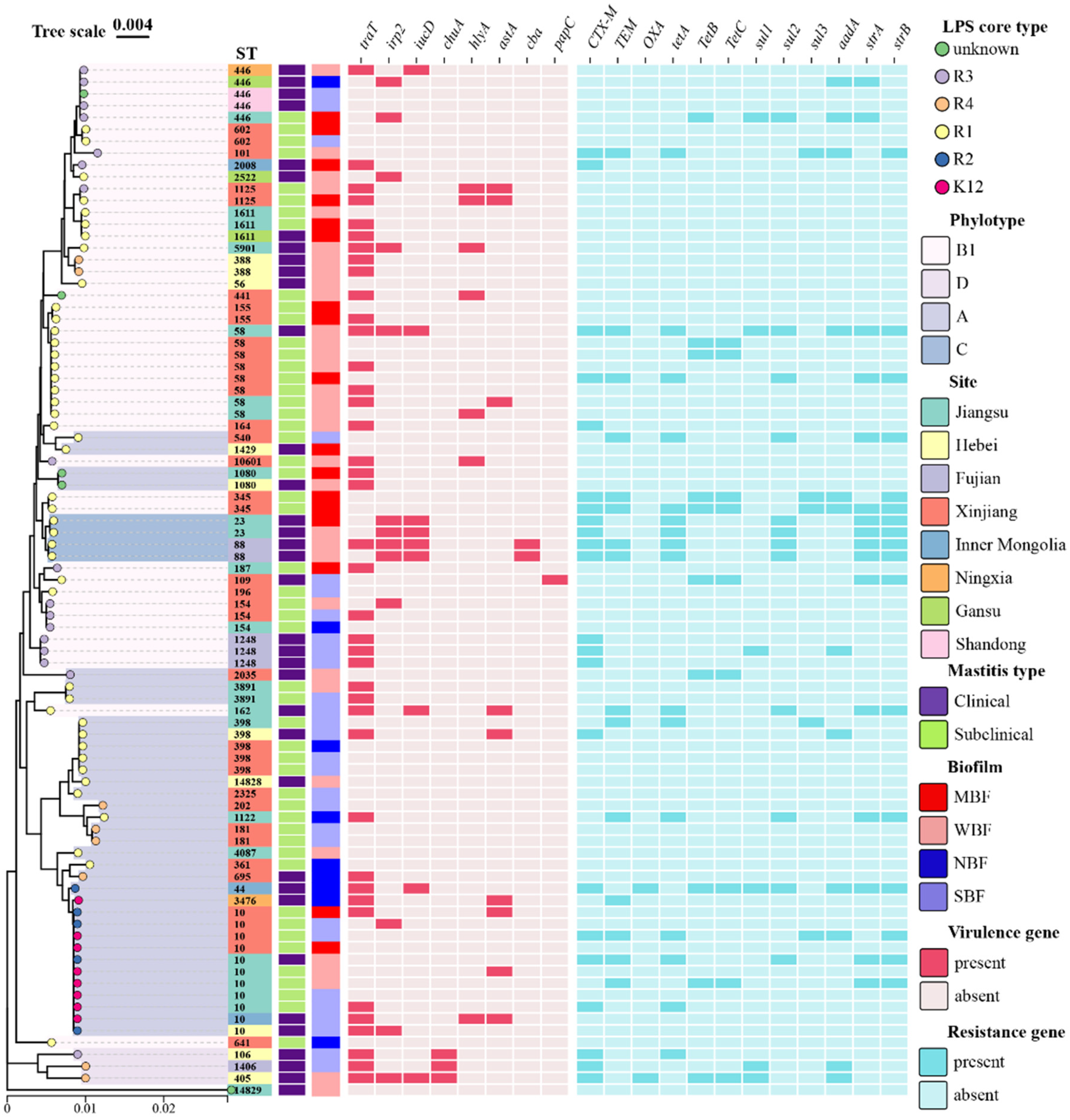
3.6. Phylogenetic Tree of Mastitis Isolates
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| CMT | California Mastitis Test |
| MLST | Multilocus sequence typing |
| LPS | Lipopolysaccharide |
| ExPEC | Extraintestinal pathogenic E. coli |
| AMR | Antimicrobial resistance |
| MDR | Multiple-drug resistance |
| ESBL-Ec | Extended-spectrum beta-lactamase-producing E. coli |
References
- Zaatout, N. An overview on mastitis-associated Escherichia coli: Pathogenicity, host immunity and the use of alternative therapies. Microbiol. Res. 2022, 256, 126960. [Google Scholar] [CrossRef] [PubMed]
- Stevens, M.; Piepers, S.; De Vliegher, S. Mastitis prevention and control practices and mastitis treatment strategies associated with the consumption of (critically important) antimicrobials on dairy herds in Flanders, Belgium. J. Dairy Sci. 2016, 99, 2896–2903. [Google Scholar] [CrossRef] [PubMed]
- Bradley, A. Bovine mastitis: An evolving disease. Vet. J. 2002, 164, 116–128. [Google Scholar] [CrossRef]
- Sharun, K.; Dhama, K.; Tiwari, R.; Gugjoo, M.B.; Iqbal Yatoo, M.; Patel, S.K.; Pathak, M.; Karthik, K.; Khurana, S.K.; Singh, R.; et al. Advances in therapeutic and managemental approaches of bovine mastitis: A comprehensive review. Vet. Q. 2021, 41, 107–136. [Google Scholar] [CrossRef] [PubMed]
- Goetz, C.; Tremblay, Y.D.N.; Lamarche, D.; Blondeau, A.; Gaudreau, A.M.; Labrie, J.; Malouin, F.; Jacques, M. Coagulase-negative staphylococci species affect biofilm formation of other coagulase-negative and coagulase-positive staphylococci. J. Dairy Sci. 2017, 100, 6454–6464. [Google Scholar] [CrossRef]
- Ashraf, A.; Imran, M. Causes, types, etiological agents, prevalence, diagnosis, treatment, prevention, effects on human health and future aspects of bovine mastitis. Anim. Health Res. Rev. 2020, 21, 36–49. [Google Scholar] [CrossRef]
- Botrel, M.A.; Haenni, M.; Morignat, E.; Sulpice, P.; Madec, J.Y.; Calavas, D. Distribution and antimicrobial resistance of clinical and subclinical mastitis pathogens in dairy cows in Rhone-Alpes, France. Foodborne Pathog. Dis. 2010, 7, 479–487. [Google Scholar] [CrossRef] [PubMed]
- Ruegg, P.L. A 100-Year Review: Mastitis detection, management, and prevention. J. Dairy Sci. 2017, 100, 10381–10397. [Google Scholar] [CrossRef] [PubMed]
- Nery Garcia, B.L.; Dantas, S.T.A.; da Silva Barbosa, K.; Mendes Mitsunaga, T.; Butters, A.; Camargo, C.H.; Nobrega, D.B. Extended-Spectrum Beta-Lactamase-Producing Escherichia coli and Other Antimicrobial-Resistant Gram-Negative Pathogens Isolated from Bovine Mastitis: A One Health Perspective. Antibiotics 2024, 13, 391. [Google Scholar] [CrossRef]
- Ren, Q.; Liao, G.; Wu, Z.; Lv, J.; Chen, W. Prevalence and characterization of Staphylococcus aureus isolates from subclinical bovine mastitis in southern Xinjiang, China. J. Dairy Sci. 2020, 103, 3368–3380. [Google Scholar] [CrossRef]
- Cheng, H.R.; Jiang, N. Extremely rapid extraction of DNA from bacteria and yeasts. Biotechnol. Lett. 2006, 28, 55–59. [Google Scholar] [CrossRef] [PubMed]
- Clermont, O.; Christenson, J.K.; Denamur, E.; Gordon, D.M. The Clermont Escherichia coli phylo-typing method revisited: Improvement of specificity and detection of new phylo-groups. Environ. Microbiol. Rep. 2013, 5, 58–65. [Google Scholar] [CrossRef] [PubMed]
- Dissanayake, D.R.; Wijewardana, T.G.; Gunawardena, G.A.; Poxton, I.R. Distribution of lipopolysaccharide core types among avian pathogenic Escherichia coli in relation to the major phylogenetic groups. Vet. Microbiol. 2008, 132, 355–363. [Google Scholar] [CrossRef] [PubMed]
- Beutin, L.; Kaulfuss, S.; Herold, S.; Oswald, E.; Schmidt, H. Genetic analysis of enteropathogenic and enterohemorrhagic Escherichia coli serogroup O103 strains by molecular typing of virulence and housekeeping genes and pulsed-field gel electrophoresis. J. Clin. Microbiol. 2005, 43, 1552–1563. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Lin, X.; Wan, Z.; Wang, S.; Zuo, J.; Wang, Z.; Xu, Y.; Han, X.; Miao, J. The Deletion of yeaJ Gene Facilitates Escherichia coli Escape from Immune Recognition. J. Bacteriol. 2021, 203, e0033621. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Liang, Y.; Lin, S.; Chen, D.; Li, B.; Li, L.; Deng, Y. Crystal Violet and XTT Assays on Staphylococcus aureus Biofilm Quantification. Curr. Microbiol. 2016, 73, 474–482. [Google Scholar] [CrossRef]
- Humphries, R.; Bobenchik, A.M.; Hindler, J.A.; Schuetz, A.N. Overview of Changes to the Clinical and Laboratory Standards Institute Performance Standards for Antimicrobial Susceptibility Testing, M100, 31st Edition. J. Clin. Microbiol. 2021, 59, e0021321. [Google Scholar] [CrossRef] [PubMed]
- Xie, J.; Chen, Y.; Cai, G.; Cai, R.; Hu, Z.; Wang, H. Tree Visualization By One Table (tvBOT): A web application for visualizing, modifying and annotating phylogenetic trees. Nucleic Acids Res. 2023, 51, W587–W592. [Google Scholar] [CrossRef] [PubMed]
- Fairbrother, J.H.; Dufour, S.; Fairbrother, J.M.; Francoz, D.; Nadeau, E.; Messier, S. Characterization of persistent and transient Escherichia coli isolates recovered from clinical mastitis episodes in dairy cows. Vet. Microbiol. 2015, 176, 126–133. [Google Scholar] [CrossRef]
- Guerra, S.T.; Orsi, H.; Joaquim, S.F.; Guimaraes, F.F.; Lopes, B.C.; Dalanezi, F.M.; Leite, D.S.; Langoni, H.; Pantoja, J.C.F.; Rall, V.L.M.; et al. Short communication: Investigation of extra-intestinal pathogenic Escherichia coli virulence genes, bacterial motility, and multidrug resistance pattern of strains isolated from dairy cows with different severity scores of clinical mastitis. J. Dairy Sci. 2020, 103, 3606–3614. [Google Scholar] [CrossRef] [PubMed]
- Seddon, C.; David, S.; Wong, J.L.C.; Ishimoto, N.; He, S.; Bradshaw, J.; Low, W.W.; Frankel, G.; Beis, K. Cryo-EM structure and evolutionary history of the conjugation surface exclusion protein TraT. Nat. Commun. 2025, 16, 659. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Wang, H.; Zhao, W.; Shan, C.; Liu, C.; Gao, L.; Zhao, R.; Ao, P.; Xiao, P.; Lv, L.; et al. New insights into the construction of wild-type Saba pig-derived Escherichia coli irp2 gene deletion strains. 3 Biotech 2021, 11, 408. [Google Scholar] [CrossRef] [PubMed]
- Cummins, M.L.; Reid, C.J.; Roy Chowdhury, P.; Bushell, R.N.; Esbert, N.; Tivendale, K.A.; Noormohammadi, A.H.; Islam, S.; Marenda, M.S.; Browning, G.F.; et al. Whole genome sequence analysis of Australian avian pathogenic Escherichia coli that carry the class 1 integrase gene. Microb. Genom. 2019, 5, e000250. [Google Scholar] [CrossRef] [PubMed]
- Goulart, D.B.; Mellata, M. Escherichia coli Mastitis in Dairy Cattle: Etiology, Diagnosis, and Treatment Challenges. Front. Microbiol. 2022, 13, 928346. [Google Scholar] [CrossRef] [PubMed]
- Jamal, M.; Ahmad, W.; Andleeb, S.; Jalil, F.; Imran, M.; Nawaz, M.A.; Hussain, T.; Ali, M.; Rafiq, M.; Kamil, M.A. Bacterial biofilm and associated infections. J. Chin. Med. Assoc. 2018, 81, 7–11. [Google Scholar] [CrossRef] [PubMed]
- Rudenko, P.; Sachivkina, N.; Vatnikov, Y.; Shabunin, S.; Engashev, S.; Kontsevaya, S.; Karamyan, A.; Bokov, D.; Kuznetsova, O.; Vasilieva, E. Role of microorganisms isolated from cows with mastitis in Moscow region in biofilm formation. Vet. World 2021, 14, 40–48. [Google Scholar] [CrossRef] [PubMed]
- Dhaouadi, S.; Romdhani, A.; Bouglita, W.; Chedli, S.; Chaari, S.; Soufi, L.; Cherif, A.; Mnif, W.; Abbassi, M.S.; Elandoulsi, R.B. High Biofilm-Forming Ability and Clonal Dissemination among Colistin-Resistant Escherichia coli Isolates Recovered from Cows with Mastitis, Diarrheic Calves, and Chickens with Colibacillosis in Tunisia. Life 2023, 13, 299. [Google Scholar] [CrossRef]
- Roth, N.; Kasbohrer, A.; Mayrhofer, S.; Zitz, U.; Hofacre, C.; Domig, K.J. The application of antibiotics in broiler production and the resulting antibiotic resistance in Escherichia coli: A global overview. Poult. Sci. 2019, 98, 1791–1804. [Google Scholar] [CrossRef] [PubMed]
- Aworh, M.K.; Kwaga, J.K.P.; Hendriksen, R.S.; Okolocha, E.C.; Thakur, S. Genetic relatedness of multidrug resistant Escherichia coli isolated from humans, chickens and poultry environments. Antimicrob. Resist. Infect. Control 2021, 10, 58. [Google Scholar] [CrossRef] [PubMed]
- Suojala, L.; Kaartinen, L.; Pyorala, S. Treatment for bovine Escherichia coli mastitis—An evidence-based approach. J. Vet. Pharmacol. Ther. 2013, 36, 521–531. [Google Scholar] [CrossRef]
- Poirel, L.; Madec, J.Y.; Lupo, A.; Schink, A.K.; Kieffer, N.; Nordmann, P.; Schwarz, S. Antimicrobial Resistance in Escherichia coli. Microbiol. Spectr. 2018, 6, 1–27. [Google Scholar] [CrossRef]
- Barrios, H.; Garza-Ramos, U.; Mejia-Miranda, I.; Reyna-Flores, F.; Sanchez-Perez, A.; Mosqueda-Garcia, D.; Silva-Sanchez, J.; Bacterial Resistance, C. ESBL-producing Escherichia coli and Klebsiella pneumoniae: The most prevalent clinical isolates obtained between 2005 and 2012 in Mexico. J. Glob. Antimicrob. Resist. 2017, 10, 243–246. [Google Scholar] [CrossRef] [PubMed]
- Pehlivanoglu, F.; Turutoglu, H.; Ozturk, D. CTX-M-15-Type Extended-Spectrum Beta-Lactamase-Producing Escherichia coli as Causative Agent of Bovine Mastitis. Foodborne Pathog. Dis. 2016, 13, 477–482. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.T.; Zhang, L.J.; Lu, Y.; Zhang, R.M.; Jiang, H.X. Genomic Insights into Global bla(CTX-M-55)-Positive Escherichia coli Epidemiology and Transmission Characteristics. Microbiol. Spectr. 2023, 11, e0108923. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibiotic Class | Drug Agents | Drug Breakpoint (R S I, mm) | Percentage of Susceptible Strains (S) | Percentage of Intermediate Strains (I) | Percentage of Resistant Strains (R) |
|---|---|---|---|---|---|
| β-Lactams | Ampicillin | ≤13 14~16 ≥17 | 49.4% (43/87) | 13.8% (12/87) | 36.8% (32/87) |
| Amoxicillin/clavulanic acid | ≤13 14~17 ≥18 | 96.6% (84/87) | 2.3% (2/87) | 1.1% (1/87) | |
| Cefepime | ≤14 15~17 ≥18 | 89.7% (79/87) | 4.6% (4/87) | 5.7% (4/87) | |
| Ceftazidime | ≤14 15~17 ≥18 | 86.2% (75/87) | 6.9% (6/87) | 6.9% (6/87) | |
| Sulfonamides | Cotrimoxazole | ≤10 11~15 ≥16 | 69.0% (60/87) | 2.3% (2/87) | 28.7% (25/87) |
| Tetracyclines | Tetracycline | ≤14 15~18 ≥19 | 41.4% (36/87) | 20.7% (18/87) | 37.9% (33/87) |
| Phenicols | Florfenicol | ≤12 13~17 ≥18 | 87.4% (76/87) | 0.0% (0/87) | 12.6% (11/87) |
| Aminoglycosides | Kanamycin | ≤13 14~17 ≥18 | 69.0% (60/87) | 16.1% (14/87) | 14.9% (13/87) |
| Gentamycin | ≤12 13~14 ≥15 | 79.3% (69/87) | 3.4% (3/87) | 17.2% (15/87) | |
| Streptomycin | ≤14 15~18 ≥19 | 50.6% (44/87) | 14.9% (13/87) | 34.5% (30/87) | |
| Fluoroquinolones | Enrofloxacin | ≤14 15~20 ≥21 | 93.1% (81/87) | 2.3% (2/87) | 4.6% (4/87) |
| Polymyxin | Polymyxin B | ≤8 9~11 ≥12 | 97.7% (85/87) | 2.3% (2/87) | 1.1% (0/87) |
| Classes of Resistant Antibiotics | Number of Strains of Mastitis-Related E. coli | Percentage of Mastitis-Related E. coli | Percentage of Clinical Strains | Percentage of Subclinical Strains |
|---|---|---|---|---|
| 0 | 32 | 36.8% | 36.1% | 37.3% |
| 1 | 19 | 21.8% | 13.9% | 27.5% |
| 2 | 10 | 11.5% | 16.7% | 7.8% |
| 3 | 10 | 11.5% | 13.9% | 9.8% |
| 4 | 7 | 8.0% | 8.3% | 7.8% |
| 5 | 6 | 6.9% | 8.3% | 5.9% |
| 6 | 3 | 3.4% | 2.8% | 3.9% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zuo, J.; Lv, Z.; Lian, L.; Wu, Z.; Fu, S.; Zhang, H.; Wu, J.; Pan, Z.; Yu, Y.; Chen, W.; et al. Difference Analysis on Virulence Genes, Biofilms and Antimicrobial Susceptibility of Escherichia coli from Clinical and Subclinical Bovine Mastitis. Vet. Sci. 2025, 12, 132. https://doi.org/10.3390/vetsci12020132
Zuo J, Lv Z, Lian L, Wu Z, Fu S, Zhang H, Wu J, Pan Z, Yu Y, Chen W, et al. Difference Analysis on Virulence Genes, Biofilms and Antimicrobial Susceptibility of Escherichia coli from Clinical and Subclinical Bovine Mastitis. Veterinary Sciences. 2025; 12(2):132. https://doi.org/10.3390/vetsci12020132
Chicago/Turabian StyleZuo, Jiakun, Zhaoyang Lv, Liyan Lian, Zihao Wu, Shaodong Fu, Haiyang Zhang, Jing Wu, Zihao Pan, Yong Yu, Wei Chen, and et al. 2025. "Difference Analysis on Virulence Genes, Biofilms and Antimicrobial Susceptibility of Escherichia coli from Clinical and Subclinical Bovine Mastitis" Veterinary Sciences 12, no. 2: 132. https://doi.org/10.3390/vetsci12020132
APA StyleZuo, J., Lv, Z., Lian, L., Wu, Z., Fu, S., Zhang, H., Wu, J., Pan, Z., Yu, Y., Chen, W., Jiang, W., Yin, H., Chen, Z., Yi, Y., Han, X., & Miao, J. (2025). Difference Analysis on Virulence Genes, Biofilms and Antimicrobial Susceptibility of Escherichia coli from Clinical and Subclinical Bovine Mastitis. Veterinary Sciences, 12(2), 132. https://doi.org/10.3390/vetsci12020132