
Galacto-Oligosaccharide (GOS) Synthesis during Enzymatic Lactose-Free Milk Production: State of the Art and Emerging Opportunities


Abstract
:1. Introduction
2. Enzyme Sources and Factors Affecting GOS Production
3. Recent Advances in β-Galactosidase Immobilization for GOS Production
4. Conclusions and Perspectives
Author Contributions
Funding
Conflicts of Interest
References
- Ugidos-Rodríguez, S.; Matallana-González, M.C.; Sánchez-Mata, M.C. Lactose malabsorption and intolerance: A review. Food Funct. 2018, 9, 4056–4068. [Google Scholar] [CrossRef]
- Brüssow, H. Nutrition, population growth and disease: A short history of lactose. Environ. Microb. 2013, 15, 2154–2161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robert, K.Y.; Nakatani, Y.; Yanagisawa, M. The role of glycosphingolipid metabolism in the developing brain. J. Lipid Res. 2009, 50, S440–S445. [Google Scholar]
- Brenkert, A.; Radin, N.S. Synthesis of galactosyl ceramide and glucosyl ceramide by rat brain: Assay procedures and changes with age. Brain Res. 1972, 36, 183–193. [Google Scholar] [CrossRef] [Green Version]
- Nilsson, Å. Sphingolipids in the gut? Which are the important issues? Eur. J. Lipid 2007, 109, 971–976. [Google Scholar] [CrossRef]
- Naim, H.Y.; Sterchi, E.E.; Lentze, M.J. Biosynthesis and maturation of lactase-phlorizin hydrolase in the human small intestinal epithelial cells. Biochem. J. 1987, 241, 427–434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elferink, H.; Bruekers, J.P.J.; Veeneman, G.H.; Boltje, T.J. A comprehensive overview of substrate specificity of glycoside hydrolases and transporters in the small intestine: ‘A gut feeling’. Cell. Mol. Life Sci. 2020, 77, 4799–4826. [Google Scholar] [CrossRef]
- Lule, V.K.; Garg, S.; Tomar, S.K.; Khedkar, C.D.; Nalage, D.N. Food Intolerance: Lactose Intolerance. In Encyclopedia of Food and Health; Elsevier Inc.: Amsterdam, The Netherlands, 2015; pp. 43–48. [Google Scholar]
- Garballo-Rubio, A.; Soto-Chinchilla, J.; Moreno, A.; Zafra-Gómez, A. Determination of residual lactose in lactose-free cow milk by hydrophilic interaction liquid chromatography (HILIC) coupled to tandem mass spectrometry. J. Food Compos. Anal. 2018, 66, 39–45. [Google Scholar] [CrossRef]
- Suri, S.; Kumar, V.; Prasad, R.; Tanwar, B.; Goyal, A.; Kaur, S.; Gat, Y.; Kumar, A.; Kaur, J.; Singh, D. Considerations for development of lactose-free food. J. Nut. Inter. Met. 2019, 15, 27–34. [Google Scholar] [CrossRef]
- Dekker, P.J.T.; Koenders, D.; Bruins, M.J. Lactose-free dairy products: Market developments, production, nutrition and health benefits. Nutrients 2019, 11, 551. [Google Scholar] [CrossRef] [Green Version]
- Raza, A.; Iqbal, S.; Ullah, A.; Khan, M.I.; Imran, M. Enzymatic conversion of milk lactose to prebiotic galacto-oligosaccharides to produce low lactose yogurt. J. Food Proc.Preserv. 2018, 42, e13586. [Google Scholar] [CrossRef]
- Adhikari, K.; Dooley, L.M.; Chambers IV, E.; Bhumiratana, N. Sensory characteristics of commercial lactose-free milks manufactured in the United States. LWT-Food Sci. Technol. 2010, 43, 113–118. [Google Scholar] [CrossRef] [Green Version]
- Harju, M. Milk sugars and minerals as ingredients. Int. J. Dairy Technol. 2001, 54, 61–63. [Google Scholar] [CrossRef]
- Dutra Rosolen, M.; Gennari, A.; Volpato, G.; Volken de Souza, C.F. Lactose Hydrolysis in Milk and Dairy Whey Using Microbial β-Galactosidases. Enzymatic Res. 2015, 2015, 806240. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez-Colinas, B.; Fernandez-Arrojo, L.; Ballesteros, A.O.; Plou, F.J. Galactooligosaccharides formation during enzymatic hydrolysis of lactose: Towards a prebiotic-enriched milk. Food Chem. 2014, 145, 388–394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gosling, A.; Stevens, G.W.; Barber, A.R.; Kentish, E.S.; Gras, S.L. Recenti progressi nella raffinazione della produzione di galattooligosaccaridi a partire dal lattosio. Food Chem. 2010, 121, 307–318. [Google Scholar] [CrossRef]
- Fischer, C.; Kleinschmidt, T. Synthesis of galactooligosaccharides in milk and whey: A review. Compr. Rev. Food Sci. Saf. 2018, 17, 678–697. [Google Scholar] [CrossRef] [PubMed]
- Jenab, E.; Omidghane, M.; Mussone, P.; Armada, D.H.; Cartmell, J.; Montemagno, C. Enzymatic conversion of lactose into galacto-oligosaccharides: The effect of process parameters, kinetics, foam architecture, and product characterization. J. Food Eng. 2018, 222, 63–72. [Google Scholar] [CrossRef]
- Park, A.-R.; Oh, D.-K. Galacto-oligosaccharide production using microbial β-galactosidase: Current state and perspectives. App. Microb. Biotechnol. 2010, 85, 1279–1286. [Google Scholar] [CrossRef]
- Movahedpour, A.; Ahmadi, N.; Ghalamfarsa, F.; Ghesmati, Z.; Khalifeh, M.; Maleksabet, A.; Shabaninejad, Z.; Taheri-Anganeh, M.; Savardashtaki, A. β-Galactosidase: From its source and applications to its recombinant form. Biotechnol. Appl. Biochem. 2021. [CrossRef]
- Huang, J.; Zhu, S.; Zhao, L.; Chen, L.; Du, M.; Zhang, C.; Yang, S.T. A novel β-galactosidase from Klebsiella oxytoca ZJUH1705 for efficient production of galacto-oligosaccharides from lactose. App. Microb. Biotechnol. 2020, 104, 6161–6172. [Google Scholar] [CrossRef] [PubMed]
- Vera, C.; Guerrero, C.; Aburto, C.; Cordova, A.; Illanes, A. Conventional and non-conventional applications of β-galactosidases. Biochim. Et Biophys. Acta (BBA) Proteins Proteom. 2020, 1868, 140271. [Google Scholar] [CrossRef] [PubMed]
- Guerrero, C.; Vera, C.; Conejeros, R.; Illanes, A. Transgalactosylation and hydrolytic activities of commercial preparations of β-galactosidase for the synthesis of prebiotic carbohydrates. Enzyme Microb. Technol. 2015, 70, 9–17. [Google Scholar] [CrossRef] [PubMed]
- Martins, G.N.; Ureta, M.M.; Tymczyszyn, E.E.; Castilho, P.C.; Gomez-Zavaglia, A. Technological aspects of the production of fructo and enzymatic synthesis and hydrolysis. Front. Nutr. 2019, 6, 78. [Google Scholar] [CrossRef]
- Otieno, D.O. Synthesis of β-galactooligosaccharides from lactose using microbial β-galactosidases. Compr. Rev. Food Sci. Food Saf. 2010, 9, 471–482. [Google Scholar] [CrossRef] [PubMed]
- Zarate, S.; Lopex-Leiva, M.H. Oligosaccharide formation during enzymatic lactose hydrolysis: A literature review. J. Food Prot. 1990, 53, 262–268. [Google Scholar] [CrossRef] [PubMed]
- Prenosil, J.E.; Stuker, E.; Bourne, J.R. Formation of oligosaccharides during enzymatic lactose. Part I: State of art. Biotechnol. Bioeng. 1987, 30, 1019–1025. [Google Scholar] [CrossRef] [PubMed]
- de Albuquerque, T.L.; de Sousa, M.; Silva, N.C.G.; Neto, C.A.C.G.; Gonçalves, L.R.B.; Fernandez-Lafuente, R.; Rocha, M.V.P. β-Galactosidase from Kluyveromyces lactis: Characterization, production, immobilization and applications-A review. Int. J. Biol. Macromol. 2021, 191, 881–898. [Google Scholar] [CrossRef]
- Santos, A.; Ladero, M.; García-Ochoa, F. Kinetic Modeling of Lactose Hydrolysis by a β-Galactosidase from Kluyveromices Fragilis. Enzyme Microb. Technol. 1998, 22, 558–567. [Google Scholar] [CrossRef]
- Saqib, S.; Akram, A.; Halim, S.A.; Tassaduq, R. Sources of β-galactosidase and its applications in food industry. Biotech 2017, 7, 79. [Google Scholar] [CrossRef] [Green Version]
- Cho, Y.-J.; Shin, H.-J.; Bucke, C. Purification and biochemical properties of a galactooligosaccharide producing β-galactosidase from Bullera singularis. Biotechnol. Lett. 2003, 25, 2107–2111. [Google Scholar] [CrossRef] [PubMed]
- Petzelbauer, I.; Reiter, A.; Splechtna, B.; Kosma, P.; Nidetzky, B. Transgalactosylation by thermostable β-glycosidases from Pyrococcus furiosus and Sulfolobus solfataricus. Eur. J. Biochem. 2000, 267, 5055–5066. [Google Scholar] [CrossRef] [PubMed]
- Hsu, C.A.; Lee, S.L.; Chou, C.C. Enzymatic production of galactooligosaccharides by β-galactosidase from Bifidobacterium longum BCRC 15708. J. Agric. Food Chem. 2007, 55, 2225–2230. [Google Scholar] [CrossRef] [PubMed]
- Boon, M.A.; Janssen, A.E.M.; van’t Riet, K. Effect of temperature and enzyme origin on the enzymatic synthesis of oligosaccharides. Enzyme Microb. Technol. 2000, 26, 271–281. [Google Scholar] [CrossRef]
- Splechtna, B.; Nguyen, T.; Steinböck, M.; Kulbe, K.D.; Lorenz, W.; Haltrich, D. Production of Prebiotic Galacto-Oligosaccharides from Lactose Using β-Galactosidases from Lactobacillus reuteri. J. Agric. Food. Chem. 2006, 54, 4999–5006. [Google Scholar] [CrossRef] [PubMed]
- Iwasaki, K.I.; Nakajima, M.; Nakao, S.I. Galacto-oligosaccharide production from lactose by an enzymic batch reaction using β-galactosidase. Proc. Biochem. 1996, 31, 69–76. [Google Scholar] [CrossRef]
- Prenosil, J.E.; Stuker, E.; Bourne, J.R. Formation of oligosaccharides during enzymatic lactose hydrolysis and their importance in a whey hydrolysis process: Part II: Experimental. Biotechnol. Bioeng. 1987, 30, 1026–1031. [Google Scholar] [CrossRef]
- Ji, E.S.; Park, N.H.; Oh, D.K. Galacto-oligosaccharide production by a thermostable recombinant β-galactosidase from Thermotoga maritima. World J. Microb. Biot. 2005, 21, 759–764. [Google Scholar] [CrossRef]
- Nakao, M.; Harada, M.; Kodama, Y.; Nakayama, T.; Shibano, Y.; Amachi, T. Purification and characterization of a thermostable β-galactosidase with high transgalactosylation activity from Saccharopolyspora rectirgula. Appl. Microbiol. Biotechnol. 1994, 40, 657–663. [Google Scholar] [CrossRef]
- Pala-Ozkok, I.; Rehman, A.; Yagci, N.; Ubay-Cokgor, E.; Jonas, D.; Orhon, D. Characteristics of mixed microbial culture at different sludge ages: Effect on variable kinetics for substrate utilization. Bioresour. Technol. 2012, 126, 274–282. [Google Scholar] [CrossRef]
- Onishi, N.; Tanaka, T. Purification and properties of a novel thermostable galacto-oligosaccharide-producing beta-galactosidase from Sterigmatomyces elviae CBS8119. Appl. Environ. Microb. 1995, 61, 4026–4030. [Google Scholar] [CrossRef] [Green Version]
- Placier, G.; Watzlawick, H.; Rabiller, C.; Mattes, R. Evolved β-galactosidases from Geobacillus stearothermophilus with improved transgalactosylation yield for galacto-oligosaccharide production. Appl. Environ. Microb. 2009, 75, 6312–6321. [Google Scholar] [CrossRef] [Green Version]
- Carević, M.; Vukašinović-Sekulić, M.; Ćorović, M.; Rogniaux, H.; Ropartz, D.; Veličković, D.; Bezbradica, D. Evaluation of β-galactosidase from Lactobacillus acidophilus as biocatalyst for galacto-oligosaccharides synthesis: Product structural characterization and enzyme immobilization. J. Biosci. Bioeng. 2018, 126, 697–704. [Google Scholar] [CrossRef]
- Albayrak, N.; Yang, S.T. Production of galacto-oligosaccharides from lactose by Aspergillus oryzae b-galactosidase immobilized on cotton cloth. Biotechnol. Bioeng. 2002, 77, 8–19. [Google Scholar] [CrossRef] [PubMed]
- Chockchaisawasdee, S.; Athanasopoulos, V.I.; Niranjan, K.; Rastall, R.A. Synthesis of galacto-oligosaccharide from lactose using β-galactosidase from Kluyveromyces lactis: Studies on batch and continuous UF membrane-fitted bioreactors. Biotechnol. Bioeng. 2005, 89, 434–443. [Google Scholar] [CrossRef] [PubMed]
- Tzortzis, G.; Goulas, A.K.; Gibson, G.R. Synthesis of prebiotic galacto-oligosaccharides using whole cells of a novel strain, Bifidobacterium bifidum NCIMB 41171. Appl. Microb. Biotechnol. 2005, 68, 412–416. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, A.; Mishra, S.; Chand, S. Transgalactosylation of lactose for synthesis of galacto-oligosaccharides using Kluyveromyces marxianus NCIM 3551. New Biotechnol. 2015, 32, 412–418. [Google Scholar] [CrossRef] [PubMed]
- Huber, R.E.; Kurz, G.; Wallenfels, K.A. A quantitation of the factors which affect the hydrolase and transgalactosylase activities of β-galactosidase (E. coli) on lactose. Biochemistry 1976, 15, 1994–2001. [Google Scholar] [CrossRef] [PubMed]
- Abdul Manas, N.H.; Md. Illias, R.; Mahadi, N.M. Strategy in manipulating transglycosylation activity of glycosyl hydrolase for oligosaccharide production. Crit. Rev. Biotechnol. 2018, 38, 272–293. [Google Scholar] [CrossRef]
- Kim, C.S.; Ji, E.S.; Oh, D.K. A new kinetic model of recombinant β-galactosidase from Kluyveromyces lactis for both hydrolysis and transgalactosylation reactions. Biochem. Biophys. Res. Commun. 2004, 316, 738–743. [Google Scholar] [CrossRef] [PubMed]
- Guerrero, C.; Vera, C.; Acevedo, F.; Illanes, A. Simultaneous synthesis of mixtures of lactulose and galacto-oligosaccharides and their selective fermentation. J. Biotechnol. 2015, 209, 31–40. [Google Scholar] [CrossRef]
- Neto, C.A.C.G.; Silva, N.C.G.; de Oliveira Costa, T.; de Albuquerque, T.L.; Gonçalves, L.R.B.; Fernandez-Lafuente, R.; Rocha, M.V.P. The β-galactosidase immobilization protocol determines its performance as catalysts in the kinetically controlled synthesis of lactulose. Int. J. Biol. Macromol. 2021, 176, 468–478. [Google Scholar] [CrossRef] [PubMed]
- Ricardi, N.C.; Arenas, L.T.; Benvenutti, E.V.; Hinrichs, R.; Flores, E.E.E.; Hertz, P.F.; Costa, T.M.H. High performance biocatalyst based on β-D-galactosidase immobilized on mesoporous silica/titania/chitosan material. Food Chem. 2021, 359, 129890. [Google Scholar] [CrossRef]
- Guerrero, C.; Vera, C.; Serna, N.; Illanes, A. Immobilization of Aspergillus oryzae β-galactosidase in an agarose matrix functionalized by four different methods and application to the synthesis of lactulose. Bioresour. Technol. 2017, 232, 53–63. [Google Scholar] [CrossRef]
- Liburdi, K.; Benucci, I.; Esti, M. Lysozyme in Wine: An Overview of Current and Future Applications. Compr. Rev. Food Sci. Food Saf. 2014, 13, 1062–1073. [Google Scholar] [CrossRef]
- Panesar, P.S.; Panesar, R.; Singh, R.S.; Kennedy, J.F.; Kumar, H. Review Microbial production, immobilization and applications of β-D-galactosidase. J. Chem. Technol. Biotechnol. Int. Res. Process Environ. Clean Technol. 2006, 81, 530–543. [Google Scholar]
- Xavier, J.R.; Ramana, K.V.; Sharma, R.K. β-galactosidase: Biotechnological applications in food processing. J. Food Biochem. 2018, 42, e12564. [Google Scholar] [CrossRef]
- Serey, M.; Vera, C.; Guerrero, C.; Illanes, A. Immobilization of Aspergillus oryzae β-galactosidase in cation functionalized agarose matrix and its application in the synthesis of lactulose. Int. J. Biol. Macromol. 2021, 67, 1564–1574. [Google Scholar] [CrossRef]
- Guerrero, C.; Aburto, C.; Suárez, S.; Vera, C.; Illanes, A. Effect of the type of immobilization of β-galactosidase on the yield and selectivity of synthesis of transgalactosylated oligosaccharides. Biocatal. Agric. Biotechnol. 2018, 16, 353–363. [Google Scholar] [CrossRef]
- Eijsink, V.; Bjørk, A.G.; Gåseidnes, S.; Sirevåg, R.; Synstad, B.; van den Burg, B.; Vriend, G. Rational engineering of enzyme stability. J. Biotechnol. 2004, 113, 105–120. [Google Scholar] [CrossRef]
- Rueda, N.; dos Santos, J.C.S.; Torres, R.; Ortiz, C.; Barbosa, O.; Fernandez-Lafuente, R. Improved performance of lipases immobilized on heterofunctional octyl-glyoxyl agarose beads. RSC Adv. 2015, 5, 11212–11222. [Google Scholar] [CrossRef] [Green Version]
- Barbosa, O.; Torres, R.; Ortiz, C.; Berenguer-Murcia, Á.; Rodrigues, R.C.; Fernandez-Lafuente, R. Heterofunctional supports in enzyme immobilization: From traditional immobilization protocols to opportunities in tuning enzyme properties. Biomacromolecules 2013, 14, 2433–2462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mateo, C.; Grazu, V.; Palomo, J.M.; Lopez-Gallego, F.; Fernandez-Lafuente, R.; Guisan, J.M. Immobilization of enzymes on heterofunctional epoxy supports. Nat. Protoc. 2007, 2, 1022–1033. [Google Scholar] [CrossRef] [PubMed]
- Urrutia, P.; Bernal, C.; Wilson, L.; Illanes, A. Use of chitosan heterofunctionality for enzyme immobilization: β-galactosidase immobilization for galacto-oligosaccharide synthesis. Int. J. Biol. Macromol. 2018, 116, 182–193. [Google Scholar] [CrossRef]
- Warmerdam, A.; Benjamins, E.; de Leeuw, T.F.; Broekhuis, T.A.; Boom, R.M.; Janssen, A.E.M. Galacto-oligosaccharide production with immobilized β-galactosidase in a packed-bed reactor vs. free β-galactosidase in a batch reactor. Food Bioprod. Process. 2014, 92, 383–392. [Google Scholar] [CrossRef]
- Carević, M.; Ćorović, M.; Mihailović, M.; Banjanac, K.; Milisavljević, A.; Veličković, D.; Bezbradica, D. Galacto-oligosaccharide synthesis using chemically modified β-galactosidase from Aspergillus oryzae immobilised onto macroporous amino resin. Int. Dairy J. 2016, 54, 50–57. [Google Scholar] [CrossRef]
- Osman, A.; Symeou, S.; Trisse, V.; Watson, K.A.; Tzortzis, G.; Charalampopoulos, D. Synthesis of prebiotic galactooligosaccharides from lactose using bifidobacterial β-galactosidase (BbgIV) immobilised on DEAE-Cellulose, Q-Sepharose and amino-ethyl agarose. Biochem. Eng. J. 2014, 82, 188–199. [Google Scholar] [CrossRef] [Green Version]
- Cappannella, E.; Benucci, I.; Lombardelli, C.; Liburdi, K.; Bavaro, T.; Esti, M. Immobilized lysozyme for the continuous lysis of lactic bacteria in wine: Bench-scale fluidized-bed reactor study. Food Chem. 2016, 210, 49–55. [Google Scholar] [CrossRef]

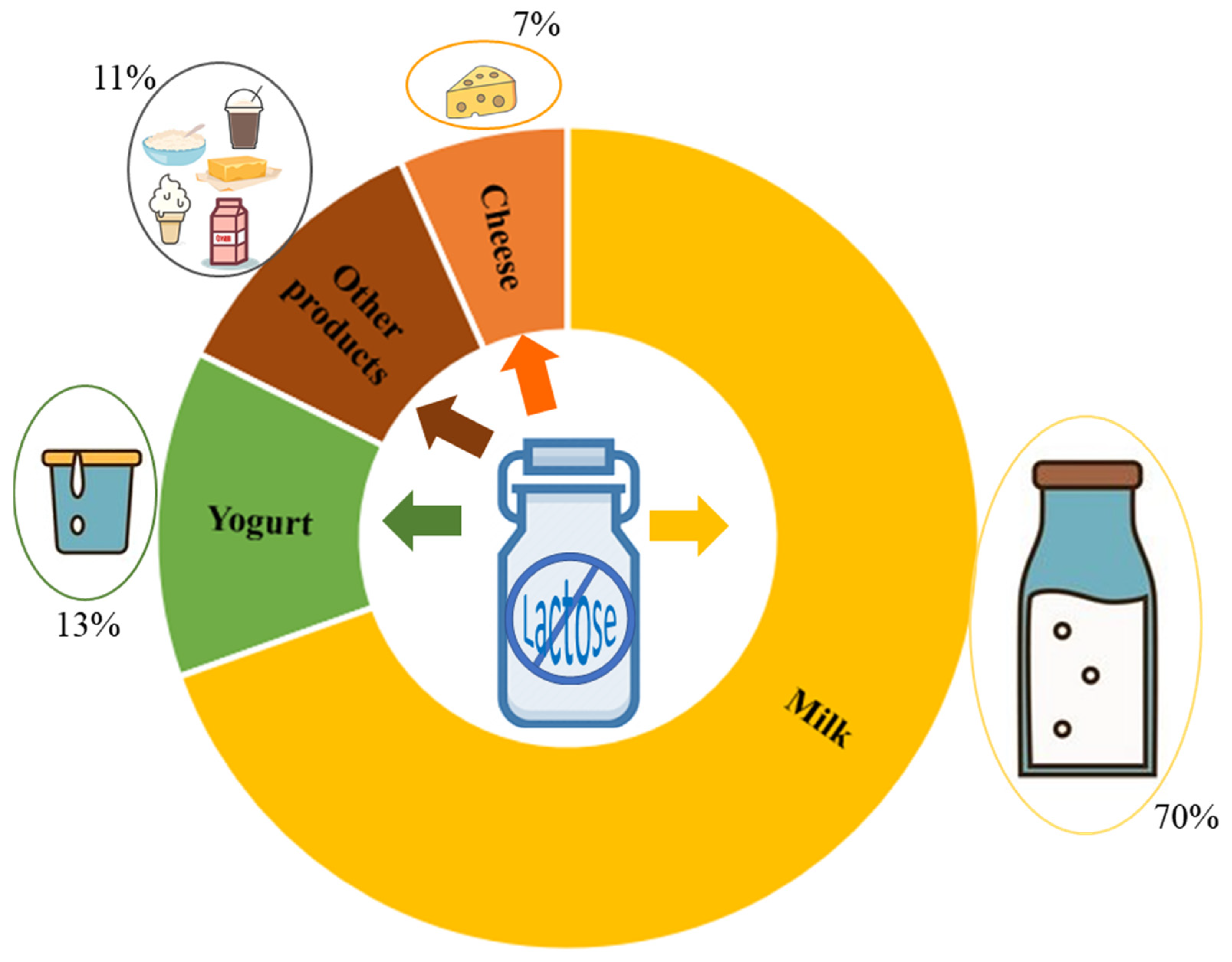

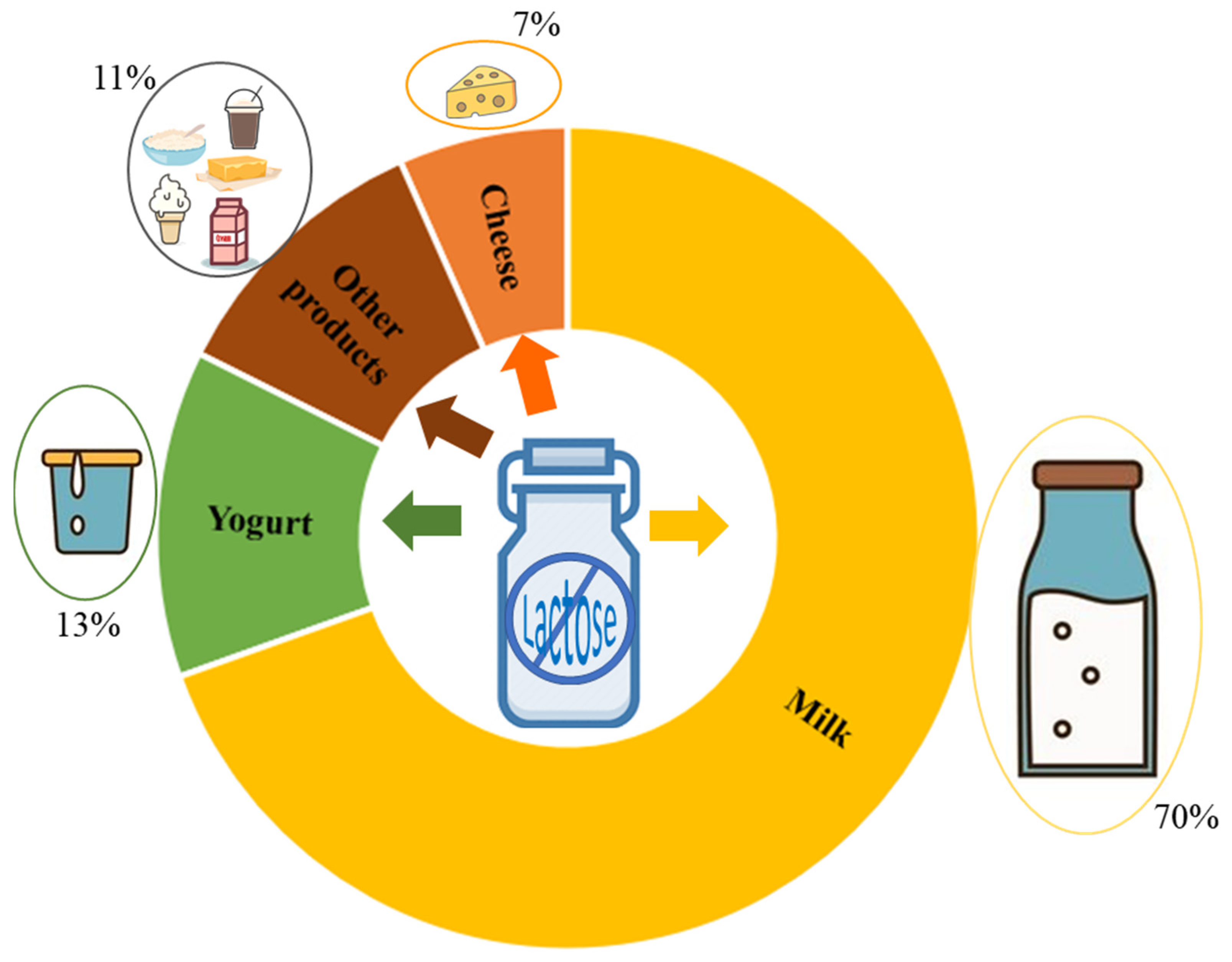
{kind=link}
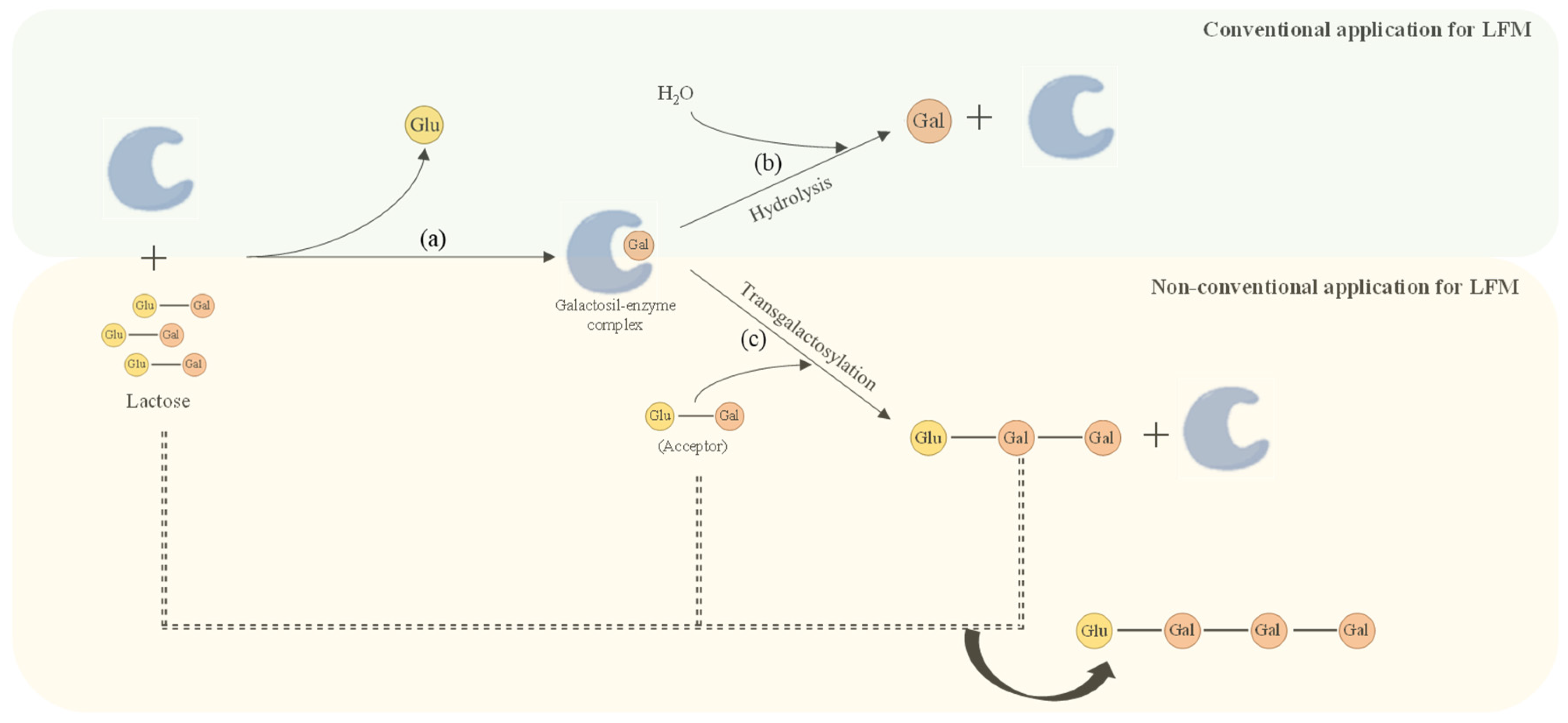
{kind=link}
{kind=link}
| Microrganisms | Topt (°C) | pHopt | Maximum GOS Yield (%, w/w) | References | |
|---|---|---|---|---|---|
| Hyperthermophiles | Thermotoga maritima | 80–85 | 6.5 | 19 | [39] |
| Extreme thermofiphiles | Sulfolobus solfataricus | 80 | 6.5 | 41 | [40] |
| Thermus aquaticus | 80 | 5.5 | 39 | [41] | |
| Pyrococcus furiosus | 80 | 5.0 | 33 | [33] | |
| Moderate thermophiles | Saccharopolyspora rectivirgula | 70 | 7.0 | 44 | [40] |
| Sterigmatomyces elviae | 60 | 5.0 | 39 | [42] | |
| Geobacillus stearothermophilus | 70 | 7.7 | 18.6 | [43] | |
| Lactobacillus acidophilus | 55 | 6.5–8 | 25.5 | [44] | |
| Bullera singularis | 50 | 6.0 | 50 | [32] | |
| Lactobacillus reuteri | 50 | 6.5–8 | 38 | [36] | |
| Bifidobacterium longum | 45 | 6.8 | 32.5 | [34] | |
| Mesophiles | Aspergillus oryzae | 40 | 4.5 | 27 | [45] |
| Kluyveromyces lactis | 40 | 7.0 | 25 | [46] | |
| Bifidobacterium bifidum | 37 | 6.5 | 20 | [47] | |
| Kluyveromyces marxianus | 30 | 6.5 | 25 | [48] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liburdi, K.; Esti, M. Galacto-Oligosaccharide (GOS) Synthesis during Enzymatic Lactose-Free Milk Production: State of the Art and Emerging Opportunities. Beverages 2022, 8, 21. https://doi.org/10.3390/beverages8020021
Liburdi K, Esti M. Galacto-Oligosaccharide (GOS) Synthesis during Enzymatic Lactose-Free Milk Production: State of the Art and Emerging Opportunities. Beverages. 2022; 8(2):21. https://doi.org/10.3390/beverages8020021
Chicago/Turabian StyleLiburdi, Katia, and Marco Esti. 2022. "Galacto-Oligosaccharide (GOS) Synthesis during Enzymatic Lactose-Free Milk Production: State of the Art and Emerging Opportunities" Beverages 8, no. 2: 21. https://doi.org/10.3390/beverages8020021
APA StyleLiburdi, K., & Esti, M. (2022). Galacto-Oligosaccharide (GOS) Synthesis during Enzymatic Lactose-Free Milk Production: State of the Art and Emerging Opportunities. Beverages, 8(2), 21. https://doi.org/10.3390/beverages8020021