Abstract
It is shown that bacteria Bradyrhizobium japonicum 273 were capable of degrading phenol at moderate concentrations either in a free cell culture or by immobilized cells on granulated activated carbon particles. The amount of degraded phenol was greater in an immobilized cell preparation than in a free culture. The application of a constant electric field during cultivation led to enhanced phenol biodegradation in a free culture and in immobilized cells on granulated activated carbon. The highest phenol removal efficiency was observed for an anode potential of 1.0 V/S.H.E. The effect was better pronounced in a free culture. The enzyme activities of free cells for phenol oxidation and benzene ring cleavage were very sensitive to the anode potential in the first two steps of the metabolic pathway of phenol biodegradation catalyzed by phenol hydroxylase—catechol-1,2-dioxygenase and catechol-2,3-dioxygenase. It was observed that at an anode potential of 0.8 V/S.H.E., the meta-pathway of cleavage of the benzene ring catalyzed by catechol-2,3-dioxygenase became competitive with the ortho-pathway, catalyzed by catechol-1,2-dioxygenase. The obtained results showed that the positive effect of constant electric field on phenol biodegradation was rather due to electric stimulation of enzyme activity than electrochemical anode oxidation.
1. Introduction
Phenol and its derivatives are widely distributed as a source and product of various human activities (plastics, pesticides, etc.) They are considered as some of the most dangerous organic pollutants released in the environment. Phenol has adverse effects on the nervous system, respiratory tract [1], skin and mucous membranes [2], liver and kidneys [3], etc. Various methods have been proposed to decrease its concentration in waste streams to harmless levels. These methods are based mostly on adsorption and ion exchange [4,5,6] or microbial processes [7,8].
Various microbial species have been tested and reported for phenol biodegradation: bacterial strains from the genus Pseudomonas [7,9,10,11], Arthrobacter [12], Bacillus [13], etc.
Under aerobic conditions, microbial degradation of phenol usually starts by oxygenation, in which the aromatic ring is initially mono-hydroxylated at an ortho- position to the pre-existing hydroxyl group to form catechol. The active enzyme in this reaction is phenol hydroxylase for different microbial strains.
Further, catechol is oxidized following two different pathways: with o-benzoquinone formation [14] or with cleavage of the benzene ring [6].
Depending on the type of the strain’s metabolism, the ring cleavage of catechol can occur either at the ortho-position or at the meta-position. The orthi-pathway leads to succinic acid and finally to acetyl CoA. The meta-pathway leads to the formation of pyruvate and acetaldehyde; cf. [9].
There is another bacterial strain—namely, Bradyrhizobium japonicum 273, which is well known for biodegrading organic substances including aromatic compounds [15]. In previous studies, we succeeded in using it for the biodegradation of phenol by a free culture; cf. Vasileva et al. [16].
For redox biochemical reactions, the idea to combine microbial processes in aqueous media with an electrochemical effect on them seems promising when intermediates of the redox pathway are inhibitors for further biodegradation. Their removal can be performed either by oxidation or reduction on an electrode. Therefore, it is expected that electrochemical reactions on the electrode might enhance the complete microbial biodegradation of the substrate. The effects of constant electric fields on microbial activity were studied for different redox systems—namely, biodegradation of organic matter by Jobin and Namour [17] and Liu et al. [18], including phenol biodegradation [19,20].
Recently, Beschkov et al. [11] have shown the enhancing effect of applying a constant electric field on microbial phenol biodegradation by the strain Pseudomonas putida. In that study, it was shown that the enhancement was not assigned to purely electrochemical processes but to enzyme stimulation by the electric field. Moreover, it was shown that the catechol oxidation was dependent on the enzyme activity stimulated by the constant electric field at certain anode potential. The ortho-cleavage pathway (cf. Figure 1) was proven.
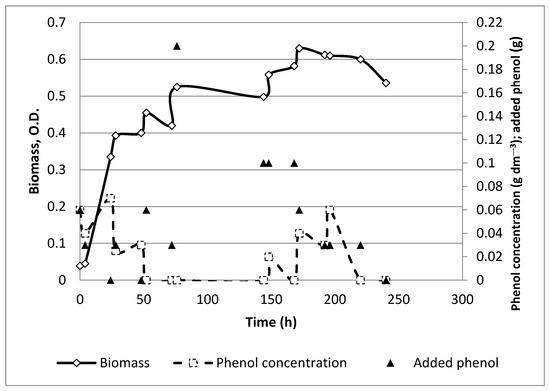
Figure 1.
Time profile for control experiment with initial phenol concentration as 0.06 g dm−3.
We could not find data for the phenol biodegradation by bacteria from the strain B. japonicum in a constant electric field in the literature. That is why it is interesting to test the effect of applying a constant electric field on the ability of the strain to degrade phenol and to check its effect on the pathway of phenol destruction, either for a free or for an immobilized culture.
The goal of this study is to test the bioelectrochemical effect on the degradation of phenol by a pure microbial monoculture of B. japonicum 273 in a potentiostatic mode for free and immobilized cultures.
Generally, the questions to answer in the present paper are as follows:
- How does the electric field affect the microbial growth of this strain?
- How is the optimum anode potential for bioelectrochemical oxidation of phenol established?
- How does the application of an electric field affect enzyme oxidation of phenol and catechol?
- How is enzyme activity affected by the electric field for free cultures?
- What is the impact of the electric field on the microbial cells immobilized on activated carbon to degrade phenol combined with bioelectrochemical stimulation?
2. Materials and Methods
2.1. Free Cell Cultivation
The strain B. japonicum 273 was obtained from the National Bank for Industrial Microorganisms and Cell Cultures, Bulgaria (NBIMCC). The strain was grown in mineral medium (MMY) containing (per liter): 1 g of yeast extract, 0.2 g of NaCl, 0.2 g of MgSO4, 0.5 g of K2HPO4, 10 g of Glucose; 1 L of tap water. The medium pH was adjusted to 7.2 before being autoclaved.
First, experiments were carried out in flasks of 250 mL in a rotary shaker at 28 °C and agitation speed of 100 rpm. In each flask, the volume of cultivated inoculums was 30 mL at different initial phenol concentrations, ranging from 0.06 to 0.15 g dm−3. It was found out that strong substrate inhibition occurred. Acceptable phenol biodegradation was observed at an initial concentration of 0.06 g dm−3. Therefore, our experiments were made at this starting concentration.
2.2. Immobilization Technique
The cells of the used strain were immobilized on granulated activated carbon (Fujikasui, Japan, with a specific area of 680 m2/g). The activated carbon was chosen as a carrier because of its high adsorption capacity, and therefore, for the protection of the cells at higher phenol concentrations.
The microbial suspension was prepared in the same way as for the experiments with free cell culture; cf. Section 2.1. The cells in their exponential phase of growth were mixed with washed and dried GAC particles at a particles/medium volume ratio of 10%. Then, the mixture was agitated in a rotary shaker at 200 rpm and 30 °C for 48 h.
The same volumetric ratio was used for the experiments in the fermentor.
2.3. Experimental Conditions
Fermentations both in free culture and by immobilized cells were carried out in a lab-scale, 0.5 L volume fermentor (Bioflo, a New Brunswick Inc. production, Sanford, FL, USA) at 30 °C and continuous stirring (100 rpm) with aeration rate 1 vvmin−1. The constant electric field was applied by anode, which was dipped in the broth. One of the stainless steel baffles of the fermentor was used as an anode. The counter-electrode was placed outside the fermentor with connection by an agar salt bridge. The anode potential was maintained constant by potentiostat to a quinhydrone reference electrode separated from the fermentation broth by a membrane of sintered glass. The applied anode potentials were 0.8, 0.9, and 1 V/S.H.E.
The fermentation media were identical to the one for strain cultivation, i.e., MMY, cf. Section 2.1. These media differed by the initially added amount of phenol. During the runs, phenol was introduced into the broth in a fed-batch mode when it was already exhausted. The total amount of degraded phenol for each run was considered as a reference for the process efficiency of the run.
Samples from the broth were analyzed for biomass and phenol concentration. For each applied voltage and for the control experiments, samples were also tested for enzyme activity, namely for phenol hydroxylase, 1,2-catechol dioxygenase, and for 2,3-catechol dioxygenase. The samples were analyzed in triplicate, and the mean values were presented.
The electric current was monitored during the runs. Its values were used for calculation of the electrochemical input in the overall biodegradation process by Faraday‘s law as follows:
where
i—electric current, A;
m—mass of reacting substance, g;
t—time, s;
M—molar mass of reacting substance, g;
n—number of exchanged electrons;
F = 96,484 C mole−1, Faraday constant.
2.4. Effect of Physical Adsorption
It must be emphasized that there can be multiple effects of the activated carbon: as adsorbent for phenol and its degradation products, as a carrier for the immobilized cells, and as a catalyst for the abiotic phenol oxidation. The abiotic contribution due to adsorption of phenol on the activated carbon was estimated in a separate test with cell-free carbon particles. For this purpose, 150 mL of granulated activated carbon were mixed with a a 500 mL cultivation medium. The mixture was stirred for 48 h with intermittent addition of 4 g of phenol in total. Afterward, the adsorbed phenol was extracted in 1 N NaOH solution and analyzed. The sample of the alkaline extract was analyzed qualitatively by NMR.
2.5. Analyses
2.5.1. Phenol Concentration
Phenol was determined photometrically by a standard colorimetric method based on the formation of a red-colored compound by treating the phenol-containing sample with 3.5% 4 amino antipyrine, 20% ammonium persulfate in the presence of a buffer (50 g of NH4Cl in 900 mL of distilled water with pH adjusted by ammonia to 9.3). The sample was centrifuged and then treated with the abovementioned reagents, and after 15 min, the sample absorbance was measured at wavelength λ = 540 nm. The concentration of phenol was calculated from a calibration straight line.
The qualitative content of the adsorbed phenol on the carbon particles and degradation products in the broth after 140 h of fermentation was determined by the NMR technique. The NMR spectra are acquired in water with presaturation of the water signal.
2.5.2. Biomass
The biomass in the samples was estimated photometrically by its optical density at wavelength λ = 610 nm on a Specol spectrophotometer (Carl Zeiss, Jena, Germany).
2.5.3. Phenol Hydroxylase Activity
This enzyme activity was measured by oxidation of NADPH, as a specific co-substrate. We studied the decrease in UV absorption at 340 nm; cf. Hintereger et al. [10]. The reaction mixture contained 2.7 mL of 0.1 M K-phosphate buffer (pH = 7.0); 0.1 mL of 2.5 mM phenol; 0.1 mL 5 mM NADPH and enzyme extract (0.1–0.15 mL cleared cell lysate). The reaction was started by introducing the substrate into the reaction mixture. Readings were carried out in 30 s for 3 min. An enzyme unit is defined as the amount of enzyme that, in the presence of phenol, oxidizes 1 μmol of NADPH for 1 min. Specific enzyme activity is calculated as enzyme units per 1 mg of protein (U/mg protein).
2.5.4. Catechol 1,2-Dioxygenase Activity
The spectrophotometrical method for the determination of cis,cis-muconic acid accumulation is a spectrophotometric one, consisting in measuring the increase in UV absorption at 260 nm in the linear range of the calibration photometric curve [21]. The reaction mixture contained 2.89 mL 0.1 M K-phosphate buffer (pH = 7.0) and 0.06 mL 0.01 M catechol. The reaction was started by introducing an enzyme extract (0.05 mL cleared cell lysate). The readings were carried out over 30 s for 3 min. An enzyme unit is defined as the amount of enzyme that catalyzes the formation of 1 μmol cis, cis-muconic acid for 1 min. The specific enzyme activity is calculated as enzyme units per 1 mg protein (U/mg protein).
2.5.5. Catechol 2,3-Dioxygenase Activity
The activity of the cell-free and immobilized catechol 2,3-dioxygenase was determined spectrophotometrically with the measurement of absorbance at λ = 375 nm, as described by Baggi et al. [22].
2.5.6. Scanning Electron Microscopy (SEM)
Microscopic examinations on the impact of the constant electric field on cell morphology were made by scanning electron microscopy. Both intact free cells and cells after application of electric field were observed.
Microscopic examinations of free and immobilized cells were performed on a scanning electron microscope by the following SEM sample procedure: The free cell samples were washed with 0.1 M sodium cacodylate (Fluka) pH 7.2. Then, they were fixed by 4% glutaraldehyde (Fluka) in 0.1 M sodium cacodylate at 4 °C for 2 h, followed by three washes with cacodyl buffer. Post-fixation of the samples was performed in 0.1% OsO4 in 0.1 M sodium cacodylate. This was followed by dehydration in an ascending ethanol series starting from 30% to 70% ethanol. The observation was made with a scanning electron microscope SEM Philips 515, digitized, with image detectors with secondary electrons (SEI) [23].
Microscopic examination of immobilized cells was performed on a scanning electron microscope (Inovenso Inc., IEM11 Philips 515, Eindhoven, The Netherlands). The particles of activated carbon with immobilized cells were taken out of a culture medium and washed with a Na cacodylate buffer. A fixation with glutaraldehyde was then performed, followed by dehydration with ethanol. The gold coating was performed in vacuo. The method has been described in detail in a previous publication [24].
3. Results
3.1. Physical Adsorption of Phenol by Granulated Activated Carbon
It was established that the residual amount of phenol in the cell-free carbon particles after 48 h of adsorption was 0.465 g. Considering the total amount of added phenol (i.e., 4 g), we concluded that the residual adsorbed phenol was 11.6% wt. from the initial amount. Obviously, the rest was oxidized with the help of the immobilized cells and on the particle’s active sites. No other compounds except phenolate were found in the alkaline extract from the used carbon particles.
3.2. Experiments with Free Cells without a Constant Electric Field
Two separate control experiments were carried out. The results for one of them are shown in Figure 1. Although the microbial growth curves were somewhat different, the total amounts of digested phenol were similar (0.93 and 0.86 g), i.e., 0.9 g in average; cf. Table 1.

Table 1.
Total amounts of degraded phenol and specific bacterial growth rates during the experiments. Initial phenol concentration, 0.06 g dm−3.
3.3. Experiments with Free Cells with a Constant Electric Field
Some experimental results on the kinetics of phenol biodegradation under the effect of a constant electric field in a free culture are shown in Figure 2A. The data for all experiments are summarized in Table 1. The specific microbial growth rates were not affected considerably by the electric field.
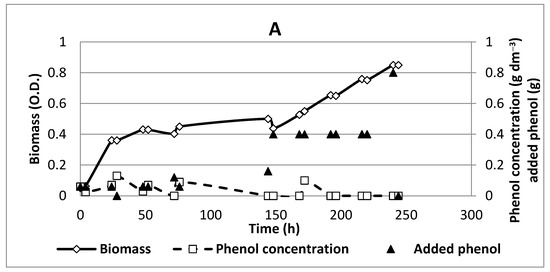
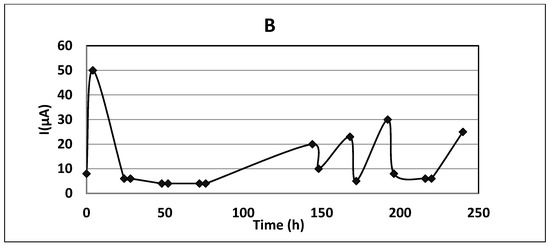
Figure 2.
Time profiles for an experiment with free cells and the application of a constant electric field: (A) anode potential 1.0 V/S.H.E; (B) the current measured at anode potential 1.0 V/S.H.E. Initial phenol concentration, 0.06 gdm−3.
Although the kinetic curves were not quite different, it is clear from the kinetic data and the ones in Table 1 that the constant electric field enhanced the biodegradation activity of the strain, particularly for the two higher anode potentials, reaching up to four times higher phenol concentrations in the broth.
The monitored current values were microamperes of orders of magnitude (Figure 3B), too low to expect electrochemical input for the total biodegradation process. Calculations using Faraday’s law (cf. Equation (1)) for the measured electric currents yielded about 1 mg degraded phenol for 240 h run, whereas the degraded amounts were 4.24 and 6.56 g phenol at free and immobilized cultures, respectively. Similar effects were observed for phenol biodegradation by P. putida [11].

Figure 3.
SEM image of free cell samples for SEM analysis: (A) control experiment; (B) with the constant electric field (0.8 V).
An illustration of the influence of the constant electric field on the morphology of the cells is shown in Figure 3. SEM observations were made on cells of Bradyrhizobium japonicum strain 273 utilizing phenol for 240 h. A constant electric field was applied during the experiment, while no electric impact was applied for the control sample.
Relatively intact morphological characteristics, shape, and size of the strain were observed in the control sample (Figure 3A). The trend indicated ubiquitous morphological damage to the cells in the sample treated by an electric field (Figure 3B). Cell surface indentations recorded in more than one area were reported, but the observed phenomenon was not ubiquitous. In some areas of the cell population, some cells emptied of cell contents were observed. As a result of longer treatment, the strain was likely to synthesize an exopolysaccharide in response to stressful conditions.
3.4. Effect of Electric Field on Enzyme Activity of Free Cells
The experimental results for the enzyme activities with and without application of an electric field in a free cell culture are shown in Table 2. For the control experiments without the application of an electric field, all enzyme activities grew in time. In cases when a constant electric field was applied, the situation became quite interesting. At an anode potential of 0.8 V/S.H.E., the phenol hydrolase was more active at the 144th hour. The catechol-1,2-dioxygenase was more active in the first two samples, whereas at t = 240 h, the control experiment provided better results. The ortho-pathway was predominant for the control experiment. Peculiarly, the catechol-2,3-dioxygenase became very active when the electric field was applied, at 0.8 and 0.9 V/S.H.E. This means that a competitive meta-pathway under these conditions was observed. In the case of anode potential of 1.0 V/S.H.E., the ortho-pathway prevailed, reaching the highest enzyme activity (1.67 U/mg protein) at the end of the process, t = 240 h. At this anode potential, the meta-pathway was not convincing. These results are illustrated in Figure S1.
Table 2.
Maximum enzyme activities at different anode potentials. Initial phenol concentration, 0.06 g dm−3.
3.5. Experiments with Immobilized Cells without a Constant Electric Field
The experimental results of phenol biodegradation by immobilized cells without the application of a constant electric field are shown in Figure 4. The cell concentration in the media was too low, and no detectable increase in biomass concentration was observed during the experiments for 250 h.
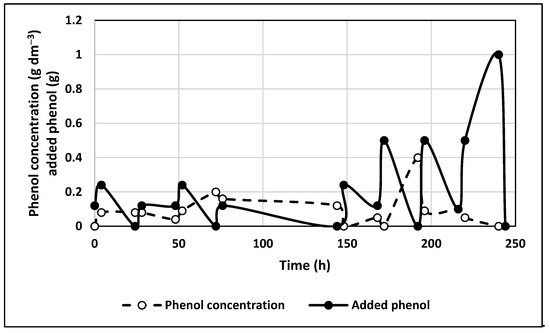
Figure 4.
Time profile for experiment with immobilized bacteria without electric field application. Initial phenol concentration, 0.06 g dm−3.
1H NMR spectra of the initial phenol solution of an alkaline extract from a carbon particle after physical adsorption and of the broth after the operation are shown in Figure S2 (Supplementary Materials). No other adsorbed compounds except phenolate in the alkaline extract were found by the NMR test; cf. Figure S2a, spectrum B. That is why the positive effect of immobilization could be sought in the microbial process of biodegradation facilitated by the parallel adsorption, leading to reduced phenol concentration in the vicinity of the immobilized cells.
3.6. Experiments in Immobilized Culture with the Application of a Constant Electric Field
One can infer that the positive effect of the applied electric field on phenol biodegradation was stronger for free cultures, compared with the immobilized ones; cf. Table 1. For free cells, the total biodegraded amount at an anode potential of 1.0 V/S.H.E. was about five times more than for the reference experiments. For immobilized cells, the electric field gave only 67% more degraded phenol than the reference experiment with immobilized cells growing without the influence of electric current.
This effect could be due to the abiotic physical adsorption present in all cases (with and without application of electric field) because of the accumulation of cells on the surface of the carrier, as well as the penetration of cells below the exposed surface of the carrier. As a result, the direct influence on all cells of the strain was limited, and accordingly, there was limited influence from the electric field. The local immobilization of microbial cells was proven by SEM after 240 h of phenol degradation. A significant amount of intact cells was observed on the carbon surface (Figure 5).

Figure 5.
SEM image of the activated carbon surface with attached bacterial cells.
The comparison of the electric current efficiency, calculated according to Faraday’s law, and the amount of biodegraded phenol versus chemical analysis, showed that the electrochemical contribution to the phenol biodegradation in the case of immobilized cells was negligible. Therefore, there was again bioelectrochemical stimulation but basically on the microbial cells’ enzyme activity.
Experimental results on phenol biodegradation by immobilized cells in a constant electric field are shown in Figure 6.
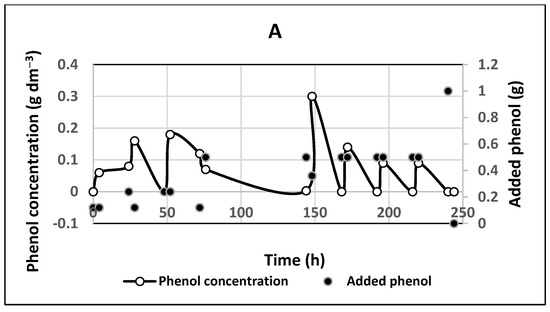
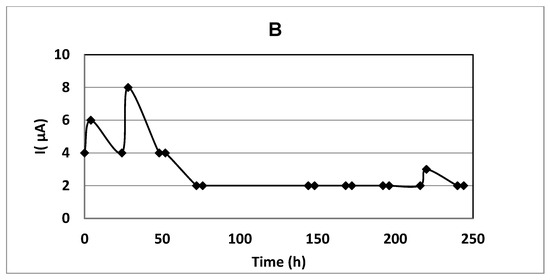
Figure 6.
Experiment with immobilized cells with a constant electric field at anode potential 1.0 V/S.H.E. Initial phenol concentration 0.06 g dm−3: (A) time profiles for added phenol and phenol concentration; (B) the electric current in immobilized cell culture.
On the other hand, no phenol or other aromatic compounds were detected in the broth after 140 h of electric field exposure; cf. sample C, Figure S2a,b. Hence, the electric field enhanced considerably the biodegradation activity even of the immobilized cells. Only traces of aliphatic compounds were present in the broth; cf. Figure S2b, spectrum C. The total digested amount was four times greater than for the free cultures for the same period; cf. Table 1. It was observed that the residual concentrations of phenol in the broth were much lower (even zero), compared with the case without an applied electric field; cf. Figure 5.
4. Discussion
The obtained experimental results showed that the constant electric field enhanced the biodegradation activity of the strain, particularly for the two higher anode potentials, reaching up to four times higher phenol concentrations in the broth. There was a synergy on phenol biodegradation of the electric field and activity of the immobilized cells by the used strain, leading to seven times higher biodegradation efficiency.
The measured very low electric current efficiency, compared with the analytically determined biodegraded amounts of phenol, led to the conclusion that there was practically no electrochemical anode oxidation of phenol. The electric field caused some biochemical effects, possibly some steric changes in the enzyme active sites, thus affecting the activation energies of the enzyme reactions of phenol oxidation and benzene ring cleavage. The results for the activity of the studied enzymes showed that their activities were very sensitive to the anode potential. The probable mechanism of the observed enzyme sensitivity toward the anode potential may be related to certain configurational changes in the active or allosteric centers of the enzymes. As a support of this explanation, a study on the effect of constant electric field on the activation energy for nitrate reduction can be indicated [25]. However, the comprehensive explanation of these effects is beyond the scope of this study.
The synergetic effect of the constant electric field and the cell immobilization could be explained by the additional physical adsorption of phenol on the carbon particles. One may suppose that adsorption played a dominant role in phenol removal from the broth. However, the experiment on phenol extraction from the carbon particles by a solution of sodium hydroxide showed a small amount of residual phenol adsorbed on the GAC surface after fermentation with no electric field (12% of the total added amount). On the other hand, no phenol was detected in the broth after 140 h of exposure to the electric field; cf. sample C, Figure S1a,b. Only traces of aliphatic compounds were present in the broth; cf. Figure S1b, spectrum C. Thus, it could be supposed that the immobilized bacteria were also considerably affected by the constant electric field.
The effect of the immobilized cells could also be explained by the accumulation of cells on the surface of the carrier, as well as the penetration of cells below the exposed surface of the carrier. As a result, the direct influence on all cells of the strain was limited, and accordingly, there was limited influence from the electric field. Obviously, the effect of the constant electric field was also similar, i.e., no electrochemical input for phenol degradation was relevant.
As a whole, the combined effect of microbial cell immobilization on activated carbon and of the applied constant electric field gave more than seven times larger amount of completely destroyed phenol than for the free culture with no electric field; cf. Table 1. This result leads to the suggestion for a practical application of immobilized cells with bioelectrochemical stimulation in continuous culture.
5. Conclusions
The following conclusions were drawn from the present experimental results:
- The bacteria Bradyrhizobium japonicum 273 were capable to degrade phenol at moderate concentrations either in a free culture or as immobilized ones on granulated activated carbon particles. The amount of degraded phenol was greater in an immobilized cell preparation than in a free culture. This fact could be explained by the additional effects of physical adsorption and, therefore, the facilitated biodegradation.
- The constant electric field applied during cells cultivation led to increased phenol degradation in both free and immobilized cells cultures. The best results were observed for an anode potential of 1.0 V/S.H.E. The effect was relatively better pronounced in the case of free cultures: the biodegraded amount of phenol was more than 4 times greater when an electric field was applied at an anode potential of 1.0 V/S.H.E, whereas for immobilized cells, the increase in this amount was 67%. The explanation is that the biodegradation rate by immobilized cells without an electric field application was already more than 4 times higher, compared with the case of the free ones. The combination of immobilization and application of a constant electric field provided synergic results: the degraded amount of phenol was 7.3 times higher than for free culture when no electric field was applied.
- It was observed that the enzyme activities for the first two steps of the phenol catabolic pathway of phenol biodegradation—phenol oxidation (phenol hydroxylase) and benzene ring cleavage (catechol-1,2-dioxygenase and catechol-2,3-dioxygenase)—in free cells were very sensitive to the anode potential. The enzyme activities varied in time and with the anode potential. It was even observed that at an anode potential of 0.8 V/S.H.E., the meta-pathway of cleavage of the benzene ring catalyzed by catechol-2,3-dioxygenase became competitive with the ortho-pathway, catalyzed by catechol-1,2-dioxygenase.
- The increased enzyme activities, together with the calculated biodegraded amounts of phenol under the influence of electric current, compared with the analytical data, showed that the positive effect on the biodegradation of phenol in a constant electric field was due to electrical stimulation of enzyme activity rather than electrochemical anode oxidation. The obtained synergic results for phenol biodegradation may induce researchers to conduct further studies on continuous processes with constant electric field application on immobilized cell cultures.
Supplementary Materials
The following is available online at https://www.mdpi.com/article/10.3390/chemengineering5040075/s1, Figure S1: Enzyme activities at different anode potentials. Initial phenol concentration, 0.06 g dm−3: (A) phenol hydroxylase; (B) catechol 1,2-dioxygenase; (C) catechol 2,3-dioxygenase. Figure S2: 1H-NMR-spectra in water (with presaturation of the water signal): (S2a) 1H NMR spectra of initial phenol solution: (A) an extract from activated carbon particles from a process with no electric field; (B) an alkaline extract after adsorption in the abiotic process; (C) sample from the broth after 140 h of fermentation in an electric field, with anode potential 1 V/S.H.E; (S2b) enlarged spectrum of a sample after 140 h (C).
Author Contributions
Conceptualization, V.B. and Z.A.; methodology, E.V. and M.G.; validation, V.B. and Z.A.; formal analysis, N.P.; investigation, E.V., T.P.-M. and M.G.; data curation, T.P.-M. and M.G.; writing—original draft preparation, V.B.; writing—review and editing, V.B. and Z.A. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Fund for Scientific Research, Republic of Bulgaria, Grant Number DN 17/4/2017.
Data Availability Statement
The presented data will be available at the Fund of Scientific Research, Republic of Bulgaria after the project completion.
Acknowledgments
The NMR analyses were made and interpreted by Svetlana Simova, Institute of Organic Chemistry with Centre of Phytochemistry, Bulgarian Academy of Sciences. The authors express deep gratitude for her valuable help.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
References
- Williams, M. The Merck Index: An Encyclopedia of Chemical, Drugs, and Biologicals, 15th ed.; O’Neil, M.J., Ed.; Royal Society of Chemistry: Cambridge, UK, 2013; ISBN 9781849736701. [Google Scholar] [CrossRef]
- Phenol. Ullmann’s Encyclopedia of Industrial Chemistry; Wiley-VCH: Weinheim, Germany, 2000; ISBN 9783527306732. [Google Scholar] [CrossRef]
- World Health Organization/International Labour Organization: International Chemical Safety Cards. 2017. Available online: http://www.inchem.org/documents/icsc/icsc/eics0070.htm (accessed on 18 August 2021).
- Caetano, M.; Valderrama, C.; Farran, A.; Cortina, J.L. Phenol removal from aqueous solution by adsorption and ion exchange mechanisms onto polymeric resins. J. Colloid Interface Sci. 2009, 338, 402–409. [Google Scholar] [CrossRef] [PubMed]
- Soto, M.L.; Moure, A.; Dominguez, H.; Parajу, J.C. Recovery, concentration and purification of phenolic compounds by adsorption: A review. J. Food Eng. 2011, 105, 1–27. [Google Scholar] [CrossRef]
- Victor-Ortega, M.D.; Ochando-Pulido, J.M.; Martinez-Ferez, A. Modeling of fixed bed column studies for iron removal from industrial effluents through ion exchange process. Chem. Eng. Trans. 2016, 47, 253–258. [Google Scholar] [CrossRef]
- Allsop, P.J.; Chisti, Y.; Moo-Young, M.; Sullivan, G.R. Dynamics of phenol degradation by Pseudomonas putida. Biotechnol. Bioeng. 1993, 41, 572–580. [Google Scholar] [CrossRef] [PubMed]
- Neumegen, R.A.; Fernandez-Alba, A.R.; Chisti, Y. Toxicities of triclosan, phenol, and copper sulfate in activated sludge. Environ. Toxicol. 2005, 20, 160–164. [Google Scholar] [CrossRef]
- Sridevi, V.; Lakshmi, M.V.V.C.; Manasa, M.; Sravani, M. Metabolic pathways for the biodegradation of phenol. Int. J. Eng. Sci. Adv. Technol. 2012, 2, 695–705. [Google Scholar]
- Hinteregger, C.; Leitner, R.; Loidl, M.; Ferschl, A.; Streichsbier, F. Degradation of phenol and phenolic compounds by Pseudomonas putida EKII. Appl. Microbiol. Biotechnol. 1992, 37, 252–259. [Google Scholar] [CrossRef]
- Beschkov, V.; Alexieva, Z.; Parvanova-Mancheva, T.; Vasileva, E.; Gerginova, M.; Peneva, N.; Stoyanova, K. Phenol biodegradation by the strain Pseudomonas putida affected by constant electric field. Int. J. Environ. Sci. Technol. 2020, 17, 1929–1936. [Google Scholar] [CrossRef]
- Lakshmi, M.V.V.C.; Sridevi, V. A review on biodegradation of phenol from industrial effluents. J. Ind. Pollut. Control 2009, 25, 13–27. [Google Scholar]
- Li, Q.; Kang, C.; Zhang, C. Waste water produced from an oilfield and continuous treatment with an oil-degrading bacterium. Process Biochem. 2005, 40, 873–877. [Google Scholar] [CrossRef]
- Dey, S.K.; Mukherjee, A. Catechol oxidase and phenoxazinone synthase: Biomimetic functional models and mechanistic studies. Coord. Chem. Rev. 2016, 310, 80–115. [Google Scholar] [CrossRef]
- Hopper, W.; Mahadevan, A. Degradation of catechin by Bradyrhizobium japonicum. Biodegradation 1997, 8, 159–165. [Google Scholar] [CrossRef]
- Vasileva, E.; Parvanova-Mancheva, T.; Beschkov, V. The Bradyrhizobium japonicum 273 strain’ s ability to degrade phenol. Part 1. J. Int. Sci. Publ. Mater. Methods Technol. 2020, 14, 165–176. Available online: https://www.scientific-publications.net/en/article/1002058/ (accessed on 8 October 2021).
- Jobin, L.; Namour, P. Bioremediation in water environment: Controlled electro-stimulation of organic matter self-purification in aquatic environments. Adv. Microbiol. 2017, 7, 813–852. [Google Scholar] [CrossRef][Green Version]
- Liu, H.; Tong, S.; Chen, N.; Liu, Y.; Feng, C.; Hu, Q. Effect of electro-stimulation on activity of heterotrophic denitrifying bacteria and denitrification performance. Bioresour. Technol. 2015, 196, 123–128. [Google Scholar] [CrossRef]
- Ailijiang, N.; Chang, J.; Wu, Q.; Li, P.; Liang, P.; Zhang, X.X.; Huang, X. Phenol degradation by suspended biomass in aerobic/anaerobic electrochemical reactor. Water Air Soil Pollut. 2016, 227, 233. [Google Scholar] [CrossRef]
- Zhou, L.; Yan, X.; Yan, Y.; Li, T.; An, J.; Liao, C.M.; Li, N.; Wang, X. Electrode potential regulates phenol degradation pathways in oxygen-diffused microbial electrochemical system. Chem. Eng. J. 2020, 381, 122663. [Google Scholar] [CrossRef]
- Nakazawa, T.; Nakazawa, A. [64] Pyrocatechase (pseudomonas). Methods Enzymol. 1970, 17, 518–522. [Google Scholar]
- Baggi, G.; Barbieri, P.; Galli, E.; Tollari, S. Isolation of a Pseudomonas stutzeri strain that degrades o-xylene. Appl. Environ. Microbiol. 1987, 53, 2129–2132. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC204069/ (accessed on 8 October 2021).
- Lazarkevich, I.; Sotirova, T.; Avramova, S.; Stoitsova, S.; Paunova-Krasteva, T.; Galabova, D. Antibacterial activity of methyltiosulfonate and its complexes with rhamnolipid and trehalose lipid against Pseudomonas aeroginosa NBMCC 1390. Res. J. Pharm. Biol. Chem. Sci. 2015, 6, 282–290. [Google Scholar]
- Sotirova, A.; Avramova, T.; Stoitsova, S.; Lazarkevich, I.; Lubenets, V.; Karpenko, E.; Galabova, D. The importance of rhamnolipid-biosurfactant-induced changes in bacterial membrane lipids of Bacillus subtilis for the antimicrobial activity of thiosulfonates, Curr. Microbiol. 2012, 65, 534–541. [Google Scholar] [CrossRef]
- Field, S.J.; Thornton, N.P.; Anderson, L.J.; Gates, A.J.; Reilly, A.; Jepson, B.J.N.; Richardson, D.J.; George, S.J.; Cheesman, M.R.; Butt, J.N. Reductive activation of nitrate reductases. Dalton Trans. 2005, 21, 3580–3586. [Google Scholar] [CrossRef] [PubMed]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).