Quercetin Enhances the Thioredoxin Production of Nasal Epithelial Cells In Vitro and In Vivo


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Mice
2.2. Reagents
2.3. Cell Culture
2.4. Assay to Assess Cytotoxicity of H2O2 and Quercetin
2.5. Assay to Assess TRX mRNA Expression
2.6. Sensitization and Treatment of Mice
2.7. Collection of Nasal Lavage Fluids
2.8. Assessment of Nasal Symptoms
2.9. TRX Assay
2.10. Oxidative Stress Assay
2.11. Statistical Analysis
3. Results
3.1. Influence of H2O2 Stimulation on TRX Production from HNEpCs in Vitro
3.2. In Vitro Influence of Quercetin on H2O2-Induced TRX Production from HNEpCs
3.3. Influence of H2O2 and Quercetin on Cell Viability
3.4. Influence of Quercetin on TRX mRNA Expression
3.5. Influence of Quercetin on Oxidative Stress Responses in Nasal Mucosa
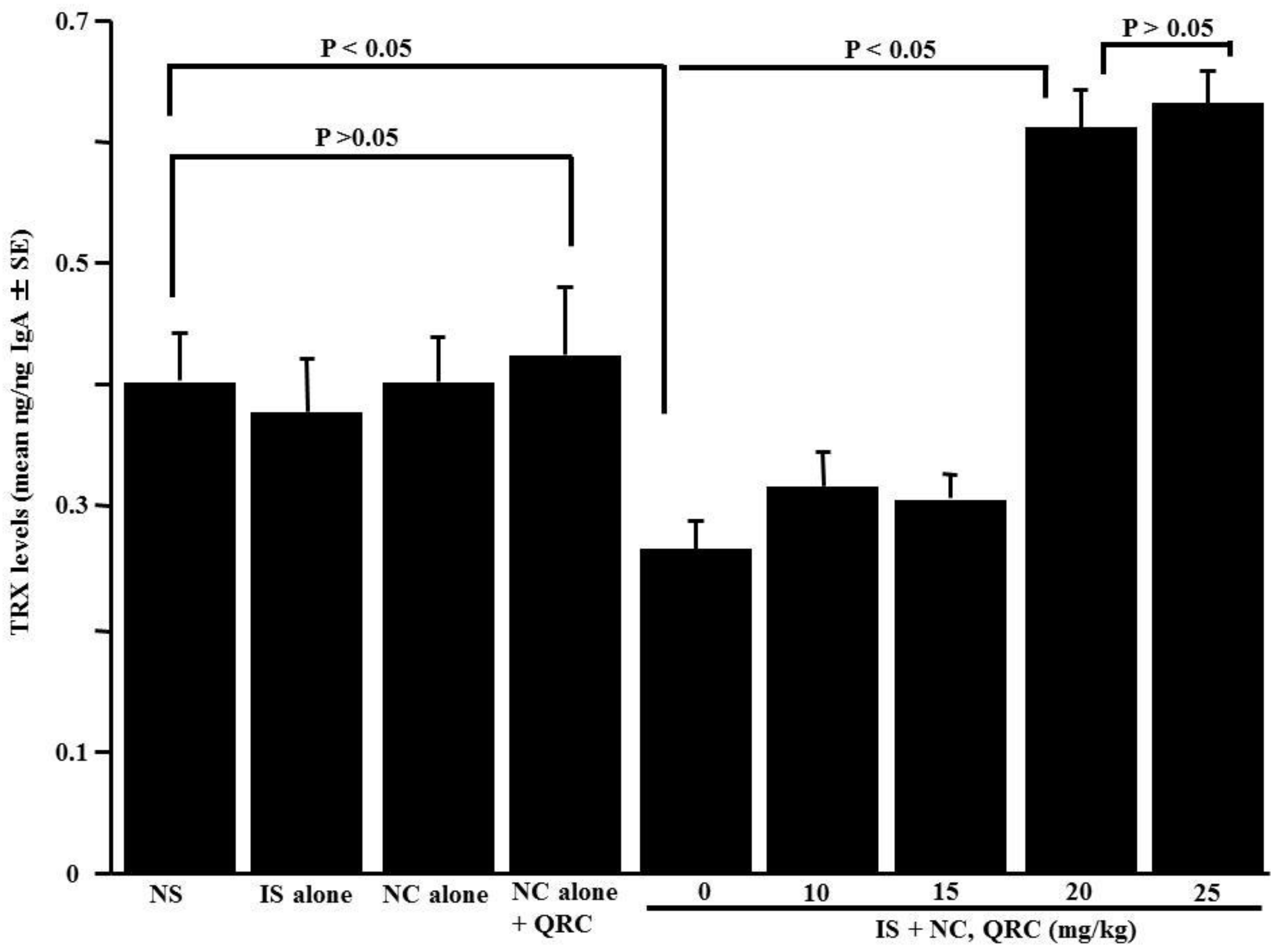
3.6. Influence of Quercetin on the Appearance of TRX in Nasal Lavage Fluids
3.7. Influence of Quercetin on the Development of OVA-Induced Nasal Allergy-Like Symptoms
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Pawankar, R.; Mori, S.; Ozu, C.; Kimura, S. Overview on the pathomechanisms of allergic rhinitis. Asia Pac. Allergy 2011, 1, 157–167. [Google Scholar] [CrossRef] [PubMed]
- Ramirez-Jimenez, F.; Pavon-Romero, G.; Juarez-Martinez, L.L.; Teran, L.M. Allergic rhinitis. J. Allergy Ther. 2012, S5, 006. [Google Scholar] [CrossRef]
- Jung, D.; Lee, S.; Hong, S. Effects of acupuncture and moxibustion in a mouse model of allergic rhinitis. Otolaryngol. Head Neck Surg. 2011, 146, 19–25. [Google Scholar] [CrossRef] [PubMed]
- Jeong, K.T.; Kim, S.G.; Lee, J.; Park, Y.N.; Park, H.H.; Parl, N.Y.; Kim, K.J.; Lee, H.; Lee, Y.J. Anti-allergic effect of a Korean traditional medicine, Biyeom-Tang on mast cells and allergic rhinitis. BMC Comp. Altern. Med. 2014, 14, 54. [Google Scholar] [CrossRef] [PubMed]
- Van Cauwenberg, P.; Bachert, C.; Passalacua, G. Consensus statement on the treatment of allergic rhinitis. European Academy of Allergology and Clinical Immunology. Allergy 2000, 55, 116–134. [Google Scholar] [CrossRef]
- Ishizawa, K.; Yoshizumi, M.; Kawai, Y.; Terao, J.; Kihira, Y.; Ikeda, Y.; Tomita, S.; Minakuchi, K.; Tsuchiya, K.; Tamaki, T. Pharmacology in health food: Metabolism of quercetin in vivo and its protective effect against arteriosclerosis. J. Pharmacol. Sci. 2011, 115, 466–470. [Google Scholar] [CrossRef] [PubMed]
- Hattori, M.; Mizuguchi, H.; Baba, Y.; Ono, S.; Nakano, T.; Zhang, Q.; Sasaki, Y.; Kobayashi, M.; Kitamura, Y.; Takeda, N.; et al. Quercetin inhibits transcriptional up-regulation of histamine H1 receptor via suppressing protein kinase C-δ/extracellular signal-regulated kinase/poly (ADP-ribose) polymerase-1 signaling pathway in Hela cells. Int. Immunopharmacol. 2013, 15, 232–239. [Google Scholar] [CrossRef] [PubMed]
- Kawada, N.; Seki, S.; Inoue, M.; Kuroki, T. Effect of antioxidants, resveratrol, quercetin, and N-acetylcysteine, on the functions of cultured rat hapatic stellate cells and kupper cells. Hepatology 1998, 27, 1265–1274. [Google Scholar] [CrossRef] [PubMed]
- Amorati, R.; Baschieri, A.; Cowde, A.; Valgimigli, L. The antioxidant activity of quercetin in water solution. Biomimetics 2017, 2, 9. [Google Scholar] [CrossRef]
- Lesjak, M.; Beara, I.; Simin, N.; Pintac, D.; Majkic, T.; Bekvalac, K.; Orcic, D.; Mimica-Dukic, N. Antioxidant and anti-inflammatory activities of quercetin and its derivatives. J. Funct. Foods 2018, 40, 68–75. [Google Scholar] [CrossRef]
- Middleton, E., Jr. Effect of plant flavonoids on immune and inflammatory cell function. Adv. Exp. Med. Biol. 1998, 439, 175–182. [Google Scholar] [PubMed]
- Min, Y.D.; Choi, C.H.; Bark, H.; Son, H.Y.; Park, H.H.; Lee, S.; Park, J.W.; Park, E.K.; Shin, H.I.; Kim, S.H. Quercetin inhibits expression of inflammatory cytokines through attenuation of NF-kB and P38 MAPK in HMC-1 human mast cell line. Inflamm. Res. 2007, 56, 210–215. [Google Scholar] [CrossRef] [PubMed]
- Hirano, T.; Kawai, M.; Arimitsu, J.; Ogawa, M.; Kuwahara, Y.; Hagihara, K.; Shima, Y.; Narazaki, M.; Ogata, A.; Koyanagi, M.; et al. Preventive effect of a flavonoid, enzymatically modified isoquercetin on ocular symptoms of Japanese cedar pollinosis. Allergol. Int. 2009, 58, 373–382. [Google Scholar] [CrossRef] [PubMed]
- Rogerio, A.P.; Kanashiro, A.; Fontanari, C.; de Silva, E.V.; Lucisano-Valim, Y.M.; Soares, E.G.; Faccioli, L.H. Anti-inflammatory activity of quercetin and isoquercetin in experimental murine allergic asthma. Inflamm. Res. 2007, 56, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Dorsch, W.; Bittinger, M.; Kaas, A.; Muller, A.; Kreher, B.; Wagner, H. Antiasthmatic effects of Galphimia glauca, gallic acid, and related compounds prevent allergen- and platelet-activating factor-induced bronchial obstruction as well as bronchial hyperreactivity in guinea pigs. Int. Arch. Allergy Immunol. 1992, 97, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Kashiwabara, M.; Asano, K.; Mizuyoshi, T.; Kobayashi, H. Suppression of neuropeptide production by quercetin in allergic rhinitis model rats. BMC Comp. Altern. Med. 2016, 16, 132. [Google Scholar] [CrossRef] [PubMed]
- Min, A.; Lee, Y.A.; Kim, K.; EI-Benna, J.; Shin, M.H. Nox2-derived ROS-mediated surface translocation of BLT1 is essential for exocytosis in human eosinophils induced by LBT4. Int. Arch. Allergy Immunol. 2014, 165, 40–51. [Google Scholar] [CrossRef] [PubMed]
- Suojalehto, H.; Vehmas, T.; Lindstrom, I.; Kennedy, D.W.; Kilpelainen, M.; Plosila, T.; Savukoski, S.; Sipilä, J.; Varpula, M.; Wolff, H.; et al. Nasal nitric oxide is dependent on sinus obstruction in allergic rhinitis. Laryngoscope 2014, 124, E231–E238. [Google Scholar] [CrossRef] [PubMed]
- Sen, S.; Chakraborty, R.; Sridhar, C.; Reddy, Y.S.R.; De, B. Free radicals, antioxidants, diseases and phytomedicines: Current status and future prospect. Int. J. Pharm. Sci. Rev. Res. 2010, 3, 91–100. [Google Scholar]
- Burke-Gaffney, A.; Callister, M.E.J.; Nakamura, H. Thioredoxin: Friend or foe in human disease. Trends Pharmacol. Sci. 2005, 26, 398–404. [Google Scholar] [CrossRef] [PubMed]
- Holmgren, A.; Lu, J. Thioredoxin and thioredoxin reductase: Current research with special reference to human disease. Biochem. Biophys. Res. Commun. 2010, 396, 120–124. [Google Scholar] [CrossRef] [PubMed]
- Ichiki, H.; Hoshino, T.; Kinoshita, T.; Imaoka, H.; Kato, S.; Inoue, H.; Nakamura, H.; Yodoi, J.; Young, H.A.; Aizawa, H. Thioredoxin suppresses airway hyperresponsiveness and airway inflammation in asthma. Biochem. Biophys. Res. Commun. 2005, 334, 1141–1148. [Google Scholar] [CrossRef] [PubMed]
- Imaoka, H.; Hoshino, T.; Takei, S.; Sakazaki, Y.; Kinoshita, T.; Okamoto, M.; Kawayama, T.; Yodoi, J.; Kato, S.; Iwanaga, T.; et al. Effects of thioredoxin on established airway remodeling in a chronic antigen exposure asthma model. Biochem. Biophys. Res. Commun. 2007, 360, 525–530. [Google Scholar] [CrossRef] [PubMed]
- Suzaki, I.; Asano, K.; Kanei, A.; Suzaki, H. Enhancement of thioredoxin production from nasal epithelial cells by the macrolide antibiotic, Clarithromycin in vitro. Vivo 2013, 27, 351–356. [Google Scholar]
- Hollman, P.C.; vd Gaag, M.; Mengelers, M.J.; van Trijp, J.M.; de Vries, J.H.; Katan, M.B. Absorption and disposition kinetics of the dietary antioxidant quercetin in man. Free Radic. Biol. Med. 1996, 21, 703–707. [Google Scholar] [CrossRef]
- Wadsworth, T.L.; Koop, D.R. Effects of Ginkgo biloba extract (EGb 761) and quercetin on lipopolysaccharide-induced release of nitric oxide. Biochem. Pharmacol. 2001, 137, 43–58. [Google Scholar] [CrossRef]
- Imoto, Y.; Yamada, T.; Tsukahara, H.; Kimura, Y.; Kato, Y.; Sakashita, M.; Fujieda, S. Nitrite/nitrate in nasal lavage fluid reflect nasal symptoms after a single nasal allergen provocation in patients with seasonal allergic rhinitis. J. Investig. Allergol. Clin. Immunol. 2015, 25, 382–384. [Google Scholar] [PubMed]
- Sannohe, S.; Adachi, T.; Hamada, K.; Honda, K.; Yamada, Y.; Saito, N.; Cui, C.H.; Kayaba, H.; Ishikawa, K.; Chihara, J. Upregulated response to chemokines in oxidative metabolism of eosinophils in asthma and allergic rhinitis. Eur. Respir. J. 2003, 21, 925–931. [Google Scholar] [CrossRef] [PubMed]
- Sakai-Kashiwabara, M.; Abe, S.; Asano, K. Suppressive activity of quercetin on the production of eosinophil chemoattractants from eosinophils in vitro. Vivo 2014, 28, 515–522. [Google Scholar]
- Irie, S.; Kashiwabara, M.; Yamada, A.; Asano, K. Suppressive activity of quercetin on periostin functions in vitro. Vivo 2016, 30, 17–25. [Google Scholar]
- Lopez, M.S.; Nieto, A. Glucocorticoids induce the expression of the uteroglobin gene in rabbit fetal lung explants cultured in vitro. Biochem. J. 1985, 225, 255–258. [Google Scholar] [CrossRef]
- Fernandez-Renau, D.; Lombardero, M.; Nieto, A. Glucocorticoid-dependent uteroglobin synthesis and uteroglobin mRNA levels in rabbit lung explants cultured in vitro. Eur. J. Biochem. 1984, 144, 523–527. [Google Scholar] [CrossRef] [PubMed]
- Price, A.; Ramachandran, S.; Ramachandran, S.; Smith, G.P.; Stevenson, M.L.; Pomeranz, M.K.; Cohen, D.E. Oral allergy syndrome (Pollen-food allergy syndrome). Dermatitis 2015, 26, 78–88. [Google Scholar] [CrossRef] [PubMed]
- Lessof, M.H. Pollen-food allergy syndrome. J. Allergy Clin. Immunol. 1996, 98, 239–240. [Google Scholar] [CrossRef]







© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Edo, Y.; Otaki, A.; Asano, K. Quercetin Enhances the Thioredoxin Production of Nasal Epithelial Cells In Vitro and In Vivo. Medicines 2018, 5, 124. https://doi.org/10.3390/medicines5040124
Edo Y, Otaki A, Asano K. Quercetin Enhances the Thioredoxin Production of Nasal Epithelial Cells In Vitro and In Vivo. Medicines. 2018; 5(4):124. https://doi.org/10.3390/medicines5040124
Chicago/Turabian StyleEdo, Yukako, Amane Otaki, and Kazuhito Asano. 2018. "Quercetin Enhances the Thioredoxin Production of Nasal Epithelial Cells In Vitro and In Vivo" Medicines 5, no. 4: 124. https://doi.org/10.3390/medicines5040124
APA StyleEdo, Y., Otaki, A., & Asano, K. (2018). Quercetin Enhances the Thioredoxin Production of Nasal Epithelial Cells In Vitro and In Vivo. Medicines, 5(4), 124. https://doi.org/10.3390/medicines5040124