
Degradation of 2,4,6-Trinitrotoluene (TNT): Involvement of Protocatechuate 3,4-Dioxygenase (P34O) in Buttiauxella sp. S19-1



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Selection of Bacterial Strains
2.2. Bacterial Culture and Cometabolic Degradation of TNT
2.3. High-Performance Liquid Chromatography (HPLC) Detection of TNT
2.4. Prokaryotic Transcriptome Analysis of Buttiauxella sp. S19-1
2.5. Cloning Vectors, Kits and Other Reagents
2.6. Obtaining the BuP34O Wildtype Gene and Generating a BuP34O Frame Shift Sequence by Polymerase Chain Reactions (PCR)
2.7. Cloning of the BuP34O Wild-Type Sequence
2.8. Subcloning of the BuP34O Wild-Type Gene and BuP34O Knockout Sequence
2.9. Construction of the BuP34O Knockout Strain (S-M1 Mutant)
2.10. Construction of BuP34O-Expressing Escherichia coli Mutant Strain (EM1)
2.11. Exposure of Wild-Type and Mutant Strains to TNT
2.12. Purification and Enzyme Activity Analysis of Recombinant P34O (rP34O)
2.12.1. Purification of rP34O
2.12.2. Enzyme Activity and Kinetic Analysis of rP34O
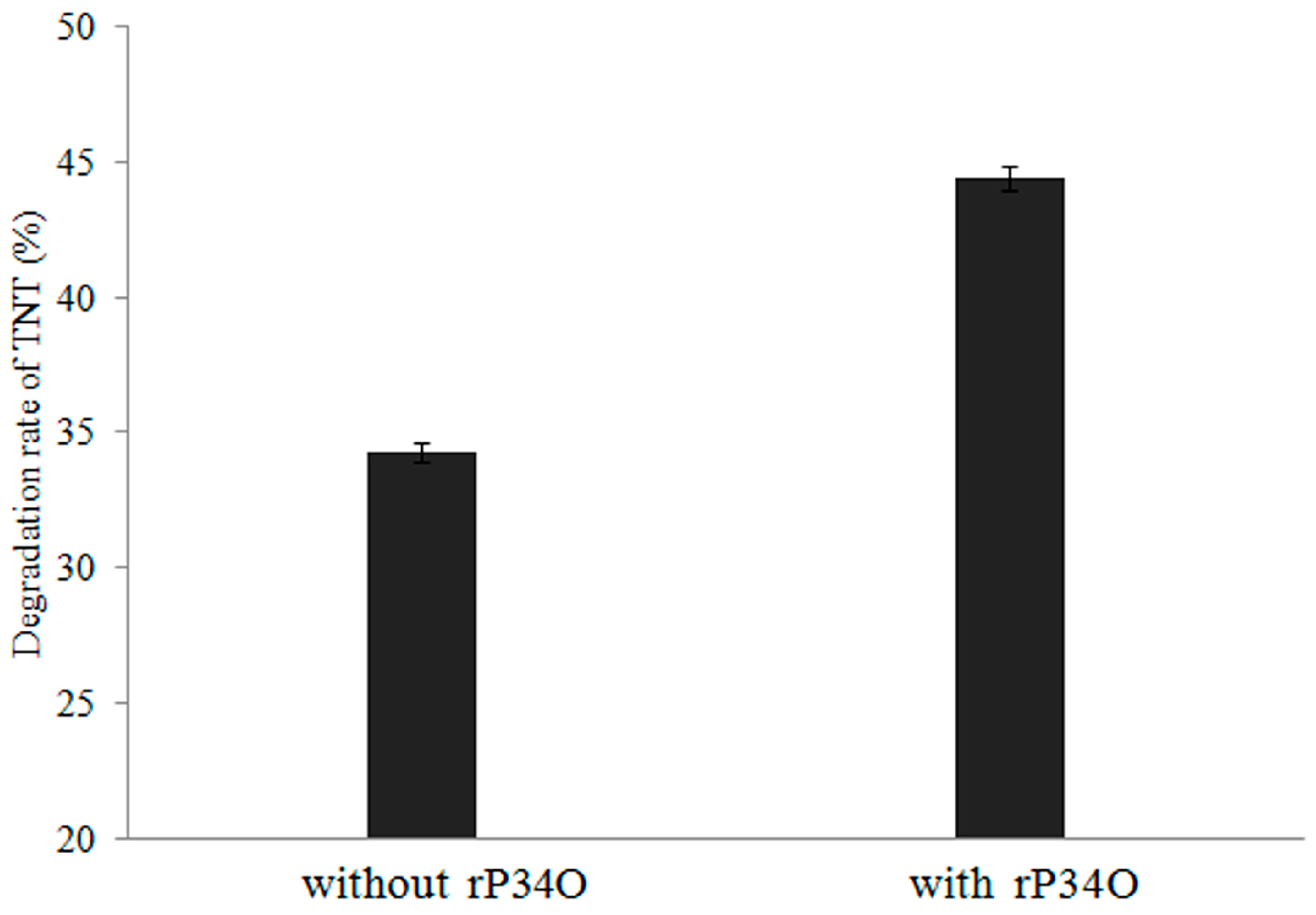
2.13. Investigating the Effect of rP34O on TNT and ADNT Degradation
2.14. GC-MS Analysis of TNT and Its Metabolite ADNT
2.15. Statistics
3. Results and Discussion
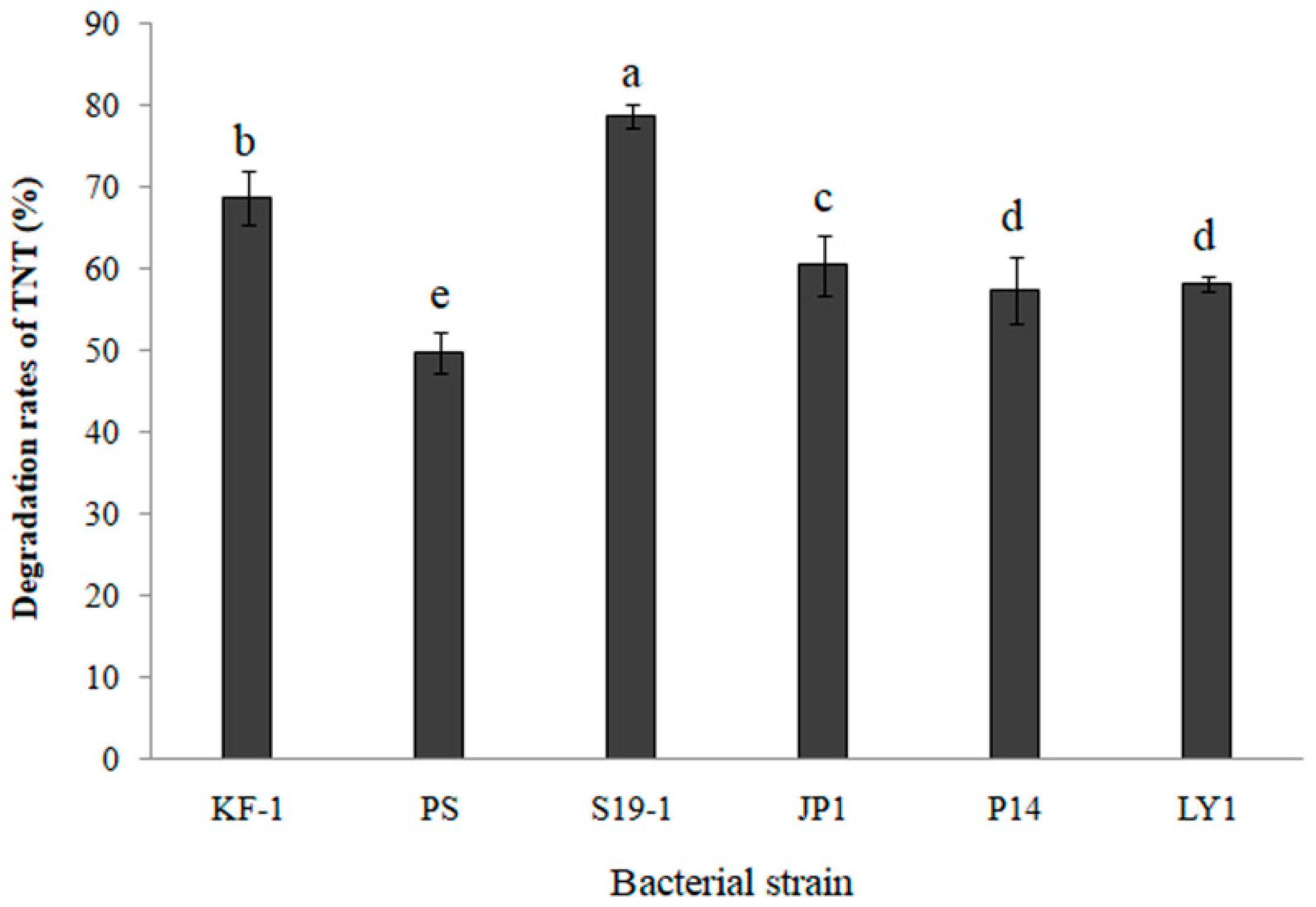
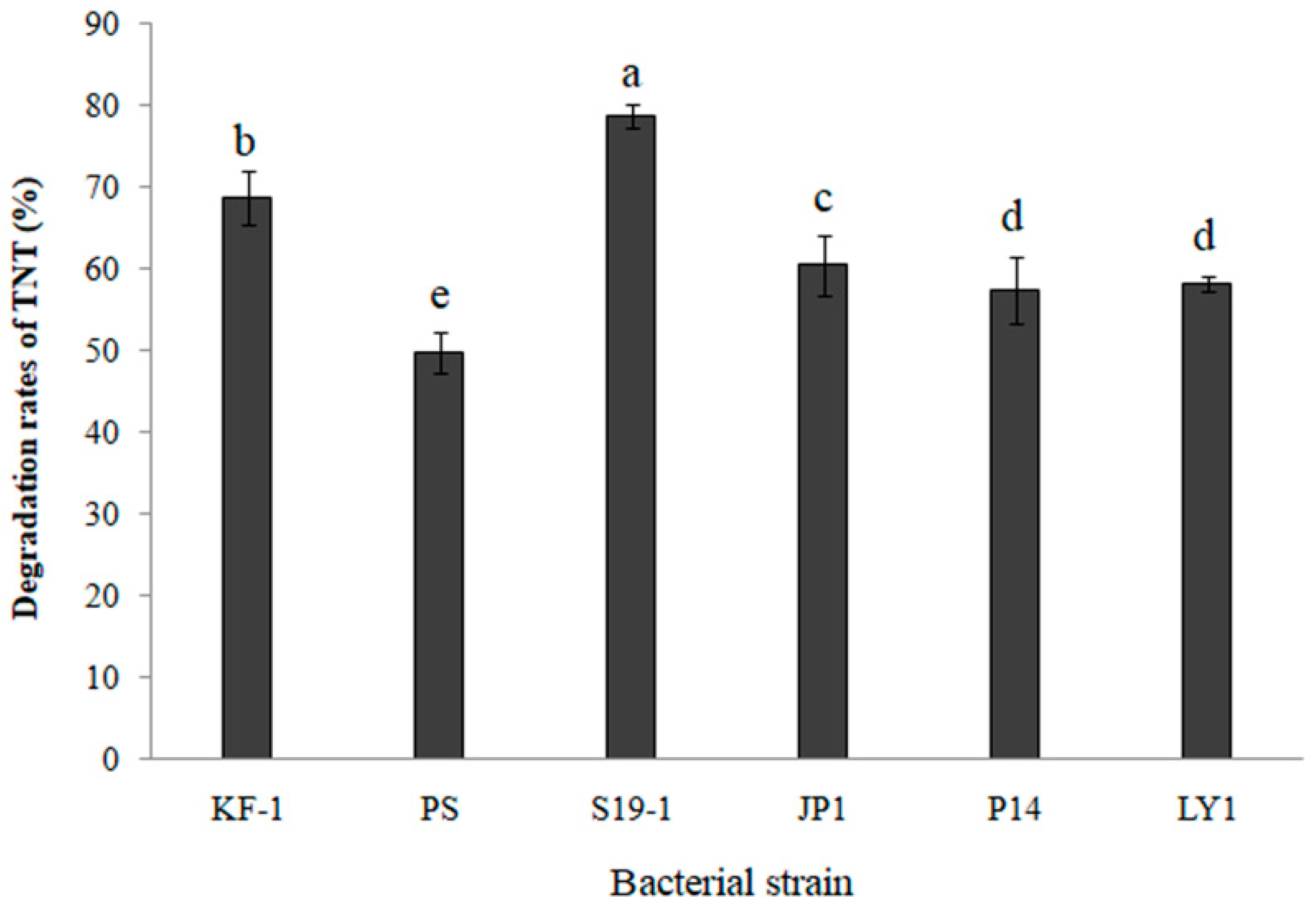
3.1. Comparing Cometabolic Degradation of TNT in Selected Bacteria
3.2. Prokaryotic Transcriptomics of Buttiauxella sp. S19-1
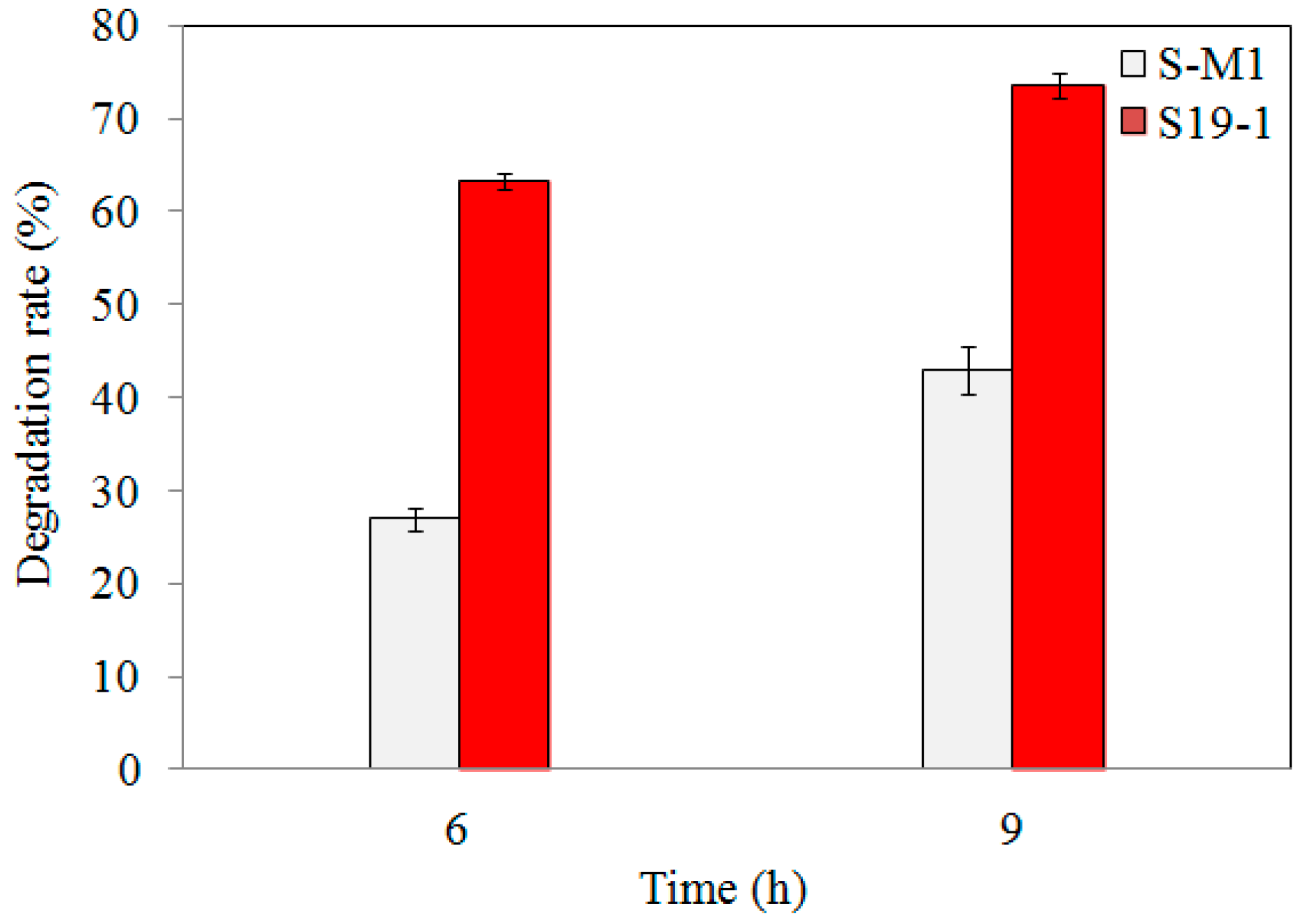
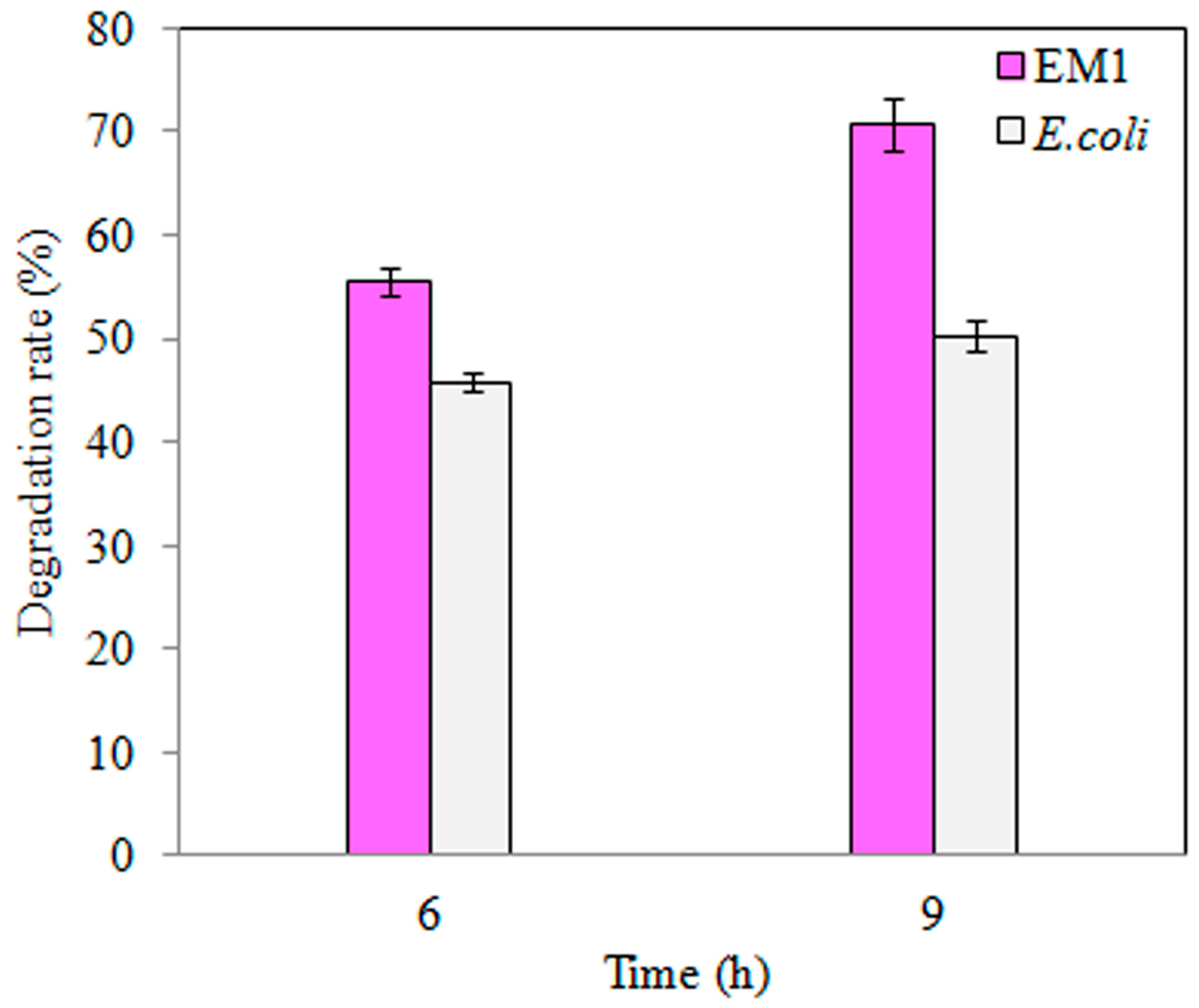
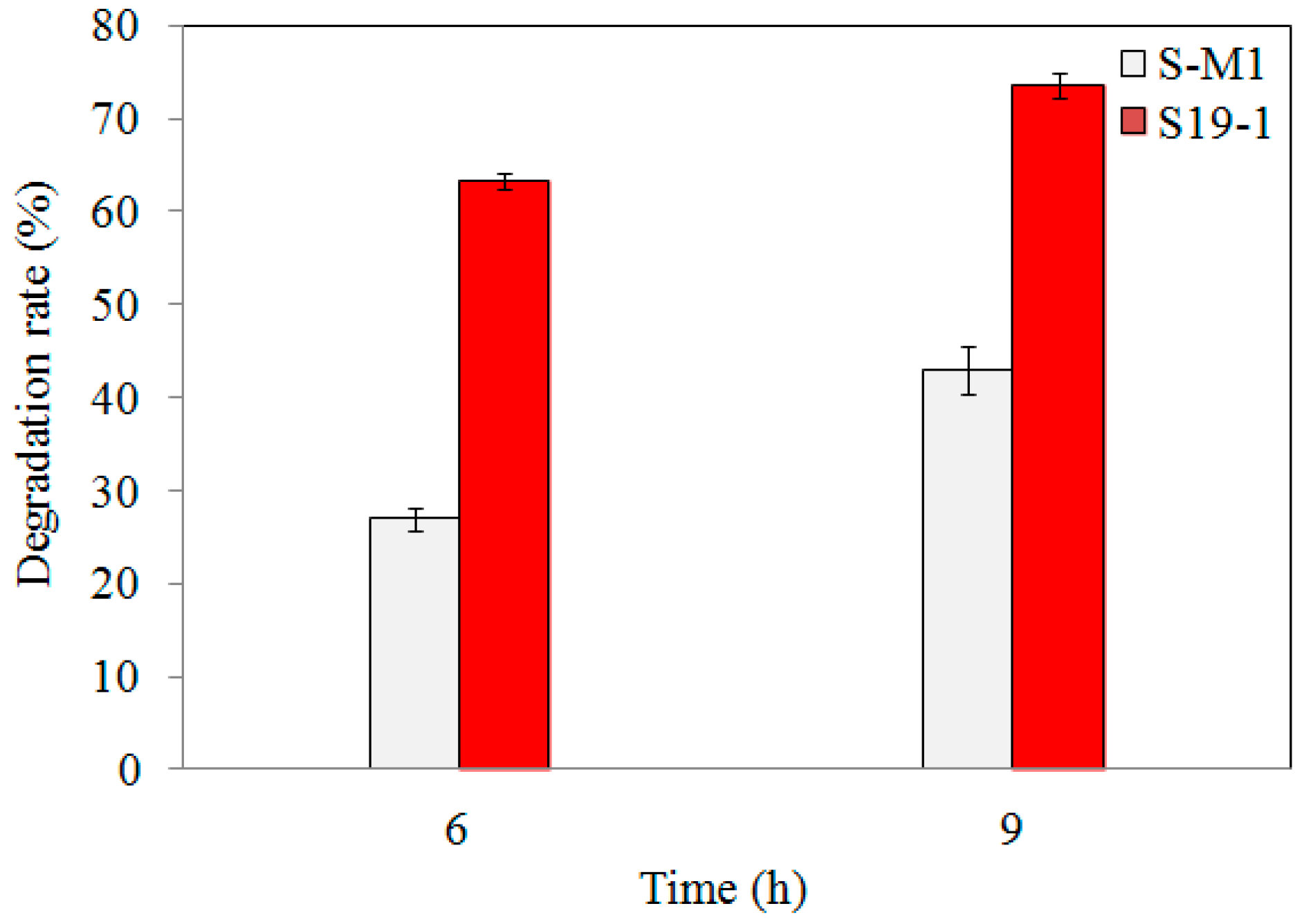
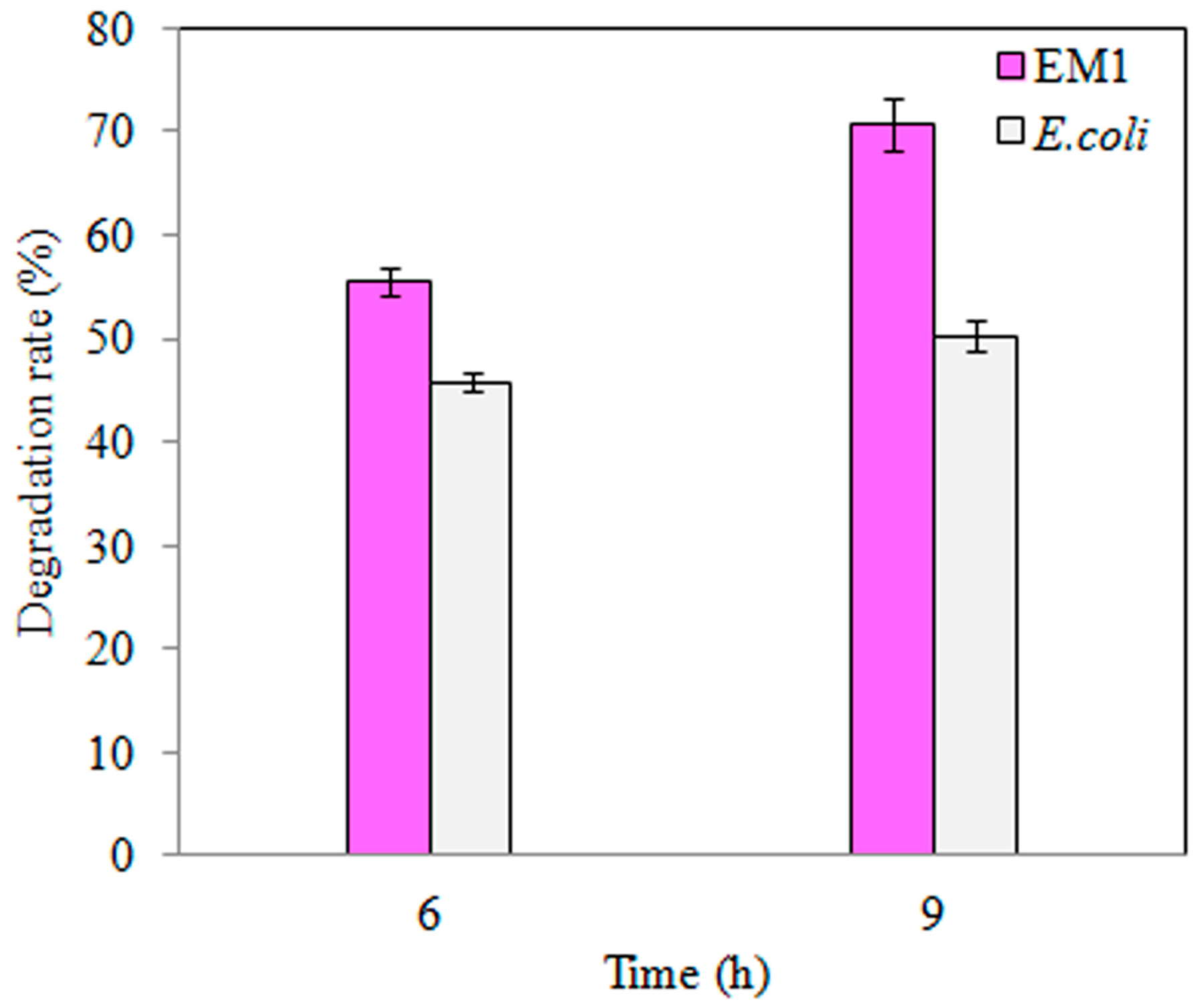
3.3. TNT Degradation by S-M1 and EM1 Mutant Strains
3.4. Expression, Identification, Enzyme Activity, and Kinetic Analysis of rP34O
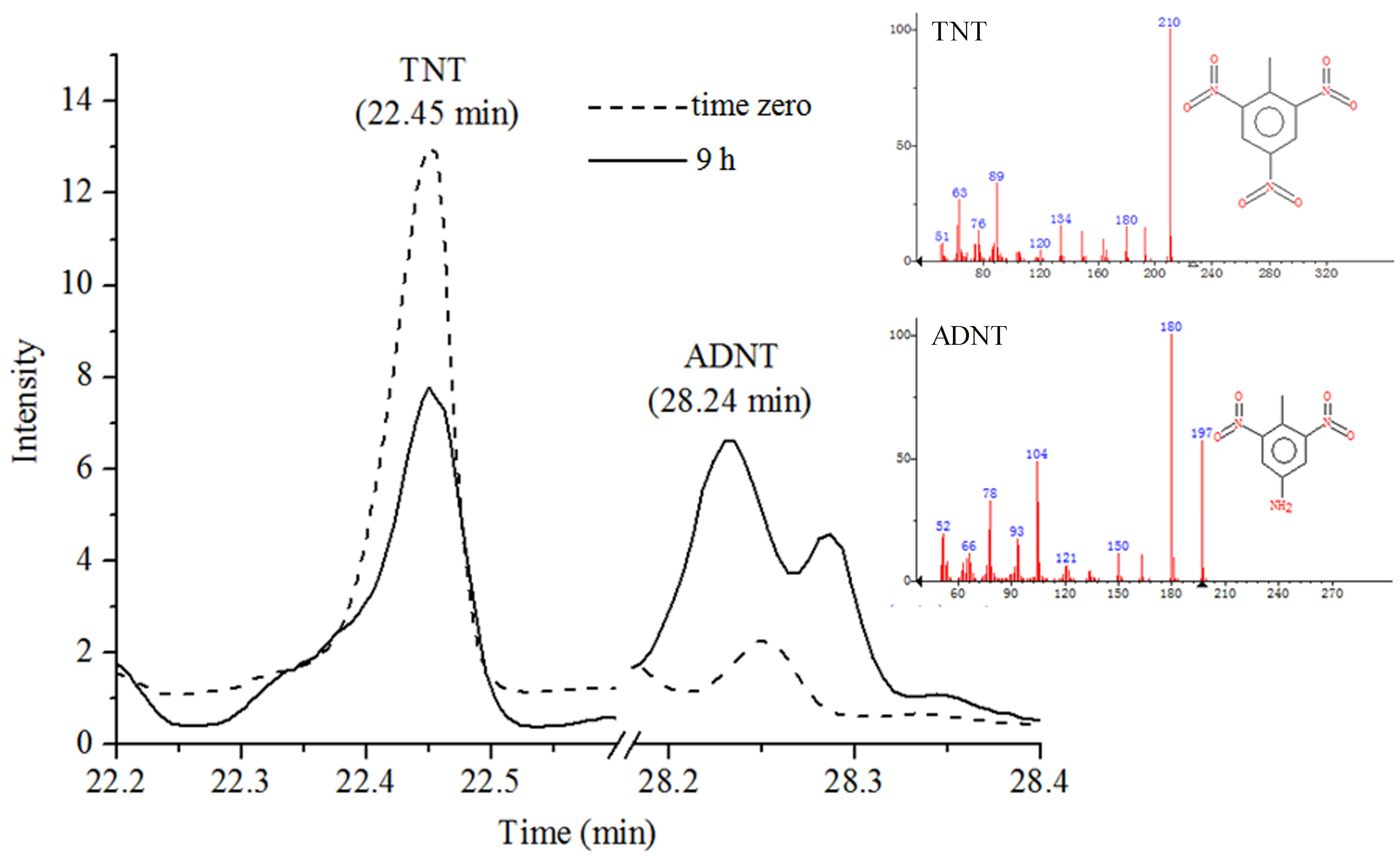
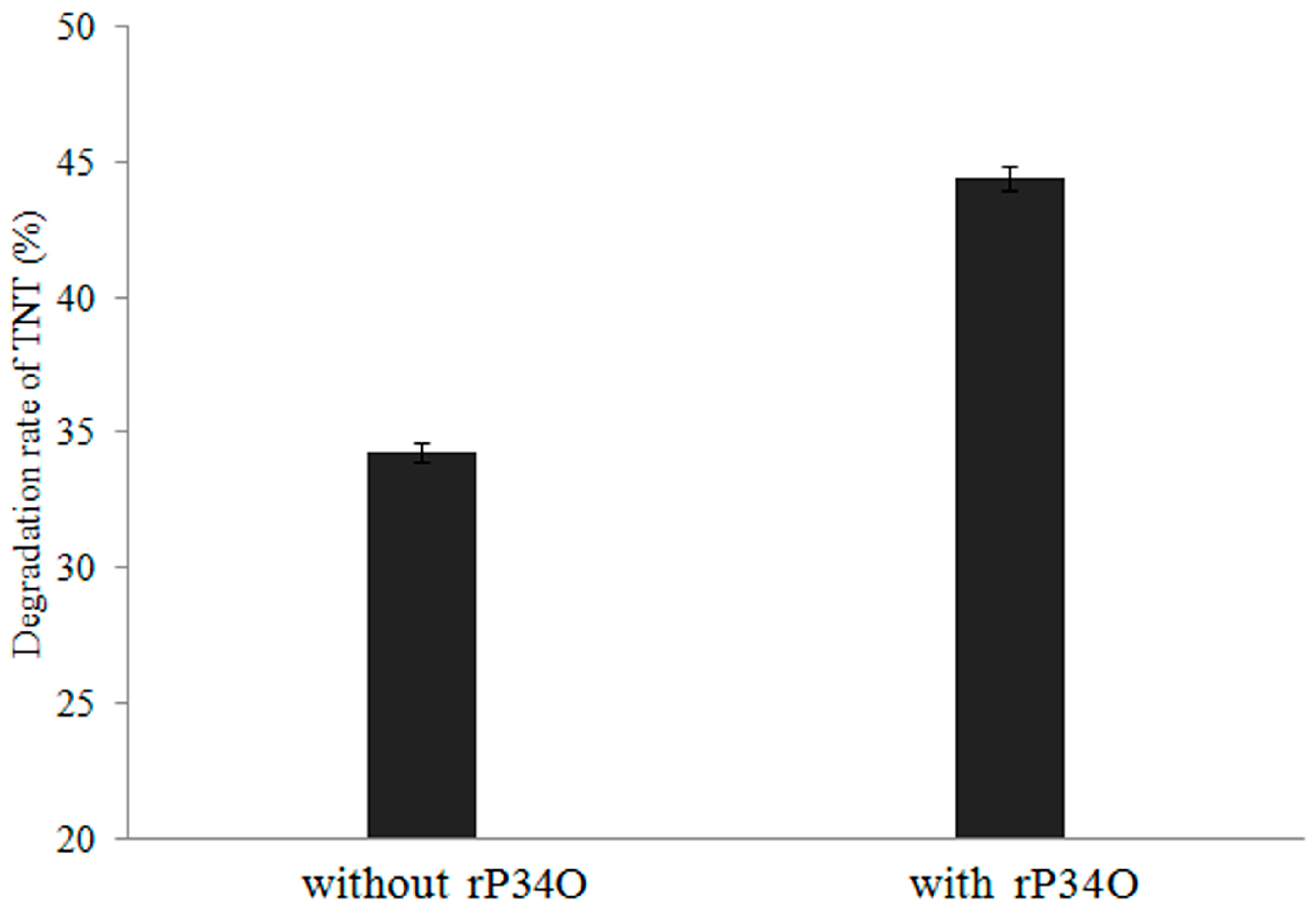
3.5. GC-MS Analysis of TNT Degradation by rP34O and Buttiauxella sp. S19-1
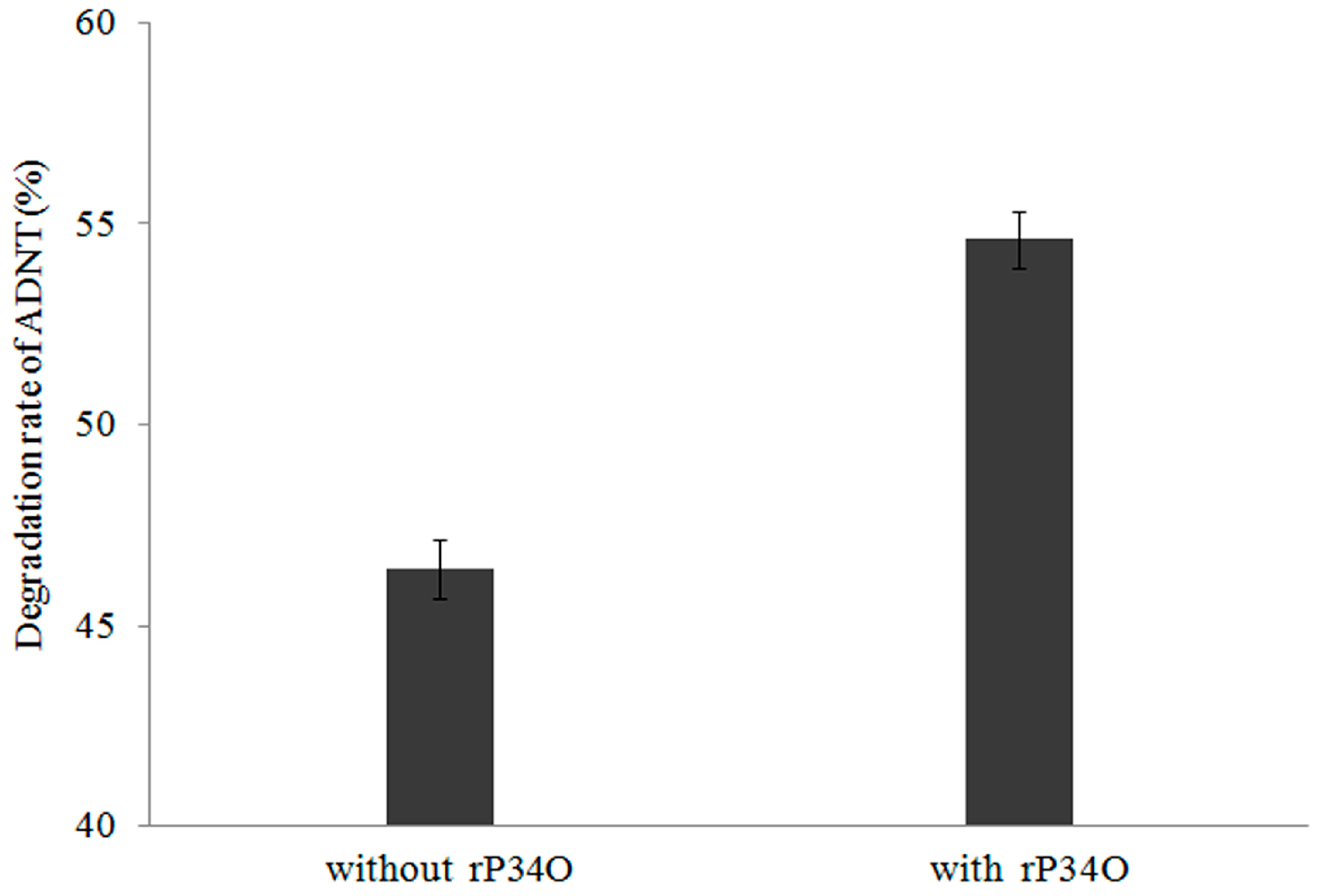
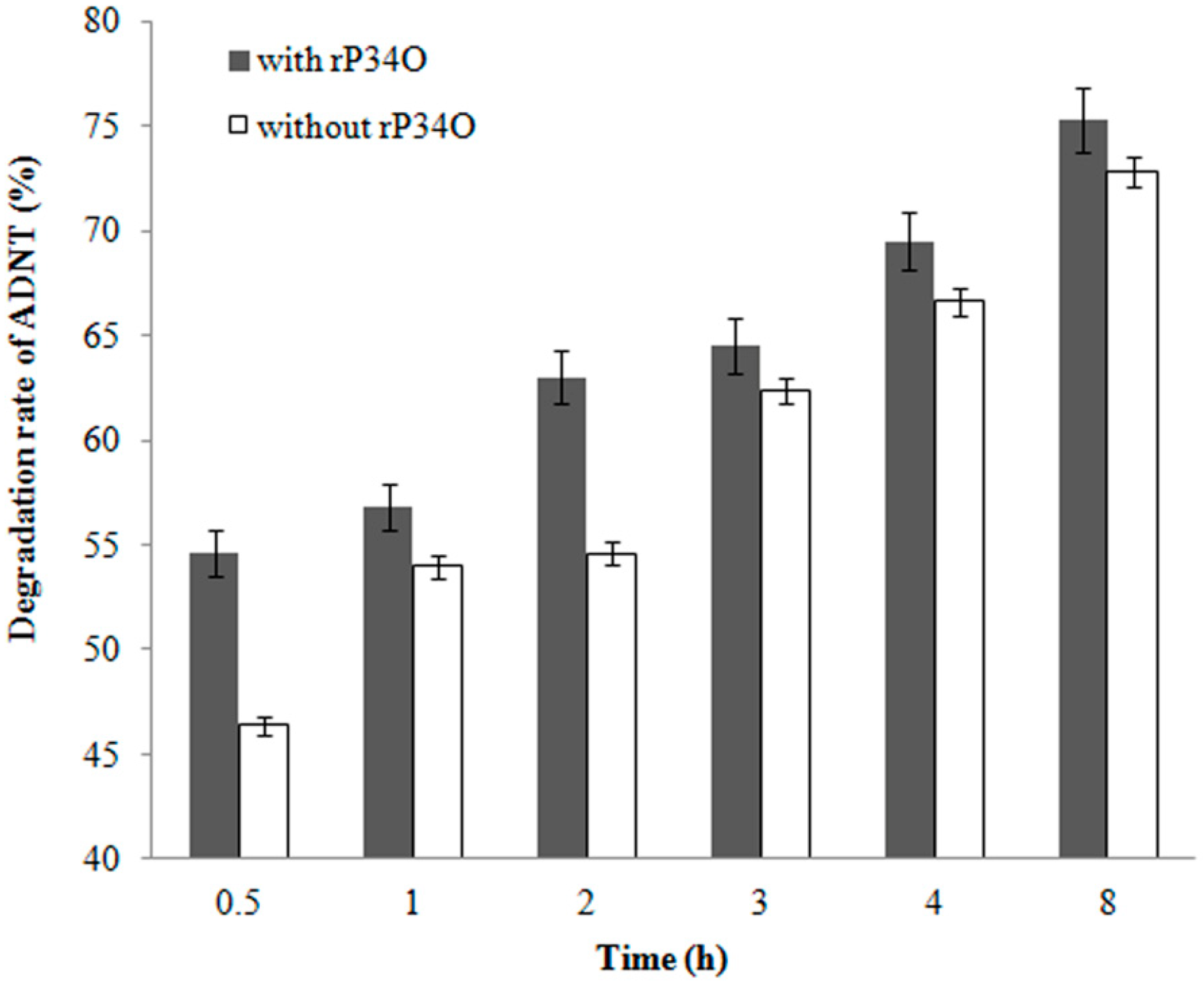
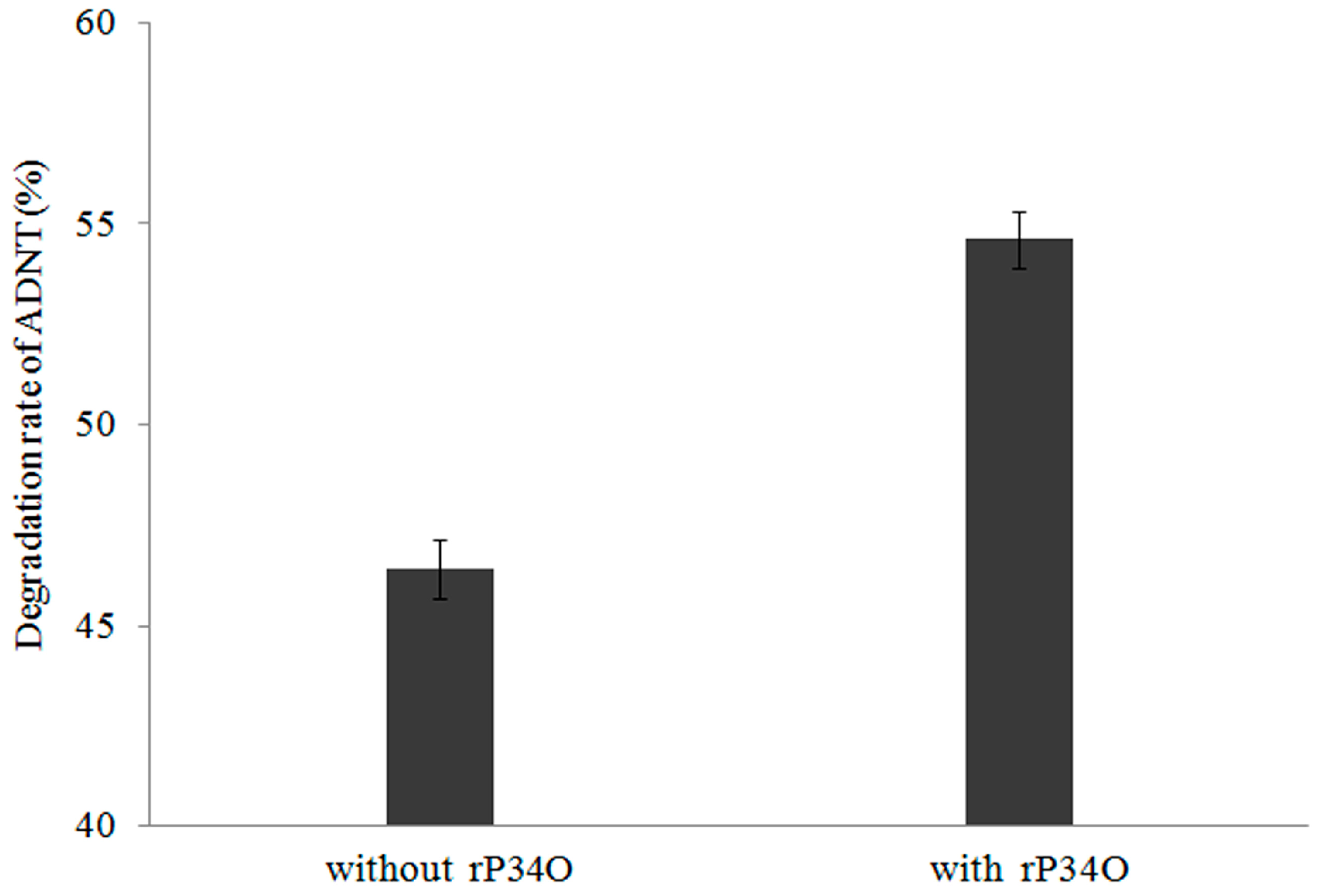
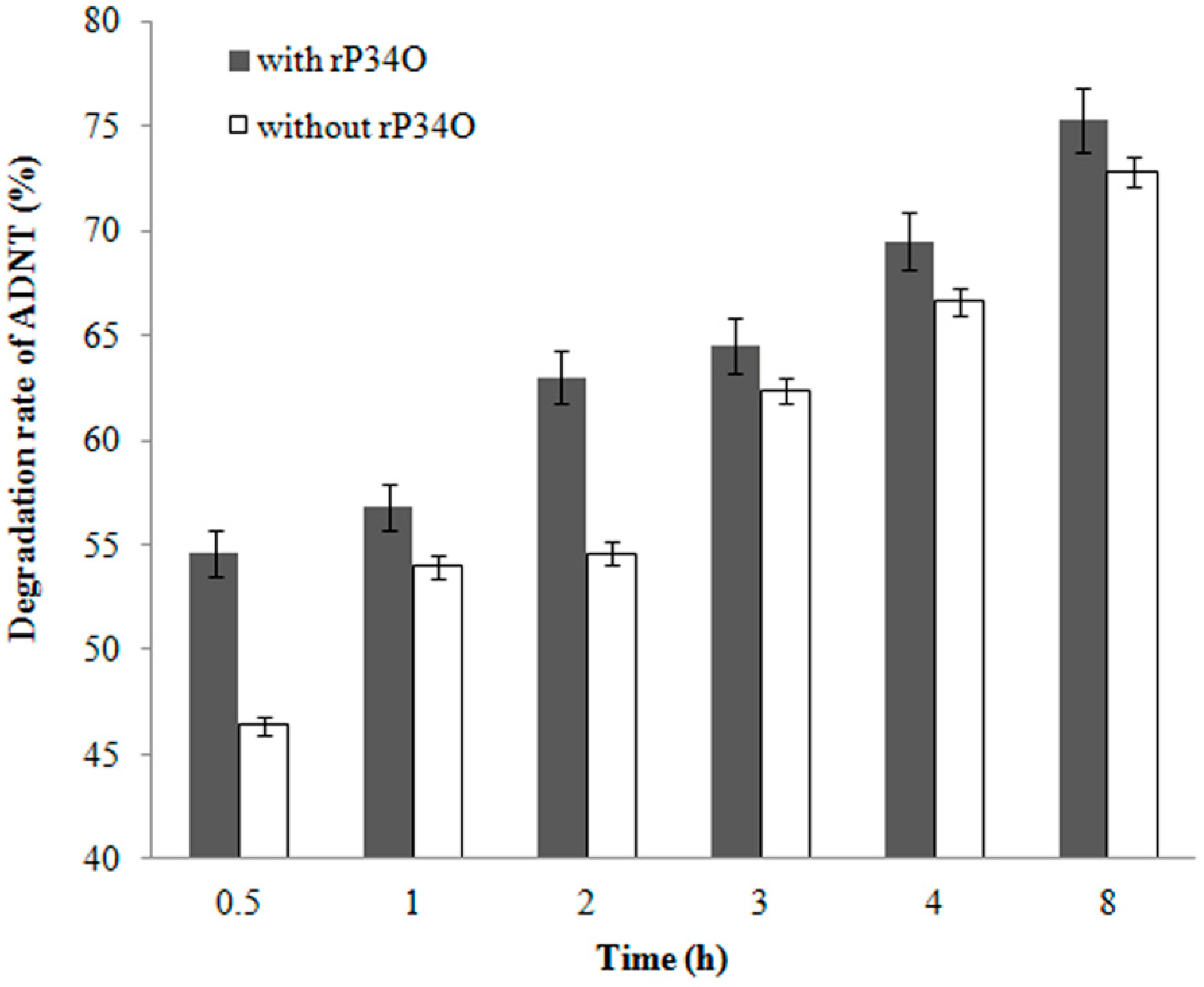
3.6. GC-MS Analysis of ADNT Degradation by Buttiauxella sp. S19-1 and rP34O
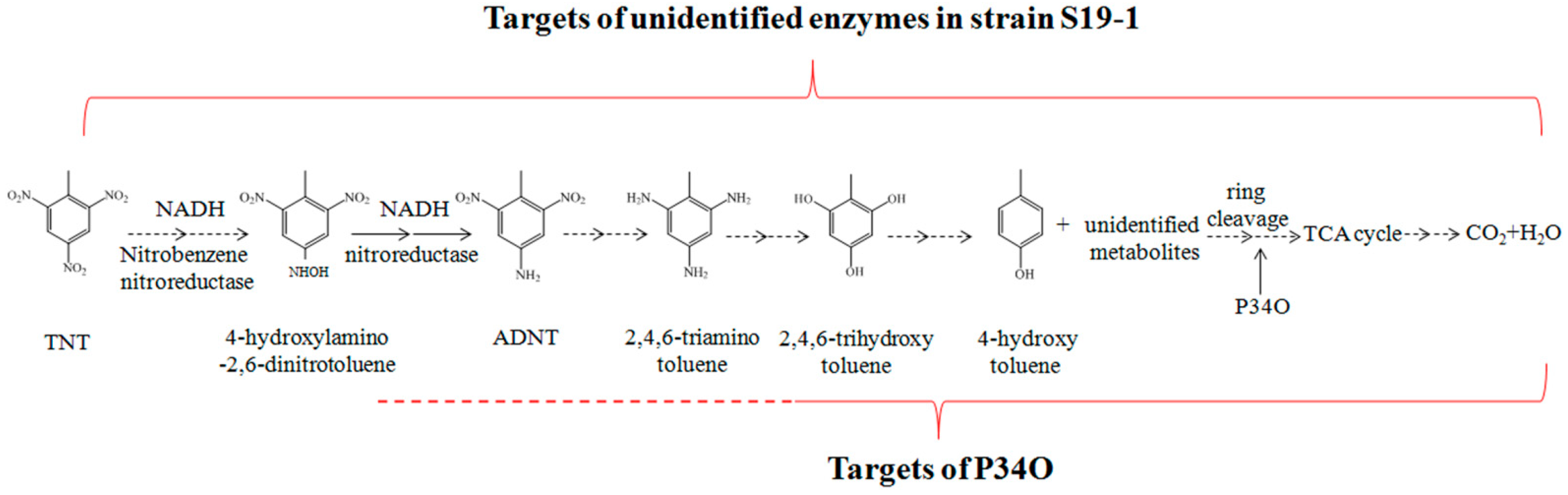
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Esteve-Núnez, A.; Caballero, A.; Ramos, J.L. Biological degradation of 2,4,6-trinitrotoluene. Microbiol. Mol. Biol. R. 2001, 65, 335–352. [Google Scholar] [CrossRef] [Green Version]
- Stenuit, B.A.; Agathos, S.N. Biodegradation and bioremediation of TNT and other nitro explosives. Compr. Biotechnol. 2011, 6, 167–181. [Google Scholar]
- Boelsterli, U.A.; Ho, H.K.; Zhou, S.; Leow, K.Y. Bioactivation and hepatotoxicity of nitroaromatic drugs. Curr. Drug Metab. 2006, 7, 715–727. [Google Scholar] [CrossRef] [PubMed]
- Whitacre, D.M. Reviews of Environmental Contamination and Toxicology, 1st ed.; Springer: New York, NY, USA, 2012. [Google Scholar]
- Alexander, M. Biological degradation and bioremediation of toxic chemicals. J. Environ. Qual. 1996, 25, 204–205. [Google Scholar] [CrossRef]
- Lewis, T.A.; Newcombe, D.A.; Crawford, R.L. Bioremediation of soils contaminated with explosives. J. Environ. Manag. 2004, 70, 291–307. [Google Scholar] [CrossRef] [PubMed]
- Meagher, R.B. Plants tackle explosive contamination. Nat. Biotechnol. 2006, 24, 161–163. [Google Scholar] [CrossRef]
- Ayoub, K.; Hullebusch, E.D.V.; Cassir, M.; Bermond, A. Application of advanced oxidation processes for TNT removal: A review. J. Hazard. Mater. 2010, 178, 10–28. [Google Scholar] [CrossRef]
- Miura, G. Plant toxicology: Defusing the explosive. Nat. Chem. Biol. 2015, 11, 829. [Google Scholar] [CrossRef]
- Gómez-Gardeñes, J.; Lotero, L.; Taraskin, S.N.; Pérez-Reche, F.J. Explosive contagion in networks. Sci. Rep. 2016, 6, 19767. [Google Scholar] [CrossRef] [Green Version]
- Bolt, H.M.; Degen, G.H.; Dorn, S.B.; Plöttner, S.; Harth, V. Genotoxicity and potential carcinogenicity of 2,4,6-trinitrotoluene: Structural and toxicological considerations. Rev. Environ. Health 2006, 21, 217–228. [Google Scholar] [CrossRef]
- Koske, D.; Goldenstein, N.I.; Kammann, U. Nitroaromatic compounds damage the DNA of zebrafish embryos (Danio rerio). Aquat. Toxicol. 2019, 217, 105345. [Google Scholar] [CrossRef] [PubMed]
- Travis, E.R.; Bruce, N.C.; Rosser, S.J. Microbial and plant ecology of a long-term TNT-contaminated site. Environ. Pollut. 2008, 153, 119–126. [Google Scholar] [CrossRef] [PubMed]
- Travis, E.R.; Bruce, N.C.; Rosser, S.J. Short term exposure to elevated trinitrotoluene concentrations induced structural and functional changes in the soil bacterial community. Environ. Pollut. 2008, 153, 432–439. [Google Scholar] [CrossRef]
- Serrano-González, M.Y.; Chandra, R.; Castillo-Zacarias, C.; Robledo-Padilla, F.; Rostro-Alanis, M.D.J.; Parra-Saldivar, R. Biotransformation and degradation of 2,4,6-trinitrotoluene by microbial metabolism and their interaction. Def. Technol. 2018, 14, 151–164. [Google Scholar] [CrossRef]
- Chauhan, A.; Jain, R.K. Biodegradation: Gaining insight through proteomics. Biodegradation 2010, 21, 861–879. [Google Scholar] [CrossRef] [PubMed]
- Aken, B.V.; Hofrichter, M.; Scheibner, K.; Hatakka, A.I.; Naveau, H.; Agathos, S.N. Transformation and mineralization of 2,4,6-trinitrotoluene (TNT) by manganese peroxidase from the white-rot basidiomycete Phlebia radiata. Biodegradation 1999, 10, 83–91. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Lee, S.Y.; Shin, K.S. Biodegradation of 2,4,6-trinitrotoluene by white-rot fungus Irpex lacteus. Mycobiology 2009, 37, 17–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ziganshin, A.M.; Naumova, R.P.; Pannier, A.J.; Gerlach, R. Influence of pH on 2,4,6-trinitrotoluene degradation by Yarrowia lipolytica. Chemosphere 2010, 79, 426–433. [Google Scholar] [CrossRef] [Green Version]
- Khilyas, I.V.; Lochnit, G.; Ilinskaya, O.N. Proteomic analysis of 2,4,6-trinitrotoluene degrading yeast Yarrowia lipolytica. Front. Microbiol. 2017, 8, 2600. [Google Scholar] [CrossRef]
- Aken, B.V.; Yoon, J.M.; Schnoor, J.L. Biodegradation of nitro-substituted explosives 2,4,6-trinitrotoluene, hexahydro-1,3,5-trinitro-1,3,5-triazine, and octahydro-1,3,5,7-tetranitro-1,3,5-tetrazocine by a phytosymbiotic Methylobacterium sp. associated with poplar tissues (Populus deltoides x nigra DN34). Appl. Environ. Microbiol. 2004, 70, 508–517. [Google Scholar]
- Hoehamer, C.F.; Wolfe, N.L.; Eriksson, K.E.L. Biotransformation of 2, 4, 6-trinitrotoluene (TNT) by the Fungus Fusarium oxysporum. Int. J. Phytoremed. 2006, 8, 95–105. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, S.; Deb, U.; Datta, S.; Walther, C.; Gupta, D.K. Common explosives (TNT, RDX, HMX) and their fate in the environment: Emphasizing bioremediation. Chemosphere 2017, 184, 438–451. [Google Scholar] [CrossRef] [PubMed]
- Kalafut, T.; Wales, M.E.; Rastogi, V.K.; Naumova, R.P.; Zaripova, S.K.; Wild, J.R. Biotransformation patterns of 2,4,6-trinitrotoluene by aerobic bacteria. Curr. Microbiol. 1998, 36, 45–54. [Google Scholar] [CrossRef]
- Vanderberg, L.A.; Perry, J.J.; Unkefer, P.J. Catabolism of 2,4,6-trinitrotoluene by Mycobacterium vaccae. Appl. Microbiol. Biotechnol. 1995, 43, 937–945. [Google Scholar] [CrossRef]
- Vorbeck, C.; Lenke, H.; Fischer, P.; Spain, J.C.; Knackmuss, H.J. Initial reductive reactions in aerobic microbial metabolism of 2,4,6-trinitrotoluene. Appl. Environ. Microbiol. 1998, 64, 246–252. [Google Scholar] [CrossRef] [Green Version]
- Vorbeck, C.; Lenke, H.; Fischer, P.; Knackmuss, H.J. Identification of a hydride-Meisenheimer complex as a metabolite of 2,4,6-trinitrotoluene by a Mycobacterium strain. J. Bacteriol. 1994, 176, 932–934. [Google Scholar] [CrossRef] [Green Version]
- Martin, J.L.; Comfort, S.D.; Shea, P.J.; Drijber, R.A.; Kokjohn, T.A. Denitration of 2,4,6-trinitrotoluene by Pseudomonas savastanoi. Can. J. Microbiol. 1997, 43, 447–455. [Google Scholar] [CrossRef]
- Rieger, P.G.; Sinnwell, V.; Preuß, A.; Francke, W.; Knackmuss, H.J. Hydride-Meisenheimer complex formation and protonation as key reactions of 2, 4, 6-trinitrophenol biodegradation by Rhodococcus erythropolis. J. Bacteriol. 1999, 181, 1189–1195. [Google Scholar] [CrossRef] [Green Version]
- Stenuit, B.A.; Agathos, S.N. Microbial 2,4,6-trinitrotoluene degradation: Could we learn from (bio) chemistry for bioremediation and vice versa? Appl. Microbiol. Biotechnol. 2010, 88, 1043–1064. [Google Scholar] [CrossRef]
- Claus, H. Microbial degradation of 2,4,6-trinitrotoluene in vitro and in natural environments. In Biological Remediation of Explosive Residues. Environmental Science and Engineering; Singh, S., Ed.; Springer: New York, NY, USA, 2014; pp. 15–38. [Google Scholar]
- Giedraityte, G.; Kalėdienė, L. Catechol 1, 2-dioxygenase from α-naphthol degrading thermophilic Geobacillus sp. strain: Purification and properties. Cent. Eur. J. Biol. 2009, 4, 68–73. [Google Scholar] [CrossRef] [Green Version]
- Wells, J.T.; Ragauskas, A.J. Biotechnological opportunities with the β-ketoadipate pathway. Trends Biotechnol. 2012, 30, 627–637. [Google Scholar] [CrossRef]
- Harwood, C.S.; Parales, R.E. The β-ketoadipate pathway and the biology of self-identity. Ann. Rev. Microbiol. 1996, 50, 553–590. [Google Scholar] [CrossRef]
- Valley, M.P.; Brown, C.K.; Burk, D.L.; Vetting, M.W.; Ohlendorf, D.H.; Lipscomb, J.D. Roles of the equatorial tyrosyl iron ligand of protocatechuate 3,4-dioxygenase in catalysis. Biochemistry 2005, 44, 11024–11039. [Google Scholar] [CrossRef] [PubMed]
- Mayilmurugan, R.; Sankaralingam, M.; Suresh, E.; Palaniandavar, M. Novel square pyramidal iron (III) complexes of linear tetradentate bis (phenolate) ligands as structural and reactive models for intradiol-cleaving 3,4-PCD enzymes: Quinine formation vs. intradiol cleavage. Dalton Trans. 2010, 39, 9611–9625. [Google Scholar] [CrossRef] [PubMed]
- Chandra, R.; Modestra, J.A.; Mohan, S.V. Biophotovoltaic cell to harness bioelectricity from acidogenic wastewater associated with microbial community profiling. Fuel 2015, 160, 502–512. [Google Scholar] [CrossRef]
- Wang, P.P.; Zheng, D.N.; Liang, R.B. Isolation and characterization of an estrogen-degrading Pseudomonas putida strain SJTE-1. 3 Biotech 2019, 9, 61. [Google Scholar] [CrossRef] [PubMed]
- Dutta, K.; Shitiokov, S.; Khalifa, I.; Mal, A.; Moulik, S.P.; Panda, A.K.; Ghosh, C. Effects of secondary carbon supplement on biofilm-mediated biodegradation of naphthalene by mutated naphthalene 1,2-dioxygenase encoded by Pseudomonas putida strain KD9. J. Hazard. Mater. 2018, 357, 187–197. [Google Scholar] [CrossRef] [PubMed]
- Schleheck, D.; Knepper, T.P.; Fischer, K.; Cook, A.M. Mineralization of individual congeners of linear alkylbenzenesulfonate by defined pairs of heterotrophic bacteria. Appl. Environ. Microbiol. 2004, 70, 4053–4063. [Google Scholar] [CrossRef] [Green Version]
- Zhang, T.; Xiong, G.; Maser, E. Characterization of the steroid degrading bacterium S19-1 from the Baltic Sea at Kiel, Germany. Chem. Biol. Interact. 2011, 191, 83–88. [Google Scholar] [CrossRef]
- Xu, M.; Han, S.N.; Lu, N.N.; Zhang, X.; Liu, J.M.; Liu, D.; Xiong, G.; Guo, L. Degradation of oestrogen and an oestrogen-like compound in chicken faeces by bacteria. Water Air Soil Pollut. 2018, 229, 337–348. [Google Scholar] [CrossRef]
- Luo, A.; Wu, Y.; Xu, Y.; Kan, J.; Qiao, J.; Liang, L.; Huang, T.; Hu, Z. Characterization of a cytochrome P450 monooxygenase capable of high molecular weight PAHs oxidization from Rhodococcus sp. P14. Process. Biochem. 2016, 51, 2127–2133. [Google Scholar] [CrossRef]
- Liang, L.; Song, X.; Kong, J.; Shen, C.; Huang, T.; Hu, Z. Anaerobic biodegradation of high-molecular-weight polycyclic aromatic hydrocarbons by a facultative anaero be Pseudomonas sp. JP1. Biodegradation 2014, 25, 825–833. [Google Scholar] [CrossRef] [PubMed]
- Li, M.T.; Zhao, X.M.; Zhang, X.F.; Wu, D.; Leng, S. Biodegradation of 17β-estradiol by bacterial co-culture isolated from manure. Sci. Rep. 2018, 8, 3787. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Howard, D.H. The preservation of bacteria by freezing in Glycerol broth. J. Bacteriol. 1956, 71, 625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sambrook, J.; Fritsch, E.F.; Maniatis, T. Molecular Cloning: A Laboratory Manual, 2nd ed.; Cold Spring Harbor Laboratory: New York, NY, USA, 1989. [Google Scholar]
- Bradford, M.M. A rapid and sensitive method of the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–258. [Google Scholar] [CrossRef]
- Buchan, A.; Collier, L.S.; Neidle, E.L.; Moran, M.A. Key aromatic-ring-cleaving enzyme, protocatechuate 3,4-dioxygenase, in the ecologically important marine Roseobacter lineage. Appl. Environ. Microbiol. 2000, 66, 4662–4672. [Google Scholar] [CrossRef] [Green Version]
- Senavirathne, G.; Lopez, M.A.; Messer, R.; Fishel, R.; Yoder, K.E. Expression and purification of nuclease-free protocatechuate 3,4-dioxygenase for prolonged single-molecule fluorescence imaging. Anal. Biochem. 2018, 556, 78–84. [Google Scholar] [CrossRef] [PubMed]
- Messer, R.K.; Lopez, M.A.; Senavirathne, G.; Yoder, K.E. Expression and purification of nuclease-free oxygen scavenger protocatechuate 3,4-dioxygenase. J. Vis. Exp. 2019, 153, e59599. [Google Scholar] [CrossRef]
- Hou, C.T.; Lillard, M.O.; Schwartz, R.D. Protocatechuate 3,4-dioxygenase from acinetobacter calcoaceticus. Biochemistry 1976, 15, 582–588. [Google Scholar] [CrossRef]
- Zhang, L.S.; Fang, Y.; Zhou, Y.; Ye, B.C. Improvement of the stabilization and activity of protocatechuate 3,4-dioxygenase isolated from Rhizobium sp. LMB-1 and immobilized on Fe3O4 nanoparticles. Appl. Biochem. Biotechnol. 2017, 183, 1035–1048. [Google Scholar] [CrossRef]
- Spain, J.C. Biodegradation of nitroaromatic compounds. Annu. Rev. Microbiol. 1995, 49, 523–555. [Google Scholar] [CrossRef]
- Oh, B.T.; Shea, P.J.; Drijber, R.A.; Vasilyeva, G.K.; Sarath, G. TNT biotransformation and detoxification by a Pseudomonas aeruginosa strain. Biodegradation 2003, 14, 309–319. [Google Scholar] [CrossRef] [PubMed]
- Fujisawa, H.; Hayaishi, O. Protocatechuate 3,4-dioxygenase: I. crystallization and characterization. J. Biol. Chem. 1968, 243, 2673–2681. [Google Scholar] [CrossRef]
- Hammer, A.; Stolz, A.; Knackmuss, H.J. Purification and characterization of a novel type of protocatechuate 3,4-dioxygenase with the ability to oxidize 4-sulfocatechol. Arch. Microbiol. 1996, 166, 92–100. [Google Scholar] [CrossRef] [PubMed]
- Guzik, U.; Hupert-Kocurek, K.; Krysiak, M.; Wojcieszyńska, D. Degradation potential of protocatechuate 3,4-dioxygenase from crude extract of Stenotrophomonas maltophilia strain KB2 immobilized in calcium alginate hydrogels and on glyoxyl agarose. Biomed. Res. Int. 2014, 2014, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guzik, U.; Hupert-Kocurek, K.; Sitnik, M.; Wojcieszyńska, D. Protocatechuate 3,4-dioxygenase: A wide substrate specificity enzyme isolated from Stenotrophomonas maltophilia KB2 as a useful tool in aromatic acid biodegradation. J. Mol. Microbiol. Biotechnol. 2014, 24, 150–160. [Google Scholar] [CrossRef]
- Maser, E.; Strehse, J.S. “Don’t Blast”: Blast-in-place (BiP) operations of dumped World War munitions in the oceans significantly increase hazards to the environment and the human seafood consumer. Arch. Toxicol. 2020, 94, 1941–1953. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Xiong, G.; Maser, E. Analysis and characterization of eight estradiol inducible genes and a strong promoter from the steroid degrading marine bacterial strain S19-1. Chem. Biol. Interact. 2013, 202, 159–167. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, M.; Liu, D.; Sun, P.; Li, Y.; Wu, M.; Liu, W.; Maser, E.; Xiong, G.; Guo, L. Degradation of 2,4,6-Trinitrotoluene (TNT): Involvement of Protocatechuate 3,4-Dioxygenase (P34O) in Buttiauxella sp. S19-1. Toxics 2021, 9, 231. https://doi.org/10.3390/toxics9100231
Xu M, Liu D, Sun P, Li Y, Wu M, Liu W, Maser E, Xiong G, Guo L. Degradation of 2,4,6-Trinitrotoluene (TNT): Involvement of Protocatechuate 3,4-Dioxygenase (P34O) in Buttiauxella sp. S19-1. Toxics. 2021; 9(10):231. https://doi.org/10.3390/toxics9100231
Chicago/Turabian StyleXu, Miao, Dong Liu, Ping Sun, Yunuo Li, Ming Wu, Wencong Liu, Edmund Maser, Guangming Xiong, and Liquan Guo. 2021. "Degradation of 2,4,6-Trinitrotoluene (TNT): Involvement of Protocatechuate 3,4-Dioxygenase (P34O) in Buttiauxella sp. S19-1" Toxics 9, no. 10: 231. https://doi.org/10.3390/toxics9100231
APA StyleXu, M., Liu, D., Sun, P., Li, Y., Wu, M., Liu, W., Maser, E., Xiong, G., & Guo, L. (2021). Degradation of 2,4,6-Trinitrotoluene (TNT): Involvement of Protocatechuate 3,4-Dioxygenase (P34O) in Buttiauxella sp. S19-1. Toxics, 9(10), 231. https://doi.org/10.3390/toxics9100231