
Zinc Ameliorate Oxidative Stress and Hormonal Disturbance Induced by Methomyl, Abamectin, and Their Mixture in Male Rats

Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Reagents (Diagnostic Kits)
2.3. Animals
2.4. Determination of Oral LD50 for the Tested Insecticides
2.5. Dosing and Treatments
2.6. Blood and Organs Collection
2.7. Biochemical Measurements
2.8. Statistics
2.9. Joint Action Analysis
- (i)
- Case of positive effect (i.e., a significant increase of the concerned biochemical parameters above the control values due to the effect of the individual compounds); where I.I. > 1 means potentiation; I.I. = 1 means additive; I.I. < 1 means antagonism.
- (ii)
- Case of negative effect (i.e., a significant decrease of the concerned biochemical parameters below the control values due to the effect of the individual compounds); where I.I. > 1 means antagonism; I.I. = 1 means additive; I.I. < 1 means potentiation.
- (iii)
- Case of no observed effect: In such a case, it is supposed that treatment with the mixture and each of its individual compounds do not induce statistically significant differences between the values of the measured parameters; and thus control and treatments have nearly the same values. Here, the interaction index (I.I.), if determined, will equal 1 (i.e., a result similar to that of an additive effect). The prior statistical examination for the dataset of a given biochemical measurement would assist the differentiation between anadditive case and that of no effect.
2.10. Oxidative Stress and Amelioration Analysis
3. Results
3.1. Acute Oral Toxicity of the Tested Insecticides
3.2. Observations on Signs of Toxicity
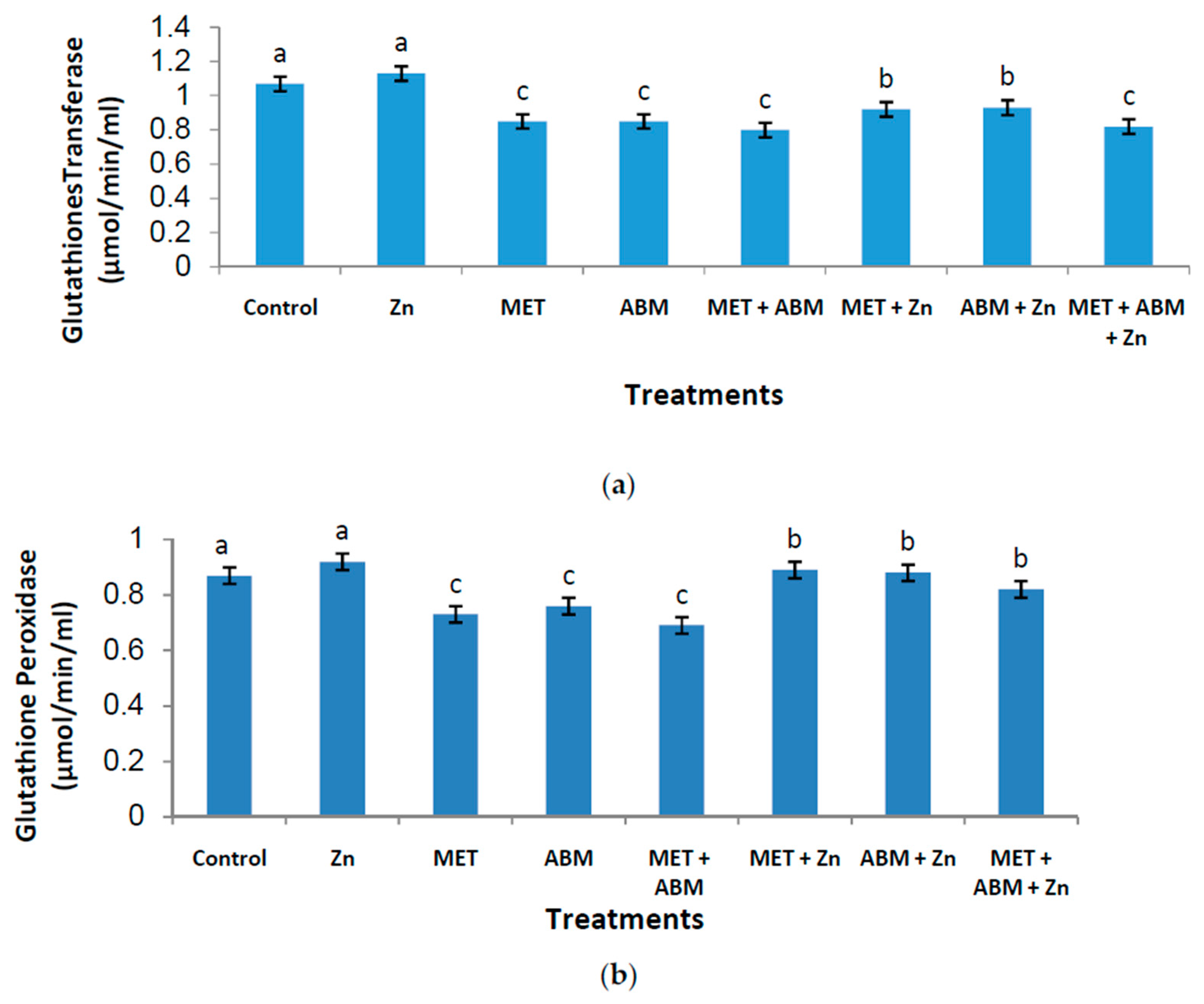
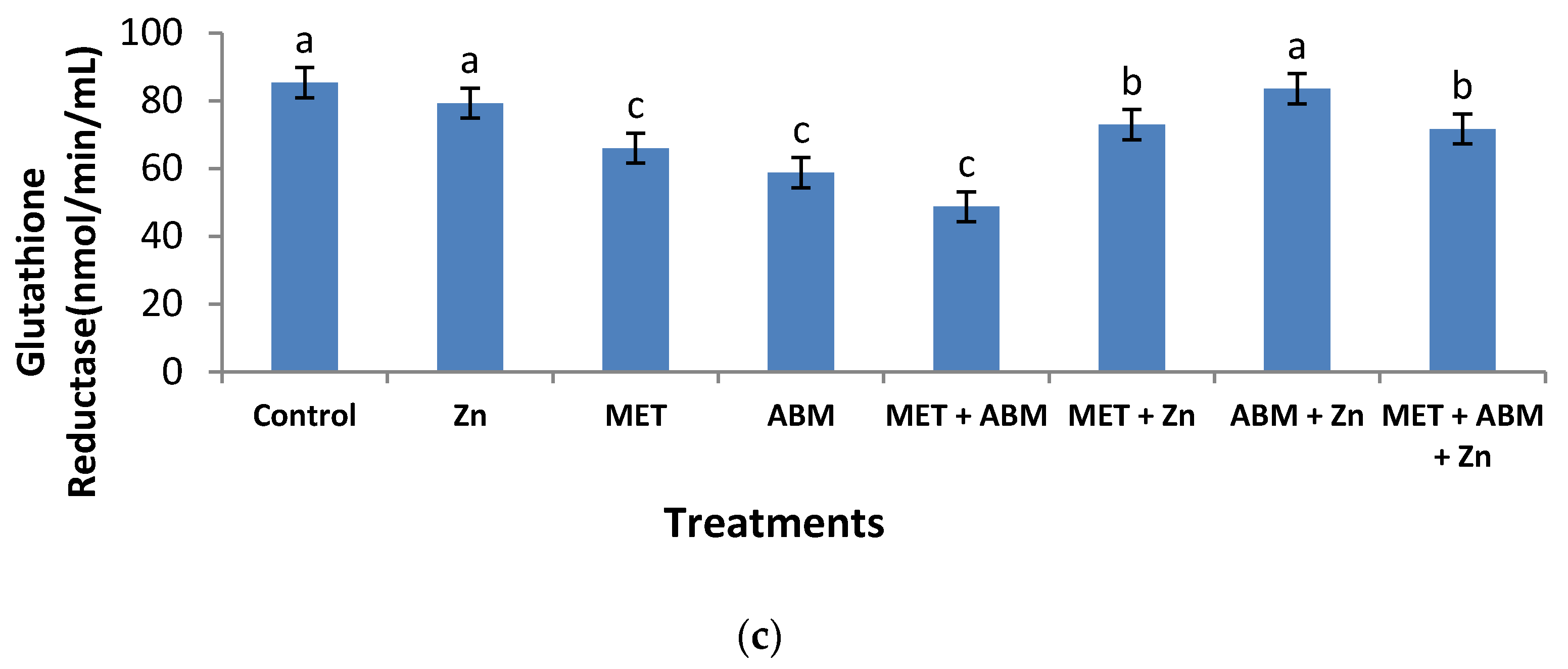
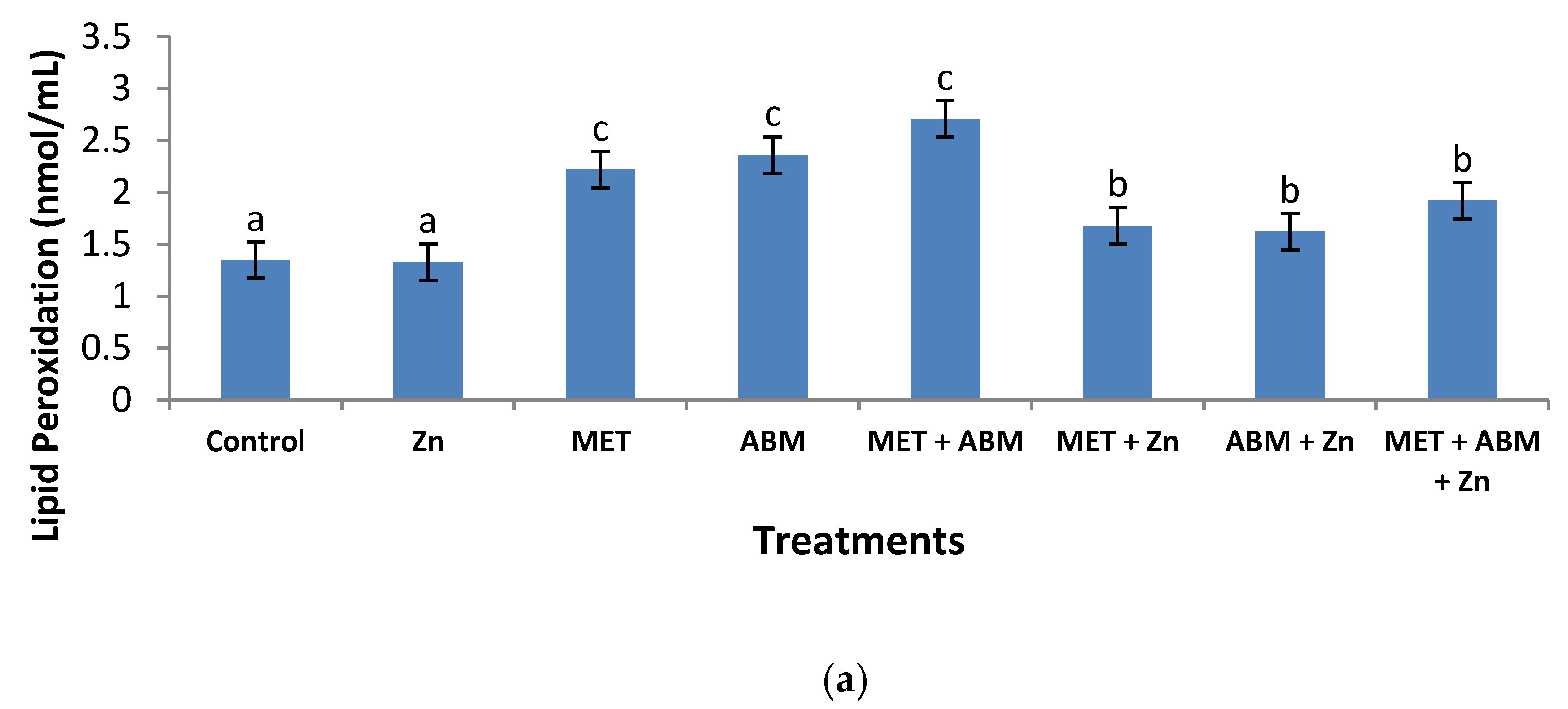
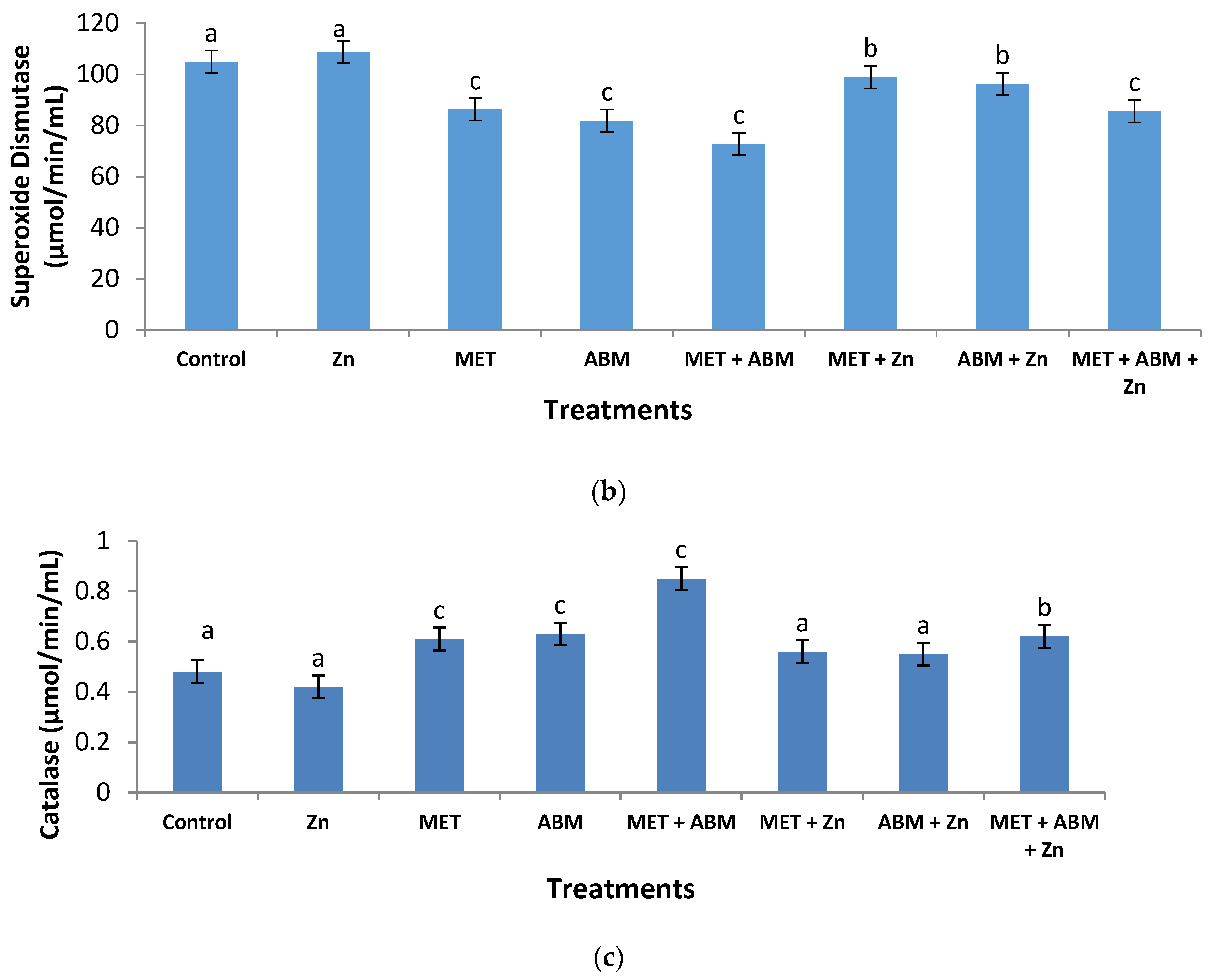
3.3. Effect on Antioxidant Enzymes and Lipid Peroxidation
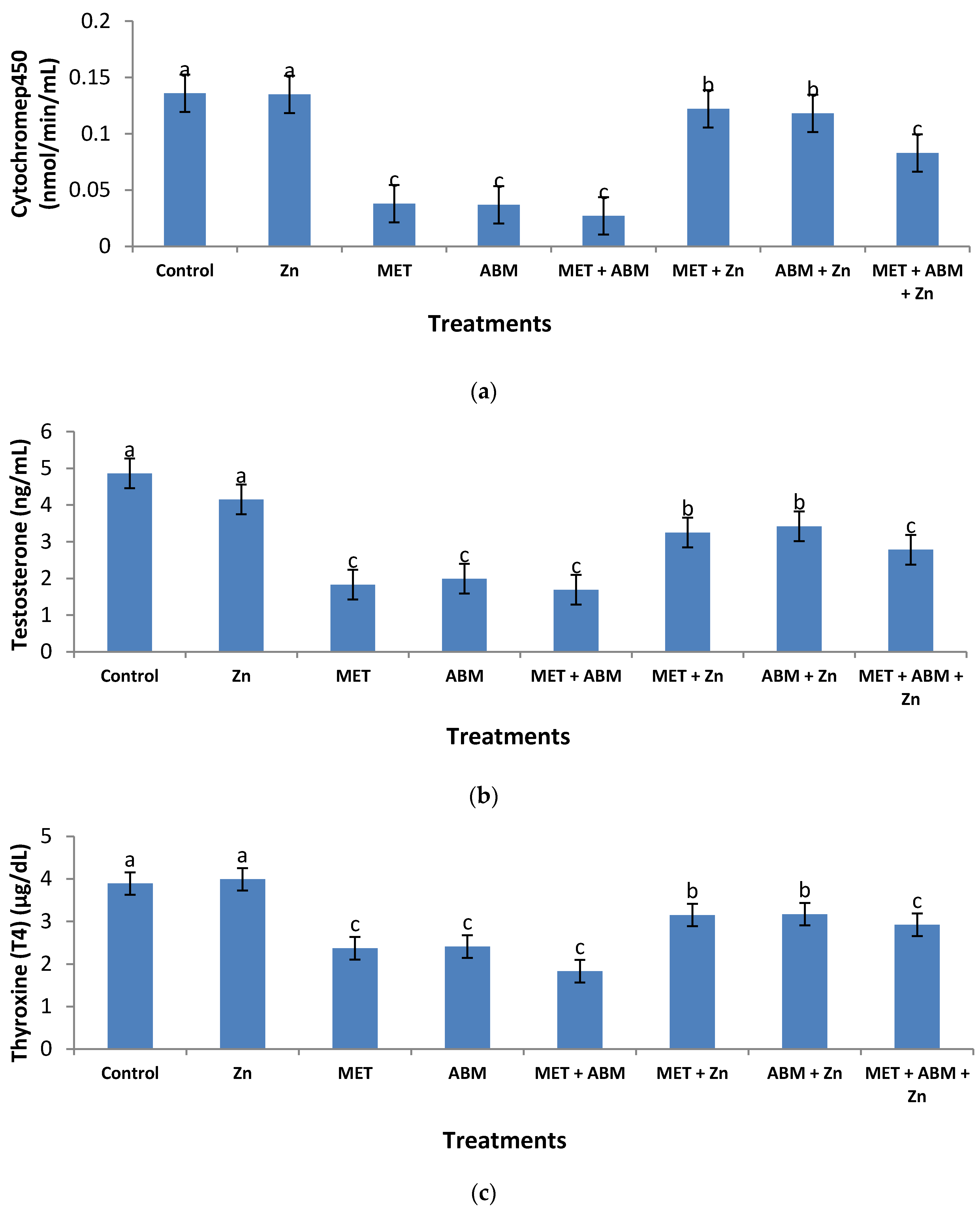
3.4. Effect on Testosterone and Thyroxine Hormones
3.5. Estimation of Joint Action
3.6. Evaluation of Oxidative Stress and Amelioration Effects
4. Discussion
5. Conclusions
6. Recommendations
- ➢
- The use of pesticide mixtures of “antagonistic action” should be encouraged to protect public health.
- ➢
- Most current global environmental agencies derive and regulate their pesticide standard values on individual pesticides. This process must take pesticide mixtures into account when developing toxicological and regulatory standards.
- ➢
- Development of protective agents against pesticide-induced oxidative stress should be among the toxicological standards sponsored by regulatory agencies.
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Kidd, H.; James, D. The Agrochemicals Handbook, 3rd ed.; Royal Society of Chemistry Information Services: Cambridge, UK, 1991. [Google Scholar]
- Moser, V.C.; Katherine, L.M.; Pamela, M.P.; Anna, B.L. Time-course, dose-response, and age comparative sensitivity of N-methyl carbamates in rats. Toxicol. Sci. 2010, 114, 113–123. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. The WHO Recommended Classification of Pesticides by Hazard and Guidelines to Classification: 2004; NLM Classification: Geneva, Switzerland, 2005; pp. 240–256.
- Mansour, S.A.; Mossa, A.H.; Heikal, T.M. Effects of methomyl on lipid peroxidation and antioxidant enzymes in rat erythrocytes: In vitro studies. Toxicol. Ind. Health 2009, 25, 557–563. [Google Scholar] [CrossRef] [PubMed]
- Burg, R.W.; Stapley, E.O. Isolation and characterization of the producing organism. In Ivermectin and Avermectin; Campbell, W.C., Ed.; Springer: New York, NY, USA, 1989; pp. 24–32. [Google Scholar]
- Kolar, L.; Erzen, N.K.; Hogerwerf, L.; Van Gestel, C.A.M. Toxicity of abamectin and doramectin to soil invertebrates. Environ. Pollut. 2008, 151, 182–189. [Google Scholar] [CrossRef] [PubMed]
- Seixas, J.N.; Peixoto, P.V.; Armién, A.G.; Jabour, F.F.; Brito, M.F. Clinical and pathogenetic aspects of abamectin poisoning in calves. Pesq. Vet. Bras. 2006, 26, 161–166. [Google Scholar] [CrossRef]
- Clark, J.M.; Scott, J.G.; Campos, F.; Bloomquist, J.R. Resistance to avermectins: Extent, mechanisms, and management implications. Annu. Rev. Entomol. 1995, 40, 1–30. [Google Scholar] [CrossRef] [PubMed]
- Andreia, N.O.; Jardim, E.; Caldas, D. Brazilian monitoring programs for pesticide residues in food e Results from 2001 to 2010. Food Control 2012, 25, 607–616. [Google Scholar]
- EL-Kabbany, S.M.; EL-Marsafy, A.M.; Mohamed, G.G.; Saadieh, M.M. Residues and pre-harvest interval of abamectin, diniconazole, methomyl and phenthoate in sweet pepper under greenhouse conditions. J. Plant Prot. Pathol. Mansoura Univ. 2013, 4, 1025–1033. [Google Scholar]
- Gad Alla, S.A.; Almaz, M.M.; Wasfi, M.; Thabet, W.F.; Nabil, M.M. Evaluation of Pesticide Residues in some Egyptian Fruits. Int. J. Environ. 2015, 4, 87–97. [Google Scholar]
- Radwan, O.M.M.; Khorshed, M.A.; El-Gohary, A.A.; Sorour, M. Monitoring of Some Pesticide Residues in Consumed Green Beans in Egypt. Mid. East J. Appl. Sci. 2016, 6, 70–77. [Google Scholar]
- Akhgari, M.; Abdollahi, M.; Kebryaeezadeh, A.; Hosseini, R.; OmidSabzevari, O. Biochemical evidence for free radical induced lipid peroxidation as a mechanism for sub chronic toxicity of malathion in blood and liver of rats. Hum. Exp. Toxicol. 2013, 22, 205–211. [Google Scholar] [CrossRef] [PubMed]
- Mansour, S.A.; Mossa, A.H. Lipid peroxidation and oxidative estress in rat erythrocytes induced by chlorpyrifos and the protective effect of zinc. Pestic. Biochem. Physiol. 2009, 93, 34–39. [Google Scholar] [CrossRef]
- Mansour, S.A.; Mossa, A.H. Oxidative damage, biochemical and histopathological alterations in rats exposed to chlorpyrifos and the antioxidant role of zinc. Pestic. Biochem. Physiol. 2010, 96, 14–23. [Google Scholar] [CrossRef]
- Mansour, S.A.; Barakat, A.A.; Mahafrash, S.M.; Heikal, T.M.; El Mahy, S.A. Ameliorative effect of selenium on the hepatotoxicity of methomyl, some common drugs and their combinations. ScienceJet 2012, 1, 8. [Google Scholar]
- Mansour, S.A.; Ali, A.R.; Mohamed, R.I. Toxicity of different doses of methomyl to male rats and the protective effect of zinc especially at high lethal doses. Curr. Top. Toxicol. 2017, 13, 81–93. [Google Scholar]
- Magdy, B.W.; Mohamed, F.E.; Amin, A.S.; Rana, S.S. Ameliorative effect of antioxidants (vitamins C and E) against abamectin toxicity in liver, kidney and testis of male albino rats. J. Basic Appl. Zool. 2016, 77, 69–82. [Google Scholar] [CrossRef]
- Abd-Elhady, H.K.; GamalElsayedAbou-Elghar, G.E. Abamectin induced biochemical and histopathological changes in the Albino rat, Rattus norvegicus. J. Plant Prot. Res. 2013, 53, 263–270. [Google Scholar] [CrossRef]
- Khaldoun-Oularbi, H.; Richeval, C.; Djenas, N.; Lhermitte, M.; Humbert, L.; Bag, A. Effect of subacute exposure to abamectin (insecticide) on liver rat (Rattusnorvegicus). Ann. Toxicol. Anal. 2013, 25, 63–70. [Google Scholar] [CrossRef]
- Mansour, S.A.; Refaie, A.A. Xenobiotics Interaction.2. An approach to the use of biochemical data measurements for interpreting interaction of insecticide mixtures in rat. Adv. Pharmacol. Toxicol. 2000, 1, 1–20. [Google Scholar]
- Nasr, H.M.; El-Demerdash, F.M.; El-Nagar, W.A. Neuro and renal toxicity induced by chlorpyrifos and abamectin in rats: Toxicity of insecticide mixture. Environ. Sci. Pollut. Res. 2016, 23, 1852–1859. [Google Scholar] [CrossRef] [PubMed]
- National Research Council (NRC). Guide for the Care and Use of Laboratory Animals; National Research Council: Washington, DC, USA, 1996; p. 12.
- Finney, D.J. Propit Analysis, 3rd ed.; Cambridge University Press: Cambridge, UK, 1971; p. xv+333. [Google Scholar]
- Goel, A.; Dani, V.; Dhawan, D.K. Protective effects of zinc on lipid peroxidation, antioxidant enzymes and hepatic histoarchitecture in chlorpyrifos-induced toxicity. Chem. Biol. Interact. 2005, 156, 131–140. [Google Scholar] [CrossRef] [PubMed]
- Mansour, S.A.; Gamet-Payrastre, L. Ameliorative effect of vitamin E to mouse dams and their pups following exposure of mothers to chlorpyrifos during gestation and lactation periods. Toxicol. Ind. Health 2016, 32, 1179–1196. [Google Scholar] [CrossRef] [PubMed]
- Akhtar, A.; Deshmukh, A.A.; Raut, C.G.; Somkuwar, A.P.; Bhagat, S.S. Prallethrin induced serum biochemical changes in Wistar rats. Pestic. Biochem. Physiol. 2012, 102, 160–168. [Google Scholar] [CrossRef]
- Evans, C.O. General Introduction. In Animal Clinical Chemistry a Primer for Toxicologists; Evans, G.O., Ed.; USA Taylor & Francis Inc.: Bristol, UK, 1969; pp. 1–9. [Google Scholar]
- Čolović, M.B.; Krstić, D.Z.; Lazarević-Pašti, T.D.; Bondžić, A.M.; Vasić, V.M. Acetylcholinesterase Inhibitors: Pharmacology and Toxicology. Curr. Neuropharmacol. 2013, 11, 315–335. [Google Scholar] [CrossRef] [PubMed]
- El-Shenawy, N.S. Effects of insecticides fenitrothion, endosulfan and abamectin on antioxidant parameters of isolated rat hepatocytes. Toxicol. In Vitro 2010, 24, 1148–1157. [Google Scholar] [CrossRef] [PubMed]
- Bhagat, J.; Ingole, B.S.; Singh, N. Glutathione S-transferase, catalase, superoxide dismutase, glutathione peroxidase, and lipid peroxidation as biomarkers of oxidative stress in snails: A review. Invertebr. Surviv. J. 2016, 13, 336–349. [Google Scholar]
- Banerjee, B.D.; Seth, V.; Ahmed, R.S. Pesticide-induced oxidative stress: Perspective and trends. Rev. Environ. Health 2001, 16, 1–40. [Google Scholar] [CrossRef] [PubMed]
- Powell, S.R. Theantioxidant properties of zinc. J. Nutr. 2000, 130, 1447S–1454S. [Google Scholar] [PubMed]
- Debnath, D.; Mandal, T.K. Study of quinalphos (an environmental oestrogenic insecticide) formulation (Ekalux 25 E.C.)—Induced damage of the testicular tissues and antioxidant defence systems in Sprague-Dawley albino rats. J. Appl. Toxicol. 2000, 20, 197–204. [Google Scholar] [CrossRef]
- Bhattacharya, S. Reactive oxygen species and cellular defense system. In Free Radicals in Human Health and Disease; Rani, V., Yadav, U.C.S., Eds.; Springer: New Delhi, India, 2015; pp. 17–29. [Google Scholar]
- Wang, X.; Kanel, G.C.; DeLeve, L.D. Support of sinusoidal endothelial cell glutathione prevents hepatic veno-occlusive disease in the rat. Hepatology 2000, 31, 428–434. [Google Scholar] [CrossRef] [PubMed]
- Rao, A.V.; Shaha, C. Role of glutathione S-transferases in oxidative stress-induced male germ cell apoptosis. Free Radic. Biol. Med. 2000, 29, 1015–1027. [Google Scholar] [CrossRef]
- Kyle, M.E.; Miccadei, S.; Nakae, D.; Farber, J.L. Superoxide dismutase and catalase protect cultured hepatocytes from the cytotoxicity of acetaminophen. Biochem. Biophys. Res. Commun. 1987, 149, 889–896. [Google Scholar] [CrossRef]
- Verma, R.S.; Srivastava, N. Effect of chlorpyrifos on thiobarbituric acid reactive substances, scavenging enzymes and glutathione in rat tissues. Ind. J. Biochem. Biophys. 2003, 40, 423–428. [Google Scholar]
- White, R.E. The involvement of free radicals in the mechanisms of monooxygenases. Pharmacol. Ther. 1991, 49, 21–42. [Google Scholar] [CrossRef]
- Yamano, T.; Morita, S. Hepatotoxicity of trichlorfon and dichlorvos in isolated rat hepatocytes. Toxicology 1992, 76, 69–77. [Google Scholar] [CrossRef]
- Timbrell, J. Principles of Biochemical Toxicology, 3rd ed.; Taylor and Francis: London, UK, 2000; 776p. [Google Scholar]
- Dias, V.; Junn, E.; Mouradian, M.M. The role of oxidative stress in Parkinson’s Disease. J. Parkinsons Dis. 2013, 3, 461–491. [Google Scholar] [PubMed]
- El-Kashoury, A.A.; Salama, A.F.; Mohamed, R.A. Chronic exposure of dicofol promotes reproductive toxicity in male rats. Life Sci. J. 2010, 7, 5–19. [Google Scholar]
- Fattahi, E.; Parivar, K.; Jorsaraei, S.G.A.; Moghadamnia, A.A. The effects of diazinon on testosterone, FSH and LH levels and testicular tissue in mice. Iran. J. Reprod. Med. 2009, 7, 59–64. [Google Scholar]
- Hassan, H.F.; Meligi, N.M. Abamectin exposure on some hormonal profiles and testicular histopathology in male albino rats and the possible ameliorative role of Eruca sativa. Environ. Sci. Pollut. Res. Int. 2017, 24, 24690–24697. [Google Scholar]
- Pereira, B.; Rosa, L.F.; Safi, D.A.; Bechara, E.J.; Curi, R. Control of superoxide dismutase, catalase and glutathione peroxidase activities in rat lymphoid organs by thyroid hormones. J. Endocrinol. 1994, 140, 73–77. [Google Scholar] [CrossRef] [PubMed]
- Asayama, K.; Dobashi, K.; Hayashibe, H.; Megata, Y.; Kato, K. Lipid peroxidation and free radical scavengers in thyroid dysfunction in the rat: A possible mechanism of injury to heart and skeletal muscles in hyperthyroidism. Endocrinology 1987, 121, 2112–2118. [Google Scholar] [CrossRef] [PubMed]
- Zia-ur-Rahman, N.A.; Bukhari, S.A.; Akhtar, N.; Haq, I.U. Serum hormonal, electrolytes and trace element profiles in the rutting and non-rutting onehumped male camel (Camelusdromedarius). Anim. Reprod. Sci. 2007, 101, 172–178. [Google Scholar] [CrossRef] [PubMed]
- El-Kashoury, A.A.; El-Far, F.A. Effect of two products of profenofos on thyroid gland, lipid profile and plasma APO-1/FAS in adult male albino rats. Egypt. J. Basic Appl. Physiol. 2004, 3, 213–226. [Google Scholar]
- De Angelis, S.; Tassinari, R.; Maranghi, F.; Eusepi, A.; Di Virgilio, A.; Chiarotti, F.; Ricceri, L.; Pesciolini, A.V.; Gilardi, E.; Moracci, G.; et al. Developmental exposure to chlorpyrifos induces alteration in thyroid and thyroid hormone levels without other toxicity signs in Cd mice. Toxicol. Sci. 2009, 108, 311–319. [Google Scholar] [CrossRef] [PubMed]
- Jeong, S.H.; Kim, B.Y.; Kang, H.G.; Ku, H.O.; Cho, J.H. Effect of chlorpyrifos-methyl on steroid and thyroid hormones in rat F0- and F1-generations. Toxicology 2006, 220, 189–202. [Google Scholar] [CrossRef] [PubMed]
- Mahgoub, A.A.; El-Medany, A.H. Evaluation of chronic exposure of the male rat reproductive system to the insecticide methomyl. Pharmacol. Res. 2001, 44, 273–280. [Google Scholar] [CrossRef] [PubMed]
- Bettger, W.J.; O’Dell, B.L. A critical physiological role of zinc in the structure and function of biomembranes. Life Sci. 1981, 28, 1425–1438. [Google Scholar] [CrossRef]
- Rouimi, P.; Zucchini-Pascal, N.; Dupont, G.; Razpotnik, A.; Fouché, E.; DeSousa, G.; Rahmani, R. Impacts of low doses of pesticide mixtures on liver cell defence systems. Toxicol. In Vitro 2012, 26, 718–726. [Google Scholar] [CrossRef] [PubMed]
- Belden, J.B.; Lydy, M.J. Impact of atrazine on organophosphate insecticide toxicity. Environ. Toxicol. Chem. 2000, 19, 2266–2274. [Google Scholar] [CrossRef]
- DeLorenzo, M.E.; Lorimar, S. Individual and mixture toxicity of three pesticides; atrazine, chlorpyrifos, and chlorothalonil to the marine phytoplankton species Dunaliellatertiolecta. J. Environ. Sci. Health Part B 2003, 5, 529–538. [Google Scholar] [CrossRef] [PubMed]
- Roloff, D.B.; Bulluck, A.D.; Meisner, F.L. Cytogenetic studies of herbicide interactions in vitro and in vivo using atrazine and linuron. Arch. Environ. Contam. Toxicol. 1992, 22, 267–271. [Google Scholar] [CrossRef] [PubMed]
- Krishnan, K.; Clewell Ill, H.J.; Anderson, M.E. Physiologically based pharmacokinetic analysis of simple mixtures. Environ. Health Perspect. 1994, 102 (Suppl. 9), 151–155. [Google Scholar] [CrossRef] [PubMed]
- Oesch, E.; Oesch, B.; Arens, J.; Fahndrich, E.; Vogel, E.; Friedberg, T.; Glatt, H. Mechanism—Based predictions of interactions. Environ. Health Perspect. 1994, 102 (Suppl. 9), 5–9. [Google Scholar] [CrossRef] [PubMed]
- Yang, R.S.H. Some critical issues and concerns related to research advances on the toxicity of chemical mixtures. Environ. Health Perspect. 1998, 106 (Suppl. 4), 1059–1063. [Google Scholar] [PubMed]
- Mansour, S.A.; Gamet-Payrastre, L.; Mohamed, D.A.; Gad, M.F. Ameliorative effect of vitamin E to mice dams and their pups following exposure of mothers to atrazine during gestation and lactation periods. ScienceJet 2013, 2, 32. [Google Scholar]
- Mansour, S.A.; Mohamed, D.A.; Gamet-Payrastre, L. Indirect exposure of mice pups to endosulfan via their dams during gestation and lactation periods and the ameliorative effect of vitamin E. Hum. Exp. Toxicol. 2014, 33, 911–927. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Biochemical Parameter | Control Value (C) | MET Value (A) | ABM Value (B) | MET+ABM Value (M) | Interaction Index (I.I.) | Joint Action |
|---|---|---|---|---|---|---|
| MDA †† | 1.35 | 2.22 | 2.36 | 2.71 | 0.89 | An |
| nmol/mL | ||||||
| SOD † | 104.87 | 86.23 | 81.85 | 72.71 | 1.06 | An |
| µmol/min/mL | ||||||
| CAT †† | 0.48 | 0.61 | 0.63 | 0.85 | 1.07 | Po |
| µmol/min/mL | ||||||
| GPx † | 0.87 | 0.73 | 0.76 | 0.69 | 1.05 | Ad |
| µmol/min/mL | ||||||
| GR † | 85.37 | 66.01 | 58.80 | 48.73 | 1.07 | An |
| nmol/min/mL | ||||||
| GST † | 1.07 | 0.85 | 0.85 | 0.80 | 1.10 | An |
| µmol/min/mL | ||||||
| CYP450 † | 0.14 | 0.04 | 0.04 | 0.03 | 2.13 | An |
| nmol/min/mL | ||||||
| T † | 4.86 | 1.83 | 1.99 | 1.69 | 1.71 | An |
| ng/mL | ||||||
| T4 † | 3.89 | 2.37 | 2.41 | 1.83 | 1.20 | An |
| µg/dL |
| Treatment | Biochemical Parameters | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| MDA | SOD | CAT | GPx | GR | GST | CYP450 | T | T4 | |
| nmol/mL | µmol/min/mL | µmol/min/mL | µmol/min/mL | nmol/min/mL | µmol/min/mL | nmol/min/mL | ng/mL | µg/dL | |
| Control (a) | 1.35 | 104.87 | 0.48 | 0.87 | 85.37 | 1.07 | 0.136 | 4.86 | 3.89 |
| Methomyl | |||||||||
| MET (b) | 2.22 | 86.23 | 0.61 | 0.73 | 66.01 | 0.85 | 0.038 | 1.83 | 2.37 |
| MET+Zn (c) | 1.68 | 98.81 | 0.56 | 0.89 | 72.95 | 0.92 | 0.122 | 3.25 | 3.15 |
| % of Change * | 64.44 | −17.77 | 27.08 | −16.09 | −22.68 | −20.56 | −72.06 | −62.35 | −39.07 |
| Ameliorative Index ** | 1.24 | 0.94 | 1.17 | 1.02 | 0.85 | 0.86 | 0.90 | 0.67 | 0.81 |
| Abamectin | |||||||||
| ABM (b) | 2.36 | 81.85 | 0.63 | 0.76 | 58.80 | 0.85 | 0.037 | 1.99 | 2.41 |
| ABM+Zn (c) | 1.62 | 96.18 | 0.55 | 0.88 | 83.55 | 0.93 | 0.118 | 3.42 | 3.17 |
| % of Change * | 74.81 | −21.95 | 31.25 | −13.8 | −31.12 | −20.56 | −72.79 | −59.06 | −38.05 |
| Ameliorative Index ** | 1.20 | 0.92 | 1.15 | 1.0 | 0.98 | 0.87 | 0.87 | 0.70 | 0.82 |
| Methomyl+Abamectin | |||||||||
| MET+ABM (b) | 2.71 | 72.71 | 0.85 | 0.69 | 48.73 | 0.80 | 0.027 | 1.69 | 1.83 |
| MET+ABM+Zn (c) | 1.92 | 85.55 | 0.62 | 0.82 | 71.68 | 0.82 | 0.083 | 2.78 | 2.92 |
| % of Change * | 100.74 | −30.67 | 77.08 | −20.69 | −42.92 | −25.23 | −80.15 | −65.23 | −52.96 |
| Ameliorative Index ** | 1.42 | 0.82 | 1.29 | 0.94 | 0.84 | 0.77 | 0.61 | 0.59 | 0.75 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mansour, S.A.; Abbassy, M.A.; Shaldam, H.A. Zinc Ameliorate Oxidative Stress and Hormonal Disturbance Induced by Methomyl, Abamectin, and Their Mixture in Male Rats. Toxics 2017, 5, 37. https://doi.org/10.3390/toxics5040037
Mansour SA, Abbassy MA, Shaldam HA. Zinc Ameliorate Oxidative Stress and Hormonal Disturbance Induced by Methomyl, Abamectin, and Their Mixture in Male Rats. Toxics. 2017; 5(4):37. https://doi.org/10.3390/toxics5040037
Chicago/Turabian StyleMansour, Sameeh A., Mostafa A. Abbassy, and Hassan A. Shaldam. 2017. "Zinc Ameliorate Oxidative Stress and Hormonal Disturbance Induced by Methomyl, Abamectin, and Their Mixture in Male Rats" Toxics 5, no. 4: 37. https://doi.org/10.3390/toxics5040037
APA StyleMansour, S. A., Abbassy, M. A., & Shaldam, H. A. (2017). Zinc Ameliorate Oxidative Stress and Hormonal Disturbance Induced by Methomyl, Abamectin, and Their Mixture in Male Rats. Toxics, 5(4), 37. https://doi.org/10.3390/toxics5040037