
Hetero-Aggregation of Nanoplastics with Freshwater Algae and the Toxicological Consequences: The Role of Extracellular Polymeric Substances


 ,
,
Abstract
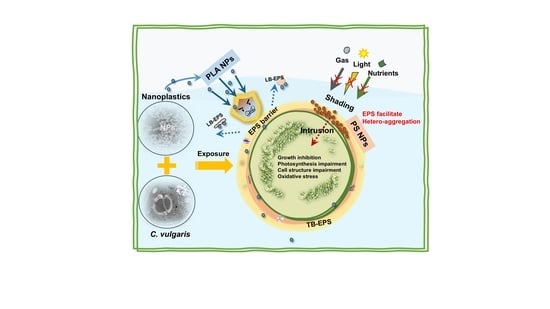
1. Introduction
2. Materials and Methods
2.1. NP Preparation
2.2. Algae Preparation
2.3. Analysis of Hetero-Aggregation Between NPs and Algae
2.3.1. Settling Experiments
2.3.2. DLVO Theory Analysis
2.3.3. EPS Analysis
2.3.4. Algal Ultrastructure Analysis
2.4. NP Toxicity Experiments
2.4.1. NP Exposures
2.4.2. Determination of Algal Growth
2.4.3. Determination of Pigment Synthesis
2.4.4. Determination of Algal Membrane Integrity
2.4.5. Determination of Oxidative Stress
2.4.6. Integrated Biomarker Response Analysis
2.5. Statistical Analysis
3. Results and Discussion
3.1. Hetero-Aggregations Between NPs and Algae in Settling Experiments
3.2. Influence of EPS on Hetero-Aggregations Between NPs and Algae
3.3. NP Toxicity to Algae
3.3.1. Effects of NPs on Algal Growth
3.3.2. Effects of NPs on Algal Pigment Synthesis
3.3.3. Effects of NPs on Algal Membrane Integrity
3.3.4. Effects of NPs on Algal Antioxidant System
3.3.5. Integrated Biomarker Response
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Huang, W.; Xia, X. Element cycling with micro (nano) plastics. Science 2024, 385, 933–935. [Google Scholar] [CrossRef]
- Gigault, J.; Ter Halle, A.; Baudrimont, M.; Pascal, P.; Gauffre, F.; Phi, T.; El Hadri, H.; Grassl, B.; Reynaud, S. Current Opinion: What is a nanoplastic? Environ. Pollut. 2018, 235, 1030–1034. [Google Scholar] [CrossRef] [PubMed]
- Gigault, J.; El Hadri, H.; Nguyen, B.; Grassl, B.; Rowenczyk, L.; Tufenkji, N.; Feng, S.; Wiesner, M. Nanoplastics are neither microplastics nor engineered nanoparticles. Nat. Nanotechnol. 2021, 16, 501–507. [Google Scholar] [CrossRef] [PubMed]
- Pradel, A.; Catrouillet, C.; Gigault, J. The environmental fate of nanoplastics: What we know and what we need to know about aggregation. NanoImpact 2023, 29, 100453. [Google Scholar] [CrossRef] [PubMed]
- Athulya, P.A.; Chandrasekaran, N. Interactions of natural colloids with microplastics in aquatic environment and its impact on FTIR characterization of polyethylene and polystyrene microplastics. J. Mol. Liq. 2023, 369, 120950. [Google Scholar] [CrossRef]
- Veclin, C.; Desmet, C.; Pradel, A.; Valsesia, A.; Ponti, J.; El Hadri, H.; Maupas, T.; Pellerin, V.; Gigault, J.; Grassl, B.; et al. Effect of the surface hydrophobicity-morphology-functionality of nanoplastics on their homoaggregation in seawater. ACS EST Water 2022, 2, 88–95. [Google Scholar] [CrossRef]
- Oriekhova, O.; Stoll, S. Heteroaggregation of nanoplastic particles in the presence of inorganic colloids and natural organic matter. Environ. Sci. Nano 2018, 5, 792–799. [Google Scholar] [CrossRef]
- Parrella, F.; Brizzolara, S.; Holzner, M.; Mitrano, D.M. Impact of heteroaggregation between microplastics and algae on particle vertical transport. Nat. Water 2024, 2, 541–552. [Google Scholar] [CrossRef]
- Déniel, M.; Errien, N.; Daniel, P.; Caruso, A.; Lagarde, F. Current methods to monitor microalgae-nanoparticle interaction and associated effects. Aquat. Toxicol. 2019, 217, 105311. [Google Scholar] [CrossRef]
- Ge, S.; Agbakpe, M.; Zhang, W.; Kuang, L. Heteroaggregation between PEI-coated magnetic nanoparticles and algae: Effect of particle size on algal harvesting efficiency. ACS Appl. Mater. Interfaces 2015, 7, 6102–6108. [Google Scholar] [CrossRef]
- Sendra, M.; Yeste, M.P.; Gatica, J.M.; Moreno-Garrido, I.; Blasco, J. Homoagglomeration and heteroagglomeration of TiO2, in nanoparticle and bulk form, onto freshwater and marine microalgae. Sci. Total Environ. 2017, 592, 403–411. [Google Scholar] [CrossRef] [PubMed]
- Lagarde, F.; Olivier, O.; Zanella, M.; Daniel, P.; Hiard, S.; Caruso, A. Microplastic interactions with freshwater microalgae: Hetero-aggregation and changes in plastic density appear strongly dependent on polymer type. Environ. Pollut. 2016, 215, 331–339. [Google Scholar] [CrossRef] [PubMed]
- Flemming, H.C.; van Hullebusch, E.D.; Little, B.J.; Neu, T.R.; Nielsen, P.H.; Seviour, T.; Stoodley, P.; Wingender, J.; Wuertz, S. Microbial extracellular polymeric substances in the environment, technology and medicine. Nat. Rev. Microbiol. 2025, 23, 87–105. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Bolan, N.; Tsang, D.C.; Sarkar, B.; Bradney, L.; Li, Y. A review of microplastics aggregation in aquatic environment: Influence factors, analytical methods, and environmental implications. J. Hazard. Mater. 2021, 402, 123496. [Google Scholar] [CrossRef] [PubMed]
- Cunha, C.; Faria, M.; Nogueira, N.; Ferreira, A.; Cordeiro, N. Marine vs freshwater microalgae exopolymers as biosolutions to microplastics pollution. Environ. Pollut. 2019, 249, 372–380. [Google Scholar] [CrossRef]
- Su, Y.; Gao, L.; Xu, E.G.; Peng, L.; Diao, X.; Zhang, Y.; Bao, R. When microplastics meet microalgae: Unveiling the dynamic formation of aggregates and their impact on toxicity and environmental health. Water Res. 2025, 273, 123008. [Google Scholar] [CrossRef]
- Sun, H.; Jiao, R.; Wang, D. The difference of aggregation mechanism between microplastics and nanoplastics: Role of Brownian motion and structural layer force. Environ. Pollut. 2021, 268, 115942. [Google Scholar] [CrossRef]
- Basuvaraj, M.; Fein, J.; Liss, S.N. Protein and polysaccharide content of tightly and loosely bound extracellular polymeric substances and the development of a granular activated sludge floc. Water Res. 2015, 82, 104–117. [Google Scholar] [CrossRef]
- Guo, X.; Wang, X.; Liu, J. Composition analysis of fractions of extracellular polymeric substances from an activated sludge culture and identification of dominant forces affecting microbial aggregation. Sci. Rep. 2016, 6, 28391. [Google Scholar] [CrossRef]
- Ma, Y.; Li, T.Y.; Meng, H.; Wang, G.X.; Zhang, L.M.; Jia, G.Z.; Ma, J.; Xiao, Y.; Li, W.H.; Xie, W.M. The contradictory roles of tightly bound and loosely bound extracellular polymeric substances of activated sludge in trimethoprim adsorption process. J. Environ. Manag. 2023, 336, 117661. [Google Scholar] [CrossRef]
- Li, S.; Bai, Y.; Li, Z.; Wang, A.; Ren, N.Q.; Ho, S.H. Overlooked role of extracellular polymeric substances in antibiotic-resistance gene transfer within microalgae-bacteria system. J. Hazard. Mater. 2025, 488, 137206. [Google Scholar] [CrossRef]
- Nigam, H.; Jain, R.; Malik, A.; Singh, V. Effect of different polystyrene nano-plastic concentrations on Chlorella pyrenoidosa. Algal Res. 2022, 67, 102835. [Google Scholar] [CrossRef]
- Nam, S.; Kim, D.; An, Y. Soil algae as a potential carrier for nanoplastics: Adsorption and internalization of nanoplastics in algal cells. Sci. Total Environ. 2022, 837, 155678. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Tao, S.; Liu, S.; Hu, T.; Zheng, K.; Shen, M.; Meng, G. Research advances on impacts micro/nanoplastics and their carried pollutants on algae in aquatic ecosystems: A review. Aquat. Toxicol. 2023, 264, 106725. [Google Scholar] [CrossRef] [PubMed]
- Schröter, L.; Ventura, N. Nanoplastic toxicity: Insights and challenges from experimental model systems. Small 2022, 18, 2201680. [Google Scholar] [CrossRef]
- Zhao, J.; Liu, S.; Liu, N.; Zhang, H.; Zhou, Q.; Ge, F. Accelerated productions and physicochemical characterizations of different extracellular polymeric substances from Chlorella vulgaris with nano-ZnO. Sci. Total Environ. 2019, 658, 582–589. [Google Scholar] [CrossRef]
- Zhang, B.; Wang, Z.; Li, D.; Li, L.; Zhao, Y.; Tang, X.; Zhao, Y. Reactive oxygen species mediated extracellular polymeric substances production assisting the recovery of Thalassiosira pseudonana from polystyrene micro and nanoplastics exposure. Environ. Pollut. 2024, 348, 123850. [Google Scholar] [CrossRef]
- Arif, Y.; Mir, A.R.; Zieliński, P.; Hayat, S.; Bajguz, A. Microplastics and nanoplastics: Source, behavior, remediation, and multi-level environmental impact. J. Environ. Manag. 2024, 356, 120618. [Google Scholar] [CrossRef]
- Liu, Z.; Zhu, Y.; Lv, S.; Shi, Y.; Dong, S.; Yan, D.; Zhu, X.; Peng, R.; Keller, A.A.; Huang, Y. Quantifying the dynamics of polystyrene microplastics UV-aging process. Environ. Sci. Technol. Lett. 2022, 9, 50–56. [Google Scholar] [CrossRef]
- Wang, Z.; Ding, J.; Song, X.; Zheng, L.; Huang, J.; Zou, H.; Wang, Z. Aging of poly (lactic acid)/poly (butylene adipate-co-terephthalate) blends under different conditions: Environmental concerns on biodegradable plastic. Sci. Total Environ. 2023, 855, 158921. [Google Scholar] [CrossRef]
- Zhao, X.; Wu, X.; Wang, Q.; Wu, F. Ecological risks of biodegradable plastics. Science 2025, 388, 1034. [Google Scholar] [CrossRef]
- Zhang, S.; Wang, C. Precise analysis of nanoparticle size distribution in TEM Image. Methods Protoc. 2023, 6, 63. [Google Scholar] [CrossRef]
- Zhou, K.; Hu, Y.; Zhang, L.; Yang, K.; Lin, D. The role of exopolymeric substances in the bioaccumulation and toxicity of Ag nanoparticles to algae. Sci. Rep. 2016, 6, 32998. [Google Scholar] [CrossRef]
- Agmo Hernández, V. An overview of surface forces and the DLVO theory. ChemTexts 2023, 9, 10. [Google Scholar] [CrossRef]
- Sadiq, I.M.; Dalai, S.; Chandrasekaran, N.; Mukherjee, A. Ecotoxicity study of titania (TiO2) NPs on two microalgae species: Scenedesmus sp. and Chlorella sp. Ecotoxicol. Environ. Saf. 2011, 74, 1180–1187. [Google Scholar] [CrossRef]
- Su, Y.; Cheng, Z.; Hou, Y.; Lin, S.; Gao, L.; Wang, Z.; Bao, R.; Peng, L. Biodegradable and conventional microplastics posed similar toxicity to marine algae Chlorella vulgaris. Aquat. Toxicol. 2022, 244, 106097. [Google Scholar] [CrossRef]
- Riccardi, C.; Nicoletti, I. Analysis of apoptosis by propidium iodide staining and flow cytometry. Nat. Protoc. 2006, 1, 1458–1461. [Google Scholar] [CrossRef]
- Beliaeff, B.; Burgeot, T. Integrated biomarker response: A useful tool for ecological risk assessment. Environ. Toxicol. Chem. 2002, 21, 1099–1322. [Google Scholar] [CrossRef]
- Sanchez, W.; Burgeot, T.; Porcher, J.M. A novel “Integrated Biomarker Response” calculation based on reference deviation concept. Environ. Sci. Pollut. Res. 2013, 20, 2721–2725. [Google Scholar] [CrossRef]
- Catteau, A.; Le Guernic, A.; Palos Ladeiro, M.; Dedourge-Geffard, O.; Bonnard, M.; Bonnard, I.; Delahaut, L.; Bado-Nilles, A.; Porcher, J.-M.; Lopes, C.; et al. Integrative biomarker response-Threshold (IBR-T): Refinement of IBRv2 to consider the reference and threshold values of biomarkers. J. Environ. Manag. 2023, 341, 118049. [Google Scholar] [CrossRef]
- Li, X.; Qiu, H.; Zhang, P.; Song, L.; Romero-Freire, A.; He, E. Role of heteroaggregation and internalization in the toxicity of differently sized and charged plastic nanoparticles to freshwater microalgae. Environ. Pollut. 2023, 316, 120517. [Google Scholar] [CrossRef]
- Grassi, G.; Gabellieri, E.; Cioni, P.; Paccagnini, E.; Faleri, C.; Lupetti, P.; Corsi, I.; Morelli, E. Interplay between extracellular polymeric substances (EPS) from a marine diatom and model nanoplastic through eco-corona formation. Sci. Total Environ. 2020, 725, 138457. [Google Scholar] [CrossRef]
- Li, Y.; Zheng, X.; Zhao, Z.; Li, W.; Huang, Y.; He, H.; Han, Z.; Tao, J.; Lin, T. Perfluorobutanoic acid weakens the heterogeneous aggregation of microplastics and microalgae: Perspective from physicochemical properties, extracellular polymeric substances secretion and DLVO theory. Sci. Total Environ. 2024, 955, 177127. [Google Scholar] [CrossRef]
- Kooi, M.; Nes, E.H.V.; Scheffer, M.; Koelmans, A.A. Ups and downs in the ocean: Effects of biofouling on vertical transport of microplastics. Environ. Sci. Technol. 2017, 51, 7963–7971. [Google Scholar] [CrossRef]
- Maqbool, T.; Hur, J. Changes in fluorescent dissolved organic matter upon interaction with anionic surfactant as revealed by EEM-PARAFAC and two dimensional correlation spectroscopy. Chemosphere 2016, 161, 190–199. [Google Scholar] [CrossRef]
- Santschi, P.H.; Xu, C.; Schwehr, K.A.; Lin, P.; Sun, L.; Chin, W.-C.; Kamalanathan, M.; Bacosa, H.P.; Quigg, A. Can the protein/carbohydrate (P/C) ratio of exopolymeric substances (EPS) be used as a proxy for their ‘stickiness’ and aggregation propensity? Mar. Chem. 2020, 218, 103734. [Google Scholar] [CrossRef]
- Xu, J.; Wang, L.; Sun, H. Adsorption of neutral organic compounds on polar and nonpolar microplastics: Prediction and insight into mechanisms based on pp-LFERs. J. Hazard. Mater. 2021, 408, 124857. [Google Scholar] [CrossRef]
- Cheng, Y.; Wang, H. Highly effective removal of microplastics by microalgae Scenedesmus abundans. Chem. Eng. J. 2022, 435, 135079. [Google Scholar] [CrossRef]
- Wang, S.; Al-Hasni, N.S.; Liu, Z.; Liu, A. Multifaceted aquatic environmental differences between nanoplastics and microplastics: Behavior and fate. Environ. Health 2024, 2, 688–701. [Google Scholar] [CrossRef]
- Feng, Y.; Feng, L.J.; Liu, S.C.; Duan, J.L.; Zhang, Y.B.; Li, S.C.; Sun, X.D.; Wang, S.G.; Yuan, X.Z. Emerging investigator series: Inhibition and recovery of anaerobic granular sludge performance in response to short-term polystyrene nanoparticle exposure. Environ. Sci. Water Res. Technol. 2018, 4, 1902–1911. [Google Scholar] [CrossRef]
- Rossi, G.; Barnoud, J.; Monticelli, L. Polystyrene nanoparticles perturb lipid membranes. J. Phys. Chem. Lett. 2014, 5, 241–246. [Google Scholar] [CrossRef]
- Su, Y.; Gao, L.; Peng, L.; Diao, X.; Lin, S.; Bao, R.; Mehmood, T. Heterogeneous aggregation between microplastics and microalgae: May provide new insights for microplastics removal. Aquat. Toxicol. 2023, 261, 106638. [Google Scholar] [CrossRef]
- Lee, S.; Ahn, C.; Kim, E.; Park, J.; Joo, J. Growth inhibition of harmful algae using TiO2-embedded expanded polystyrene balls in the hypereutrophic stream. J. Hazard. Mater. 2020, 398, 123172. [Google Scholar] [CrossRef]
- Yan, Z.; Xu, L.; Zhang, W.; Yang, G.; Zhao, Z.; Wang, Y.; Li, X. Comparative toxic effects of microplastics and nanoplastics on Chlamydomonas reinhardtii: Growth inhibition, oxidative stress, and cell morphology. J. Water Process Eng. 2021, 43, 102291. [Google Scholar] [CrossRef]
- Hu, M.; Palić, D. Micro- and nano-plastics activation of oxidative and inflammatory adverse outcome pathways. Redox Biol. 2020, 37, 101620. [Google Scholar] [CrossRef]
- Junaid, M.; Wang, J. Interaction of nanoplastics with extracellular polymeric substances (EPS) in the aquatic environment: A special reference to eco-corona formation and associated impacts. Water Res. 2021, 201, 117319. [Google Scholar] [CrossRef]
- Gao, X.; Zhou, K.; Zhang, L.; Yang, K.; Lin, D. Distinct effects of soluble and bound exopolymeric substances on algal bioaccumulation and toxicity of anatase and rutile TiO2 nanoparticles. Environ. Sci. Nano 2018, 5, 720–729. [Google Scholar] [CrossRef]
- Xu, H.; Pan, J.; Zhang, H.; Yang, L. Interactions of metal oxide nanoparticles with extracellular polymeric substances (EPS) of algal aggregates in an eutrophic ecosystem. Ecol. Eng. 2016, 94, 464–470. [Google Scholar] [CrossRef]
- Wang, C.; Jiang, L.; Huang, W.; Wang, C.; He, M. Light availability modulates the responses of the microalgae Desmodesmus sp. to micron-sized polyvinyl chloride microplastics. Aquat. Toxicol. 2022, 249, 106234. [Google Scholar] [CrossRef]
- Zhang, X.; Ma, G.; Wei, W. Simulation of nanoparticles interacting with a cell membrane: Probing the structural basis and potential biomedical application. NPG Asia Mater. 2021, 13, 52. [Google Scholar] [CrossRef]
- Giangreco, G.; Malabarba, M.G.; Sigismund, S. Specialised endocytic proteins regulate diverse internalisation mechanisms and signalling outputs in physiology and cancer. Biol. Cell 2021, 113, 165–182. [Google Scholar] [CrossRef]
- Tsikas, D. Assessment of lipid peroxidation by measuring malondialdehyde (MDA) and relatives in biological samples: Analytical and biological challenges. Anal. Biochem. 2017, 524, 13–30. [Google Scholar] [CrossRef]
- Li, X.; Luo, J.; Zeng, H.; Zhu, L.; Lu, X. Microplastics decrease the toxicity of sulfamethoxazole to marine algae (Skeletonema costatum) at the cellular and molecular levels. Sci. Total Environ. 2022, 824, 153855. [Google Scholar] [CrossRef] [PubMed]
- Tang, B.; Zhang, L.; Salam, M.; Yang, B.; He, Q.; Yang, Y.; Li, H. Revealing the environmental hazard posed by biodegradable microplastics in aquatic ecosystems: An investigation of polylactic acid’s effects on Microcystis aeruginosa. Environ. Pollut. 2024, 344, 123347. [Google Scholar] [CrossRef] [PubMed]
- Hessler, C.M.; Wu, M.Y.; Xue, Z.; Choi, H.; Seo, Y. The influence of capsular extracellular polymeric substances on the interaction between TiO2 nanoparticles and planktonic bacteria. Water Res. 2012, 46, 4687–4696. [Google Scholar] [CrossRef] [PubMed]
- Planchon, M.; Jittawuttipoka, T.; Cassier-Chauvat, C.; Guyot, F.; Gelabert, A.; Benedetti, M.F.; Chauvat, F.; Spalla, O. Exopolysaccharides protect Synechocystis against the deleterious effects of Titanium dioxide nanoparticles in natural and artificial waters. J. Colloid Interface Sci. 2013, 405, 35–43. [Google Scholar] [CrossRef]
- Petersen, E.J.; Barrios, A.C.; Henry, T.B.; Johnson, M.E.; Koelmans, A.A.; Montoro Bustos, A.R.; Matheson, J.; Roesslein, M.; Zhao, J.; Xing, B. Potential artifacts and control experiments in toxicity tests of nanoplastic and microplastic particles. Environ. Sci. Technol. 2022, 56, 15192–15206. [Google Scholar] [CrossRef]
- Huang, W.; Zhao, T.; Zhu, X.; Ni, Z.; Guo, X.; Tan, L.; Wang, J. The effects and mechanisms of polystyrene and polymethyl methacrylate with different sizes and concentrations on Gymnodinium aeruginosum. Environ. Pollut. 2021, 287, 117626. [Google Scholar] [CrossRef]
- Besseling, E.; Wang, B.; Lürling, M.; Koelmans, A.A. Nanoplastic affects growth of S. obliquus and reproduction of D. magna. Environ. Sci. Technol. 2014, 48, 12336–12343. [Google Scholar] [CrossRef]
- Lechner, A.; Keckeis, H.; Lumesberger-Loisl, F.; Zens, B.; Krusch, R.; Tritthart, M.; Glas, M.; Schludermann, E. The Danube so colourful: A potpourri of plastic litter outnumbers fish larvae in Europe’s second largest river. Environ. Pollut. 2014, 188, 177–181. [Google Scholar] [CrossRef]
- Sjollema, S.B.; Redondo-Hasselerharm, P.; Leslie, H.A.; Kraak, M.H.S.; Vethaak, A.D. Do plastic particles affect microalgal photosynthesis and growth? Aquat. Toxicol. 2016, 170, 259–261. [Google Scholar] [CrossRef]
- Su, L.; Xue, Y.; Li, L.; Yang, D.; Kolandhasamy, P.; Li, D.; Shi, H. Microplastics in Taihu Lake, China. Environ. Pollut. 2016, 216, 711–719. [Google Scholar] [CrossRef]
- Yonkos, L.T.; Friedel, E.A.; Perez-Reyes, A.C.; Ghosal, S.; Arthur, C.D. Microplastics in four estuarine rivers in the Chesapeake Bay, U.S.A. Environ. Sci. Technol. 2014, 48, 14195–14202. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| NPs | EPS Condition | ODplateau | ODreduced | v | R2 | ΔODreduced | |
|---|---|---|---|---|---|---|---|
| PS | EPS-C | Sum | −0.076 | 1.076 | 0.037 | 0.998 | 0.74 |
| Mix | −0.818 | 1.817 | 0.033 | 0.998 | |||
| EPS-F | Sum | −0.227 | 1.227 | 0.041 | 0.998 | 0.53 | |
| Mix | −0.758 | 1.758 | 0.031 | 0.997 | |||
| PLA | EPS-C | Sum | −0.071 | 1.071 | 0.037 | 0.998 | 0.33 |
| Mix | −0.404 | 1.404 | 0.043 | 0.997 | |||
| EPS-F | Sum | −0.174 | 1.174 | 0.044 | 0.998 | 0.49 | |
| Mix | −0.669 | 1.669 | 0.033 | 0.991 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ding, J.; Yang, J.; Song, X.; Liu, S.; Wang, Z.; Zou, H. Hetero-Aggregation of Nanoplastics with Freshwater Algae and the Toxicological Consequences: The Role of Extracellular Polymeric Substances. Toxics 2025, 13, 980. https://doi.org/10.3390/toxics13110980
Ding J, Yang J, Song X, Liu S, Wang Z, Zou H. Hetero-Aggregation of Nanoplastics with Freshwater Algae and the Toxicological Consequences: The Role of Extracellular Polymeric Substances. Toxics. 2025; 13(11):980. https://doi.org/10.3390/toxics13110980
Chicago/Turabian StyleDing, Jiannan, Jiaxin Yang, Xiaojun Song, Shuo Liu, Zhenguo Wang, and Hua Zou. 2025. "Hetero-Aggregation of Nanoplastics with Freshwater Algae and the Toxicological Consequences: The Role of Extracellular Polymeric Substances" Toxics 13, no. 11: 980. https://doi.org/10.3390/toxics13110980
APA StyleDing, J., Yang, J., Song, X., Liu, S., Wang, Z., & Zou, H. (2025). Hetero-Aggregation of Nanoplastics with Freshwater Algae and the Toxicological Consequences: The Role of Extracellular Polymeric Substances. Toxics, 13(11), 980. https://doi.org/10.3390/toxics13110980