Immunohistochemical Study of Glucose Transporter GLUT-5 in Duodenal Epithelium in Norm and in T-2 Mycotoxicosis

 , ,
, ,
Abstract
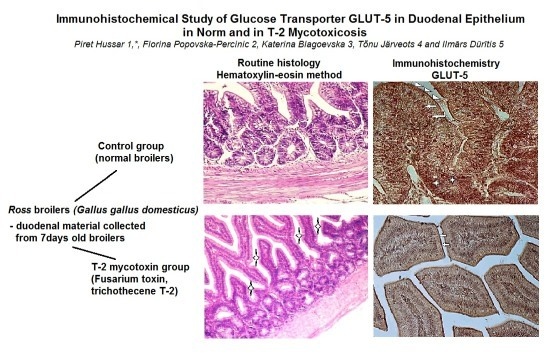
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
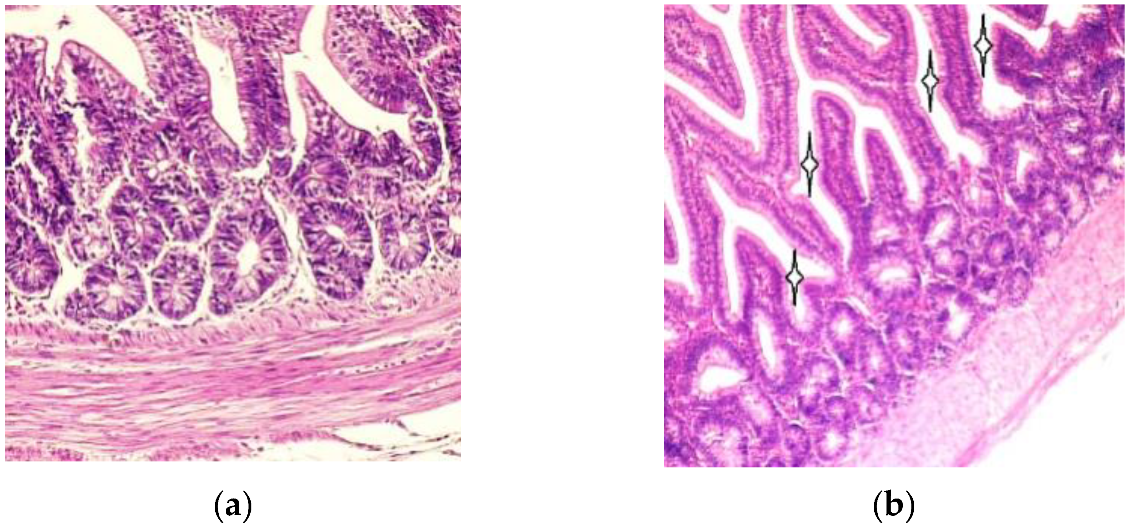
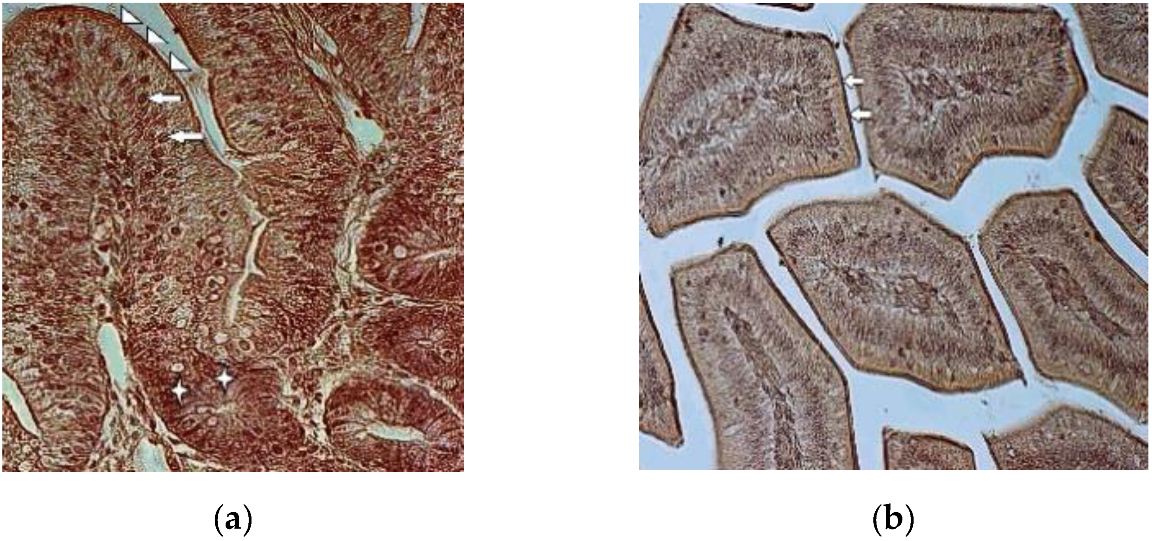
3. Results
3.1. Light Microscopy
3.2. Immunohistochemistry
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Stevens, L. Avian Biochemistry and Molecular Biology; Cambridge University Press: Cambridge, UK, 1996; pp. 29–45. [Google Scholar]
- Welch, K.C., Jr.; Allalou, A.; Sehgal, P.; Cheng, J.; Ashok, A. Glucose transporter expression in an avian nectarivore: The ruby-throated hummingbird (Archilochus colubris). PLoS ONE 2013, 8, e77003. [Google Scholar] [CrossRef] [PubMed]
- Takata, K. Glucose transporters in the transepithelial transport of glucose. J. Electron. Microsc. 1996, 45, 275–284. [Google Scholar] [CrossRef] [PubMed]
- Braun, E.J.; Sweazea, K.L. Glucose regulation in birds. Comp. Biochem. 2008, 151, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Wood, I.S.; Trayhurn, P. Glucose transporters (GLUT and SGLT): Expanded families of sugar transport proteins. Br. J. Nutr. 2003, 89, 3–9. [Google Scholar] [CrossRef] [PubMed]
- Mueckler, M.; Thorens, B. The SLC2 (GLUT) family of membrane transporters. Mol. Aspects Med. 2013, 34, 121–138. [Google Scholar] [CrossRef]
- Ferraris, R.P. Dietary and developmental regulation of intestinal sugar transport. Biochem. J. 2001, 360, 265–276. [Google Scholar] [CrossRef]
- Yoshikawa, T.; Inoue, R.; Matsumoto, M.; Yajima, T.; Ushida, K.; Iwanaga, T. Comparative expression of hexose transporters (SGLT1, GLUT1, GLUT2 and GLUT5) throughout the mouse gastrointestinal tract. Histochem. Cell Biol. 2011, 135, 183–194. [Google Scholar] [CrossRef]
- Mueckler, M. Facilitative glucose transporters. Eur. J. Biochem. 1994, 219, 713–725. [Google Scholar] [CrossRef]
- Augustin, R. The protein family of glucose transport facilitators: It’s not only about glucose after all. IUBMB Life 2010, 62, 315–333. [Google Scholar] [CrossRef]
- Thorens, B. Facilitated glucose transporters in epithelial cells. Annu. Rev. Physiol. 1993, 55, 591–608. [Google Scholar] [CrossRef]
- Kellett, G.L.; Brot-Laroche, E. Apical GLUT2. A major pathway of intestinal sugar absorption. Diabetes 2005, 54, 3056–3062. [Google Scholar] [CrossRef] [PubMed]
- Ang, B.R.G.; Yu, G.F. The role of fructose in type 2 diabetes and other metabolic diseases. J. Nutr. Food Sci. 2018, 8, 659. [Google Scholar]
- Gilbert, E.R.; Li, H.; Emmerson, D.A.; Webb, K.E., Jr.; Wong, E.A. Developmental regulation of nutrient transporter and enzyme mRNA abundance in the small intestine of broilers. Poult. Sci. 2007, 86, 1739–1753. [Google Scholar] [CrossRef] [PubMed]
- Wasserman, D.; Hoekstra, J.H.; Tolia, V.; Taylor, C.J.; Kirschner, B.S.; Takeda, J.; Bell, G.I.; Taub, R.; Rand, E.B. Molecular analysis of the fructose transporter gene (GLUT5) in isolated fructose malabsorption. J. Clin. Investig. 1996, 98, 2398–2402. [Google Scholar] [CrossRef]
- Lertanekawattana, S.; Wichatrong, T.; Chaisiri, K.; Uchikawa, R.; Arizono, N. Expression of cytokines and monosaccharide transporters in the duodenal mucosa of patients with gastrointestinal symptoms in rural Thailand. Southeast Asian J. Trop. Med. Public Health 2005, 36, 923–930. [Google Scholar]
- Kumagai, S.; Shimizu, T. Effects of Fusarenon-X and T-2 toxin on intestinal absorption of monosaccharide in rats. Arch. Toxicol. 1998, 61, 489–495. [Google Scholar] [CrossRef]
- Zhang, Y.; Han, Y.; Zhu, C.C.; Tang, F.; Cui, X.-S.; Kim, N.; Sun, S.-C. Exposure to HT-2 toxin causes oxidative stress induced apoptosis/autophagy in porcine oocytes. Sci. Rep. 2016, 6, 33904. [Google Scholar] [CrossRef]
- Sokolović, M.; Garaj-Vrhovac, V.; Šimpraga, B. T-2 toxin incidence and toxicity in poultry. Arh. Hig. Rada Toksikol. 2008, 59, 43–52. [Google Scholar] [CrossRef]
- Yuan, G.; Wang, Y.; Yuan, X.; Zhang, T.; Zhao, Y.; Huang, L.; Peng, S. T-2 toxin induces developmental toxicity and apoptosis in zebrafish embryos. J. Environ. Sci. 2014, 26, 917–925. [Google Scholar] [CrossRef]
- Kachuei, R.; Rezaie, S.; Hossein Yadegari, M.; Safaie, N.; Allameh, A.-A.; Aref-Poor, M.-A.; Imani Fooladi, A.-A.; Riazipour, M.; Mohammad Abadi, H.M. Determination of T-2 Mycotoxin in Fusarium strains by HPLC with fluorescence detector. J. Appl. Biotechnol. Rep. 2014, 1, 38–43. [Google Scholar]
- Carson, F.L. Histotechnology: A Self-Instructional Text, 2nd ed.; ASCP Press: Chicago, IL, USA, 1997. [Google Scholar]
- Nasrin, M.; Siddiqi, M.N.H.; Masum, M.A.; Wares, M.A. Gross and histological studies of digestive tract of broilers during postnatal growth and development. J. Bangladesh Agric. Univ. 2012, 10, 69–77. [Google Scholar] [CrossRef]
- Aitken, R.N.C. A histochemical study of the stomach and intestine of the chicken. J. Anat. 1958, 92, 453–466. [Google Scholar] [PubMed]
- Calhoun, M.L. Microscopic Anatomy of the Digestive System of the Chicken; Iowa State Colleage Press: Ames, IA, USA, 1954. [Google Scholar]
- Solcan, C.; Pavel, G.; Floristean, V.; Chiriac, I.; Şlencu, B.; Solcan, G. Effect of ochratoxin A on the intestinal mucosa and mucosa-associated lymphoid tissues in broiler chickens. Acta Vet. Hugarica 2015, 63, 30–40. [Google Scholar] [CrossRef] [PubMed]
- Tawfeek, F.A.; Hassan, R.A.; Eid, Y. Evaluation of antimycotoxin effects of humate and hydrated sodium calcium aluminosilicate on broilers toxicated with aflatoxin. Alex. J. Vet. Sci. 2017, 54, 45–56. [Google Scholar]
- Merigo, F.; Brandolese, A.; Facchin, S.; Missaggia, S.; Bernardi, P.; Boschi, F.; D’Inca, R.; Savarino, E.V.; Sbarbati, A.; Sturniolo, G.C. Glucose transporter expression in the human colon. World J. Gastroenterol. 2018, 24, 775–793. [Google Scholar] [CrossRef]
- Hussar, P.; Kärner, M.; Järveots, T.; Pendovski, L.; Dūrītis, I.; Popovska-Percinic, F. Comparative study of glucose transporters GLUT-2 and GLUT-5 in ostriches gastrointestinal tract. Maced. Vet. Rev. 2016, 39, 225–231. [Google Scholar] [CrossRef]
- Hussar, P.; Kärner, M.; Dūrītis, I.; Plivča, A.; Pendovski, L.; Järveots, T.; Popovska-Percinic, F. Temporospatial study of hexose transporters and mucin in the epithelial cells of chicken (Gallus gallus domesticus) small intestine. Pol. J. Vet. Sci. 2017, 20, 627–633. [Google Scholar]
- Dong, R.; Srai, S.K.; Debnam, E.; Smith, M. Transcriptional andtranslational control over sodium-glucose-linked transporter(SGLT1) gene expression in adult rat small intestine. FEBS Lett. 1997, 406, 79–82. [Google Scholar] [CrossRef]
- Kojima, T.; Nishimura, M.; Yajima, T.; Kuwata, T.; Suzuki, Y.; Goda, T.; Takase, S.; Harada, E. Developmental changes in theregional Na+/glucose transporter mRNA along the smallintestine of suckling rats. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 1998, 122, 89–95. [Google Scholar] [CrossRef]
- Burant, C.F.; Saxena, M. Rapid reversible substrate regulation of fructose transporter expression in rat small intestine and kidney. Am. J. Physiol. Gastrointest. Liver Physiol. 1994, 267, G71–G79. [Google Scholar] [CrossRef]
- Burant, C.F.; Takeda, J.; Brot-Laroche, E.; Bell, G.I.; Davidson, N.O. Fructose transporter in human spermatozoa and small intestine is GLUT5. J. Biol. Chem. 1992, 267, 14523–14526. [Google Scholar] [PubMed]
- Douard, V.; Ferraris, R.P. Regulation of the fructose transporter GLUT5 in health and disease. Am. J. Physiol. Endocrinol. Metab. 2008, 295, E227–E237. [Google Scholar] [CrossRef]
- Garcia-Herrera, J.; Abad, B.; Rodriguez-Yoldi, M.J. Effect of lipopolysaccharide on d-fructose transport across rabbit jejunum. Inflamm. Res. 2003, 52, 177–184. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Herrera, J.; Navarro, M.A.; Marca, M.C.; de la Osada, J.; Rodriguez-Yoldi, M.J. The effect of tumor necrosis factor-alpha on d-fructose intestinal transport in rabbits. Cytokine 2004, 25, 21–30. [Google Scholar] [CrossRef]
- Afsah-Hejri, L.; Jinap, S.; Hajeb, P.; Radu, S.; Shakibazadeh, S.H. A review on mycotoxins in food and feed: Malaysia case study. Compr. Rev. Food Sci. Food Saf. 2013, 12, 629–651. [Google Scholar] [CrossRef]
- Boonen, J.; Malysheva, S.V.; Taevernier, L.; Di Mavungu, J.D.; De Saeger, S.; De Spiegeleer, B. Human skin penetration of selected model mycotoxins. Toxicology 2012, 301, 21–32. [Google Scholar] [CrossRef] [PubMed]
- Caldwell, R.D. Yellow rain’ or natural toxins? Nature 1983, 301, 651. [Google Scholar] [CrossRef]
- Adegoke, G.O.; Letuma, P. Strategies for the prevention and reduction of mycotoxins in developing countries, Mycotoxin and Food Safety in Developing Countries. In Mycotoxin and Food Safety in Developing Countries; IntechOpen: Rijeka, Croatia, 2013; pp. 123–136. [Google Scholar]
- Marin, S.; Ramos, A.J.; Cano-Sancho, G.; Sanchis, V. Mycotoxins: Occurrence, toxicology, and exposure assessment. Food Chem. Toxicol. 2013, 60, 218–237. [Google Scholar] [CrossRef]
- Torp, M.; Langseth, W. Production of T-2 toxin by a Fusarium resembling Fusarium poae. Mycopathologia 1999, 147, 89–96. [Google Scholar] [CrossRef]
- Obremski, K.; Podlasz, P.; Żmigrodzka, M.; Winnicka, A.; Woźny, M.; Brzuzan, P.; Jakimiuk, E.; Wojtacha, P.; Gajęcka, M.; Zielonka, L.; et al. The effect of T-2 toxin on percentages of CD4+, CD8+, CD4+CD8+ and CD21+ lymphocytes, and mRNA expression levels of selected cytokines in porcine ileal Peyer’s patches. Pol. J. Vet. Sci. 2013, 16, 341–349. [Google Scholar] [CrossRef]
- Seeboth, R.; Solinhac, I.P.; Oswald, L.; Guzylack, P. The fungal T-2 toxin alters the activation of primary macrophages induced by TLR-agonists resulting in a decrease of the inflammatory response in the pig. Vet. Res. 2012, 43, 35. [Google Scholar] [CrossRef] [PubMed]
- Sweazea, K.L.; Braun, E.J. Glucose transporter expression in English sparrows (Passer domesticus). Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2006, 144, 263–270. [Google Scholar] [CrossRef]
- Wang, F.; Zuo, Z.; Chen, K.; Gao, C.; Yang, Z.; Zhao, S.; Li, J.; Song, H.; Peng, X.; Fang, J.; et al. Histopathological injuries, ultrastructural changes, and depressed TLR expression in the small intestine of broiler chickens with aflatoxin B1. Toxins 2018, 10, 131. [Google Scholar] [CrossRef] [PubMed]
- Phletus, P.W. Effects of T-2 mycotoxin on gastrointestinal tissues: A Review of in vivo and in vitro models. Arch. Environ. Contam. Toxicol. 1989, 18, 374–387. [Google Scholar]
- Suneja, S.K.; Ram, G.C.; Wagle, D.S. Effects of T-2 toxin on glucose and tryptophan uptake and intestinal mucosal enzymes. Toxicon 1984, 22, 39–43. [Google Scholar] [CrossRef]


© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hussar, P.; Popovska-Percinic, F.; Blagoevska, K.; Järveots, T.; Dūrītis, I. Immunohistochemical Study of Glucose Transporter GLUT-5 in Duodenal Epithelium in Norm and in T-2 Mycotoxicosis. Foods 2020, 9, 849. https://doi.org/10.3390/foods9070849
Hussar P, Popovska-Percinic F, Blagoevska K, Järveots T, Dūrītis I. Immunohistochemical Study of Glucose Transporter GLUT-5 in Duodenal Epithelium in Norm and in T-2 Mycotoxicosis. Foods. 2020; 9(7):849. https://doi.org/10.3390/foods9070849
Chicago/Turabian StyleHussar, Piret, Florina Popovska-Percinic, Katerina Blagoevska, Tõnu Järveots, and Ilmārs Dūrītis. 2020. "Immunohistochemical Study of Glucose Transporter GLUT-5 in Duodenal Epithelium in Norm and in T-2 Mycotoxicosis" Foods 9, no. 7: 849. https://doi.org/10.3390/foods9070849
APA StyleHussar, P., Popovska-Percinic, F., Blagoevska, K., Järveots, T., & Dūrītis, I. (2020). Immunohistochemical Study of Glucose Transporter GLUT-5 in Duodenal Epithelium in Norm and in T-2 Mycotoxicosis. Foods, 9(7), 849. https://doi.org/10.3390/foods9070849