The Relationship between Phytochemical Composition and Biological Activities of Differently Pigmented Varieties of Berry Fruits; Comparison between Embedded in Food Matrix and Isolated Anthocyanins


Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Plant Material
2.3. Plant Extract Preparation
2.4. Cell Culture
2.5. Composition of Phenolic Compounds
2.6. Profiles of Antioxidants and Antioxidant Capacity
2.7. Cytotoxic Activity towards HT29 Cells
2.8. Antigenotoxic Activity
2.9. Influence on Activity of Phase II Detoxification Enzymes
2.9.1. Preparation of Cytosolic Fractions
2.9.2. GST Activity
2.9.3. NQO1 Activity
2.10. Antidiabetic Activity
2.10.1. Inhibition of α-Amylase Activity
2.10.2. Inhibition of α-Glucosidase Activity
2.11. Statistical Analysis
3. Results
3.1. Composition of Phenolic Compounds
3.2. Antioxidant Capacity and Profiles of Antioxidants
3.3. Chemopreventive Potential of Tested Plant Extracts in Cellular Model
3.4. Inhibition of α-Amylase and α-Glucosidase in a Cell-Free Model
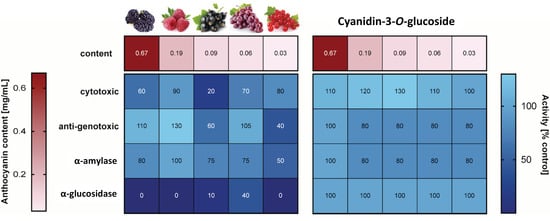
3.5. Chemopreventive Properties of Cy-3-O-Glu
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Rahman, T.; Hosen, I.; Islam, M.M.T.; Shekhar, I. Oxidative stress and human health. Adv. Biosci. Biotechnol. 2012, 3, 997–1019. [Google Scholar] [CrossRef]
- Miyazawa, T.; Nakagawa, K.; Kudo, M.; Muraishi, K.; Someya, K. Direct intestinal absorption of red fruit anthocyanins, cyanidin-3-glucoside and cyanidin-3, 5-diglucoside, into rats and humans. J. Agric. Food Chem. 1999, 47, 1083–1091. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.S.; Hecht, S.S.; Carmella, S.G.; Yu, N.; Larue, B.; Henry, C.; McIntyre, C.; Rocha, C.; Lechner, J.F.; Stoner, G.D. Anthocyanins in black raspberries prevent esophageal tumors in rats. Cancer Prev. Res. 2009, 2, 84–93. [Google Scholar] [CrossRef] [PubMed]
- Faria, A.; Pestana, D.; Teixeira, D.; de Freitas, V.; Mateus, N.; Calhau, C. Blueberry anthocyanins and pyruvic acid adducts: Anticancer properties in breast cancer cell lines. Phytother. Res. 2010, 24, 1862–1869. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Zhang, Y.; Liu, Y.; Sun, R.; Xia, N. Purified anthocyanin supplementation reduces dyslipidemia, enhances anti- oxidant capacity, and prevents insulin resistance in diabetic patients. J. Nutr. 2015, 145, 742–748. [Google Scholar] [CrossRef]
- Kwon, S.H.; Ahn, I.S.; Kim, S.O.; Kong, C.; Chung, H.; Do, M.; Park, K. Anti-obesity and hypolipidemic effects of black soybean anthocyanins. J. Med. Food 2007, 10, 552–556. [Google Scholar] [CrossRef]
- Cooke, D.; Steward, W.P.; Gescher, A.J.; Marczyło, T. Anthocyanins from fruits and vegetables—Does bright colour signal cancer chemopreventive activity? Eur. J. Cancer 2005, 41, 1931–1940. [Google Scholar] [CrossRef]
- Dudley, J.I.; Lekli, I.; Mukhrjee, S.; Das, M.; Bartelli, A.A.; Das, D.K. Does white wine qualify for French paradox? Comparison of the cardioprotective effects of red and white wines and their constituents: Resveratrol, tyrosol and hydroxytyrosol. J. Agric. Food Chem. 2008, 20, 9362–9373. [Google Scholar] [CrossRef]
- Koss-Mikołajczyk, I.; Kusznierewicz, B.; Wiczkowski, W.; Płatosz, N.; Bartoszek, A. Phytochemical composition and biological activities of differently pigmented cabbage (Brassica oleracea var. capitata) and cauliflower (Brassica oleracea var. botrytis) varieties. J. Sci. Food Agric. 2019, 99, 5499–5507. [Google Scholar]
- Määttä, K.; Kamal-Eldin, A.; Törrönen, R. Phenolic compounds in berries of black, red, green and white currants (Ribes sp.). Antioxid. Redox Signal. 2001, 3, 981–993. [Google Scholar] [CrossRef]
- Cejudo-Bastante, M.J.; Chaalal, M.; Louileche, H.; Parrado, J.; Heredia, F.J. Betalain profile, phenolic content, and color characterization of different parts and varieties of Opuntia ficus-indica. J. Agric. Food Chem. 2015, 62, 8491–8499. [Google Scholar] [CrossRef] [PubMed]
- Ercisli, S.; Orhan, E. Chemical composition of white (Morus alba), red (Morus rubra) and black (Morus nigra) mulberry fruits. Food Chem. 2007, 103, 1380–1384. [Google Scholar] [CrossRef]
- Matsufuji, H.; Ishikawa, K.; Nunomura, O.; Chino, M.; Takeda, M. Antioxidant content of different coloured sweet peppers, white, green, yellow, orange and red (Capsicum annum L.). Int. J. Food Sci. Technol. 2007, 42, 1482–1488. [Google Scholar] [CrossRef]
- Zhou, C.; Xu, C.; Sun, C.; Li, X.; Chen, K. Carotenoids in white and red-fleshed loquat fruits. J. Agric. Food Chem. 2007, 55, 7822–7830. [Google Scholar] [CrossRef] [PubMed]
- Sumaya-Martínez, M.T.; Cruz-Jaime, S.; Madrigal-Santillán, E.; García-Peredes, J.D.; Coriño-Cortés, R.; Cruz-Cansino, N.; Valadez-Vega, C.; Martinez-Cardenas, L.; Alanís-García, E. Betalain, ascorbic acid, phenolic contents and antioxidant properties of purple, red, yellow and white cactus pear. Int. J. Mol. Sci. 2011, 12, 6452–6468. [Google Scholar] [CrossRef]
- Sánchez-Salceo, E.M.; Mena, P.; García-Viguera, C.; Martínez, J.J.; Hernandéz, F. Phytochemical evaluation of white (Morus alba L.) and black (Morus nigra L.) mulberry fruits, a starting point of the assessment of their beneficial properties. J. Funct. Foods 2015, 12, 399–408. [Google Scholar] [CrossRef]
- Shon, M.; Choi, S.; Kahng, G.; Nam, S.; Sung, N. Antimutagenic, antioxidant and free radical scavenging activity of ethyl acetate extracts from white, yellow and red onions. Food Chem. Toxicol. 2004, 42, 659–666. [Google Scholar] [CrossRef]
- García-Lafuente, A.; Moro, C.; Manchón, N.; Gonzalo-Ruiz, A.; Villares, A.; Guillamón, E. In vitro anti-inflammatory activity of phenolic rich extracts from white and red common beans. Food Chem. 2014, 161, 216–223. [Google Scholar] [CrossRef]
- Koss-Mikołajczyk, I.; Kusznierewicz, B.; Wiczkowski, W.; Sawicki, T.; Bartoszek, A. The comparison of betalain composition and chosen biological activities for differently pigmented prickly pear (Opuntia ficus-indica) and beetroot (Beta vulgaris) varieties. Int. J. Food Sci. Nutr. 2019, 70, 442–452. [Google Scholar] [CrossRef]
- Kim, H.; Choi, H.; Moon, J.Y.; Kim, Y.S.; Mosaddik, A.; Cho, S.K. Comparative antioxidant and antiproliferative activities of red and white pitayas and their correlation with flavonoid and polyphenol content. J. Food Sci. 2011, 76, 38–45. [Google Scholar] [CrossRef]
- Kusznierewicz, B.; Piekarska, A.; Mrugalska, B.; Konieczka, P.; Namieśnik, J.; Bartoszek, A. Phenolic Composition and Antioxidant Properties of Polish Blue-Berried Honeysuckle Genotypes by HPLC-DAD-MS, HPLC Postcolumn Derivatization with ABTS or FC, and TLC with DPPH Visualization. J. Agric. Food Chem. 2012, 60, 1755–1760. [Google Scholar] [CrossRef] [PubMed]
- Kusznierewicz, B.; Piasek, A.; Bartoszek, A.; Namieśnik, J. Application of a commercially available derivatization instrument and commonly used reagents to HPLC on-line determination of antioxidants. J. Food Compos. Anal. 2011, 24, 1073–1080. [Google Scholar] [CrossRef]
- Koss-Mikołajczyk, I.; Pilipczuk, T.; Lewandowska, A.; Kusznierewicz, B.; Bartoszek, A. Composition of bioactive secondary metabolites and mutagenicity of elderberry (Sambucus nigra L.) fruits at different stages of ripeness. Acta Aliment. 2016, 45, 442–451. [Google Scholar] [CrossRef]
- Benzie, I.F.; Strain, J.J. The ferric reducing ability of plasma (FRAP) as a measure of “antioxidant power”: The FRAP assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef]
- Koss-Mikołajczyk, I.; Kusznierewicz, B.; Namieśnik, J.; Bartoszek, A. Juices from non-typical edible fruits as health-promoting acidity regulators for food industry. LWT Food Sci. Technol. 2015, 64, 845–852. [Google Scholar] [CrossRef]
- Koss-Mikołajczyk, I.; Baranowska, M.; Todorovic, V.; Albini, A.; Sansone, C.; Andreoletti, P.; Cherkaoui-Malki, M.; Lizard, G.; Noonan, D.; Sobajic, S.; et al. Prophylaxis of Non-communicable Diseases: Why Fruits and Vegetables may be Better Chemopreventive Agents than Dietary Supplements Based on Isolated Phytochemicals? Curr. Pharm. Des. 2019, 25, 1847–1860. [Google Scholar] [CrossRef]
- Piekarska, A.; Kołodziejski, D.; Pilipczuk, T.; Bodnar, M.; Konieczka, P.; Kusznierewicz, B.; Hanschen, F.S.; Schreiner, M.; Cyprys, J.; Groszewska, M.; et al. The influence of selenium addition during germination of Brassica seeds on health-promoting potential of sprouts. Int. J. Food Sci. Nutr. 2014, 65, 692–702. [Google Scholar] [CrossRef]
- Zia-Ul-Haq, M.; Ćavar, S.; Qayum, M.; Imran, I.; De Feo, V. Compositional studies: Antioxidant and antidiabetic activities of Capparis decidua (Forsk.) Edgew. Int. J. Mol. Sci. 2011, 12, 8846–8861. [Google Scholar] [CrossRef]
- Jia, N.; Xiongm, Y.; Kong, B.; Liu, Q.; Xia, X. Radical scavenging activity of black currant (Ribes nigrum L.) extract and its inhibitory effect on gastric cancer cell proliferation via induction of apoptosis. J. Funct. Foods 2012, 4, 382–390. [Google Scholar] [CrossRef]
- Rubinskiene, M.; Viskelis, P.; Jasutiene, I.; Viskeliene, R.; Bobinas, C. Impact of various factors on the composition and stability of black currant anthocyanins. Food Res. Int. 2005, 38, 867–871. [Google Scholar] [CrossRef]
- Borges, G.; Degeneve, A.; Mullen, W.; Crozier, A. Impact of various factors on the composition and stability of black currant anthocyanins. Food Res. Int. 2010, 58, 3901–3909. [Google Scholar]
- Katsube, N.; Iwashita, K.; Tsashida, T.; Yamaki, K.; Kobori, M. Induction of apoptosis in cancer cells by bilberry (Vaccinium myrtillus) and the anthocyanins. J. Agric. Food Chem. 2003, 51, 68–75. [Google Scholar] [CrossRef] [PubMed]
- Olsson, M.E.; Gustavsson, K.; Andersson, S.; Nilsson, A.; Duan, R. Inhibition of cancer cell proliferation in vitro by fruit and berry extracts and correlations with antioxidant levels. J. Agric. Food Chem. 2004, 52, 7264–7271. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.K.; Koponen, J.M.; Mykkänen, H.M.; Törrönen, A.R. Berry phenolic extracts modulate the expression of p21WAF1 and Bax but not Bcl-2 in HT-29 colon cancer cells. J. Agric. Food Chem. 2007, 55, 1156–1163. [Google Scholar] [CrossRef]
- Bishayee, A.; Háznagy-Radnai, E.; Mbimba, T.; Sipos, P.; Morazzoni, P.; Darvesh, A.S.; Bhatia, D.; Hohmann, J. Anthocyanin-rich black currant extract suppresses the growth of human hepatocellular carcinoma cells. Nat. Prod. Commun. 2010, 5, 1613–1618. [Google Scholar] [CrossRef]
- Moon, Y.J.; Wang, X.; Morris, M.E. Dietary flavonoids: Effects on xenobiotic and carcinogen metabolism. Toxicol. In Vitro 2006, 20, 187–210. [Google Scholar] [CrossRef]
- Shih, P.H.; Yeh, C.T.; Yen, G.C. Anthocyanins induce theactivation of phase II enzymes through the antioxidantresponse element pathway against oxidative stress inducedapoptosis. J. Agric. Food Chem. 2007, 55, 9427–9435. [Google Scholar] [CrossRef]
- Soriano Sancho, R.A.; Pastore, G.M. Evaluation of the effects of anthocyanins in type 2 diabetes. Food Res. Int. 2012, 46, 378–386. [Google Scholar] [CrossRef]
- McDougal, G.J.; Shpiro, F.; Dobson, P.; Smith, P.; Blake, A.; Stewart, D. Different polyphenolic components of soft fruits inhibit α-amylase and α-glucosidase. J. Agric. Food Chem. 2005, 53, 2760–2766. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| a Peak No | Compounds (mg/g d.w.) | YRas | RRas | WGra | RGra | WMul | BMul | WCur | RCur | BCur |
|---|---|---|---|---|---|---|---|---|---|---|
| 1–4, 6–10, 13, 18, 20, 22, 25, 28, 41–42, 54, 62, 65, 69, 71 | Σ Hydroxybenzoic acid derivatives | 0.976 ± 0.033 | 1.078 ± 0.153 | 0.911 ± 0.035 | 0.346 ± 0.083 | 0 | 2.872 ± 0.223 | 0.895 ± 0.023 | 0.968 ± 0.048 | 0.478 ± 0.064 |
| 5, 11, 14–15, 17, 21, 23–24, 26 | Σ Hydroxycinnamic acid derivatives | 1.978 ± 0.083 | 6.009 ± 0.646 | 0.861 ± 0.022 | 0.203 ± 0.040 | 0 | 1.744 ± 0.157 | 0.465 ± 0.101 | 0.791 ± 0.085 | 0.650 ± 0.077 |
| 12, 16, 19, 27 | Flavan-3-ols | |||||||||
| 12 | Catechin | nd | 0.738 ± 0.045 | nd | 0.278 ± 0.074 | 0.177 ± 0.005 | 1.888 ± 0.161 | nd | nd | nd |
| 27 | Epicatechin | nd | nd | nd | 0.642 ± 0.096 | nd | 2.836 ± 0.282 | nd | nd | nd |
| Σ Flavan-3-ols | 0 | 0.738 ± 0.045 | 0 | 2.008 ± 0.085 | 1.888 ± 0.161 | 8.465 ± 0.826 | 0 | 0 | 0 | |
| 38, 40, 44, 47–51, 55–57, 59–60, 64, 67 | Flavonols | |||||||||
| 60 | Quercetin | nd | nd | nd | nd | nd | 0.195 ± 0.003 | nd | nd | nd |
| 56 | Rutin | nd | nd | 0.134 ± 0.006 | nd | nd | 0.201 ± 0.005 | 0.111 ± 0.007 | 0.180 ± 0.024 | 0.065 ± 0.006 |
| Σ Flavonols | 0 | 0.193 ± 0.015 | 0.274 ± 0.010 | 0.259 ± 0.009 | 0 | 1.487 ± 0.156 | 0.181 ± 0.005 | 0.359 ± 0.031 | 0.833 ± 0.132 | |
| Anthocyanins | ||||||||||
| 29 | Cy-3-O-Soph | nd | 1.970 ± 0.045 | nd | nd | nd | 5.686 ± 0.609 | nd | nd | nd |
| 30 | Delph-3-O-Glu | nd | nd | nd | 0.216 ± 0.016 | nd | nd | nd | nd | 0.315 ± 0.049 |
| 31 | Cy-3-O-(Glu-Rut) | nd | 0.507 ± 0.021 | nd | nd | nd | 0.521 ± 0.063 | nd | nd | nd |
| 32 | Delph-3-O-Rut | nd | nd | nd | nd | nd | nd | nd | nd | 0.569 ± 0.068 |
| 33 | Cy-3-O-Samb | nd | nd | nd | nd | nd | nd | nd | 0.035 ± 0.001 | nd |
| 34 | Cy-3-O-(Xyl-Rut) | nd | nd | nd | nd | nd | nd | nd | 0.314 ± 0.018 | nd |
| 35 | Cy-3-O-Glu | nd | 0.458 ± 0.005 | nd | 0.073 ± 0.003 | nd | 4.636 ± 0.586 | nd | nd | 0.161 ± 0.024 |
| 36 | Cy-3-O-Rut | nd | 0.206 ± 0.003 | nd | nd | nd | 0.111 ± 0.018 | nd | 0.077 ± 0.004 | 0.483 ± 0.086 |
| 37 | Pet-3-O-Glu | nd | nd | nd | 0.205 ± 0.012 | nd | nd | nd | nd | nd |
| 39 | Pel-3-O-Hex | nd | nd | nd | nd | nd | 0.381 ± 0.012 | nd | nd | nd |
| 43 | Peo-3-O-Glu | nd | nd | nd | 0.011 ± 0.001 | nd | nd | nd | nd | nd |
| 45 | Malv-3-O-Glu | nd | nd | nd | 0.142 ± 0.007 | nd | nd | nd | nd | nd |
| 46 | Cy-3-O-(Mal-diGlu) | nd | nd | nd | nd | nd | 0.111 ± 0.002 | nd | nd | nd |
| 52 | Delph-3-O-(6-O-acetyl)-Glu | nd | nd | nd | 0.033 ± 0.003 | nd | nd | nd | nd | nd |
| 53 | Delph-3-O-(6-O-p-Coum)-5-diGlu | nd | nd | nd | 0.035 ± 0.002 | nd | nd | nd | nd | nd |
| 58 | Pet-3-O-(6-O-p-Coum)-5-diGlu | nd | nd | nd | 0.046 ± 0.003 | nd | nd | nd | nd | nd |
| 61 | Pet-3-O-(6-O-acetyl)-Glu | nd | nd | nd | 0.014 ± 0.001 | nd | nd | nd | nd | nd |
| 63 | Malv-3-O-(6-O-p-Coum)-5-diGlu | nd | nd | nd | 0.025 ± 0.001 | nd | nd | nd | nd | nd |
| 66 | Malv-3-O-(6-O-acetyl)-Glu | nd | nd | nd | 0.061 ± 0.007 | nd | nd | nd | nd | nd |
| 68 | Delph-3-O-(6-O-p-Coum)-Glu | nd | nd | nd | 0.015 ± 0.002 | nd | nd | nd | nd | nd |
| 70 | Peo-3-O-Rut | nd | nd | nd | nd | nd | nd | nd | nd | 0.102 ± 0.003 |
| 73 | Pet- 3-O-(6-O-p-Coum)-Glu | nd | nd | nd | 0.056 ± 0.003 | nd | nd | nd | nd | nd |
| 74 | Malv-3-O-(6-O-p-Coum)-Glu | nd | nd | nd | 0.031 ± 0.002 | nd | nd | nd | nd | nd |
| Σ Anthocyanins | 0 | 3.142 ± 0.074 | 0 | 0.962 ± 0.064 | 0 | 11.708 ± 1.329 | 0 | 0.425 ± 0.023 | 1.631 ± 0.105 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Koss-Mikołajczyk, I.; Kusznierewicz, B.; Bartoszek, A. The Relationship between Phytochemical Composition and Biological Activities of Differently Pigmented Varieties of Berry Fruits; Comparison between Embedded in Food Matrix and Isolated Anthocyanins. Foods 2019, 8, 646. https://doi.org/10.3390/foods8120646
Koss-Mikołajczyk I, Kusznierewicz B, Bartoszek A. The Relationship between Phytochemical Composition and Biological Activities of Differently Pigmented Varieties of Berry Fruits; Comparison between Embedded in Food Matrix and Isolated Anthocyanins. Foods. 2019; 8(12):646. https://doi.org/10.3390/foods8120646
Chicago/Turabian StyleKoss-Mikołajczyk, Izabela, Barbara Kusznierewicz, and Agnieszka Bartoszek. 2019. "The Relationship between Phytochemical Composition and Biological Activities of Differently Pigmented Varieties of Berry Fruits; Comparison between Embedded in Food Matrix and Isolated Anthocyanins" Foods 8, no. 12: 646. https://doi.org/10.3390/foods8120646
APA StyleKoss-Mikołajczyk, I., Kusznierewicz, B., & Bartoszek, A. (2019). The Relationship between Phytochemical Composition and Biological Activities of Differently Pigmented Varieties of Berry Fruits; Comparison between Embedded in Food Matrix and Isolated Anthocyanins. Foods, 8(12), 646. https://doi.org/10.3390/foods8120646