1. Introduction
Since ancient times down to the present, wine has been prepared via biochemical transformation of grape juice, known as controlled alcoholic fermentation. Most of the enzymes necessary for the fermentation to occur originate from the grapes themselves, the grapes’ own microflora and the microorganisms present during the process [
1,
2]. Due to the ever-growing increase in the production levels of modern winemaking, these endogenous enzymes of grapes, yeasts and other microorganisms that are present naturally in musts and wines are often neither efficient enough, nor sufficient to effectively catalyze the corresponding reactions. For this reason, the use of commercial enzymes as supplements at different stages of the winemaking process is a well-established practice in the large-scale production of wine. Nowadays, the big production volumes combined with the very demanding quality control of the final product have transformed modern winemaking into a high-tech industry that makes use of the latest achievements in biology and enzymology [
3,
4].
Wine production consists of four main stages [
2]: (i) pressing and maceration with the purpose of extracting as much grape juice as possible for must formation; (ii) alcoholic fermentation, forming the alcohol content and many other specific features of wine; (iii) clarification, i.e., reducing the turbidity so as to achieve a clear, visually appealing and stable wine; (iv) aging and stabilization operations to optimize the physicochemical properties of the final product. All these stages play crucial roles in winemaking and their relative importance can vary depending on the style of wine being produced.
Exogenous pectinases are always needed for the clarification step as they are not produced by the yeasts used in fermentation [
2]. Pectinases (E.C.3.2.1.15) are a heterogeneous enzyme group used for controlled degradation of the high-molecular-weight pectic substances contained in grape cell walls that transit into must. The main purpose of the clarification step is to reduce the must viscosity and the content of insoluble fractions in it, as well as to intensify the grape juice extraction process and maximize its yield. There are indications that pectinases can also increase the production and retention of certain volatile substances, thus releasing in wine more of the color- and flavor-determining compounds contained in grape skin [
5], and making more effective the liberation of phenolic compounds [
6].
Pectin is a complex substance representing methylated esters of polygalacturonic acid. Therefore, based on their mode of action, pectinases are classified as (i) deesterifying enzymes (pectin esterase) and (ii) depolymerizing enzymes (hydrolases, lyases). Depolymerizing enzymes are further classified based on their specific action site into endo- and exo-polygalacturonases and rhamnogalacturonases [
7], or by the nature of the action mechanism, primary substrate and products obtained [
8]. Commercial pectinase preparations should contain enzymes from each of these groups.
Apart from winemaking, pectinases are applied on a large scale in other industrial processes such as fruit juice clarification and viscosity reduction, in tomato pulp extraction, chocolate and tea fermentation, vegetable waste treatment and fiber degumming in textile and paper industries, among others. Due to their universal applications in various branches of the food industry, pectinases have a share of 25% in the global sales of food enzymes [
9].
At the present time, industrial enological pectinases are most frequently used as free enzymes, i.e., soluble in the reaction medium. In this form, however, they are often unable to meet the necessary short-term operational stability and are very hard to recover and reuse [
10]. Moreover, enzymatic processes with free enzymes could be difficult to make continuous and automatize [
11]. To overcome these limitations, immobilization of the enzymatic catalysts can be performed. The exhaustive review by Ottone et al. [
2] and other studies [
12] discuss innumerous reports on diverse immobilized enzymes used at various stages of wine production. In general, immobilization of enzymes can be made on different supports (carriers) including natural or synthetic polymers, as well as inorganic substances. Physical methods (adsorption, entrapment) or chemical reactions (covalent binding to insoluble carriers, creation of crosslinked enzyme aggregates) can be implemented during immobilization. This leads to increased enzyme stability, to a final product free of biocatalysts and, finally, to the possibility of multiple uses of one and the same enzyme preparation.
In general, supports for the immobilization of enzymes that can be applied in the food industry must be nontoxic, biocompatible, insoluble under reaction conditions and mechanically and thermally stable [
13]. For the particular case of pectinases’ immobilization, a group of natural biopolymers capable of easy gelation and known as alginates seem to be quite useful as supports. For example, de Oliveira et al. [
14] demonstrated that enological pectinase extracted from
Aspergillus aculeatus and immobilized in calcium alginate beads via entrapment can result in a multiple-use and thermally stable enzyme preparation applicable for clarification of fruit juices. A commercial enological pectinase (Extrazyme
® by Épernay, France) entrapped in alginate hydrogels [
10] showed that the clarification biocatalysts so produced kept its original activity after 8 months of storage. The activity of the entrapped pectinase was retained after six reaction cycles, with 37% residual activity. Grape must turbidity decreased rapidly in the presence of this immobilized pectinase, which was more effective than the free enzyme. Immobilization of the Extrazyme
® enological preparation via entrapment in alginate hydrogel reinforced mechanically by addition of agar-agar was reported as a possible way to effectively and mechanically create a more resistant biocatalyst for wine clarification [
15]. The pectinolytic activity of the entrapped enzyme was appropriate and it was retained after six reaction cycles, with 61% residual activity.
The search for mechanically more stable and cheaper supports readily available in various forms (granules, powders, threads, etc.) led to immobilization on synthetic polymers. Special attention was given to polyamide 6 (PA6), which is probably the closest synthetic analogue of protein biomolecules and can be obtained via ring-opening polymerization. Thus, Omelková et al. [
16] and Rexová-Benková et al. [
17] disclosed PA6 powders as supports of endo-polygalacturonase immobilized via covalent binding by means of glutaraldehyde. The enzyme activity decreased from 1.35 µkat/mg to 0.67 µkat/mg after immobilization, whereby the sample that was not activated by glutaraldehyde did not show any enzyme activity [
16]. The changes in the action pattern of the covalently immobilized endo-polygalacturonase were ascribed to steric hindrance resulting from new covalent bonds in the enzyme molecule in the proximity of its active site [
17]. More recently, Shukla et al. [
18] covalently immobilized polygalacturonase from
Aspergillus Niger on PA6 beads using a similar glutaraldehyde activation. This immobilization yielded a protein loading of 70 μg/g of PA6. The immobilized enzyme showed maximum activity at 50 °C and pH 5.0 and could be reused through four cycles of apple juice clarification with almost 50% retention of its original activity. Ben-Othman and Rinken [
19] studied the immobilization of pectinolytic enzyme preparations on polyamide 66 (PA66) granules or threads applying covalent immobilization by means of dimethyl sulfate or glutaraldehyde. The immobilization yield on threads was over 40 times higher than that on pellets. The activity of the immobilized pectinase preparation for clarification of apple juice was similar to that of the free enzyme in the temperature range of 10–30 °C. The immobilized pectinase exhibited good reusability, retaining 40% of its initial activity after five successive cycles and more than 20% after twenty successive cycles. PA6 and PA66 pellets were also used by other authors as supports for pectolytic enzymes [
20,
21]. Covalent binding was applied in both cases through activation by dimethyl sulfate or glutaraldehyde.
There exist a number of studies on the immobilization of pectolytic enzymes on inorganic supports. Among them, the magnetic sensitive substances permitting complete and fast removal of the biocatalyst via magnetic separation possess good potential for industrial application. For example, Fang et al. [
22] prepared Fe
3O
4 magnetic nanoparticles coated with amorphous SiO
2 that were functionalized with NH
2 groups. They were used as a support for covalent immobilization of pectinase with glutaraldehyde as a coupling agent. The resulting biocatalyst retained 64% of its starting activity after seven consecutive uses, and only about 21% of the total activity was lost after a 30-day storage.
Summarizing the above, the advantages of the immobilized pectinolytic enzymes are clear; however, every type of support has its own advantages and disadvantages. The polyamide supports studied so far are mechanically more robust than alginates, being also biocompatible and readily available, but they seem to require covalent immobilization. The latter, as a rule, decreases the enzyme activity and needs activation by toxic compounds, which is not acceptable in the food industry. These two problems may be resolved via effective noncovalent immobilization of the pectinases on polyamide microparticles, creating conditions for multiple hydrogen bond formations between the protein-based enzymes and a highly porous, scaffoldlike support. As shown in our previous works, such porous PA6 microparticulate supports that could also have magnetic susceptibility can be produced via activated anionic ring-opening polymerization (AAROP) of lactams [
23]. The PA6 microparticles possess a suitable morphology, and their effectiveness as carriers for noncovalent immobilization of a single enzyme [
24] or of enzyme dyads [
25] has been demonstrated and the biocatalytic properties of these systems have been studied.
The main objective of this study was to perform noncovalent immobilization of a commercial pectinolytic preparation on PA6 microparticles with or without magnetic properties obtained via AAROP. The resulting pectinase complexes (Pec@PA6) were subjected to comparative activity and kinetic studies with the free enzyme preparation and applied for clarification of industrial rosé must. Their reusability was also investigated.
2. Materials and Methods
2.1. Materials
The ε-caprolactam (ECL) monomer with reduced moisture content for anionic polymerization (AP-Nylon®) was purchased from Brüggemann Chemical (Heilbronn, Germany). Before use, it was kept under vacuum for 1 h at 23 °C. As a polymerization activator, Brüggolen C20® (C20) from the same company was employed. The initiator sodium dicaprolactamato-bis-(2-methoxy-ethoxo)-aluminate (Dilactamate®, DL, 85% solution in toluene) was purchased from Katchem (Prague, Czech Republic) and applied without further treatment. The soft, non-insulated iron particles (Fe content > 99.8%), with average diameters of 3–5 µm, were kindly donated by the manufacturer BASF, Ludwigshafen, Germany. The enological pectinase preparation Viazym Clarif Extrem® derived from Aspergillus Niger and used for industrial clarification of white and rosé musts was a product of Martin Vialatte (Magenta, France). This pectinase preparation and sample amounts of industrial rosé musts were kindly donated by Sogrape Vinhos SA (Avintes, Portugal). The pectin from citrus peel and all other simple chemical reagents and solvents employed in this study were of analytical grade and were supplied by Sigma-Aldrich-Merck (Lisbon, Portugal).
2.2. Characterization Methods
Scanning electron microscopy (SEM) studies were performed with a NanoSEM-200 apparatus from FEI Nova (Hillsboro, OR, USA) using mixed secondary electron/backscattered electron in-lens detection. All the samples were observed after sputter coating with Au/Pd alloy with a 208 HR instrument from Cressington Scientific Instruments (Watford, UK) with high-resolution thickness control. The UV-VIS spectral measurements were carried out with a Shimadzu model 1900i double-beam spectrophotometer (Tokyo, Japan) working in photometric, spectral or kinetic modes. The turbidity measurements were made with a METRIA M10 portable turbidimeter (Labbox Labware, Barcelona, Spain) according to ISO 7027 performing up to 5 parallel measurements with every sample.
2.3. Synthesis of PA6 Microparticulate Supports
The PA6 microparticles (PA6 MPs) were synthesized via a proper method based on AAROP as previously described by Dencheva et al. [
23]. First, 0.3 mol of ECL was added to 100 mL of a mixed hydrocarbon solvent (toluene/xylene 1:1 by volume) while being stirred under a nitrogen atmosphere, and the reaction mixture was refluxed for 10–15 min. Subsequently, 3.0 mol% of DL and 1.5 mol% of C20 were introduced simultaneously. The reaction time was 1 h from the point of the catalytic system addition, and the temperature was maintained in the 125–135 °C range with constant stirring at about 800 rpm. The final MPs were formed as a fine powder that was separated from the reaction mixture via hot vacuum filtration, washed several times with methanol and dried for 30 min in a vacuum oven at 60 °C. To remove the low-molecular-weight PA6 fractions, further Soxhlet extraction for 4 h with methanol was employed. The resulting neat PA6 MPs were kept in a desiccator for further treatment. The scheme of PA6 MP synthesis via AAROP is presented in
Figure S1 of the Supplementary Materials. Two types of microparticle supports were synthesized, namely neat PA6 MPs and PA6-Fe MPs. In the latter case, 3 wt.% of Fe particles (in relation to the ECL monomer) was added to the ECL/C20/DL/solvent reaction mixture, all other polymerization, isolation and purification procedures being the same as in neat PA6 MPs.
2.4. Pectinase Immobilization via Physical Adsorption
The Viazym manufacturer recommends a 10-fold dilution of the preparation for industrial application. For this reason, PA6 MP or PA6-Fe MP supports (100 mg) were immobilized using 5 mL of a 10-fold diluted Viazym pectinase. This solution that shows a clear absorption maximum at 263 nm (see
Figure S2 of the Supplementary Materials) was used for the subsequent protein quantifications. It was assumed that the initial 10-fold diluted Vyazim contains 10 arbitrary units (a.u.) of protein in 1 mL. The PA6 and PA6-Fe MPs were incubated at two different temperatures (23 °C or 4 °C) under gentle agitation for 24 h. Thereafter, the supernatant was decanted and stored for further analyses and use. The final Pec@PA6 and Pec@PA6-Fe complexes were washed two times with double-distilled water to remove the non-immobilized enzymes and were stored at 4 °C for further analyses.
2.5. Determination of the Total Amount of Protein
After isolation of Pec@PA6 and Pec@PA6-Fe at the end of the physical immobilization, the respective supernatants were subjected to UV analysis to determine the residual protein in them and calculate on this basis the amount of the total protein (TP) incorporated into the pectinase-containing complexes, expressed as
where
is the starting protein content before immobilization, and
is the protein content in the resultant supernatant after immobilization. The latter was determined via direct quantification of the UV absorption peak at λ ≈ 263 nm.
2.6. Pectinase Activity Assay
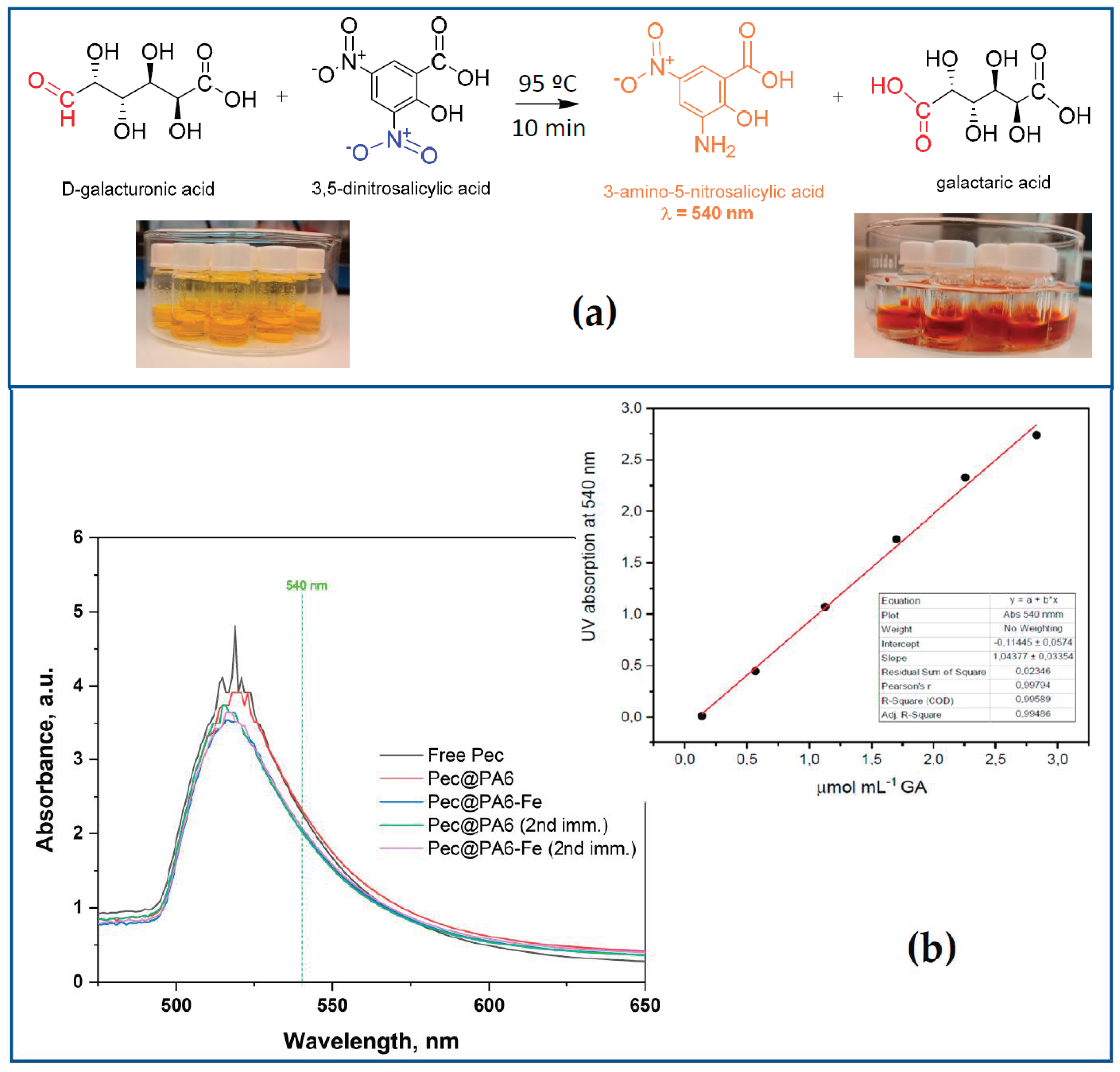
Pectinolytic activity was assayed by measuring the amount of reducing sugars released from a pectin solution in water using the DNS reagent according to the method described by Miller [
26]. The DNS reagent is composed of 2-hydroxy-3,5-dinitrobenzoic acid (1%
w/v), potassium sodium tartrate (30%
w/v) and sodium hydroxide (1%
w/v). In a typical assay, 0.05 mL of the commercial pectinolytic preparation (10-fold dilution) or 0.010 g of the wet pectinase-immobilized complexes were mixed with 0.45 mL substrate (0.25%
w/v citric pectin in 50 mM citrate buffer, pH 3.8) and incubated at 50 °C for 15 min. Subsequently, 0.50 mL of DNS solution was added to the reaction mixture, and the vials with the samples were kept in water at 90 °C for 10 min and then cooled on ice. The UV absorbance of the dark yellow solution was measured at 540 nm against the neat DNS reagent [
10]. The amount of released reducing sugar was quantified from a standard calibration plot constructed with D-galacturonic acid in the 0.14–2.23 µmol/mL range. One unit of pectinase activity is defined as the amount of enzyme required to release 1 μmol of galacturonic acid per minute under the assay conditions.
2.7. Kinetic Studies
The kinetic experiments were performed with the free commercial Viazym preparation (10-fold diluted) and its two immobilized complexes Pec@PA6 and Pec@PA6-Fe by varying the initial pectin concentration in the 0.05–5 mg/mL range and measuring the initial rates (i.e., enzyme activity) as indicated above. The total amount of enzyme added to the reaction mixture was maintained constant. Certain deviations from the conventional Michaelis–Menten kinetics were observed with the free and immobilized enzymatic complexes, i.e., substrate inhibition was present. Based on previous studies, three kinetic models, i.e., of Michaelis–Menten and two reflecting the substrate inhibition, were selected for calculation of the kinetic parameters. The functions of these models are presented in Equations (2)–(4).
The kinetic parameters (maximal rate Vmax, Michaelis–Menten constant Km and Ki, which is a constant characterizing the formation of an inactive enzyme–substrate complex) were estimated via nonlinear multiple regression analysis using the commercial package OriginPro, version 9.8.0.200 by OriginLab corporation (Northampton, MA, USA).
2.8. Application of Immobilized Pectinase for Raw Grape Must Clarification
The industrially recommended concentration of free pectinase preparation or the equivalent concentrations of the Pec@PA6 or Pec@PA6-Fe complexes were mixed with 14 mL of rosé must. The turbidity of the must samples was determined in nephelometric turbidity units (NTUs) every 15 min for 3–5 h. After clarification completion, the presence of residual pectin was tested by adding acidified alcohol (5% v/v HCl, 5 mL) to 2.5 mL of the must sample under gentle agitation. After 10 min, the must was visually inspected. In the case of pectin-free samples, clear liquid was observed. The formation of insoluble flakes indicated the presence of residual pectin.
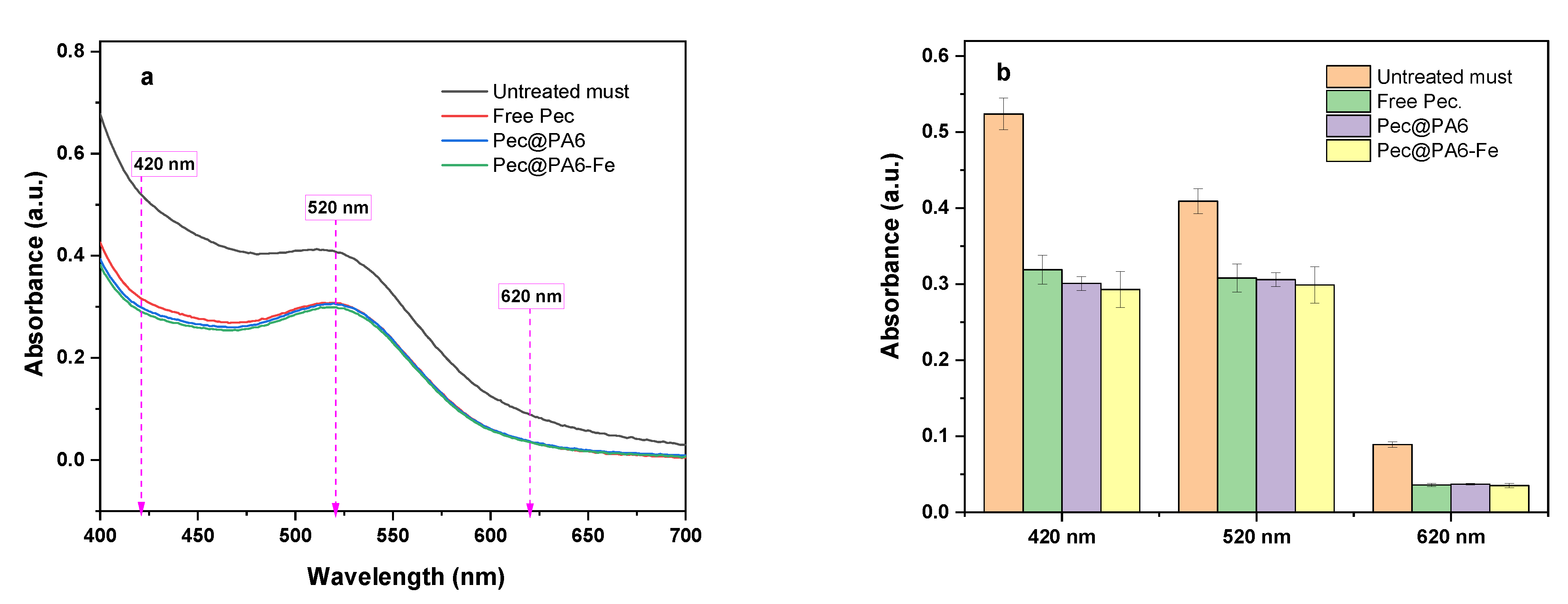
To evaluate the color changes in the must as a result of the enzymatic clarification, complete UV/VIS spectra of all treated must samples before and after the clarification were obtained, with their absorbances compared at 420, 520 and 620 nm.
2.9. Reusability Studies
The reusability of the Pec@PA6 or Pec@PA6-Fe complexes was tested by performing three consecutive clarification cycles with industrial rosé must at 23 °C, measuring turbidity every 15 min until reaching levels around 20 NTUs, according to
Section 2.8. After the first clarification cycle, the two complexes were recovered from the must via centrifugation and washed two times with 1 mL of deionized water, and their clarification capacity was tested with fresh rosé must, applying the same procedure.
4. Conclusions
This is the first study on noncovalent immobilization of commercial pectinolytic preparations (Viazym) upon specially synthesized microparticulate porous PA6-based supports via AAROP.
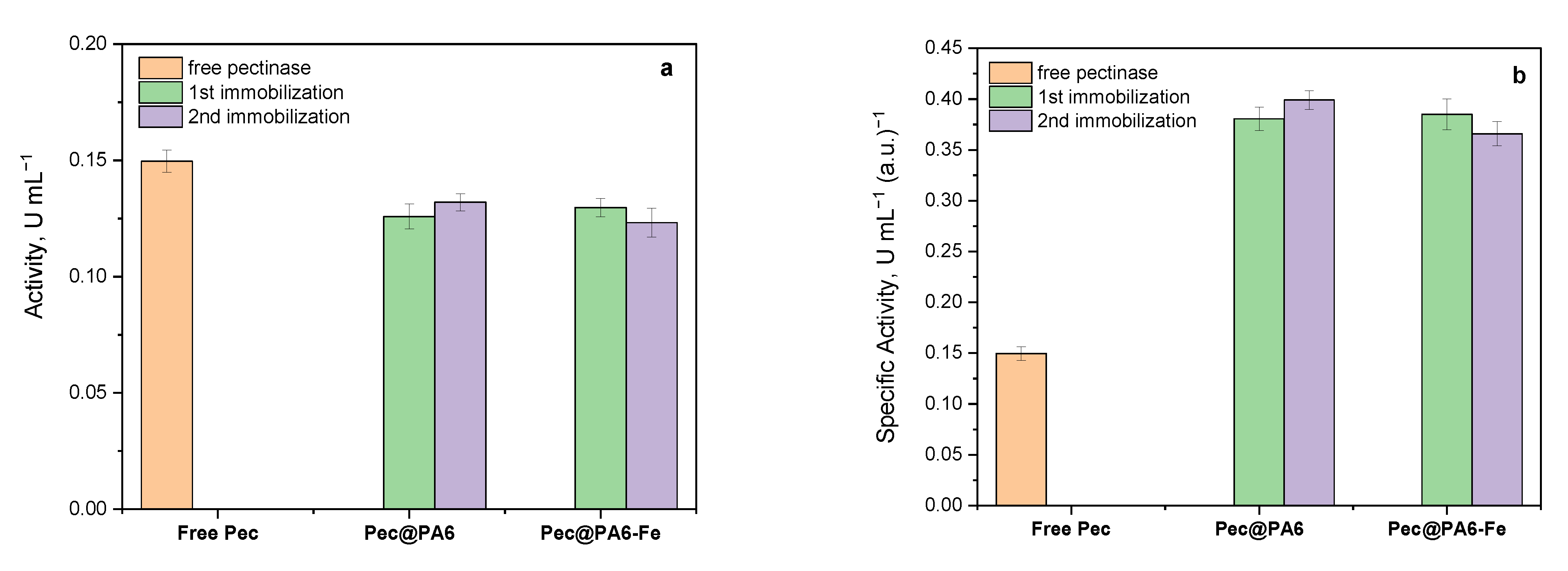
The pectinase immobilization suggested in this work is a very simple one-pot process occurring through incubation of the microparticulate supports in buffered aqueous enzyme solutions. Morphological studies with SEM-EDX indicate that the enzyme is deposited upon the surface of the PA6 or PA6-Fe microparticles, filling their pores and channels. Each round of immobilization typically removes above 15% of the enzyme contained in the initial stock solution. This process can be performed various times with fresh support particles without a detrimental effect upon the enzymatic activity of the resulting Pec@PA complexes.
The activity studies performed with the Pec@PA6 and Pec@PA6-Fe conjugates showed that they possess a specific activity (i.e., normalized by the protein content) that is up to 2.5 times higher than that of the free enzyme. This effect was explained by comparative kinetic studies that rendered the kinetic parameters of the free enzyme and the two immobilized conjugates. It was found that the immobilization of pectinase on polyamide microparticles results in faster biocatalysts, with a slightly higher affinity to the pectin substrate, being more difficult to inhibit as compared with the respective free enzyme preparation.
The rosé must clarification studies performed at room temperature with the free pectinase and the two immobilized conjugates based on nephelometric measurements showed that complete clarification reaching 20 NTUs for all catalytic systems was achieved within 120–150 min. The reusability of the immobilized pectinase conjugates was tested in rosé must during three consecutive clarification runs. While in the first cycle complete clarification was achieved within 3 h, in the second and third cycles 16 and 36 h were needed, respectively. It should be noted that the implementation of microparticulate polyamide supports for pectinase does not change the color of the must, as measured with UV/VIS in three characteristic wavelength areas.
Currently, an enological evaluation of the wines obtained from musts clarified by means of PA6-supported enological preparations is in progress. Following this evaluation, more extensive tests will be carried out under industrial conditions using the Pec@PA6 and Pec@PA6-Fe conjugates from this study, so as to arrive at a conclusive decision regarding their potential application in the winemaking industry.











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}