
Research Progress on the Effect and Mechanism of Tea Products with Different Fermentation Degrees in Regulating Type 2 Diabetes Mellitus

 , ,
, ,  , and
, and 
Abstract
1. Introduction
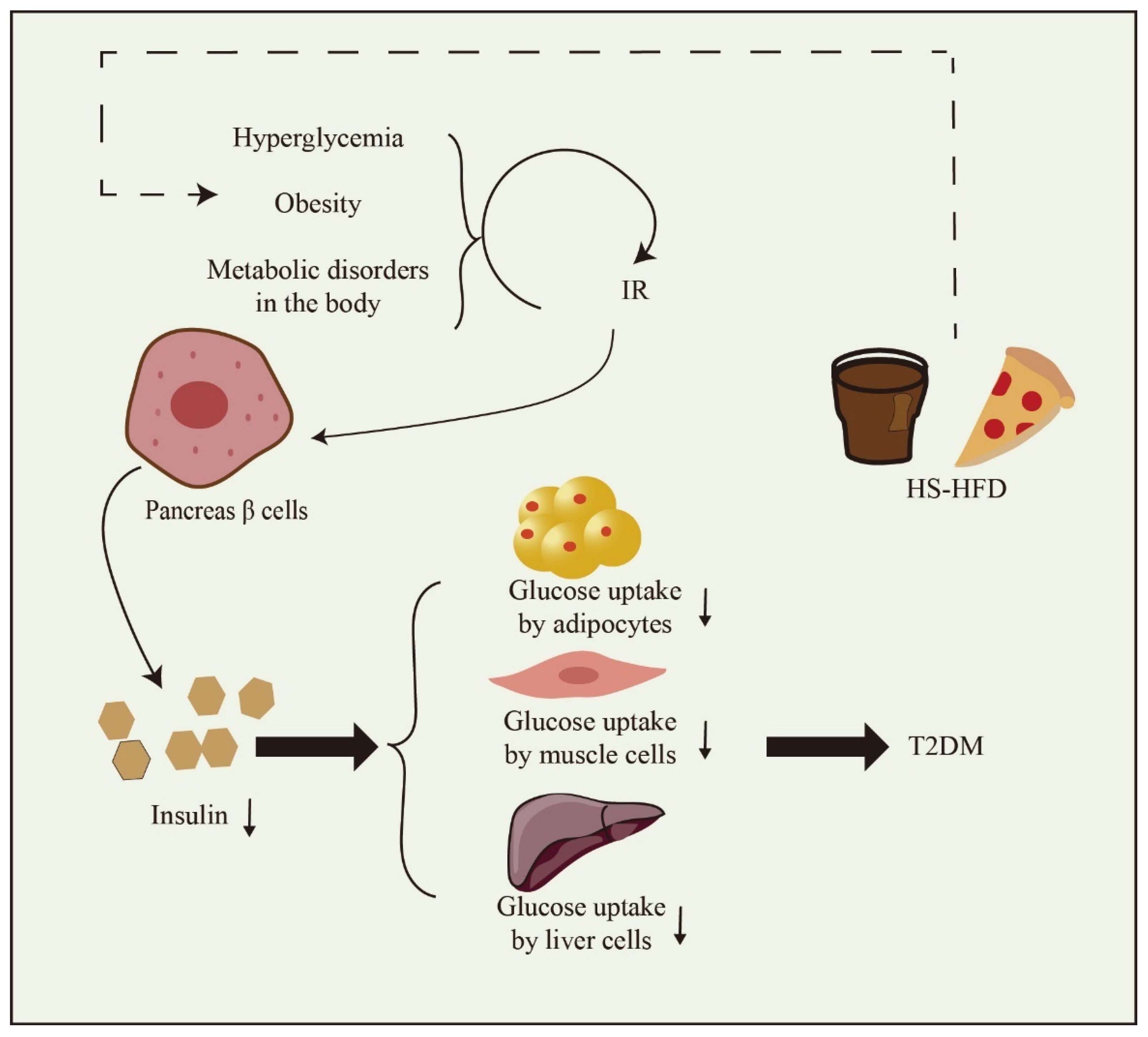
2. The Main Factors of T2DM and Prevention Methods
2.1. The Main Factors of T2DM Mellitus
2.2. The Main Prevention and Treatment Methods of T2DM
2.2.1. Exercise Prevention
2.2.2. Drug Therapy
2.2.3. Application of Natural Plant Extracts in Blood Glucose Regulation
3. The Biochemical Characteristics and Hypoglycemic Effect of Tea with Different Fermentation Degrees
3.1. Biochemical Characteristics of Tea with Different Fermentation Degrees
3.2. Effect of Tea Products with Different Fermentation Degrees on Blood Glucose Balance
3.2.1. Study on Hypoglycemic Activity of Tea with Different Fermentation Degrees in Cell Model
3.2.2. Study on Hypoglycemic Activity of Tea with Different Fermentation Degrees in Animal Models
3.2.3. Epidemiological Investigation on Hypoglycemic Effect of Tea with Different Fermentation Degrees
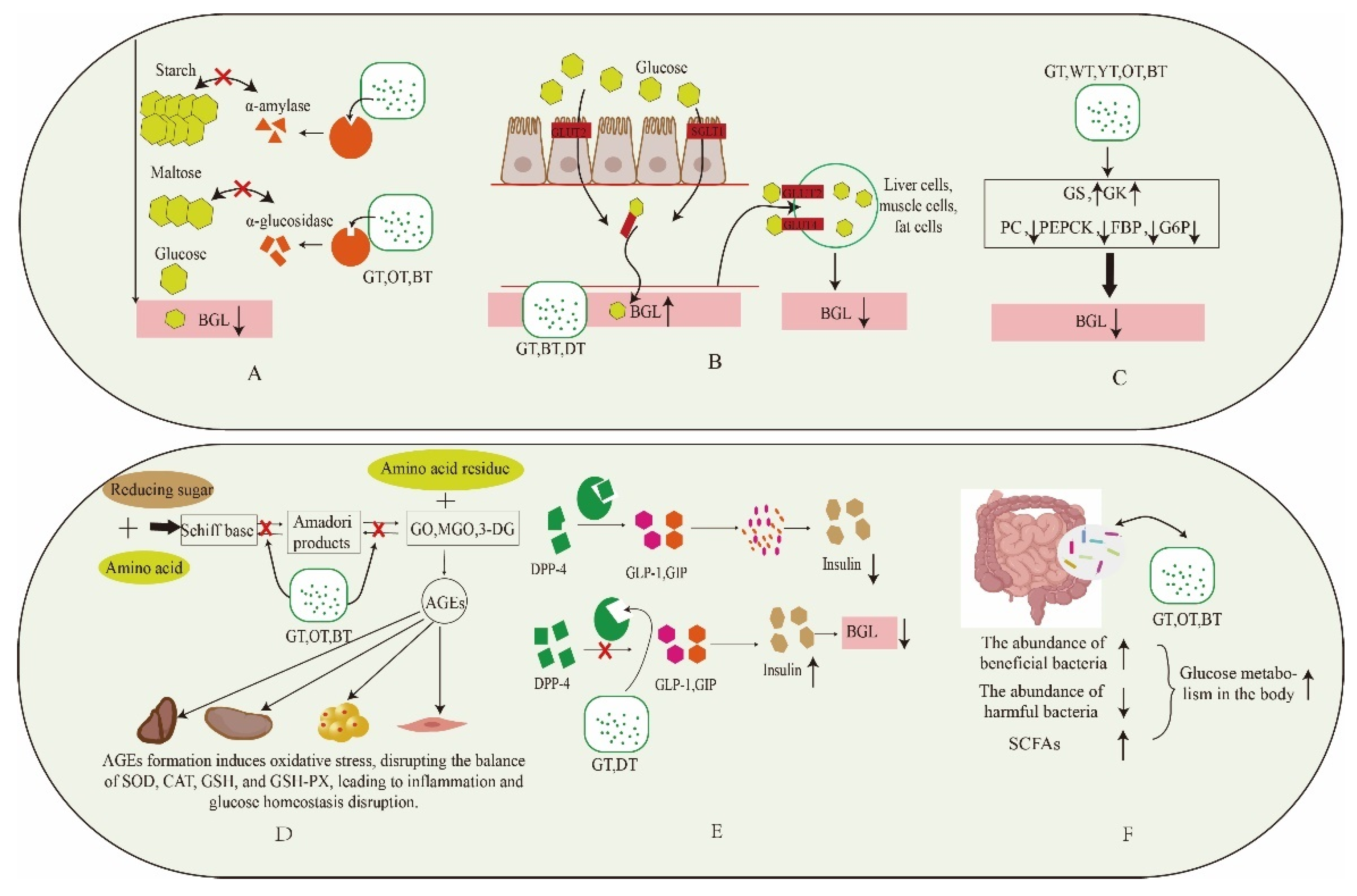
4. The Potential Mechanism of Different Fermented Tea Regulating T2DM
4.1. Inhibition of Digestive Enzymes
4.2. Effect on Glucose Transporters
4.3. Inhibition of Gluconeogenesis Pathway
4.4. Inhibit the Formation of AGEs
4.5. Inhibition of DPP-4 Activity
4.6. Regulation of Gut Microbiota
4.7. Reduce Oxidative Stress
5. Conclusions and Prospect
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Li, H.; Guo, H.; Luo, Q.; Wu, D.-T.; Zou, L.; Liu, Y.; Li, H.-B.; Gan, R.-Y. Current Extraction, Purification, and Identification Techniques of Tea Polyphenols: An Updated Review. Crit. Rev. Food Sci. Nutr. 2023, 63, 3912–3930. [Google Scholar] [CrossRef]
- Greenwalt, C.J.; Steinkraus, K.H.; Ledford, R.A. Kombucha, the Fermented Tea: Microbiology, Composition, and Claimed Health Effects. J. Food Prot. 2000, 63, 976–981. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.; Wang, Y.; Wang, J.; Geng, W. Kombucha Reduces Hyperglycemia in Type 2 Diabetes of Mice by Regulating Gut Microbiota and Its Metabolites. Foods 2022, 11, 754. [Google Scholar] [CrossRef]
- Mendelson, C.; Sparkes, S.; Merenstein, D.J.; Christensen, C.; Sharma, V.; Desale, S.; Auchtung, J.M.; Kok, C.R.; Hallen-Adams, H.E.; Hutkins, R. Kombucha Tea as an Anti-Hyperglycemic Agent in Humans with Diabetes—A Randomized Controlled Pilot Investigation. Front. Nutr. 2023, 10, 1190248. [Google Scholar] [CrossRef]
- Constantino, M.I.; Molyneaux, L.; Limacher-Gisler, F.; Al-Saeed, A.; Luo, C.; Wu, T.; Twigg, S.M.; Yue, D.K.; Wong, J. Long-Term Complications and Mortality in Young-Onset Diabetes: Type 2 Diabetes Is More Hazardous and Lethal than Type 1 Diabetes. Diabetes Care 2013, 36, 3863–3869. [Google Scholar] [CrossRef]
- Saeedi, P.; Petersohn, I.; Salpea, P.; Malanda, B.; Karuranga, S.; Unwin, N.; Colagiuri, S.; Guariguata, L.; Motala, A.A.; Ogurtsova, K.; et al. Global and Regional Diabetes Prevalence Estimates for 2019 and Projections for 2030 and 2045: Results from the International Diabetes Federation Diabetes Atlas, 9th Edition. Diabetes Res. Clin. Pract. 2019, 157, 107843. [Google Scholar] [CrossRef] [PubMed]
- Ji, X.; Guo, J.; Cao, T.; Zhang, T.; Liu, Y.; Yan, Y. Review on Mechanisms and Structure-Activity Relationship of Hypoglycemic Effects of Polysaccharides from Natural Resources. Food Sci. Hum. Wellness 2023, 12, 1969–1980. [Google Scholar] [CrossRef]
- Messina, A.; Monda, V. Role of the Orexin System on Arousal, Attention, Feeding Behaviour and Sleep Disorders. Acta Medica Mediterr. 2017, 33, 645–649. [Google Scholar] [CrossRef]
- Johnson, A.M.F.; Olefsky, J.M. The Origins and Drivers of Insulin Resistance. Cell 2013, 152, 673–684. [Google Scholar] [CrossRef]
- Czech, M.P. Insulin Action and Resistance in Obesity and Type 2 Diabetes. Nat. Med. 2017, 23, 804–814. [Google Scholar] [CrossRef]
- Rhodes, C.J. Type 2 Diabetes-a Matter of ß-Cell Life and Death? Science 2005, 307, 380–384. [Google Scholar] [CrossRef] [PubMed]
- Dempsey, P.C.; Larsen, R.N.; Sethi, P.; Sacre, J.W.; Straznicky, N.E.; Cohen, N.D.; Cerin, E.; Lambert, G.W.; Owen, N.; Kingwell, B.A.; et al. Benefits for Type 2 Diabetes of Interrupting Prolonged Sitting with Brief Bouts of Light Walking or Simple Resistance Activities. Diabetes Care 2016, 39, 964–972. [Google Scholar] [CrossRef] [PubMed]
- Henson, J.; Davies, M.J.; Bodicoat, D.H.; Edwardson, C.L.; Gill, J.M.R.; Stensel, D.J.; Tolfrey, K.; Dunstan, D.W.; Khunti, K.; Yates, T. Breaking Up Prolonged Sitting with Standing or Walking Attenuates the Postprandial Metabolic Response in Postmenopausal Women: A Randomized Acute Study. Diabetes Care 2015, 39, 130–138. [Google Scholar] [CrossRef] [PubMed]
- Savikj, M.; Gabriel, B.M.; Alm, P.S.; Smith, J.; Caidahl, K.; Björnholm, M.; Fritz, T.; Krook, A.; Zierath, J.R.; Wallberg-Henriksson, H. Afternoon Exercise Is More Efficacious than Morning Exercise at Improving Blood Glucose Levels in Individuals with Type 2 Diabetes: A Randomised Crossover Trial. Diabetologia 2019, 62, 233–237. [Google Scholar] [CrossRef]
- Davies, M.J.; Aroda, V.R.; Collins, B.S.; Gabbay, R.A.; Green, J.; Maruthur, N.M.; Rosas, S.E.; Del Prato, S.; Mathieu, C.; Mingrone, G.; et al. Management of Hyperglycaemia in Type 2 Diabetes, 2022. A Consensus Report by the American Diabetes Association (ADA) and the European Association for the Study of Diabetes (EASD). Diabetologia 2022, 65, 1925–1966. [Google Scholar] [CrossRef]
- Tasali, E.; Wroblewski, K.; Kahn, E.; Kilkus, J.; Schoeller, D.A. Effect of Sleep Extension on Objectively Assessed Energy Intake Among Adults with Overweight in Real-Life Settings: A Randomized Clinical Trial. JAMA Intern. Med. 2022, 182, 365–374. [Google Scholar] [CrossRef]
- Sondrup, N.; Termannsen, A.-D.; Eriksen, J.N.; Hjorth, M.F.; Færch, K.; Klingenberg, L.; Quist, J.S. Effects of Sleep Manipulation on Markers of Insulin Sensitivity: A Systematic Review and Meta-Analysis of Randomized Controlled Trials. Sleep Med. Rev. 2022, 62, 101594. [Google Scholar] [CrossRef]
- Wan, C.; Ouyang, J.; Li, M.; Rengasamy, K.R.R.; Liu, Z. Effects of Green Tea Polyphenol Extract and Epigallocatechin-3-O-Gallate on Diabetes Mellitus and Diabetic Complications: Recent Advances. Crit. Rev. Food Sci. Nutr. 2022, 1–29. [Google Scholar] [CrossRef]
- Lorenzati, B.; Zucco, C.; Miglietta, S.; Lamberti, F.; Bruno, G. Oral Hypoglycemic Drugs: Pathophysiological Basis of Their Mechanism of ActionOral Hypoglycemic Drugs: Pathophysiological Basis of Their Mechanism of Action. Pharmaceuticals 2010, 3, 3005–3020. [Google Scholar] [CrossRef]
- Sanchez-Rangel, E.; Inzucchi, S.E. Metformin: Clinical Use in Type 2 Diabetes. Diabetologia 2017, 60, 1586–1593. [Google Scholar] [CrossRef]
- Hashimoto, Y.; Takahashi, F.; Okamura, T.; Hamaguchi, M.; Fukui, M. Diet, Exercise, and Pharmacotherapy for Sarcopenia in People with Diabetes. Metabolism 2023, 144, 155585. [Google Scholar] [CrossRef]
- Fujiwara, Y.; Eguchi, S.; Murayama, H.; Takahashi, Y.; Toda, M.; Imai, K.; Tsuda, K. Relationship between Diet/Exercise and Pharmacotherapy to Enhance the GLP-1 Levels in Type 2 Diabetes. Endocrinol. Diabetes Metab. 2019, 2, e00068. [Google Scholar] [CrossRef]
- Clemmensen, C.; Smajilovic, S.; Smith, E.P.; Woods, S.C.; Bräuner-Osborne, H.; Seeley, R.J.; D’Alessio, D.A.; Ryan, K.K. Oral L-Arginine Stimulates GLP-1 Secretion to Improve Glucose Tolerance in Male Mice. Endocrinology 2013, 154, 3978–3983. [Google Scholar] [CrossRef]
- Dollet, L.; Kuefner, M.; Caria, E.; Rizo-Roca, D.; Pendergrast, L.; Abdelmoez, A.M.; Karlsson, H.K.R.; Bjrnholm, M.; Dalbram, E.; Treebak, J.T.; et al. Glutamine Regulates Skeletal Muscle Immunometabolism in Type 2 Diabetes. Diabetes 2022, 71, 624–636. [Google Scholar] [CrossRef]
- He, Q.; Wang, X.; Yang, C.; Zhuang, X.; Yue, Y.; Jing, H.; Hu, J.; Sun, M.; Guo, L. Metabolic and Nutritional Characteristics in Middle-Aged and Elderly Sarcopenia Patients with Type 2 Diabetes. J. Diabetes Res. 2020, 2020, e6973469. [Google Scholar] [CrossRef]
- Serina, J.J.C.; Castilho, P.C.M.F. Using Polyphenols as a Relevant Therapy to Diabetes and Its Complications, a Review. Crit. Rev. Food Sci. Nutr. 2022, 62, 8355–8387. [Google Scholar] [CrossRef]
- Chan-Zapata, I.; Sandoval-Castro, C.; Segura-Campos, M.R. Proteins and Peptides from Vegetable Food Sources as Therapeutic Adjuvants for the Type 2 Diabetes Mellitus. Crit. Rev. Food Sci. Nutr. 2022, 62, 2673–2682. [Google Scholar] [CrossRef]
- Waddell, I.S.; Orfila, C. Dietary Fiber in the Prevention of Obesity and Obesity-Related Chronic Diseases: From Epidemiological Evidence to Potential Molecular Mechanisms. Crit. Rev. Food Sci. Nutr. 2022, 63, 8752–8767. [Google Scholar] [CrossRef] [PubMed]
- Yi, T.; Zhu, L.; Peng, W.-L.; He, X.-C.; Chen, H.-L.; Li, J.; Yu, T.; Liang, Z.-T.; Zhao, Z.-Z.; Chen, H.-B. Comparison of Ten Major Constituents in Seven Types of Processed Tea Using HPLC-DAD-MS Followed by Principal Component and Hierarchical Cluster Analysis. LWT-Food Sci. Technol. 2015, 62, 194–201. [Google Scholar] [CrossRef]
- Zuo, H.; Si, X.; Li, P.; Li, J.; Chen, Z.; Li, P.; Chen, C.; Liu, Z.; Zhao, J. Dynamic Change of Tea (Camellia sinensis) Leaf Cuticular Wax in White Tea Processing for Contribution to Tea Flavor Formation. Food Res. Int. 2023, 163, 112182. [Google Scholar] [CrossRef] [PubMed]
- Zhou, S.; Zhang, J.; Ma, S.; Ou, C.; Feng, X.; Pan, Y.; Gong, S.; Fan, F.; Chen, P.; Chu, Q. Recent Advances on White Tea: Manufacturing, Compositions, Aging Characteristics and Bioactivities. Trends Food Sci. Technol. 2023, 134, 41–55. [Google Scholar] [CrossRef]
- Wei, Y.; Li, T.; Xu, S.; Ni, T.; Deng, W.-W.; Ning, J. The Profile of Dynamic Changes in Yellow Tea Quality and Chemical Composition during Yellowing Process. LWT 2021, 139, 110792. [Google Scholar] [CrossRef]
- Feng, X.; Yang, S.; Pan, Y.; Zhou, S.; Ma, S.; Ou, C.; Fan, F.; Gong, S.; Chen, P.; Chu, Q. Yellow Tea: More than Turning Green Leaves to Yellow. Crit. Rev. Food Sci. Nutr. 2023, 1–18. [Google Scholar] [CrossRef]
- Chen, Y.L.; Duan, J.; Jiang, Y.M.; Shi, J.; Peng, L.; Xue, S.; Kakuda, Y. Production, Quality, and Biological Effects of Oolong Tea (Camellia sinensis). Food Rev. Int. 2010, 27, 1–15. [Google Scholar] [CrossRef]
- Cao, Q.-Q.; Fu, Y.-Q.; Wang, J.-Q.; Zhang, L.; Wang, F.; Yin, J.-F.; Xu, Y.-Q. Sensory and Chemical Characteristics of Tieguanyin Oolong Tea after Roasting. Food Chem. X 2021, 12, 100178. [Google Scholar] [CrossRef]
- Long, P.; Rakariyatham, K.; Ho, C.-T.; Zhang, L. Thearubigins: Formation, Structure, Health Benefit and Sensory Property. Trends Food Sci. Technol. 2023, 133, 37–48. [Google Scholar] [CrossRef]
- Zhu, K.; Ouyang, J.; Huang, J.; Liu, Z. Research Progress of Black Tea Thearubigins: A Review. Crit. Rev. Food Sci. Nutr. 2021, 61, 1556–1566. [Google Scholar] [CrossRef] [PubMed]
- Lin, F.-J.; Wei, X.-L.; Liu, H.-Y.; Li, H.; Xia, Y.; Wu, D.-T.; Zhang, P.-Z.; Gandhi, G.R.; Li, H.-B.; Gan, R.-Y. State-of-the-Art Review of Dark Tea: From Chemistry to Health Benefits. Trends Food Sci. Technol. 2021, 109, 126–138. [Google Scholar] [CrossRef]
- Zhu, M.; Li, N.; Zhou, F.; Ouyang, J.; Lu, D.; Xu, W.; Li, J.; Lin, H.; Zhang, Z.; Xiao, J.; et al. Microbial Bioconversion of the Chemical Components in Dark Tea. Food Chem. 2020, 312, 126043. [Google Scholar] [CrossRef]
- Jiang, H.; Yu, F.; Qin, L.; Zhang, N.; Cao, Q.; Schwab, W.; Li, D.; Song, C. Dynamic Change in Amino Acids, Catechins, Alkaloids, and Gallic Acid in Six Types of Tea Processed from the Same Batch of Fresh Tea (Camellia sinensis L.) Leaves. J. Food Compos. Anal. 2019, 77, 28–38. [Google Scholar] [CrossRef]
- Vuong, Q.V.; Golding, J.B.; Stathopoulos, C.E.; Nguyen, M.H.; Roach, P.D. Optimizing Conditions for the Extraction of Catechins from Green Tea Using Hot Water. J. Sep. Sci. 2011, 34, 3099–3106. [Google Scholar] [CrossRef] [PubMed]
- Xie, G.; Yan, J.; Lu, A.; Kun, J.; Wang, B.; Song, C.; Tong, H.; Meng, Q. Characterizing Relationship between Chemicals and in Vitro Bioactivities of Teas Made by Six Typical Processing Methods Using a Single Camellia sinensis Cultivar, Meizhan. Bioengineered 2021, 12, 1251–1263. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Ai, Z.; Meng, Y.; Chen, Y.; Ni, D. Comparative Studies on the Physicochemical Profile and Potential Hypoglycemic Activity of Different Tea Extracts: Effect on Sucrase-Isomaltase Activity and Glucose Transport in Caco-2 Cells. Food Res. Int. 2021, 148, 110604. [Google Scholar] [CrossRef] [PubMed]
- Xiang, G.; Sun, H.; Chen, Y.; Guo, H.; Liu, Y.; Li, Y.; Lu, C.; Wang, X. Antioxidant and Hypoglycemic Activity of Tea Polysaccharides with Different Degrees of Fermentation. Int. J. Biol. Macromol. 2023, 228, 224–233. [Google Scholar] [CrossRef]
- Zhou, H.; Li, F.; Wu, M.; Zhu, J.; Wang, Y.; Wei, X. Regulation of Glucolipid Metabolism and Gut Microbiota by Green and Black Teas in Hyperglycemic Mice. Food Funct. 2023, 14, 4327–4338. [Google Scholar] [CrossRef]
- Fujimura, Y.; Watanabe, M.; Morikawa-Ichinose, T.; Fujino, K.; Yamamoto, M.; Nishioka, S.; Inoue, C.; Ogawa, F.; Yonekura, M.; Nakasone, A.; et al. Metabolic Profiling for Evaluating the Dipeptidyl Peptidase-IV Inhibitory Potency of Diverse Green Tea Cultivars and Determining Bioactivity-Related Ingredients and Combinations. J. Agric. Food Chem. 2022, 70, 6455–6466. [Google Scholar] [CrossRef]
- Da Silva, T.B.V.; Castilho, P.A.; de Sá-Nakanishi, A.B.; Seixas, F.A.V.; Dias, M.I.; Barros, L.; Ferreira, I.C.F.R.; Bracht, A.; Peralta, R.M. The Inhibitory Action of Purple Tea on in Vivo Starch Digestion Compared to Other Camellia sinensis Teas. Food Res. Int. 2021, 150, 110781. [Google Scholar] [CrossRef]
- Ni, D.; Ai, Z.; Munoz-Sandoval, D.; Suresh, R.; Ellis, P.R.; Yuqiong, C.; Sharp, P.A.; Butterworth, P.J.; Yu, Z.; Corpe, C.P. Inhibition of the Facilitative Sugar Transporters (GLUTs) by Tea Extracts and Catechins. FASEB J. 2020, 34, 9995–10010. [Google Scholar] [CrossRef]
- Lee, C.-Y.; Oh, J.-H.; Chung, J.-O.; Rha, C.-S.; Park, M.-Y.; Hong, Y.-D.; Kim, W.-K.; Shim, S.-M. Effect of Whole Green Tea Products Including Catechins, Polysaccharides, and Flavonols on the Metabolism of Added Sugars. Food Biosci. 2021, 41, 100936. [Google Scholar] [CrossRef]
- Orita, T.; Chogahara, S.; Okuda, M.; Sakao, K.; Miyata, T.; Hou, D.-X. Extraction Efficiency and Alpha-Glucosidase Inhibitory Activities of Green Tea Catechins by Different Infusion Methods. Foods 2023, 12, 2611. [Google Scholar] [CrossRef]
- Li, S.; Zhang, W.; Wang, R.; Li, C.; Lin, X.; Wang, L. Screening and Identification of Natural α-Glucosidase and α-Amylase Inhibitors from Partridge Tea (Mallotus furetianus Muell-Arg) and in Silico Analysis. Food Chem. 2022, 388, 133004. [Google Scholar] [CrossRef]
- Ueda, M.; Furuyashiki, T.; Yamada, K.; Aoki, Y.; Sakane, I.; Fukuda, I.; Yoshida, K.; Ashida, H. Tea Catechins Modulate the Glucose Transport System in 3T3-L1 Adipocytes. Food Funct. 2010, 1, 167–173. [Google Scholar] [CrossRef]
- Meng, Q.; Qi, X.; Chao, Y.; Chen, Q.; Cheng, P.; Yu, X.; Kuai, M.; Wu, J.; Li, W.; Zhang, Q.; et al. IRS1/PI3K/AKT Pathway Signal Involved in the Regulation of Glycolipid Metabolic Abnormalities by Mulberry (Morus alba L.) Leaf Extracts in 3T3-L1 Adipocytes. Chin. Med. 2020, 15, 1. [Google Scholar] [CrossRef] [PubMed]
- Qu, Z.; Liu, A.; Liu, C.; Tang, Q.; Zhan, L.; Xiao, W.; Huang, J.; Liu, Z.; Zhang, S. Theaflavin Promotes Mitochondrial Abundance and Glucose Absorption in Myotubes by Activating the CaMKK2-AMPK Signal Axis via Calcium-Ion Influx. J. Agric. Food Chem. 2021, 69, 8144–8159. [Google Scholar] [CrossRef] [PubMed]
- Xia, B.; Liu, Q.; Sun, D.; Wang, Y.; Wang, W.; Liu, D. Ultrasound-Assisted Deep Eutectic Solvent Extraction of Polysaccharides from Anji White Tea: Characterization and Comparison with the Conventional Method. Foods 2023, 12, 588. [Google Scholar] [CrossRef]
- Xu, L.; Chen, Y.; Chen, Z.; Gao, X.; Wang, C.; Panichayupakaranant, P.; Chen, H. Ultrafiltration Isolation, Physicochemical Characterization, and Antidiabetic Activities Analysis of Polysaccharides from Green Tea, Oolong Tea, and Black Tea. J. Food Sci. 2020, 85, 4025–4032. [Google Scholar] [CrossRef] [PubMed]
- Tong, T.; Ren, N.; Soomi, P.; Wu, J.; Guo, N.; Kang, H.; Kim, E.; Wu, Y.; He, P.; Tu, Y.; et al. Theaflavins Improve Insulin Sensitivity through Regulating Mitochondrial Biosynthesis in Palmitic Acid-Induced HepG2 Cells. Molecules 2018, 23, 3382. [Google Scholar] [CrossRef]
- Zhou, H.; Wu, Y.; Kim, E.; Pan, H.; He, P.; Li, B.; Chen, Y.C.; Tu, Y. Simultaneous Tests of Theaflavin-3,3′-Digallate as an Anti-Diabetic Drug in Human Hepatoma G2 Cells and Zebrafish (Danio rerio). Nutrients 2021, 13, 4379. [Google Scholar] [CrossRef]
- Tenore, G.C.; Stiuso, P.; Campiglia, P.; Novellino, E. In Vitro Hypoglycaemic and Hypolipidemic Potential of White Tea Polyphenols. Food Chem. 2013, 141, 2379–2384. [Google Scholar] [CrossRef]
- Du, W.; Peng, S.-M.; Liu, Z.; Shi, L.; Tan, L.-F.; Zou, X.-Q. Hypoglycemic Effect of the Water Extract of Pu-Erh Tea. J. Agric. Food Chem. 2012, 60, 10126–10132. [Google Scholar] [CrossRef]
- Shimizu, M.; Kobayashi, Y.; Suzuki, M.; Satsu, H.; Miyamoto, Y. Regulation of Intestinal Glucose Transport by Tea Catechins. BioFactors 2000, 13, 61–65. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Q.Y.; Zhang, A.; Tsang, D.; Huang, Y.; Chen, Z.-Y. Stability of Green Tea Catechins. J. Agric. Food Chem. 1997, 45, 4624–4628. [Google Scholar] [CrossRef]
- Park, C.W.; Kim, H.W.; Ko, S.H.; Lim, J.H.; Ryu, G.R.; Chung, H.W.; Han, S.W.; Shin, S.J.; Bang, B.K.; Breyer, M.D.; et al. Long-Term Treatment of Glucagon-Like Peptide-1 Analog Exendin-4 Ameliorates Diabetic Nephropathy through Improving Metabolic Anomalies in Db/Db Mice. J. Am. Soc. Nephrol. 2007, 18, 1227. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Lv, Q.; Jia, S.; Chen, Y.; Sun, C.; Li, X.; Chen, K. Effects of Flavonoid-Rich Chinese Bayberry (Morella rubra Sieb. et Zucc.) Fruit Extract on Regulating Glucose and Lipid Metabolism in Diabetic KK-Ay Mice. Food Funct. 2016, 7, 3130–3140. [Google Scholar] [CrossRef] [PubMed]
- Ishii, Y.; Ohta, T.; Sasase, T.; Morinaga, H.; Hata, T.; Miyajima, K.; Katusda, Y.; Masuyama, T.; Shinohara, M.; Kakutani, M.; et al. A High-Fat Diet Inhibits the Progression of Diabetes Mellitus in Type 2 Diabetic Rats. Nutr. Res. 2010, 30, 483–491. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Fu, L.; Kojima, R.; Yamamoto, A.; Ueno, T.; Matsui, T. Theaflavins Prevent the Onset of Diabetes through Ameliorating Glucose Tolerance Mediated by Promoted Incretin Secretion in Spontaneous Diabetic Torii Rats. J. Funct. Foods 2021, 86, 104702. [Google Scholar] [CrossRef]
- Ueda-Wakagi, M.; Nagayasu, H.; Yamashita, Y.; Ashida, H. Green Tea Ameliorates Hyperglycemia by Promoting the Translocation of Glucose Transporter 4 in the Skeletal Muscle of Diabetic Rodents. Int. J. Mol. Sci. 2019, 20, 2436. [Google Scholar] [CrossRef] [PubMed]
- Sampath, C.; Rashid, M.R.; Sang, S.; Ahmedna, M. Green Tea Epigallocatechin 3-Gallate Alleviates Hyperglycemia and Reduces Advanced Glycation End Products via Nrf2 Pathway in Mice with High Fat Diet-Induced Obesity. Biomed. Pharmacother. 2017, 87, 73–81. [Google Scholar] [CrossRef]
- Shang, L.; Li, F.; Zhu, J.; Sun, C.; Wang, Y. Selenium-Enriched and Ordinary Black Teas Regulate the Metabolism of Glucose and Lipid and Intestinal Flora of Hyperglycemic Mice. Plant Foods Hum. Nutr. 2023, 78, 61–67. [Google Scholar] [CrossRef]
- Gao, Y.; Xu, Y.; Ruan, J.; Yin, J. Selenium Affects the Activity of Black Tea in Preventing Metabolic Syndrome in High-Fat Diet-Fed Sprague–Dawley Rats. J. Sci. Food Agric. 2020, 100, 225–234. [Google Scholar] [CrossRef]
- Han, M.; Zhao, G.; Wang, Y.; Wang, D.; Sun, F.; Ning, J.; Wan, X.; Zhang, J. Safety and Anti-Hyperglycemic Efficacy of Various Tea Types in Mice. Sci. Rep. 2016, 6, 31703. [Google Scholar] [CrossRef] [PubMed]
- Imran, A.; Butt, M.S.; Arshad, M.S.; Arshad, M.U.; Saeed, F.; Sohaib, M.; Munir, R. Exploring the Potential of Black Tea Based Flavonoids against Hyperlipidemia Related Disorders. Lipids Health Dis. 2018, 17, 57. [Google Scholar] [CrossRef]
- Xu, W.; Zhou, Y.; Lin, L.; Yuan, D.; Peng, Y.; Li, L.; Xiao, W.; Gong, Z. Hypoglycemic Effects of Black Brick Tea with Fungal Growth in Hyperglycemic Mice Model. Food Sci. Hum. Wellness 2022, 11, 711–718. [Google Scholar] [CrossRef]
- Cai, X.; Liu, Z.; Dong, X.; Wang, Y.; Zhu, L.; Li, M.; Xu, Y. Hypoglycemic and Lipid Lowering Effects of Theaflavins in High-Fat Diet-Induced Obese Mice. Food Funct. 2021, 12, 9922–9931. [Google Scholar] [CrossRef] [PubMed]
- Deng, X.; Zhang, N.; Wang, Q.; Huang, Y.; Huang, Y.; Lin, Y.; Huang, M.; Zheng, F.; Xiao, M.; Ye, J. Theabrownin of Raw and Ripened Pu-Erh Tea Varies in the Alleviation of HFD-Induced Obesity via the Regulation of Gut Microbiota. Eur. J. Nutr. 2023, 62, 2177–2194. [Google Scholar] [CrossRef]
- Su, K.; Mao, X.; Zhang, X. Glucose-Lowering Activity of Dark Tea Protein Extract by Modulating Spleen–Brain Axis of Diabetic Mice. Br. J. Nutr. 2021, 126, 961–969. [Google Scholar] [CrossRef]
- Li, H.; Fang, Q.; Nie, Q.; Hu, J.; Yang, C.; Huang, T.; Li, H.; Nie, S. Hypoglycemic and Hypolipidemic Mechanism of Tea Polysaccharides on Type 2 Diabetic Rats via Gut Microbiota and Metabolism Alteration. J. Agric. Food Chem. 2020, 68, 10015–10028. [Google Scholar] [CrossRef]
- Wu, Z.; Zeng, W.; Zhang, X.; Yang, J. Characterization of Acidic Tea Polysaccharides from Yellow Leaves of Wuyi Rock Tea and Their Hypoglycemic Activity via Intestinal Flora Regulation in Rats. Foods 2022, 11, 617. [Google Scholar] [CrossRef]
- Huang, H.; Chen, J.; Chen, Y.; Xie, J.; Xue, P.; Ao, T.; Chang, X.; Hu, X.; Yu, Q. Metabonomics Combined with 16S rRNA Sequencing to Elucidate the Hypoglycemic Effect of Dietary Fiber from Tea Residues. Food Res. Int. 2022, 155, 111122. [Google Scholar] [CrossRef]
- Hajizadeh Tekmeh, H.; Vanizor Kural, B.; Kör, S.; Arıkan Malkoç, M.; Yuluğ, E.; Kutlu, A.; Abidin, İ.; Orem, A. How Does L-Theanine Treatment Affect the Levels of Serum and Hippocampal BDNF, Insulin and Adipocytokines in Diabetic Rats? Biochem. Biophys. Res. Commun. 2023, 667, 95–103. [Google Scholar] [CrossRef]
- Ma, L.; Hu, Y.; Alperet, D.J.; Liu, G.; Malik, V.; Manson, J.E.; Rimm, E.B.; Hu, F.B.; Sun, Q. Beverage Consumption and Mortality among Adults with Type 2 Diabetes: Prospective Cohort Study. BMJ 2023, 381, e073406. [Google Scholar] [CrossRef] [PubMed]
- Imamura, F.; Schulze, M.B.; Sharp, S.J.; Guevara, M.; Romaguera, D.; Bendinelli, B.; Salamanca-Fernández, E.; Ardanaz, E.; Arriola, L.; Aune, D.; et al. Estimated Substitution of Tea or Coffee for Sugar-Sweetened Beverages Was Associated with Lower Type 2 Diabetes Incidence in Case–Cohort Analysis across 8 European Countries in the EPIC-InterAct Study. J. Nutr. 2019, 149, 1985–1993. [Google Scholar] [CrossRef]
- Li, Y.; Wang, C.; Huai, Q.; Guo, F.; Liu, L.; Feng, R.; Sun, C. Effects of Tea or Tea Extract on Metabolic Profiles in Patients with Type 2 Diabetes Mellitus: A Meta-Analysis of Ten Randomized Controlled Trials: Effects of Tea on T2DM Patients. Diabetes Metab. Res. Rev. 2016, 32, 2–10. [Google Scholar] [CrossRef] [PubMed]
- Iso, H.; Date, C.; Wakai, K.; Fukui, M.; Tamakoshi, A. The Relationship between Green Tea and Total Caffeine Intake and Risk for Self-Reported Type 2 Diabetes among Japanese Adults. Ann. Intern. Med. 2006, 144, 554–562. [Google Scholar] [CrossRef] [PubMed]
- Fuchs, D.; Nyakayiru, J.; Draijer, R.; Mulder, T.P.J.; Hopman, M.T.E.; Eijsvogels, T.M.H.; Thijssen, D.H. Impact of Flavonoid-Rich Black Tea and Beetroot Juice on Postprandial Peripheral Vascular Resistance and Glucose Homeostasis in Obese, Insulin-Resistant Men: A Randomized Controlled Trial. Nutr. Metab. 2016, 13, 34. [Google Scholar] [CrossRef] [PubMed]
- Neyestani, T.R.; Shariatzade, N.; Kalayi, A.; Gharavi, A.; Khalaji, N.; Dadkhah, M.; Zowghi, T.; Haidari, H.; Shab-bidar, S. Regular Daily Intake of Black Tea Improves Oxidative Stress Biomarkers and Decreases Serum C-Reactive Protein Levels in Type 2 Diabetic Patients. Ann. Nutr. Metab. 2010, 57, 40–49. [Google Scholar] [CrossRef]
- Odegaard, A.O.; Pereira, M.A.; Koh, W.-P.; Arakawa, K.; Lee, H.-P.; Yu, M.C. Coffee, Tea, and Incident Type 2 Diabetes: The Singapore Chinese Health Study1. Am. J. Clin. Nutr. 2008, 88, 979–985. [Google Scholar] [CrossRef]
- Liu, J.; Liu, S.; Zhou, H.; Hanson, T.; Yang, L.; Chen, Z.; Zhou, M. Association of Green Tea Consumption with Mortality from All-Cause, Cardiovascular Disease and Cancer in a Chinese Cohort of 165,000 Adult Men. Eur. J. Epidemiol. 2016, 31, 853–865. [Google Scholar] [CrossRef]
- Deng, D.; Yan, N. GLUT, SGLT, and SWEET: Structural and Mechanistic Investigations of the Glucose Transporters. Protein Sci. 2016, 25, 546–558. [Google Scholar] [CrossRef]
- Mueckler, M.; Thorens, B. The SLC2 (GLUT) Family of Membrane Transporters. Mol. Asp. Med. 2013, 34, 121–138. [Google Scholar] [CrossRef]
- Huang, S.; Czech, M.P. The GLUT4 Glucose Transporter. Cell Metab. 2007, 5, 237–252. [Google Scholar] [CrossRef] [PubMed]
- Wright, E.M.; Loo, D.D.F.; Hirayama, B.A. Biology of Human Sodium Glucose Transporters. Physiol. Rev. 2011, 91, 733–794. [Google Scholar] [CrossRef]
- Zhang, X.; Yang, S.; Chen, J.; Su, Z. Unraveling the Regulation of Hepatic Gluconeogenesis. Front. Endocrinol. 2019, 9, 802. [Google Scholar] [CrossRef] [PubMed]
- Xia, X.; Xu, J.; Wang, X.; Wang, H.; Lin, Z.; Shao, K.; Fang, L.; Zhang, C.; Zhao, Y. Jiaogulan Tea (Gpostemma pentaphyllum) Potentiates the Antidiabetic Effect of White Tea via the AMPK and PI3K Pathways in C57BL/6 Mice. Food Funct. 2020, 11, 4339–4355. [Google Scholar] [CrossRef] [PubMed]
- Zhao, G.; Teng, J.; Dong, R.; Ban, Q.; Yang, L.; Du, K.; Wang, Y.; Pu, H.; Yang, C.S.; Ren, Z. Alleviating Effects and Mechanisms of Action of Large-Leaf Yellow Tea Drinking on Diabetes and Diabetic Nephropathy in Mice. Food Sci. Hum. Wellness 2023, 12, 1660–1673. [Google Scholar] [CrossRef]
- Dong, L.; Li, Y.; Chen, Q.; Liu, Y.; Wu, Z.; Pan, D.; Yan, N.; Liu, L. Cereal Polyphenols Inhibition Mechanisms on Advanced Glycation End Products and Regulation on Type 2 Diabetes. Crit. Rev. Food Sci. Nutr. 2023, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Hemmler, D.; Roullier-Gall, C.; Marshall, J.W.; Rychlik, M.; Taylor, A.J.; Schmitt-Kopplin, P. Evolution of Complex Maillard Chemical Reactions, Resolved in Time. Sci. Rep. 2017, 7, 3227. [Google Scholar] [CrossRef] [PubMed]
- Tonelli, C.; Chio, I.I.C.; Tuveson, D.A. Transcriptional Regulation by Nrf2. Antioxid. Redox Signal. 2018, 29, 1727–1745. [Google Scholar] [CrossRef]
- Röhrborn, D.; Wronkowitz, N.; Eckel, J. DPP4 in Diabetes. Front. Immunol. 2015, 6, 386. [Google Scholar] [CrossRef]
- Zhao, B.; Su, K.; Mao, X.; Zhang, X. Separation and Identification of Enzyme Inhibition Peptides from Dark Tea Protein. Bioorg. Chem. 2020, 99, 103772. [Google Scholar] [CrossRef]
- Wang, K.; Zhang, Z.; Hang, J.; Liu, J.; Guo, F.; Ding, Y.; Li, M.; Nie, Q.; Lin, J.; Zhuo, Y.; et al. Microbial-Host-Isozyme Analyses Reveal Microbial DPP4 as a Potential Antidiabetic Target. Science 2023, 381, eadd5787. [Google Scholar] [CrossRef] [PubMed]
- Yaribeygi, H.; Sathyapalan, T.; Atkin, S.L.; Sahebkar, A. Molecular Mechanisms Linking Oxidative Stress and Diabetes Mellitus. Oxidative Med. Cell. Longev. 2020, 2020, e8609213. [Google Scholar] [CrossRef] [PubMed]
- Maritim, A.C.; Sanders, R.A.; Watkins, J.B. Diabetes, Oxidative Stress, and Antioxidants: A Review. J. Biochem. Mol. Toxicol. 2003, 17, 24–38. [Google Scholar] [CrossRef] [PubMed]
- Hu, S.; Li, S.; Liu, Y.; Sun, K.; Luo, L.; Zeng, L. Aged Ripe Pu-Erh Tea Reduced Oxidative Stress-Mediated Inflammation in Dextran Sulfate Sodium-Induced Colitis Mice by Regulating Intestinal Microbes. J. Agric. Food Chem. 2021, 69, 10592–10605. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
| GT | WT | YT | OT | BT | DT | FL | Reference | |
|---|---|---|---|---|---|---|---|---|
| Total catechins | 55.16–198.4 | 42.81–175.1 | 52.24–107.9 | 48.46–159.1 | 11.39–3.5 | 30.17–135.9 | 58.60 | [40,42] |
| Tea polyphenols | 238.20 | 236.36 | 233.03 | 224.47 | 133.46 | 218.98 | 245.15 | [40,42] |
| Total free amino acid | 21.856 | 29.642 | 21.596 | 26.050 | 13.382 | 4.288 | 28.896 | [40,42] |
| Caffeine | 15.6–37.3 | 15.2–36.8 | 13.7–38.2 | 14.4–36.1 | 13.4–35.7 | 14.0–36.3 | 15.5 | [40,42] |
| Theobromine | 0.28 | 0.10 | 0.17 | 0.21 | 0.08 | 0.19 | 0.13 | [40,42] |
| Theaflavin | 2.34 | 5.55 | 2.22 | 3.68 | 9.3 | 2.0 | \ | [40,42] |
| Tea | Essential Component | Cell Type | Types | Result/Pathway | References |
|---|---|---|---|---|---|
| GT, OT, BT | Catechin | Caco-2 | Inhibition of glucose uptake | SGLT1 gene expression decreases, and GLUT2 gene and protein expression are inhibited within the first two hours. | [48] |
| GT, OT, BT, DT | Theaflavins, catechins | Caco-2 | Inhibition of digestive enzyme activity and reduction of glucose uptake | BT extract strongly inhibits digestive enzyme activity, while green tea extract has the highest effect on glucose transport (SGLT1 and GLUT2). | [43] |
| GT | Green tea extract (GTE); green tea polysaccharide (CTP); green tea flavonols(FVN) | Caco-2 | Digestive enzyme inhibition; reduce glucose and fructose uptake | GTE can significantly inhibit glucose transport; GTE + CTP + FVN can significantly inhibit fructose transport. | [49] |
| GT | Catechins | Caco-2 | α-glucosidase inhibition | EGCG is the most potent inhibitor for rat α-glucosidase, while ECG is the strongest inhibitor for human Caco-2 cell α-glucosidas. | [50] |
| Partridge tea | Partridge tea water extract | 3T3-L1 | α-amylase and α-glucosidase inhibition; enhance glucose uptake | Partridge tea water extract inhibits digestive enzymes, enhances glucose uptake. | [51] |
| GT | Catechin | 3T3-L1 | Enhance glucose uptake | GT catechins (EC, EGC) can regulate adipose tissue glucose uptake and lower postprandial blood glucose. | [52] |
| Mulberry leaf tea extract | Flavonoid | 3T3-L1 | Enhance glucose uptake | Mulberry leaf flavonoids can reduce the level of free fatty acids and alleviate IR in 3T3-L1 adipocytes./ Activation of IRS1/PI3K/AKT/ GLUT4 pathway. | [53] |
| BT | Theaflavin | C2C12 | Muscle protection | Protect skeletal muscle and maintain muscle health./ Activation of CaMKK2-AMPK signaling axis via Ca2+ influx. | [54] |
| Anji WT | Tea polysaccharide | L6 | Inhibit digestive enzyme activity; enhance glucose uptake | Ultrasound-assisted deep eutectic solvent extraction of Anji WT polysaccharide exhibits strong digestive enzyme inhibitory activity, enhancing glucose uptake by L6 cells. | [55] |
| GT, OT, BT | Tea polysaccharide | L6 | α-glucosidase inhibition; inhibition of AGE formation; enhance glucose uptake | The degree of fermentation is directly linked to the biological activity of tea polysaccharides, including antioxidant, anti-glycosylation, α-glucosidase inhibition, and hypoglycemic effects on L6 myotubes. | [56] |
| BT | Theaflavin | HepG2 | Enhance glucose uptake | Theaflavins promote GLUT4 translocation, boost glucose uptake, reduce IR, and enhance mitochondrial biogenesis while reducing adipogenesis./ Activation of IRS-1/Akt/GLUT4 pathway. | [57] |
| BT | Theaflavin-3,3′-digallate | HepG2 /Zebrafish | Enhance glucose uptake and protect pancreas β cells | Regulation of phosphoenolpyruvate carboxykinase and glucokinase promotes islet β cell regeneration, reducing blood glucose. | [58] |
| GT, WT, BT | Tea polyphenol | HepG2 | Enhance glucose uptake | WT polyphenols had the strongest glucose uptake in HepG2 cells. | [59] |
| Puer tea | Puer tea water extract | HepG2 | Enhance glucose uptake | At a 0.1 g/L concentration, the water extract exhibited higher glucose uptake than 10 μmol/L of acarbose. | [60] |
| Tea | Main Components | Time, Way | Animal Type | Diabetes Induction Mode | Result/ Pathway | References |
|---|---|---|---|---|---|---|
| Roasted GT, kung fu BT | Tea water extract | 5 weeks, free to drink | SPF C57BL/6J male mice | HSFD | It can promote liver glycogen synthesis and inhibit gluconeogenesis./ Activation of IRS-1-PI3K/AKT-GLUT2 pathway. | [45] |
| GT | Green tea extract | 19 days, free to drink | Male Wistar/ST rats; KK-Ay mice | STZ | It can promote GLUT4 translocation in skeletal muscle; enhance glucose uptake. | [67] |
| GT | EGCG | 17 weeks, free to drink | C57BL/6 mice | HFD | It can reduce AGEs to lower plasma glucose and alleviate diabetic complications. | [68] |
| Selenium-rich BT and ordinary BT | Tea water extract | 5 weeks, free to drink | C57BL/6J mice | STZ/HFD | It can inhibit digestive enzymes, regulate glucose metabolism, and alleviate liver injury and inflammation./ Activation of PI3K/Akt pathway. | [69] |
| Selenium-rich BT, selenium-deficient BT | Tea powder | 4 weeks, free intake | Male Sprague-Dawley rats | HFD | It can improve weight, lower total triglycerides and fasting blood glucose, enhance insulin sensitivity, relieve liver and intestinal injury, reduce inflammation, and enrich beneficial intestinal bacteria. | [70] |
| GT, YT, BT | Water extract | 25 days, free to drink | Male ICR mice | HFD | GT and BT maintain body weight in high-fat diet mice, while YT significantly lowers blood glucose. | [71] |
| BT | Theaflavins, thearubigins | 56 days, free to drink | Male Sprague Dawley rats | HSD | Theaflavins and thearubigins reduce plasma glucose and boost insulin release. | [72] |
| BT, DT | Tea water extract | 28 days, gavage (1000, 500, and 300 mg/kg). | Male SPF Kunming mice | Intraperitoneal injection of STZ | Enhances glucose transport, reduces postprandial blood glucose, and inhibits liver glycogen synthesis./ Up-regulation of PI3K, AKT, IRS1, GLUT2; down-regulation of GSK3β protein and gene expression. | [73] |
| GT, OT, BT, DT | Tea polysaccharide | 4 weeks, gavage (0.2 mL, 150 mg/kg, 200 mg/kg, 300 mg/kg) | Male ICR mice | HSFD | BT polysaccharide is highly effective in inhibiting α-glucosidase and reducing triglyceride, total cholesterol, LDL cholesterol, creatinine, alanine aminotransferase, and aspartate aminotransferase. | [44] |
| GT, WT, OT, BT, purple tea | Tea water extract | Gavage, Samples were taken once every 30 min for two hours (500 mg/kg). | Normal male Swiss mice | Normal male Swiss mice | Purple tea shows the highest starch digestion inhibition. BT shows the most significant improvement in glucose tolerance. | [47] |
| BT | Theaflavins | 7 weeks, oral | Male C5BL mice | HFD | It can reduce blood glucose, improve IR, alleviate liver injury, and lower serum triglycerides, total cholesterol, LDL cholesterol, as well as alanine aminotransferase and aspartate aminotransferase levels./ Activation of SIRT6/AMPK/SREBP-1/FASN pathway. | [74] |
| Raw Pu‘er tea, cooked Pu‘er tea | Theabrownin | 24 weeks, gavage (400 mg/kg) | C57BL/6J male mice | HFD | Both inhibit weight gain and maintain glucose homeostasis./ Up-regulation of GLUT4 and IRS1. | [75] |
| DT | Tea protein | 21 days, gavage (50 mg/kg, 100 mg/kg, 125 mg/kg) | SPF C6BL/8 male mice | alloxan | It can inhibit weight, enhance glucose tolerance, and reduce fasting blood glucose. Activation of spleen–brain axis to alleviate hyperglycemia. | [76] |
| GT | Tea polysaccharide | 3 weeks, gavage (100 mg/kg, 200 mg/kg, 400 mg/kg) | Male Wistar rats | HFD | It can reduce blood glucose, promote SCFAs production, alleviate IR, mitigate pancreatic and liver damage, and increase beneficial intestinal bacteria. | [77] |
| Wuyi rock tea (OT) | Tea polysaccharide | 40 days, gavage | Male Wistar rats | HFD/STZ | It can regulate gut flora, boost beneficial bacteria, lower fasting blood glucose, improve glucose tolerance, and ease liver and pancreatic damage. | [78] |
| BT | Theaflavins | 20 weeks, gavage (25 mg/kg/day) | SDT rats | SDT rats | It can improve glucose tolerance, promote incretin secretion and improve IR | [66] |
| Tea grounds | Tea dietary fiber | 28 days, gavage (250 mg/kg, 500 mg/kg, 1000 mg/kg) | Male wistar rats | HFD/STZ | It can improve hyperglycemia, enhance insulin sensitivity, ease pancreatic injury, boost SCFAs levels, and stimulate insulin secretion. | [79] |
| L-theanine | L-theanine | 28 days, gavage (200 mg/kg) | Male Wistar rats | Nicotinamide/STZ | It can reduce leptin and adiponectin levels in the hippocampus of diabetic mice, alleviate hippocampal damage, promote overall blood glucose metabolism, and maintain the glucose balance in the body. | [80] |
| Country | Year | Tea, Method | Sample | Results | References | |||
|---|---|---|---|---|---|---|---|---|
| Total | Males (%) | Females (%) | Average Age | |||||
| USA | 1980–2018 | >2 times/day | 15,486 | 26.4 | 73.6 | 61.3 | Substituting sugar-sweetened beverages with tea, coffee, or plain water can lower mortality and reduce cardiovascular disease incidence in adults with T2DM. | [81] |
| Europe | 1992–2007 | 152 ± 282 g/day | 12,333 | 50 | 50 | 56.0 | Switching to tea from sugary drinks reduces the risk of T2DM by 22%. | [82] |
| Iran | 2009 | 4 cups/day (600 mL) | 23 | / | / | 57.0 | It inhibits serum malondialdehyde, reduces C-reactive protein, increases glutathione levels, and protects the cardiovascular system in diabetic patients. | [86] |
| Netherlands | 2012–2013 | 100 mL BT | 16 | 100 | 0 | / | Consuming BT in diabetic patients lowers peripheral vascular resistance in limbs after glucose intake, improves postprandial blood glucose, insulin concentration, and mitigates IR. | [85] |
| Singapore | 1999–2004 | 1 cup/day or more | 36,908 | / | / | 54.8 | Daily consumption of BT (>1 cup/day) reduces diabetes risk by 14%. | [87] |
| Japan | 5 years random access | 6 cups/day or more | 17,413 | 38.6 | 61.4 | 53.0 | GT reduces diabetes prevalence by 33%. | [84] |
| China | 1991–2006 | 5 g/day or more. | 164,681 | 100 | / | 54.0 | GT consumption lowers all-cause mortality and reduces cardiovascular disease risk in healthy adult males compared to non-drinkers. | [88] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, G.; Zhang, J.; Cui, H.; Feng, Z.; Gao, Y.; Wang, Y.; Chen, J.; Xu, Y.; Niu, D.; Yin, J. Research Progress on the Effect and Mechanism of Tea Products with Different Fermentation Degrees in Regulating Type 2 Diabetes Mellitus. Foods 2024, 13, 221. https://doi.org/10.3390/foods13020221
Li G, Zhang J, Cui H, Feng Z, Gao Y, Wang Y, Chen J, Xu Y, Niu D, Yin J. Research Progress on the Effect and Mechanism of Tea Products with Different Fermentation Degrees in Regulating Type 2 Diabetes Mellitus. Foods. 2024; 13(2):221. https://doi.org/10.3390/foods13020221
Chicago/Turabian StyleLi, Guangneng, Jianyong Zhang, Hongchun Cui, Zhihui Feng, Ying Gao, Yuwan Wang, Jianxin Chen, Yongquan Xu, Debao Niu, and Junfeng Yin. 2024. "Research Progress on the Effect and Mechanism of Tea Products with Different Fermentation Degrees in Regulating Type 2 Diabetes Mellitus" Foods 13, no. 2: 221. https://doi.org/10.3390/foods13020221
APA StyleLi, G., Zhang, J., Cui, H., Feng, Z., Gao, Y., Wang, Y., Chen, J., Xu, Y., Niu, D., & Yin, J. (2024). Research Progress on the Effect and Mechanism of Tea Products with Different Fermentation Degrees in Regulating Type 2 Diabetes Mellitus. Foods, 13(2), 221. https://doi.org/10.3390/foods13020221