Abstract
This study aimed to assess the impact of Chlorella vulgaris supplementation in broilers’ diet, alone or in combination with vitamin E, on meat quality parameters, nutritional value, and oxidative stability during storage time. An experiment was conducted on 180 COBB 500 broiler chickens (14 days old), assigned into six treatments, following a 2 × 3 factorial arrangement. A corn–soybean meal diet was supplemented with three levels of C. vulgaris (0% in group C1, 1% in E1, 2% in E2), two levels of vitamin E (0% in C1, 250 ppm in C2), and a combination of them (1% C. vulgaris + 250 ppm vitamin (E3), 2% C. vulgaris + 250 ppm vitamin (E4)). Dietary incorporation of C. vulgaris, including those supplemented with vitamin E, resulted in a significant increase in meat protein content. DPA and DHA levels increased by 2.01-fold and 1.60-fold in the 2% C. vulgaris + vitamin E group. The PUFA/SFA ratio was increased across all dietary treatments (p < 0.0001). HPI and h/H registered the highest values as a result of 2% C. vulgaris supplementation, being linked with a positive effect in lowering cholesterol levels. Supplementation with 2% C. vulgaris and vitamin E exhibited a 1.45-fold increase in vitamin E concentration in thigh meat compared to the control group, being the highest level registered in thigh meat in this experiment. Metmyoglobin concentrations registered lower values in the thigh meat of the experimental groups, while deoxymyoglobin increased in the same groups when compared to the control group. The inclusion of C. vulgaris (1% and 2%) in combination with vitamin E (250 mg/kg) in broiler diets exhibited the best prevention of lipid oxidation after 7 days of refrigerated storage, defined by the highest efficiency factors assessed in terms of secondary oxidation products.
1. Introduction
Microalgae are a diverse group of photosynthetic organisms that are known as a source of valuable bioactive compounds with numerous benefits for applications in the pharmaceuticals, food, cosmetics, and bioenergy sectors [1]. Chlorella vulgaris is a unicellular green microalga in the division Chlorophyta and a rich source of high-quality protein due to the important concentrations of essential amino acids. The protein content can range between 42 and 65.5% [2,3], with a lysine level of up to 13.2% [4]. C. vulgaris also contains important amounts of lipids, with a high content of polyunsaturated fatty acids and carbohydrates, minerals, vitamins, phenolic compounds, and pigments, such as chlorophylls and carotenoids [5,6]. During the final stage of their metabolism, microalgae undergo a transition to the carotenoid production stage, accumulating elevated quantities of pigments [7]. Carotenoids frequently present in C. vulgaris biomass include β-carotene, a precursor of vitamin A, along with lutein, astaxanthin, canthaxanthin, and violaxanthin. Chlorophylls are the predominant pigments within C. vulgaris cells, surpassing synthetic dyes and being a preferred alternative for food and cosmetic applications [8]. Additionally, chlorophyll exhibits therapeutic properties that are beneficial for ulcer treatment and liver recovery, promoting enhanced cell growth and repair [9]. Carotenoids and chlorophylls possess notable antioxidant and radical scavenging activity and have transitioned from being regarded solely as bioactive compounds to emerging as biomarkers signaling the onset of diseases associated with oxidative stress [10]. Moreover, C. vulgaris cells contain phenolic compounds recognized for their antioxidant, antifungal, and antibacterial activity. Additionally, it has been reported that the presence of phenolics in C. vulgaris formulations contributes to their antidiabetic effects [11].
Microalgae cultivation is recognized as an environmentally friendly process [12] because it requires simple conditions (CO2 and sunlight) and can be carried out in non-agricultural lands. Furthermore, microalgae cultivation does not necessitate the use of pesticides, unlike conventional crops, and it yields higher production outputs [13]. As photosynthetic organisms, microalgae efficiently convert atmospheric carbon dioxide into high-value compounds, playing a role in reducing CO2 levels in the atmosphere [14].
Due to its rich composition of essential nutrients, including high levels of natural antioxidants, C. vulgaris has been used in the nutraceutical industries as a replacement for synthetic antioxidants [15], and also in animal feeding as an alternative sustainable high-quality protein feed resource [16]. The growth of the poultry industry increases the demand for agricultural output for feed production. Proteins are the most expensive and most critical components in feed formulation. To address these issues, substantial efforts have been directed towards identifying alternative sustainable high-quality protein feed resources. C. vulgaris emerges as a valuable source of protein and other nutrients for animal feeding [17]. Additionally, its abundance of biologically active compounds makes these organisms highly appealing as feed ingredients, offering benefits that extend beyond mere nutrient supply [18]. Moreover, C. vulgaris shows potential to mitigate the environmental footprint of poultry production. Studies have shown that incorporating C. vulgaris into 10-day-old broilers’ diets can result in enhancing growth performance, gut functional characteristics, and bacterial communities [19]. Moreover, [20] explored the potential of C. vulgaris as a candidate alternative to antibiotics in broiler chickens, revealing its influence on affecting the humoral immune response in broiler chicks. The impact of C. vulgaris in broilers’ feed on the digestibility and mineral bioaccessibility in meat has been studied in [21], showing an improved nutritional composition of meat in terms of minerals and essential amino acid content.
This study aimed to assess the impact of C. vulgaris supplementation in broilers’ diet, alone or in combination with vitamin E, on meat quality parameters, nutritional value, and oxidative stability during storage time.
2. Materials and Methods
The experimental trial was carried out with the approval (no. 2761/03.03.2021) of the Ethics Committee of the National Research and Development Institute of Animal Biology and Nutrition in Romania. The experimental procedures followed the guidelines specified in Romanian Law 43/2014 regarding the handling and protection of animals used for experimental purposes and complied with Directive 2010/63/EU on the protection of animals used for scientific purposes.
2.1. Experimental Design
An experiment was conducted on 180 COBB 500 broiler chickens (14 days old), assigned into six treatments, following a 2 × 3 factorial arrangement. The initial average weight of birds at 14 days of age was 421.48 g. A corn–soybean meal diet was supplemented with 3 levels of C. vulgaris (0% (C1), 1% (E1), 2% (E2)), 2 levels of vitamin E (0% (C1), 250 ppm (C2)), and a combination of the two supplements (1% C. vulgaris + 250 ppm vitamin (E3), 2% C. vulgaris + 250 ppm vitamin (E4)), as presented in Table 1. Each group included five replicates with six birds per replicate and thirty chicks per treatment. The plant material used in this study consisted of a powder of microalga Chlorella vulgaris, supplied by Vivio (Brzozów, Poland).

Table 1.
The structure of the experimental diets.
The chicks were housed in digestibility pens in a poultry experimental hall with controlled environmental conditions (average temperature/total period: 25.08 ± 0.58 °C; humidity: 61.27 ± 4.79%; ventilation/broiler: 0.61 ± 0.16%). The light regimen was 23 h light/1 h darkness. The chicks had free access to feed and water. At the end of the feeding trial, 6 broilers (42 days old) were randomly selected from each treatment group and slaughtered by cervical dislocation. The thigh meat samples without skin were collected to assess the impact of the dietary supplements on the meat quality in terms of proximate composition, vitamin E, fatty acid profile, and oxidative stability parameters. Samples were stored in plastic bags at −80 °C until further analysis. Instrumental color measurements and pH assessment were performed on fresh meat samples.
2.2. Determination of Meat Quality Traits
The pH was measured on thigh muscles from the right side of the bird using a glass penetration pH electrode (HI9025, Hanna Instruments, Woonsocket, RI, USA) at 45 min postmortem. The pH value was determined as the average of three replicate measurements taken on the same muscle. The standard color parameters, lightness (L*), redness (a*), and yellowness (b*), were determined for thigh meats with a Minolta CR-400 Chroma Meter (Minolta Camera Co. Ltd., Osaka, Japan). The final CIELAB color parameters were the average of 3 readings, taken from 3 spots at the surface of thigh meats.
The chroma value (C*) was calculated as explained in [22], using the following equation:
where a* = redness and b* = yellowness.
C* = (a*2 + b*2)1/2,
The hue angle (h*) was assessed with the following formula:
where a* = redness and b* = yellowness.
h* = tan−1(b*/a*) × (360/2π),
The total color difference (ΔE*) in the thigh meat samples between the control diet (C1) and the experimental diets was calculated according to the following equation:
where L* = lightness, a* = redness, and b* = yellowness.
ΔE* = (ΔL* + Δa* + Δb*)1/2,
Cooking loss was assessed as described in [23]. Individual samples of thigh meat with similar weight and size were packed in plastic bags and cooked in a water bath (Memmert, Schwabach, Germany) for 30 min at 85 °C. After cooling at room temperature, the samples were dried with a paper towel and reweighed. The cooking loss was calculated by the following equation:
where W1 = initial sample weight before cooking and W2 = sample weight after cooking.
Cooking loss % = (W1 − W2) × 100/W1,
To evaluate drip loss, samples of fresh thigh muscle were weighed (W1), placed into a plastic bag, and hung in a refrigerator at 4 °C for 24 h [24]. Subsequently, samples were dried and weighed again (W2), using the following equation:
where W1 = initial sample weight and W2 = final sample weight.
Drip loss (%) = (W1 − W2) × 100/W1,
2.3. Chemical Analysis
2.3.1. Proximate Composition
The proximate composition of C. vulgaris and meat samples was determined using the reference method as described below: crude protein (ISO 5983-2/2009 [25]) with the Kjeldahl method and a semiautomatic Kjeltec auto 1030 Tecator Instruments (Höganäs, Sweden), crude fat (SR ISO 6492/2001 [26]) with the method of continuous solvent extraction and the equipment Soxtec 2055 Foss Tecator (Höganäs, Sweden), crude fiber using the intermediary filtration method and the equipment Fibertec 2010 System Foss Tecator (Höganäs, Sweden), and dry matter (ISO 6496/2001 [27]) and ash (ISO 2171/2010 [28]) using the gravimetric method and a Nabertherm calcination furnace (Nabertherm GmbH, Lilienthal, Germany).
2.3.2. Fatty Acid Determination
The fatty acid profiling of C. vulgaris and thigh meat was assessed as outlined in [29]. Fatty acids (FAs) were converted into FA methyl ester (FAME). Afterward, derivatives of FAs were analyzed using a Perkin Elmer Clarus 500 gas chromatograph (Waltham, MA, USA) equipped with a capillary column with a highly polar stationary phase (Thermo Electron, Waltham, MA, USA), with dimensions of 60 m × 0.25 mm × 0.25 µm film. Detection was conducted using a flame ionization detector (FID), and identification and quantification were achieved by referencing analytical standards.
Lipid quality indexes in thigh meat were determined based on the data obtained from the fatty acid composition of thigh samples, and their relevance to human nutrition was assessed. They were calculated using the following equations [30,31,32,33,34,35]:
AI (Atherogenic Index) = (C12:0 + 4 × C14:0 + C16:0)/(ΣMUFA + Σn6 + Σn3)
TI (Thrombogenic Index) = (C14:0 + C16:0 + C18:0)/(0.5 × ΣMUFA + 0.5 × Σn6 + 3 × Σn3 + Σn3/Σn6)
OFA (hypocholesterolemic fatty acids) = (C14:0 + C16:0) dietary FA with an undesirable effect of increasing cholesterol levels in humans
DFA (hypocholesterolemic fatty acids) = (MUFA + PUFA + C18:0) dietary fatty acids with a beneficial neutral effect on lowering cholesterol levels in humans
HPI (Health-Promoting Index) = UFA/[C12:0 + (4 × C14:0) + C16:0])
PI (Peroxidisability Index) = (monoenoic acid × 0.025) + (dienoic acid × 1) + (trienoic acid × 2) + (tetraenoic acid × 4) + (pentaenoic acid × 6) + (hexaenoic acid × 8)
DBI (Double Bond Index) = (% monoenoic acids) + 2 (% dienoic acids) + 3 (% trienoic acids)/100
IV (Iodine Value) = (0.95 × C16:1) + (0.86 × C18:1n-9) + (1.732 × C18:2n-6) + (2.616 × C18:3n-3) + (0.785 × C20:1).
h/H-hypocholesterolemic FAs/hypercholesterolemic FAs ratio = 5 [C18:1 cis n-9 + C18:2n-6 + C18:3n-6 + C18:3n-3 + C20:3n-6 + C20:4n-6 + C20:5n-3 + C22:4n-6 + C22:5n-3 + C22:6n-3)]/(C14:0 + C16:0).
COX (Calculated oxidizability) = (1 × C18:1 + 10.3 × C18:2 + 21.6 × C18:3)/100
OS (Oxidative susceptibility) = MUFA + 45 × C18:2 + 100 × C18:3
2.3.3. Liposoluble Antioxidants
The liposoluble antioxidants (vitamin E and xanthophylls) were analyzed as previously described in [36]. The extraction procedure involved saponification with ethanolic KOH and extraction with petroleum ether. The final extract was washed with distilled water to remove any alkaline traces and evaporated under vacuum until dry. The concentrated extracts were dissolved in ethanol.
Xanthophylls were analyzed using an HPLC (Finningan Surveyor Plus, Thermo-Electron Corporation, Waltham, MA, USA) with a PDA-UV detector at wavelength 445 nm, and a C18 reversed-phase column with a stationary phase of 5 µm (250 × 4.60 mm i.d.) (Nucleodur, Macherey-Nagel, Duren, Germany). The chromatographic analysis was performed in isocratic conditions, involving a mobile phase which consists of 87% acetone and 13% water, at a flow rate of 1 mL/min.
Vitamin E was analyzed using an HPLC (Vanquish Thermo-Electron Corporation, Waltham, MA, USA), a PDA-UV detector at wavelength 292 nm, and a HyperSil BDS C18 column, with silica gel, dimensions 250 × 4.6 mm, and particle size 5 µm (Thermo-Electron Corporation, Waltham, MA, USA). The mobile phase used was 96% methanol and 4% water, with a flow rate of 1.5 mL/min, in isocratic conditions.
2.3.4. Water-Soluble Antioxidants
The total polyphenol content (TPC) was measured using the Folin–Ciocalteu spectrophotometric method [30]. A calibration curve of gallic acid was employed to quantify the total phenol content, and the results were expressed as mg of gallic acid equivalents per g of dried sample (mg GAE/g).
The total flavonoid content was determined using the aluminum chloride colorimetric method described in [30]. The absorbance was measured at 410 nm against a blank using a UV-VIS spectrophotometer (Jasco V-530, Japan Servo Co. Ltd., Tokyo, Japan). A calibration curve with quercetin as the standard was used and the flavonoid content was expressed as mg of quercetin equivalent (QE) per g.
2.3.5. Antioxidant Capacity Analysis
The antioxidant capacity of C. vulgaris was evaluated using two different spectrophotometric methods for the determination of DPPH and iron chelating ability. A DPPH solution prepared in methanol (0.2 mM) was mixed with the sample extract and distilled water in a ratio of 2:0.4:1.6 (v/v/v). The absorbance was then measured at 517 nm using a spectrophotometer (Jasco V-530, Japan Servo Co., Ltd., Tokyo, Japan). Trolox was used as a reference for standard calibration curves to analyze the DPPH concentration [37]. The results were expressed as mmol of Trolox equivalents per kg of sample (mmol eq Trolox/kg sample).
To assess the chelating effect on ferrous ions, a method described by [38] was used. The method involves measuring the absorbance of the purple complex formed when an extract competes with ferrozine for ferrous ions. This method is based on measuring the absorbance of the purple complex formed when an extract competes with ferrozine for ferrous ions. The absorbance was measured using a UV-VIS spectrophotometer (JASCO V-560, Japan Servo Co. Ltd., Tokyo, Japan) at 562 nm, compared to a blank.
2.3.6. Oxidative Stability of Meat
The oxidative stability of the thigh meat samples was assessed by evaluating primary lipid degradation parameters, including peroxide values (PVs), conjugated dienes (CDs), and conjugated trienes (CTs), as well as secondary parameters such as p-anisidine values and thiobarbituric acid-reactive substances (TBARSs). These measurements were conducted using previously described methods [39].
Additionally, markers of lipid peroxidation included myoglobin derivatives (metmyoglobin, deoxymyoglobin, and oxymyoglobin). All the aforementioned parameters were measured spectrophotometrically using a V-530 Jasco spectrophotometer (Tokyo, Japan), following the methods described in [39].
2.4. Relative Prevention of Lipid Oxidation—Efficiency Factor
The relative inhibition of lipid oxidation was quantified as described in [40], as an efficacy ratio on day 7 of storage, calculated by dividing the PV or TBARS value of the control group by that of the experimental groups. A higher efficiency factor indicates greater effectiveness of the dietary treatments in decreasing lipid oxidation (PV or TBARS) in the thigh meat of broilers.
2.5. Statistical Analysis
The analytical data regarding the nutritional composition, lipid quality indexes, and markers of lipid peroxidation assessed in thigh meat samples were analyzed using a 2-way ANOVA, followed by Tukey’s HSD test, with XLStat (Addinsoft, New York, NY, USA). The statistical model included the fixed effects of vitamin E (0 and 250 ppm) and C. vulgaris (0%, 1%, and 2%) as well as the interaction between them. p-values less than 0.05 were considered significant. Each broiler that was chosen for collecting samples of thigh meat was treated as an experimental unit.
3. Results
3.1. Nutritional Composition of C. vulgaris
The results regarding the nutritional composition of Chlorella vulgaris are presented in Table 2.
Table 2.
Nutritional composition of C. vulgaris.
C. vulgaris was found to be a rich source of essential nutrients, including proteins, fatty acids, vitamins, and antioxidants. The nutritional composition analysis highlighted the high protein content of the analyzed microalga, making it a valuable source of high-protein nutritional supplements. The antioxidant analysis of C. vulgaris showed elevated concentrations of vitamin E and xanthophylls, which increase the antioxidant capacity and enhance the nutraceutical potential of C. vulgaris.
The FA profile of Chlorella vulgaris allowed for the identification of 14 FAs (Table 2), including saturated (C12:0, C14:0, C15:0, C16:0, C17:0, C18:0 and C20:0), monounsaturated (C14:1, C15:1, C16:1, C17:1 and C18:1), and polyunsaturated (C18:2, C18:3) FAs. The primary fatty acids of Chlorella were linoleic, palmitic, alpha-linolenic, oleic, heptadecenoic, and heptadecanoic acids.
3.2. Meat Quality Traits
The effect of dietary treatments on the meat quality traits of broilers is shown in Table 3. Thighs from birds fed with diets supplemented with C. vulgaris and vitamin E (E3 and E4 groups) had significantly higher values of pH than birds from the control group (C1). Thigh lightness (L*) registered a slight increase in the E3 and E4 groups compared to the control, but the values were not statistically different.
Table 3.
Effects of dietary treatments on the physicochemical properties of thigh muscle of broiler chickens.
Dietary incorporation of C. vulgaris, including those supplemented with vitamin E, resulted in a minor but statistically significant (p < 0.05) increase in b*, C*, and h*, while a* registered lowered values (p < 0.05) when compared to the control group. Nevertheless, the calculated color differences (ΔE*) between the samples of the studied groups showed a minor variation, which led to the conclusion that no differences in color could be observed. Neither cooking loss nor drip loss was affected by dietary treatments (p > 0.05) in thigh meat.
3.3. Proximate Composition of Meat
The effect of dietary microalgae on the proximate composition of thigh meat is presented in Figure 1. The incorporation of 1 and 2% C. vulgaris, alone or with vitamin E, resulted in an increase in protein content in the meat (p < 0.0001). Ash content was increased in all the groups, although no significant differences were found between them, except for the 2% C. vulgaris and vitamin E group (E4). Fat content was significantly lower (p < 0.0001) in the thigh muscle of broilers that were fed a diet with C. vulgaris or vitamin E or a combination of them, in comparison to those fed the control diet.
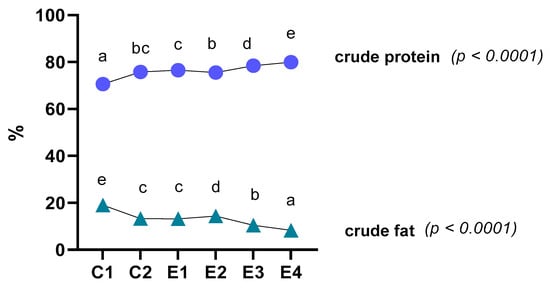
Figure 1.
Effect of dietary treatments on the levels of crude protein and crude fat of thigh muscle of broilers. Different letters indicate a significant difference (p < 0.05).
3.4. Fatty Acid Profiling of Meat
The effect of C. vulgaris incorporation, alone or combined with vitamin E, on the fatty acid composition of thigh meats is presented in Table 4. Dietary treatments induced changes in the fatty acid profile of thigh meat. The percentage of palmitic acid was lower (p < 0.0001) in the thighs of broilers fed C. vulgaris and C. vulgaris + vitamin E diets. Chlorella-fed broilers had a higher concentration of α-linolenic, erucic, and eicosatrienoic acids (p < 0.0001) in their thigh meat compared to the control group. Linolenic acid was detected in a small amount only in groups supplemented with C. vulgaris and vitamin E (E3 and E4 groups). The proportions of docosapentaenoic acid (DPA) and docosahexaenoic acid (DHA) were enhanced in the thighs of broilers fed with C. vulgaris, with significant increases being observed in the group supplemented with 2% C. vulgaris and vitamin E (E4). In fact, both DPA and DHA levels increased by 2.01-fold and by 1.60-fold in the 2% C. vulgaris + vitamin E group (E4). Moreover, the total amount of omega-3 FAs was higher in the thighs of broilers fed with C. vulgaris.
Table 4.
Fatty acid profile in thigh meat.
Concerning the fatty acid ratios, a significantly decreased n-6/n-3 ratio (p < 0.05) in the thigh muscle was observed in groups supplemented with 2% C. vulgaris, alone or with vitamin E (E2 and E4 groups), and also in the 1% C. vulgaris with vitamin E group (E3), when compared with the control. The PUFA/SFA ratio was increased across all dietary treatments (p < 0.0001). Both DFAs and OFAs were influenced by the C. vulgaris supplementation. The highest percentage of hypocholesterolemic fatty acids (DFAs) and the lowest of hypercholesterolemic fatty acids (OFAs) were shown for thighs from the C. vulgaris supplementation groups. The DFA/OFA index registered the highest increase in the 2% C. vulgaris with vitamin E group (E4).
3.5. Lipid Quality Indexes
To estimate the lipid nutritional and health benefits of C. vulgaris, alone or combined with vitamin E, the lipid quality indexes were calculated (Table 5). The values of oxidative susceptibility (OS), double-bond index (DBI), iodine value (IV), and peroxidisability index (PI) from meat samples of experimental groups presented a significant (p < 0.05) increase compared to the control group. A decreasing tendency was observed in the AI of the thighs from broilers fed with C. vulgaris, a significant reduction (p < 0.05) being observed in the groups with 2% C. vulgaris supplementation (E2 and E4). In the same manner, the TI registered the highest significant (p < 0.05) decrease as an effect of the 2% C. vulgaris supplementation in broilers’ diet (E2 and E4). The h/H values were significantly higher in C. vulgaris-supplemented groups when compared to the two control groups. The calculated oxidizability value (COX) was also measured for all meat samples. The results showed a significant increase (p < 0.05) in the COX value in all the groups, except the E3 group where the differences were not significantly higher. Therefore, the thigh meat from the experimental groups (including C1 and excluding E3) exhibits better oxidative stability and thus has a longer shelf life.
Table 5.
Lipid quality indices in thigh meat.
The health-promoting index (HPI) enables the assessment of the impact of fatty acids on cardiovascular diseases. It represents the inverse of the atherogenicity index, indicating inverse values compared to the AI, and, in the present study, registered a favorable increase in the groups with supplemented C. vulgaris. The significant variation in values obtained from the different indexes highlights their distinct meanings and the necessity of using each index under specific conditions.
3.6. Vitamin E Concentration in Thigh Meat
The effect of feeding treatments on the thigh meat content of vitamin E is presented in Table 6. The concentration of vitamin E was influenced by feeding treatments (p < 0.05), showing a significant increase in all the groups compared to the control group (C1); the increase ranged between 21.7 and 44.69%. Supplementation of broiler diet with 2% C. vulgaris and 250 mg/kg vitamin E exhibited a 1.45-fold increase in vitamin E concentration in thigh meat compared to the control group, being the highest level registered in thigh meat in this experiment.
Table 6.
Effects of dietary treatments on the vitamin E concentration in the thigh muscle of broiler chickens.
3.7. Oxidative Stability of Meat
Supplementation of broilers’ diets with C. vulgaris, alone or combined with vitamin E, had no significant effect on the primary oxidation products in thigh meat stored for 7 days in refrigerated conditions (Table 7), although a decreasing tendency was observed for conjugated dienes, conjugated trienes, and peroxide value in the experimental groups compared to the control group (C1). Regarding the secondary oxidation products, TBARS levels were significantly reduced (p < 0.05) in the experimental groups relative to the reference group (C1) after 7 days of refrigeration. For p-anisidine, lower concentrations were also observed in the experimental groups, but without statistical significance.
Table 7.
Effects of dietary treatments on the markers of lipid oxidation of thigh muscle of broiler chickens.
Incorporation of C. vulgaris, alone or combined with vitamin E, in broilers’ diet led to changes in the hem pigments in thigh meat stored for 7 days in refrigerated conditions (Figure 2). The results showed a significant increase (p > 0.05) in the deoxymyoglobin percentage in all the experimental groups compared to the control group, with no significant differences observed between the experimental groups. Another positive effect was the decrease in metmyoglobin concentrations in the thigh meat of the experimental groups, with significant differences noted (p < 0.05) when compared to the control group. The oxymyoglobin levels did not register any significant differences between the studied groups.
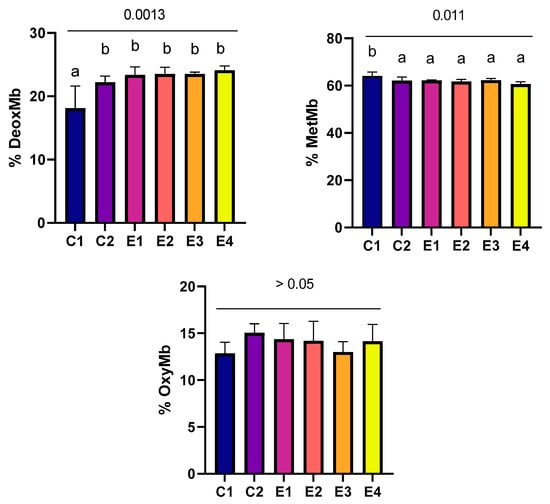
Figure 2.
Concentrations of myoglobin derivatives in thigh meat. Data represented as means ± standard deviation (SD); means with no common superscript are significantly different (p < 0.05).
Figure 3 shows the efficiency factor of the dietary treatments in relation to PV and TBARS measurements on day 7 of refrigerated storage of thigh meat. A higher efficiency factor indicates a greater ability of the antioxidants to reduce lipid oxidation (as measured by PV or TBARS) in the meat samples.
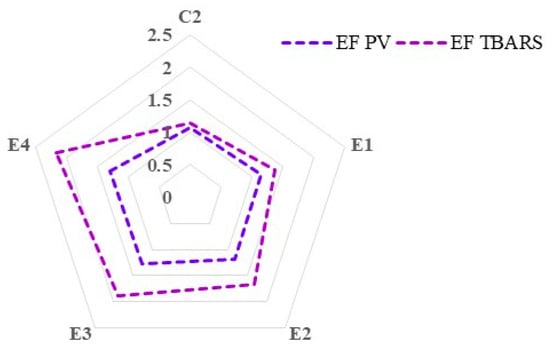
Figure 3.
Relative prevention of lipid oxidation expressed as the efficiency factor (EF) from thigh meat samples on day 7 of refrigerated storage, calculated as the ability of the contained antioxidants to inhibit lipid oxidation, measured as peroxide value (PV) and thiobarbituric acid reactive substances (TBARS) in thigh meat sample.
After 7 days of storage in refrigerated conditions, thigh meat from the group receiving a diet supplemented with 2% C. vulgaris and vitamin E (E4) showed the highest efficiency factor (lowest degree of lipid oxidation compared to the control group C1), followed by the group fed a diet with 1% C. vulgaris and vitamin E (E3), for both PV and TBARS measurements. The inclusion of C. vulgaris (1% and 2%) in combination with vitamin E (250 mg/kg) in broiler diets exhibited the best prevention of lipid oxidation after 7 days of refrigerated storage, defined by the highest efficiency factors assessed in terms of secondary oxidation products.
4. Discussion
4.1. Nutritional Composition of C. vulgaris
C. vulgaris is considered a promising feed source due to its high growth rate and rich content of essential nutrients, particularly amino acids, fatty acids, and antioxidants. C. vulgaris has been used for partial dietary replacement of conventional protein sources like soybean meal, to enhance sustainability and lower the environmental impact of animal production [41]. In this study, a high protein level was assessed (around 50% DM), which was in line with the previously reported data [3]. The predominant fatty acids in C. vulgaris are the essential PUFAs 18:2n-6 and 18:3n-3, which have recognized positive effects on health for both humans and animals. C. vulgaris contains large amounts of various pigments, including chlorophylls a and b, as well as carotenoids such as β-carotene and lutein [8]. The total carotenoid content in C. vulgaris can vary, with levels reaching up to 3.49 g/kg DM. In this study, only the xanthophylls were assessed from the total carotenoids, since lutein is considered the predominant carotenoid in C. vulgaris [42]. A xanthophyll concentration of 0.98 g/kg DM was analyzed, which was below the concentration of total carotenoid content reported previously. It was shown that different carotenoid concentrations may be observed depending on the drying process, culturing conditions, and harvest time [43]. Additionally, the analytical results of this study have shown that C. vulgaris is a valuable source of vitamin E, as was also reported in [44]. These compounds possess significant antioxidant and radical scavenging properties [10].
4.2. Meat Quality Traits
Meat color is essential for the meat industry because it significantly shapes consumers’ perceptions of product quality and influences their purchasing decisions. In meat and meat products, color is a key parameter that consumers associate with quality, particularly in fresh meat products [45]. An elevated yellow color of thigh meat is preferred by consumers from various regions around the globe [46].
In the present study, dietary incorporation of C. vulgaris resulted in a minor increase in b*, C*, and h*, while a* registered a slight decrease when compared to the control group. The low L* values observed in the present study were comparable with the ones reported in [47], which registered an L* value of 32.53 and a pH of 6.62, in line with the results of the current study. Lower redness values are usually linked to higher concentrations of MetMb. Nevertheless, no differences in MetMb levels were observed between the groups that received dietary supplements, which can be explained by the minor variation in the color differences (ΔE*) calculated between the samples of the studied groups. A study conducted in [48] reported that color changes measured instrumentally are considered visible if ∆E > 2. In this study, the supplementation of broiler diets with 1% or 2% C. vulgaris, alone or in combination with vitamin E, resulted in no visible change of color, with a maximum ∆E of 1.72 observed in the E3 group. In contrast to the present study, [16] reported a yellower breast muscle (p < 0.0001) after the incorporation of a higher proportion of C. vulgaris (10%) in broilers’ diet compared to the present study (1% and 2% C. vulgaris), while the redness decreased with the addition of C. vulgaris in higher concentrations.
The postmortem pH value significantly affects color stability. Immediately after slaughter, meat has a pH of around 7, and it drops to approximately 5.5–5.8 due to glycolysis, increasing the likelihood of autoxidation [49]. In this study, pH registered higher values in meat from the experimental groups, with statistical significance only in groups that received C. vulgaris with vitamin E (E3 and E4 groups). Drip and cooking loss were not significantly changed by the 1% and 2% C. vulgaris inclusion levels, with or without vitamin E, but a numerical reduction in cooking loss was observed in groups fed C. vulgaris with vitamin E (E3 and E4 groups). In line with these results, [16] also reported no significant changes in drip loss and cooking loss in breast meat after 10 and 15% C. vulgaris dietary supplementation in broilers’ diets.
4.3. Proximate Composition of Meat
The inclusion of C. vulgaris in poultry diets has demonstrated a significant increase (p < 0.05) in the protein concentration of thigh meat, simultaneously with a significant decrease in the crude fat of thigh meat. This enhancement is thought to be attributed to Chlorella’s abundant amino acid profile and bioactive compounds, which support muscle growth and protein synthesis mechanisms. Similar results were reported by [21] who observed an increase in protein content in breast meat as a consequence of 15 and 20% incorporation of C. vulgaris in broiler’s diet. The same authors assumed that the increased protein content could be linked to the greater availability of amino acids from Chlorella proteins, enhanced nutrient absorption, or a combination of both. In another study [50], it was shown that an increase in the villus heights and crypt depths and, consequently, increased absorption area can be observed when supplementing broiler diets with 1% Chlorella by-products. Moreover, it has been reported that the inclusion of Chlorella in broilers’ diet can increase Lactobacillus spp. in broilers’ intestines, thus enhancing digestion and nutrient absorption [51]. In addition, it has been proposed that DHA supplementation enhances muscle protein synthesis in growing pigs [52], and can reduce lipid content in pig muscle [53]. This is consistent with the results of the current study, which showed significantly higher DHA levels in the thigh meat of the experimental groups fed C. vulgaris, compared to the control group.
The results of this study have shown a significantly lower fat content in the thigh muscle of broilers in the experimental groups compared to the control group, which was also reported by [21] after incorporation of 15 and 20% C. vulgaris in the broilers’ diet. A lower fat content in muscle can be attributed to dietary changes in fatty acids. Diets enriched with n-3 PUFAs are linked to decreased fat deposition [54]. In the current study, data regarding the fatty acid profile of C. vulgaris show that this microalga is a rich source of n-3 PUFAs, as [55] also reported.
4.4. Fatty Acid Profiling of Meat
The minor variations registered in SFA and MUFA concentrations in the thigh meat of broilers fed with diets supplemented with C. vulgaris, alone or with vitamin E, compared to the control group, were consistent with previously published reports [56]. In contrast, the concentrations of n-3 FA, PUFAs, and arachidonic acid increased in the thigh meat of broiler chickens that received C. vulgaris, when compared to the control group, results that align with previous findings in broiler chickens [57,58]. In this study, the DHA level registered a slight increase in the experimental groups, being significantly higher (p < 0.05) in the group that received 2% C. vulgaris and vitamin E.
C. vulgaris is a microalga that contains high levels of methionine and lysine. The authors of [59] suggested that the high level of n-3 fatty acids in the meat of broilers that received a diet supplemented with microalgae may be linked to its high methionine and lysine contents. The authors of [60] have shown that methionine and lysine can enhance the concentrations of n-3 fatty acids in broiler meat. Alongside the changes observed in the fatty acid profile, in this study, a significant reduction in total fat content was also noted in the thigh of broilers fed C. vulgaris. Methionine functions as a lipotropic agent by serving as a methyl donor for phospholipid synthesis and acting as a precursor for lipoproteins essential for lipid transport from the liver [61]. Lipid accumulation in tissues is mainly linked to lipid synthesis and breakdown, indicating that dietary methionine influences lipid metabolism in broilers.
The proportion of hypocholesterolemic to hypercholesterolemic fatty acids (DFA/OFA indexes) reveals the influence of particular fatty acids on cholesterol metabolism. Higher DFA/OFA values are deemed more favorable for human health [62]. The DFA/OFA indexes obtained in the current study ranged from 3.799 to 4.038 for the thigh meat of broilers that received diets with C. vulgaris, being higher compared to the control group which registered a value of 3.756.
4.5. Lipid Quality Indexes
AI and TI, which assess the impact of individual fatty acids on the likelihood of atheroma and thrombus formation, are additional markers relevant to consumer health. An AI above 1.0 is considered potentially harmful to health [63]. To our knowledge, there are no studies on the effects of a C. vulgaris supplementation in broilers’ diet on atherogenic and thrombogenic activities. The results of AI in the thigh meat of broilers obtained in the present study were beneficial for human health, regardless of dietary treatments applied. Nevertheless, the lowest significant values (p < 0.05) were noted in the meat from groups receiving 2% C. vulgaris (E2 and E4 groups). Similarly, significantly reduced TI values (p < 0.05) were observed in the thigh meat of broilers from the same experimental groups (E2 and E4).
The PI indicates the sensitivity of PUFAs to oxidation. Elevated PI levels correlate with increased fatty acid oxidation. Nevertheless, high PI levels resulting from substantial omega-3 and omega-6 PUFAs contribute to enhanced antioxidant and anti-inflammatory effects [64]. As expected, in the current study, the PI levels were increased, with significantly higher values being observed in the group with 2% C. vulgaris and vitamin E when compared to the control group. Moreover, the results showed that the DBI, IV, and Cox values were higher in the experimental groups, with significantly increased values when 2% C. vulgaris was included in the broilers’ diet (E2 and E4 groups). On the contrary, HPI and h/H registered the highest values as a result of 2% C. vulgaris supplementation, being linked with a positive effect in lowering cholesterol levels. These findings highlight the nutraceutical value of fatty acids from C. vulgaris, in agreement with the outcomes reported in [65].
4.6. Vitamin E Assessment in Meat
Vitamin E concentrations increased significantly (p < 0.05) in all the groups compared to the control group (C1), with the highest value being registered in the meat of broilers that received 2% C. vulgaris and vitamin E (E4 group). There is a notable lack of research on the impact of incorporating C. vulgaris into broiler diets on meat vitamin E levels. A study conducted in [55] investigated the effects of 10% C. vulgaris in broiler diets, alone or combined with exogenous CAZymes, on meat quality. Unlike our results, the dietary treatments did not affect vitamin E in breast meat. Moreover, in the thigh, the dietary supplementation with C. vulgaris, with and without exogenous CAZymes, decreased the levels of α-tocopherol and γ-tocopherol relative to the control. Supplementation of diets with high levels of C. vulgaris can lead to high amounts of carotenoids in meat [44,55], which can interfere with the absorption of vitamin E. Previous research indicated that α-tocopherol competes for absorption with other lipid micronutrients such as γ-tocopherol, carotenoids, and vitamins A, D, and K. These competitive interactions are probably caused by common uptake pathways [66]. The findings of the current study highlight that the inclusion of 1% and 2% C. vulgaris in broiler diets, alone or with vitamin E, enhanced the vitamin E content of chicken thighs, thereby providing additional benefits for consumers. Vitamin E plays a role in preventing lipid peroxidation by scavenging lipid peroxyl radicals, which otherwise propagate lipid peroxidation [67]. Its antioxidant capacity hinges on its ability to donate phenolic hydrogen to free radicals. Studies have demonstrated that supplementing the diet with vitamin E leads to an accumulation of α-tocopherol in muscle tissue, which delays the oxidation of deoxymyoglobin to metmyoglobin and consequently enhances color stability [68].
4.7. Oxidative Stability of Meat
Increasing α-tocopherol concentrations in thigh meat can significantly improve its antioxidant stability. Numerous studies have demonstrated that dietary supplementation with vitamin E reduces muscle membrane susceptibility to Fe2+-induced lipid oxidation [69,70].
Data regarding the markers of lipid oxidation in thigh meat revealed a significant decrease in TBARSs in the thigh tissues of birds supplemented with C. vulgaris compared to the control. This finding aligns with previous research highlighting the strong antioxidant capacity of Chlorella in broiler chickens [59]. TBARS levels exceeding 0.5 mg of malondialdehyde per kilogram of fresh meat are considered critical, as consumers can easily detect rancid off-flavors at this degree of lipid oxidation [71]. In the current study, TBARS levels were under this threshold in all the studied groups after 7 days of storage, with the highest TBARS concentration of 0.37 mg/kg registered in the thigh meat of the control group. Research on C. vulgaris supplementation in broiler chicks revealed that Chlorella increased total white blood cell count and hemoglobin levels and decreased the serum malondialdehyde concentration, further supporting its antioxidant properties [72]. Broilers fed with 20 g/kg Chlorella by-products exhibited increased serum superoxide dismutase activity and decreased serum malondialdehyde concentration, suggesting an improvement in the antioxidant status of broilers [50]. Dietary supplementation of Se-enriched Chlorella in broiler diets improved the oxidative stability of chicken meat during storage in a refrigerator, as indicated by reduced malondialdehyde values in breast meat after storage [73].
Lipid oxidation is directly linked to pigment oxidation. Deoxymyoglobin (DeoxyMb) is the purple pigment seen in freshly cut meat. After a few minutes of exposure to air, DeoxyMb becomes oxygenated, transforming into oxymyoglobin (OxyMb), which imparts the characteristic bright, cherry-red color. After several hours to days of air exposure, OxyMb converts to metmyoglobin (MetMb), where a water molecule replaces an oxygen molecule, resulting in brown pigmentation. Both DeoxyMb and OxyMb are heme proteins with iron in the ferrous (Fe+2) form, whereas MetMb contains iron in the ferric (Fe+3) form. The conversion from the ferrous to ferric form occurs due to oxidation [74]. In this study, the metmyoglobin percentage was positively influenced by the dietary treatments, showing significantly (p = 0.011) lower values in the experimental groups compared to the control group (C1). Moreover, there was a significant increase in the deoxymyoglobin percentage in all the experimental groups when referred to the control group (C1). On the other hand, no significant differences were found in the relative concentration of oxymyoglobin between the studied groups. Supplementation of diets with vitamin E has been shown to enhance meat quality by decreasing lipid oxidation in skeletal muscle. This, in turn, positively impacts meat discoloration by delaying the oxidation of myoglobin or oxymyoglobin to metmyoglobin, which leads to a brown color that is unappealing to consumers [75]. In this study, supplementation of broilers’ diet with C. vulgaris led to a decrease in redness, contrasting with the increasing effect on yellowness in thigh muscles. Generally, lower redness values are linked to higher concentrations of metmyoglobin. Nevertheless, in the current study, lower metmyoglobin levels were registered in the experimental groups. Beyond the impact of lipid peroxidation on meat flavor, color, and texture, lipid peroxidation poses a health risk when meat that has undergone this process is consumed, due to the autoxidation of unsaturated lipids and cholesterol that leads to the formation of atherogenic compounds [76].
5. Conclusions
The results from the current study demonstrate that C. vulgaris incorporation in broilers’ diet can enrich chicken meat with n-3 FA and vitamin E. Moreover, it can be considered an effective and practical approach to not only enhance the nutritional value and health benefits of meat lipids by decreasing the n-6/n-3 PUFA ratio but also to delay the meat’s susceptibility to lipid oxidation.
The current results may recommend supplementation with 2% Chlorella vulgaris and 250 ppm vitamin E in broiler chickens’ diet for meat quality improvement in terms of proximate composition, lipid quality, antioxidant status, and oxidative stability during storage in refrigerated conditions.
Author Contributions
Conceptualization, I.V. and A.E.U.; methodology, I.V., A.E.U. and T.D.P.; software, I.V. and A.E.U.; formal analysis, I.V., A.E.U., R.T., M.S., A.G.O. and P.A.V.; data curation, I.V.; writing—original draft preparation, I.V.; writing—review and editing, I.V. and A.E.U. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Romanian Ministry of Research, Innovation and Digitalization, Project PN 23.20.03.01.
Institutional Review Board Statement
The animal study protocol was approved by the Ethics Committee of the National Research and Development Institute of Animal Biology and Nutrition in Romania (protocol code no. 2761/03.03.2021 and 3 March 2021 of approval). The experimental procedures followed the guidelines specified in Romanian Law 43/2014 regarding the handling and protection of animals used for experimental purposes and complied with Directive 2010/63/EU on the protection of animals used for scientific purposes.
Informed Consent Statement
Not applicable.
Data Availability Statement
The original contributions presented in the study are included in the article; further inquiries can be directed to the corresponding author.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Shaikh, R.; Rizvi, A.; Pandit, S.; Desai, N.; Patil, R. Microalgae: Classification, bioactives, medicinal properties, industrial applications, and future prospectives. In An Integration of Phycoremediation Processes in Wastewater Treatment; Elsevier: Amsterdam, The Netherlands, 2022; pp. 451–486. [Google Scholar]
- Coronado-Reyes, J.A.; Salazar-Torres, J.A.; Juarez-Campos, B.; Gonzalez-Hernandez, J.C. Chlorella vulgaris, a microalgae important to be used in Biotechnology: A review. Food Sci. Technol. 2020, 42, e37320. [Google Scholar] [CrossRef]
- Canelli, G.; Tarnutzer, C.; Carpine, R.; Neutsch, L.; Bolten, C.J.; Dionisi, F.; Mathys, A. Biochemical and nutritional evaluation of Chlorella and Auxenochlorella biomasses relevant for food application. Front. Nutr. 2020, 7, 565996. [Google Scholar] [CrossRef] [PubMed]
- Spínola, M.P.; Costa, M.M.; Prates, J.A. Enhancing Digestibility of Chlorella vulgaris Biomass in Monogastric Diets: Strategies and Insights. Animals 2023, 13, 1017. [Google Scholar] [CrossRef] [PubMed]
- Georgiopoulou, I.; Tzima, S.; Pappa, G.D.; Louli, V.; Voutsas, E.; Magoulas, K. Experimental design and optimization of recovering bioactive compounds from chlorella vulgaris through conventional extraction. Molecules 2021, 27, 29. [Google Scholar] [CrossRef]
- Dineshbabu, G.; Goswami, G.; Kumar, R.; Sinha, A.; Das, D. Microalgae–nutritious, sustainable aqua-and animal feed source. J. Funct. Foods 2019, 62, 103545. [Google Scholar] [CrossRef]
- Bhosale, P. Environmental and cultural stimulants in the production of carotenoids from microorganisms. Appl. Microbiol. Biotechnol. 2004, 63, 351–361. [Google Scholar] [CrossRef] [PubMed]
- Safi, C.; Zebib, B.; Merah, O.; Pontalier, P.Y.; Vaca-Garcia, C. Morphology, composition, production, processing and applications of Chlorella vulgaris: A review. Renew. Sustain. Energy Rev. 2014, 35, 265–278. [Google Scholar] [CrossRef]
- Zhang, T.; Zhang, R.; Zhao, G.; Liu, W.; Pan, L.; Tong, Y.; Jiang, M.; Zhang, H.; Xiao, Z.; Pandol, S.J.; et al. Plant green pigment of chlorophyllin attenuates inflammatory bowel diseases by suppressing autophagy activation in mice. Am. J. Physiol. Gastrointest. Liver Physiol. 2022, 323, G102–G113. [Google Scholar] [CrossRef]
- Pérez-Gálvez, A.; Viera, I.; Roca, M. Carotenoids and chlorophylls as antioxidants. Antioxidants 2020, 9, 505. [Google Scholar] [CrossRef]
- Ramos-Romero, S.; Torrella, J.R.; Pagès, T.; Viscor, G.; Torres, J.L. Edible microalgae and their bioactive compounds in the prevention and treatment of metabolic alterations. Nutrients 2021, 13, 563. [Google Scholar] [CrossRef] [PubMed]
- de Medeiros, V.P.B.; Pimentel, T.C.; Sant’Ana, A.S.; Magnani, M. Microalgae in the meat processing chain: Feed for animal production or source of techno-functional ingredients. Curr. Opin. Food Sci. 2021, 37, 125–134. [Google Scholar] [CrossRef]
- Tan, J.S.; Lee, S.Y.; Chew, K.W.; Lam, M.K.; Lim, J.W.; Ho, S.H.; Show, P.L. A review on microalgae cultivation and harvesting, and their biomass extraction processing using ionic liquids. Bioeng. 2020, 11, 116–129. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.; Wang, J.; Chen, P.; Ji, C.; Kang, Q.; Lu, B.; Li, K.; Liu, J.; Ruan, R. Bio-mitigation of carbon dioxide using microalgal systems: Advances and perspectives. Renew. Sustain. Energy Rev. 2017, 76, 1163–1175. [Google Scholar] [CrossRef]
- Kaur, M.; Bhatia, S.; Gupta, U.; Decker, E.; Tak, Y.; Bali, M.; Gupta, V.K.; Dar, R.A.; Bala, S. Microalgal bioactive metabolites as promising implements in nutraceuticals and pharmaceuticals: Inspiring therapy for health benefits. Phytochem. Rev. 2023, 22, 903–933. [Google Scholar] [CrossRef] [PubMed]
- Boskovic Cabrol, M.B.; Martins, J.C.; Malhão, L.P.; Alves, S.P.; Bessa, R.J.; Almeida, A.M.; Raymundo, A.; Lordelo, M. Partial replacement of soybean meal with Chlorella vulgaris in broiler diets influences performance and improves breast meat quality and fatty acid composition. Poult. Sci. 2022, 101, 101955. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Wareth, A.A.; Williams, A.N.; Salahuddin, M.; Gadekar, S.; Lohakare, J. Algae as an alternative source of protein in poultry diets for sustainable production and disease resistance: Present status and future considerations. Front. Vet. Sci. 2024, 11, 1382163. [Google Scholar] [CrossRef] [PubMed]
- Parisi, G.; Tulli, F.; Fortina, R.; Marino, R.; Bani, P.; Dalle Zotte, A.; De Angelis, A.; Piccolo, G.; Pinotti, L.; Schiavone, A.; et al. Protein hunger of the feed sector: The alternatives offered by the plant world. Ital. J. Anim. Sci. 2020, 19, 1204–1225. [Google Scholar] [CrossRef]
- Lee, J.Y.; Yoon, J.H.; An, S.H.; Cho, I.H.; Lee, C.W.; Jeon, Y.J.; Joo, S.S.; Ban, B.C.; Lee, J.-Y.; Jung, H.J.; et al. Intestinal Immune Cell Populations, Barrier Function, and Microbiomes in Broilers Fed a Diet Supplemented with Chlorella vulgaris. Animals 2023, 13, 2380. [Google Scholar] [CrossRef]
- An, B.K.; Kim, K.E.; Jeon, J.Y.; Lee, K.W. Effect of dried Chlorella vulgaris and Chlorella growth factor on growth performance, meat qualities and humoral immune responses in broiler chickens. Springerplus 2016, 5, 718. [Google Scholar] [CrossRef]
- Boskovic Cabrol, M.; Martins, J.C.; Malhão, L.P.; Alfaia, C.M.; Prates, J.A.; Almeida, A.M.; Lordelo, M.; Raymundo, A. Digestibility of meat mineral and proteins from broilers fed with graded levels of Chlorella vulgaris. Foods 2022, 11, 1345. [Google Scholar] [CrossRef]
- Zhang, L.; Sun, B.; Xie, P.; Li, H.; Su, H.; Sha, K.; Huang, C.; Lei, Y.; Liu, X.; Wang, H. Using near infrared spectroscopy to predict the physical traits of Bos grunniens meat. LWT-Food Sci. Technol 2015, 64, 602–608. [Google Scholar] [CrossRef]
- Oliveira, R.F.D.; Mello, J.L.M.D.; Ferrari, F.B.; Souza, R.A.D.; Pereira, M.R.; Cavalcanti, E.N.F.; Villegas-Cayllahua, E.A.; Fidelis, H.d.A.; Giampietro-Ganeco, A.; Fávero, M.S.; et al. Effect of aging on the quality of breast meat from broilers affected by wooden breast myopathy. Animals 2021, 11, 1960. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Yan, X.; Wang, L.; Zhu, R.; Chen, M.; Yin, J.; Zhang, X. Insights into the mechanism of L-malic acid on drip loss of chicken meat under commercial conditions. J. Anim. Sci. Biotechnol. 2024, 15, 14. [Google Scholar] [CrossRef] [PubMed]
- ISO 5983-2:2009; Animal Feeding Stuffs—Determination of Nitrogen Content and Calculation of Crude Protein Content. Part 2: Block Digestion and Steam Distillation Method. ISO: Geneva, Switzerland, 2009.
- ISO 6492:2001; Animal Feeding Stuffs—Determination of Fat Content. ISO: Geneva, Switzerland, 2001.
- ISO 6496:2001; Animal Feeding Stuffs—Determination of Moisture and Other Volatile Matter Content. ISO: Geneva, Switzerland, 2001.
- ISO, SREN. 2171:2010; Cereals. Legumes and Derived Products—Determination of Ash Content via Calcination. ISO: Geneva, Switzerland, 2010.
- Turcu, R.P.; Panaite, T.D.; Untea, A.E.; Vlaicu, P.A.; Badea, I.A.; Mironeasa, S. Effects of grape seed oil supplementation to broilers diets on growth performance, meat fatty acids, health lipid indices and lipid oxidation parameters. Agriculture 2021, 11, 404. [Google Scholar] [CrossRef]
- Untea, A.E.; Varzaru, I.; Panaite, T.D.; Gavris, T.; Lupu, A.; Ropota, M. The effects of dietary inclusion of bilberry and walnut leaves in laying hens’ diets on the antioxidant properties of eggs. Animals 2020, 10, 191. [Google Scholar] [CrossRef] [PubMed]
- Wereńska, M.; Wołoszyn, J.; Okruszek, A.; Marcinkowska, W.; Haraf, G. The effects of sous-vide, microwave cooking and stewing of goose meat on fatty acid profile and lipid indices. Poult Sci. 2023, 102, 102337. [Google Scholar] [CrossRef]
- Chen, J.; Liu, H. Nutritional indices for assessing fatty acids: A mini-review. Int. J. Mol. Sci. 2020, 21, 5695. [Google Scholar] [CrossRef]
- Cavaco, A.R.; Laureano, G.; Duarte, B.; da Silva, J.M.; Gameiro, C.; Cunha, J.; Dias, J.E.; Matos, A.R.; Figueiredo, A. First assessment of leaf lipids and fatty acids as biomarkers of grapevine tolerance/susceptibility to Plasmopara viticola. Physiol. Mol. Plant Pathol. 2023, 124, 101948. [Google Scholar] [CrossRef]
- Cecchi, T.; Passamonti, P.; Alfei, B.; Cecchi, P. Monovarietal extra virgin olive oils from the Marche region, Italy: Analytical and sensory characterization. Int. J. Food Prop. 2011, 14, 483–495. [Google Scholar] [CrossRef]
- Kotsou, K.; Chatzimitakos, T.; Athanasiadis, V.; Bozinou, E.; Rumbos, C.I.; Athanassiou, C.G.; Lalas, S.I. Enhancing the Nutritional Profile of Tenebrio molitor Using the Leaves of Moringa oleifera. Foods 2023, 12, 2612. [Google Scholar] [CrossRef]
- Varzaru, I.; Untea, A.E.; Saracila, M. In vitro antioxidant properties of berry leaves and their inhibitory effect on lipid peroxidation of thigh meat from broiler chickens. Eur. J. Lipid Sci. Technol. 2020, 122, 1900384. [Google Scholar] [CrossRef]
- Untea, A.E.; Oancea, A.G.; Vlaicu, P.A.; Varzaru, I.; Saracila, M. Blackcurrant (Fruits, Pomace, and Leaves) Phenolic Characterization before and after In Vitro Digestion, Free Radical Scavenger Capacity, and Antioxidant Effects on Iron-Mediated Lipid Peroxidation. Foods 2024, 13, 1514. [Google Scholar] [CrossRef] [PubMed]
- Saracila, M.; Untea, A.E.; Varzaru, I.; Panaite, T.D.; Vlaicu, P.A. Comparative effects on using bilberry leaves in broiler diet reared under thermoneutral conditions vs. heat stress on performance, health status and gut microbiota. Life 2023, 14, 39. [Google Scholar] [CrossRef] [PubMed]
- Untea, A.E.; Panaite, T.D.; Dragomir, C.; Ropota, M.; Olteanu, M.; Varzaru, I. Effect of dietary chromium supplementation on meat nutritional quality and antioxidant status from broilers fed with Camelina-meal-supplemented diets. Animal 2019, 13, 2939–2947. [Google Scholar] [CrossRef] [PubMed]
- Bak, K.H.; Bauer, S.; Bauer, F. Effect of different genotypes and harvest times of sage (Salvia spp. labiatae) on lipid oxidation of cooked meat. Antioxidants 2023, 12, 616. [Google Scholar] [CrossRef] [PubMed]
- Almomani, F.; Hosseinzadeh-Bandbafha, H.; Aghbashlo, M.; Omar, A.; Joo, S.W.; Vasseghian, Y.; Karimi-Maleh, H.; Lam, S.S.; Tabatabaei, M.; Rezania, S. Comprehensive insights into conversion of microalgae to feed, food, and biofuels: Current status and key challenges towards implementation of sustainable biorefineries. J. Chem. Eng. 2023, 455, 140588. [Google Scholar] [CrossRef]
- Miyazawa, T.; Nakagawa, K.; Kimura, F.; Nakashima, Y.; Maruyama, I.; Higuchi, O.; Miyazawa, T. Chlorella is an effective dietary source of lutein for human erythrocytes. J. Oleo Sci. 2013, 62, 773–779. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, M.T.; Shariff, M.; Yusoff, F.M.; Goh, Y.M.; Banerjee, S. Applications of microalga Chlorella vulgaris in aquaculture. Rev. Aquacult. 2020, 12, 328–346. [Google Scholar] [CrossRef]
- Coelho, D.; Pestana, J.; Almeida, J.M.; Alfaia, C.M.; Fontes, C.M.; Moreira, O.; Prates, J.A. A high dietary incorporation level of Chlorella vulgaris improves the nutritional value of pork fat without impairing the performance of finishing pigs. Animals 2020, 10, 2384. [Google Scholar] [CrossRef]
- Fletcher, D.L. Poultry meat quality. J. World’s Poult. Sci. 2002, 58, 131–145. [Google Scholar] [CrossRef]
- Grashorn, M. Feed additives for influencing chicken meat and egg yolk color. In Handbook on Natural Pigments in Food and Beverages; Woodhead Publishing: Cambridge, UK, 2016; pp. 283–302. [Google Scholar]
- Wattanachant, S.; Benjakul, S.; Ledward, D.A. Composition, colour, and texture of Thai indigenous and broiler chicken muscles. Poult. Sci. 2004, 83, 123–128. [Google Scholar] [CrossRef] [PubMed]
- Savadkoohi, S.; Hoogenkamp, H.; Shamsi, K.; Farahnaky, A. Color, sensory and textural attributes of beef frankfurter, beef ham and meat-free sausage containing tomato pomace. Meat Sci. 2014, 97, 410–418. [Google Scholar] [CrossRef]
- Bischof, G.; Witte, F.; Terjung, N.; Heinz, V.; Juadjur, A.; Gibis, M. Metabolic, proteomic and microbial changes postmortem and during beef aging. Crit. Rev. Food Sci. Nutr. 2024, 64, 1076–1109. [Google Scholar] [CrossRef] [PubMed]
- Mirzaie, S.; Sharifi, S.D.; Zirak-Khattab, F. The effect of a Chlorella by-product dietary supplement on immune response, antioxidant status, and intestinal mucosal morphology of broiler chickens. J. Appl. Phycol. 2020, 32, 1771–1777. [Google Scholar] [CrossRef]
- Kang, H.K.; Park, S.B.; Kim, C.H. Effects of dietary supplementation with a chlorella by-product on the growth performance, immune response, intestinal microflora and intestinal mucosal morphology in broiler chickens. J. Anim. Physiol. Anim. Nutr. 2017, 101, 208–214. [Google Scholar] [CrossRef]
- Wei, H.K.; Zhou, Y.; Jiang, S.; Tao, Y.X.; Sun, H.; Peng, J.; Jiang, S. Feeding a DHA-enriched diet increases skeletal muscle protein synthesis in growing pigs: Association with increased skeletal muscle insulin action and local mRNA expression of insulin-like growth factor 1. Br. J. Nutr. 2013, 110, 671–680. [Google Scholar] [CrossRef]
- De Tonnac, A.; Labussière, E.; Vincent, A.; Mourot, J. Effect of α-linolenic acid and DHA intake on lipogenesis and gene expression involved in fatty acid metabolism in growing-finishing pigs. Br. J. Nutr. 2016, 116, 7–18. [Google Scholar] [CrossRef]
- Kim, M.; Voy, B.H. Fighting fat with fat: N-3 polyunsaturated fatty acids and adipose deposition in broiler chickens. Front. physiol. 2021, 12, 755317. [Google Scholar] [CrossRef] [PubMed]
- Alfaia, C.M.; Pestana, J.M.; Rodrigues, M.; Coelho, D.; Aires, M.J.; Ribeiro, D.M.; Major, V.T.; Martins, C.F.; Santos, H.; Lopes, P.A.; et al. Influence of dietary Chlorella vulgaris and carbohydrate-active enzymes on growth performance, meat quality and lipid composition of broiler chickens. Poult. Sci. 2021, 100, 926–937. [Google Scholar] [CrossRef]
- Madeira, M.S.; Cardoso, C.; Lopes, P.A.; Coelho, D.; Afonso, C.; Bandarra, N.M.; Prates, J.A. Microalgae as feed ingredients for livestock production and meat quality: A review. Livest. Sci. 2017, 205, 111–121. [Google Scholar] [CrossRef]
- Moran, C.A.; Currie, D.; Keegan, J.D.; Knox, A. Tolerance of Broilers to Dietary Supplementation with High Levels of the DHA-Rich Microalga, Aurantiochytrium Limacinum: Effects on Health and Productivity. Animals 2018, 8, 180. [Google Scholar] [CrossRef] [PubMed]
- Yan, L.; Kim, I.H. Effects of dietary ω-3 fatty acid-enriched microalgae supplementation on growth performance, blood profiles, meat quality, and fatty acid composition of meat in broilers. J. Appl. Anim. Res. 2013, 41, 392–397. [Google Scholar] [CrossRef]
- El-Bahr, S.; Shousha, S.; Shehab, A.; Khattab, W.; Ahmed-Farid, O.; Sabike, I.; El-Garhy, O.; Albokhadaim, I.; Albosadah, K. Effect of dietary microalgae on growth performance, profiles of amino and fatty acids, antioxidant status, and meat quality of broiler chickens. Animals 2020, 10, 761. [Google Scholar] [CrossRef] [PubMed]
- Moghadam, M.B.; Shehab, A.; Cherian, G. Methionine supplementation augments tissue n-3 fatty acid and tocopherol content in broiler birds fed flaxseed. Anim. Feed Sci. Technol. 2017, 228, 149–158. [Google Scholar] [CrossRef]
- Huang, T.; Wahlqvist, M.L.; Li, D. Effect of n-3 polyunsaturated fatty acid on gene expression of the critical enzymes involved in homocysteine metabolism. Nutr. J. 2012, 11, 6. [Google Scholar] [CrossRef] [PubMed]
- Wereńska, M.; Haraf, G.; Wołoszyn, J.; Goluch, Z.; Okruszek, A.; Teleszko, M. Fatty acid profile and health lipid indices of goose meat in relation to various types of heat treatment. Poult. Sci. 2021, 100, 101237. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, R.; Lourenço, S.; Lopes, A.; Andrade, C.; Câmara, J.S.; Castilho, P.; Perestrelo, R. Evaluation of fatty acids profile as a useful tool towards valorization of by-products of agri-food industry. Foods 2021, 10, 2867. [Google Scholar] [CrossRef] [PubMed]
- Dawczynski, C.; Hackermeier, U.; Viehweger, M.; Stange, R.; Springer, M.; Jahreis, G. Incorporation of n-3 PUFA and γ-linolenic acid in blood lipids and red blood cell lipids together with their influence on disease activity in patients with chronic inflammatory arthritis-a randomized controlled human intervention trial. Lipids Health Dis. 2011, 10, 130. [Google Scholar] [CrossRef] [PubMed]
- Maurício, T.; Couto, D.; Lopes, D.; Conde, T.; Pais, R.; Batista, J.; Melo, T.; Pinho, M.; Moreira, A.S.P.; Trovão, M.; et al. Differences and similarities in lipid composition, nutritional value, and bioactive potential of four edible Chlorella vulgaris strains. Foods 2023, 12, 1625. [Google Scholar] [CrossRef]
- Reboul, E. Vitamin E bioavailability: Mechanisms of intestinal absorption in the spotlight. Antioxidants 2017, 6, 95. [Google Scholar] [CrossRef]
- Das, K. Ascorbate and Tocopherols in Mitigating Oxidative Stress. In Protective Chemical Agents in the Amelioration of Plant Abiotic Stress: Biochemical and Molecular Perspectives; John Wiley & Sons: Hoboken, NJ, USA, 2020; pp. 102–121. [Google Scholar]
- Lahučký, R.; Novotná, K.; Zaujec, K.; Mojto, J.; Pavlič, M.; Roa, N.B. Effects of dietary vitamin E supplementation on α-tocopherol content and antioxidative status of beef muscles. Czech J. Anim. Sci. 2002, 47, 381–386. [Google Scholar]
- Delosière, M.; Durand, D.; Bourguet, C.; Terlouw, E.C. Lipid oxidation, pre-slaughter animal stress and meat packaging: Can dietary supplementation of vitamin E and plant extracts come to the rescue? Food Chem. 2020, 309, 125668. [Google Scholar] [CrossRef] [PubMed]
- Lahucky, R.; Nuernberg, K.; Kovac, L.; Bucko, O.; Nuernberg, G. Assessment of the antioxidant potential of selected plant extracts–in vitro and in vivo experiments on pork. Meat Sci. 2010, 85, 779–784. [Google Scholar] [CrossRef]
- Wood, J.D.; Enser, M.; Fisher, A.V.; Nute, G.R.; Sheard, P.R.; Richardson, R.I.; Hugher, S.I.; Whittington, F.M. Fat deposition, fatty acid composition and meat quality: A review. Meat Sci. 2008, 78, 343–358. [Google Scholar] [CrossRef]
- Niamat, M.; Ragaa, E.-A.; Hamouda, A.; Dawoud, G.T.M. Impacts of chlorella vulgaris supplementation to chicken drinking water on amino acids, fatty acids, minerals content of broiler chicken meats. Egypt. J. Nutr. Feed 2018, 21, 509–518. [Google Scholar] [CrossRef]
- Dlouhá, G.; Sevcikova, S.; Dokoupilova, A.; Zita, L.; Heindl, J.; Skrivan, M. Effect of dietary selenium sources on growth performance, breast muscle selenium, glutathione peroxidase activity and oxidative stability in broilers. Czech J. Anim. Sci. 2008, 53, 265. [Google Scholar] [CrossRef]
- Ruedt, C.; Gibis, M.; Weiss, J. Meat color and iridescence: Origin, analysis, and approaches to modulation. Compr. Rev. Food Sci. Food Saf. 2023, 22, 3366–3394. [Google Scholar] [CrossRef]
- Salami, S.A.; Majoka, M.A.; Saha, S.; Garber, A.; Gabarrou, J.F. Efficacy of dietary antioxidants on broiler oxidative stress, performance and meat quality: Science and market. Avian Biol. Res. 2015, 8, 65–78. [Google Scholar] [CrossRef]
- Wang, D.; Xiao, H.; Lyu, X.; Chen, H.; Wei, F. Lipid oxidation in food science and nutritional health: A comprehensive review. Oil Crop Sci. 2023, 8, 35–44. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).