
Volatile Organic Compounds of Scheffersomyces spartinae W9 Have Antifungal Effect against Botrytis cinerea on Strawberry Fruit


Abstract
:1. Introduction
2. Materials and Methods
2.1. Fruit
2.2. Pathogen and the Yeast
2.3. Effects of VOCs Produced by S. spartinae W9 on Mycelium Growth and Spore Germination of B. cinerea
2.4. Effect of VOCs on Strawberry Gray Mold
2.5. Microscopic Observation of Hyphae Morphology and Ultrastructure
2.6. Identification of VOC Components
2.7. Effect of Selected Individual VOCs on Mycelial Growth of B. cinerea
2.8. Statistical Analysis
3. Results
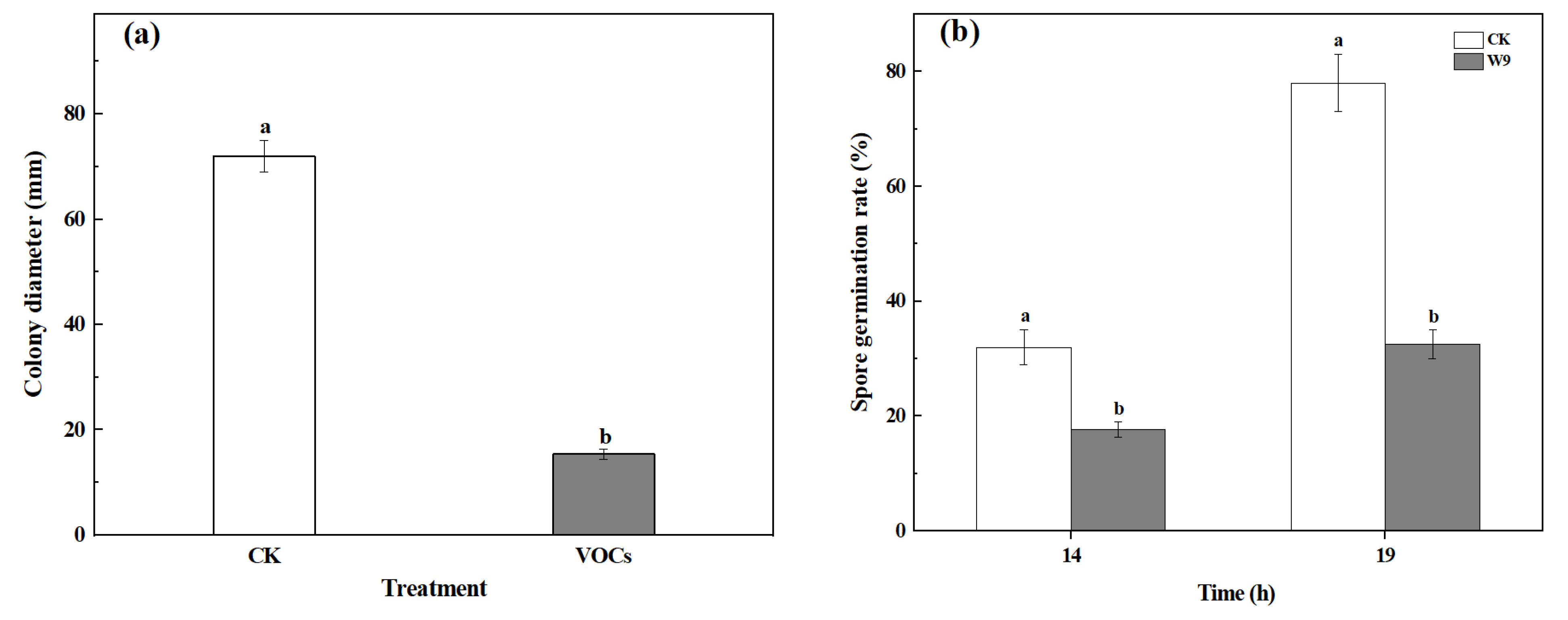
3.1. VOCs Produced by S. spartinae W9 Inhibited Mycelium Growth and Spore Germination of B. cinerea
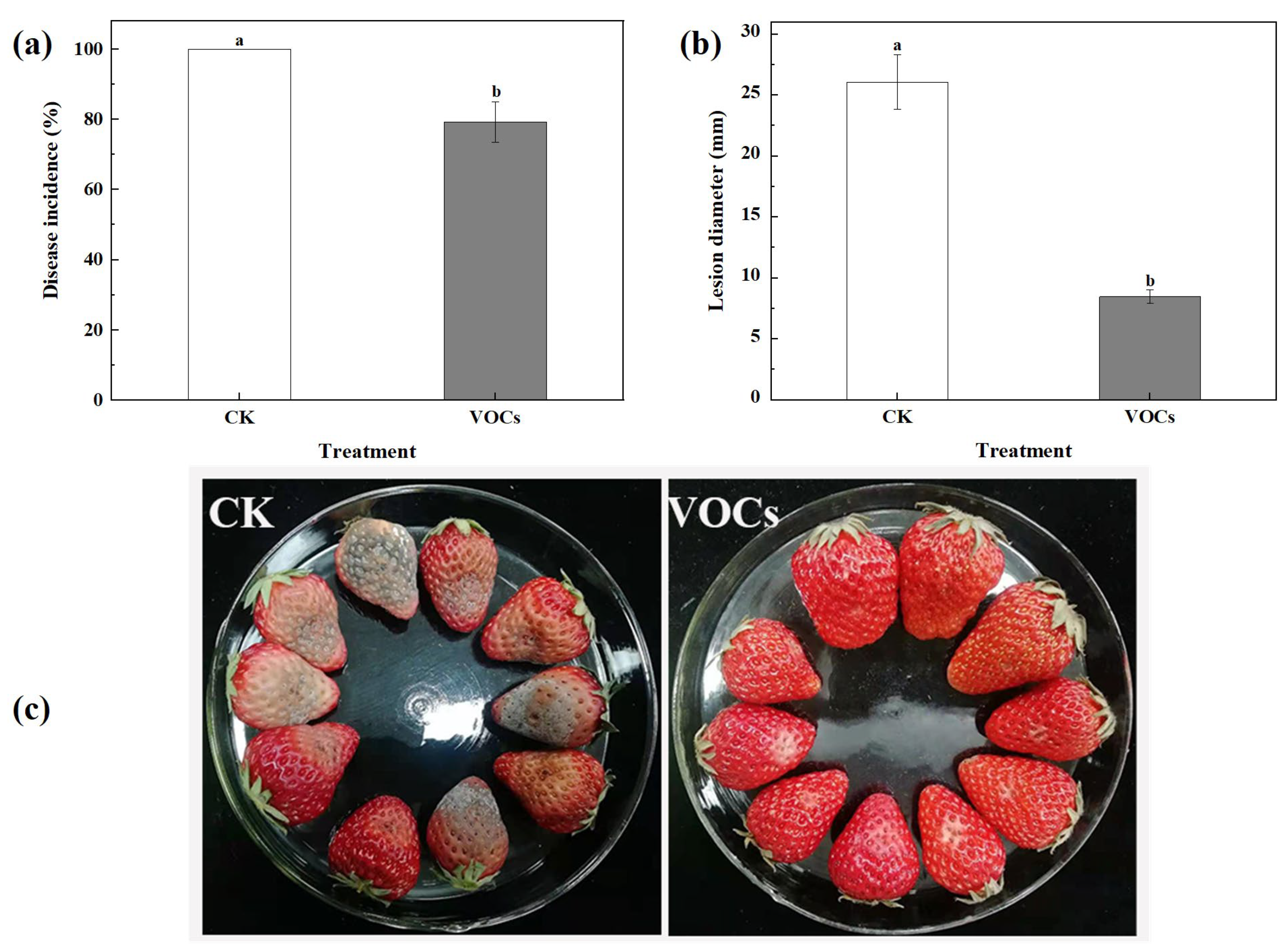
3.2. VOCs Produced by S. spartinae W9 Suppressed Strawberry Gray Mold
3.3. VOCs produced by S. spartinae W9 Destroyed the Mycelium Surface Morphology of B. cinerea
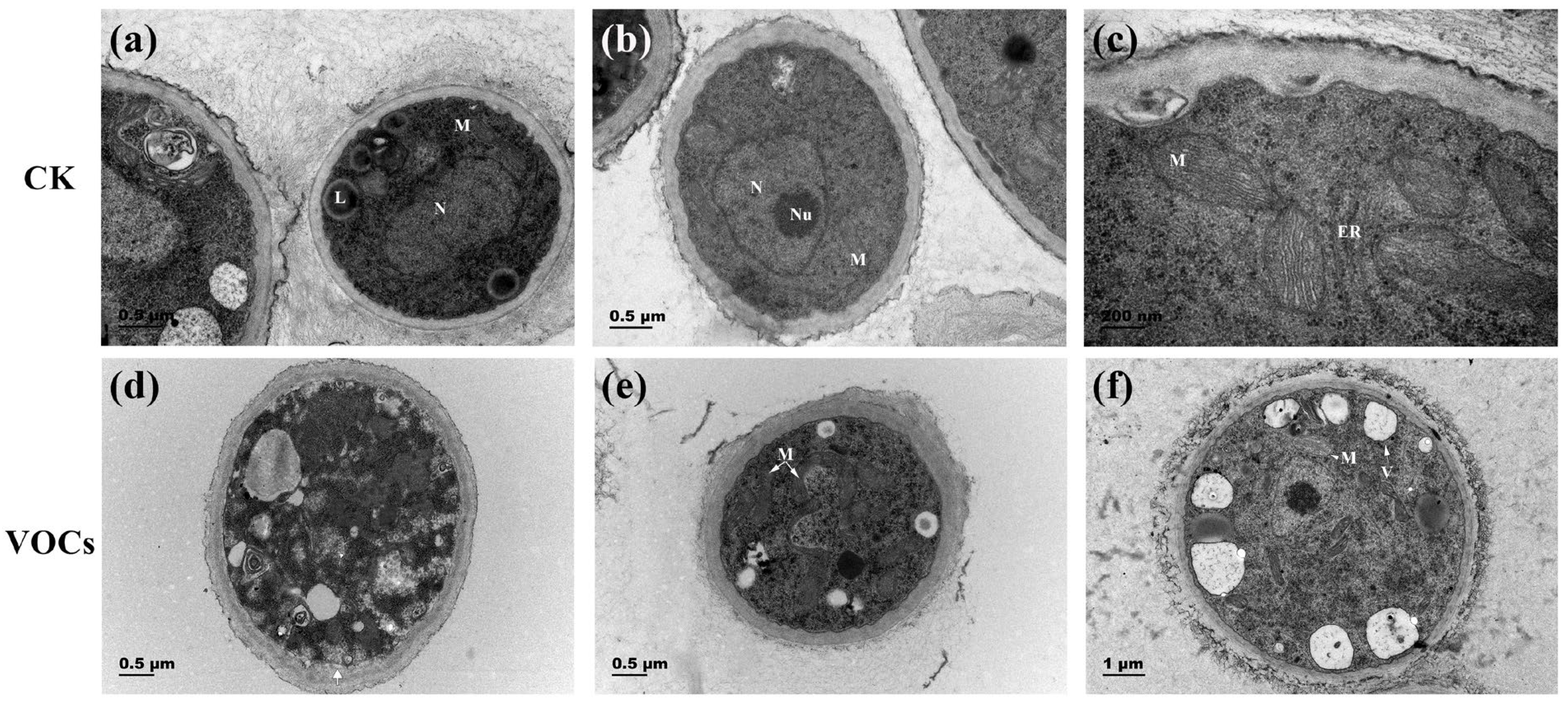
3.4. VOCs Produced by S. spartinae W9 Damaged the Cell Ultrastructure of B. cinerea
3.5. Composition of VOCs Produced by S. spartinae W9
3.6. Antifungal Effect of Main VOCs Produced by S. spartinae W9
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Dukare, A.S.; Paul, S.; Nambi, V.E.; Gupta, R.K.; Singh, R.; Sharma, K.; Vishwakarma, R.K. Exploitation of microbial antagonists for the control of postharvest diseases of fruits: A review. Crit. Rev. Food Sci. Nutr. 2019, 59, 1498–1513. [Google Scholar] [CrossRef] [PubMed]
- Freimoser, F.M.; Rueda-Mejia, M.P.; Tilocca, B.; Migheli, Q. Biocontrol yeasts: Mechanisms and applications. World J. Microbiol. Biotechnol. 2019, 35, 154. [Google Scholar] [CrossRef] [PubMed]
- Farbo, M.G.; Urgeghe, P.P.; Fiori, S.; Marcello, A.; Oggiano, S.; Balmas, V.; Hassan, Z.U.; Jaoua, S.; Migheli, Q. Effect of yeast volatile organic compounds on ochratoxin A-producing Aspergillus carbonarius and A. ochraceus. Int. J. Food Microbiol. 2018, 284, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Oro, L.; Feliziani, E.; Ciani, M.; Romanazzi, G.; Comitini, F. Volatile organic compounds from Wickerhamomyces anomalus, Metschnikowia pulcherrima and Saccharomyces cerevisiae inhibit growth of decay causing fungi and control postharvest diseases of strawberries. Int. J. Food Microbiol. 2018, 265, 18–22. [Google Scholar] [CrossRef] [PubMed]
- Parafati, L.; Vitale, A.; Restuccia, C.; Cirvilleri, G. Performance evaluation of volatile organic compounds by antagonistic yeasts immobilized on hydrogel spheres against gray, green and blue postharvest decays. Food Microbiol. 2017, 63, 191–198. [Google Scholar] [CrossRef] [PubMed]
- Pereyra, M.M.; Garmendia, G.; Rossini, C.; Meinhardt, F.; Vero, S.; Dib, J.R. Volatile organic compounds of Clavispora lusitaniae AgL21 restrain citrus postharvest pathogens. Biol. Control 2022, 174, 105025. [Google Scholar] [CrossRef]
- Di Francesco, A.; Di Foggia, M.; Baraldi, E. Aureobasidium pullulans volatile organic compounds as alternative postharvest method to control brown rot of stone fruits. Food Microbiol. 2020, 87, 103395. [Google Scholar] [CrossRef]
- Rios-Navarro, A.; Gonzalez, M.; Carazzone, C.; Celis Ramirez, A.M. Learning about microbial language: Possible interactions mediated by microbial volatile organic compounds (VOCs) and relevance to understanding Malassezia spp. metabolism. Metabolomics 2021, 17, 39. [Google Scholar] [CrossRef]
- Oufensou, S.; Ul Hassan, Z.; Balmas, V.; Jaoua, S.; Migheli, Q. Perfume guns: Potential of yeast volatile organic compounds in the biological control of mycotoxin-producing fungi. Toxins 2023, 15, 45. [Google Scholar] [CrossRef]
- Tilocca, B.; Cao, A.C.; Migheli, Q. Scent of a killer: Microbial volatilome and its role in the biological control of plant pathogens. Front. Microbiol. 2020, 11, 41. [Google Scholar] [CrossRef]
- Nasanit, R.; Jaibangyang, S.; Onwibunsiri, T.; Khunnamwong, P. Screening of volatile organic compound-producing yeasts and yeast-like fungi against aflatoxigenic Aspergillus flavus. Microbiol. Biotechnol. Lett. 2022, 50, 202–210. [Google Scholar] [CrossRef]
- Fan, L.; Wei, Y.; Chen, Y.; Jiang, S.; Xu, F.; Zhang, C.; Wang, H.; Shao, X. Epinecidin-1, a marine antifungal peptide, inhibits Botrytis cinerea and delays gray mold in postharvest peaches. Food Chem. 2023, 403, 134419. [Google Scholar] [CrossRef] [PubMed]
- Zou, X.; Wei, Y.; Dai, K.; Xu, F.; Wang, H.; Shao, X. Yeasts from intertidal zone marine sediment demonstrate antagonistic activities against Botrytis cinerea in vitro and in strawberry fruit. Biol. Control 2021, 158, 104612. [Google Scholar] [CrossRef]
- Zou, X.; Wei, Y.; Jiang, S.; Cao, Z.; Xu, F.; Wang, H.; Zhan, P.; Shao, X. Volatile organic compounds and rapid proliferation of Candida pseudolambica W16 are modes of action against gray mold in peach fruit. Postharvest Biol. Technol. 2022, 183, 111751. [Google Scholar] [CrossRef]
- Xu, Y.; Wei, J.; Wei, Y.; Han, P.; Dai, K.; Zou, X.; Jiang, S.; Xu, F.; Wang, H.; Sun, J.; et al. Tea tree oil controls brown rot in peaches by damaging the cell membrane of Monilinia fructicola. Postharvest Biol. Technol. 2021, 175, 111474. [Google Scholar] [CrossRef]
- Shao, X.; Cao, B.; Xu, F.; Xie, S.; Yu, D.; Wang, H. Effect of postharvest application of chitosan combined with clove oil against citrus green mold. Postharvest Biol. Technol. 2015, 99, 37–43. [Google Scholar] [CrossRef]
- Yalage Don, S.M.; Schmidtke, L.M.; Gambetta, J.M.; Steel, C.C. Aureobasidium pullulans volatilome identified by a novel, quantitative approach employing SPME-GC-MS, suppressed Botrytis cinerea and Alternaria alternata in vitro. Sci. Rep. 2020, 10, 4498. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.; Liu, X.-B.; Zhu, W.-W.; Zhong, X.; Sun, Q.; Liang, Y.-Z. Retention Indices for Identification of Aroma Compounds by GC: Development and Application of a Retention Index Database. Chromatographia 2014, 78, 89–108. [Google Scholar] [CrossRef]
- Lyu, A.; Yang, L.; Wu, M.; Zhang, J.; Li, G. High efficacy of the volatile organic compounds of Streptomyces yanglinensis 3–10 in suppression of Aspergillus contamination on peanut kernels. Front. Microbiol. 2020, 11, 142. [Google Scholar] [CrossRef]
- Xing, M.; Zheng, L.; Deng, Y.; Xu, D.; Xi, P.; Li, M.; Kong, G.; Jiang, Z. Antifungal activity of natural volatile organic compounds against litchi downy blight pathogen Peronophythora litchii. Molecules 2018, 23, 358. [Google Scholar] [CrossRef]
- Wang, Z.; Zhong, T.; Chen, K.; Du, M.; Chen, G.; Chen, X.; Wang, K.; Zalán, Z.; Takács, K.; Kan, J. Antifungal activity of volatile organic compounds produced by Pseudomonas fluorescens ZX and potential biocontrol of blue mold decay on postharvest citrus. Food Control 2021, 120, 107499. [Google Scholar] [CrossRef]
- Arrarte, E.; Garmendia, G.; Rossini, C.; Wisniewski, M.; Vero, S. Volatile organic compounds produced by Antarctic strains of Candida sake play a role in the control of postharvest pathogens of apples. Biol. Control 2017, 109, 14–20. [Google Scholar] [CrossRef]
- Qin, X.; Xiao, H.; Cheng, X.; Zhou, H.; Si, L. Hanseniaspora uvarum prolongs shelf life of strawberry via volatile production. Food Microbiol. 2017, 63, 205–212. [Google Scholar] [CrossRef] [PubMed]
- Lemos, W.J.F.; Binati, R.L.; Felis, G.E.; Slaghenaufi, D.; Ugliano, M.; Torriani, S. Volatile organic compounds from Starmerella bacillaris to control gray mold on apples and modulate cider aroma profile. Food Microbiol. 2020, 89, 103446. [Google Scholar]
- Elsherbiny, E.A.; Amin, B.H.; Aleem, B.; Kingsley, K.L.; Bennett, J.W. Trichoderma volatile organic compounds as a biofumigation tool against late blight pathogen Phytophthora infestans in postharvest potato tubers. J. Agric. Food Chem. 2020, 68, 8163–8171. [Google Scholar] [CrossRef] [PubMed]
- Rajani, P.; Rajasekaran, C.; Vasanthakumari, M.M.; Olsson, S.B.; Ravikanth, G.; Uma Shaanker, R. Inhibition of plant pathogenic fungi by endophytic Trichoderma spp. through mycoparasitism and volatile organic compounds. Microbiol. Res. 2021, 242, 126595. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, R.; Cordovez, V.; de Boer, W.; Raaijmakers, J.; Garbeva, P. Volatile affairs in microbial interactions. ISME J. 2015, 9, 2329–2335. [Google Scholar] [CrossRef] [PubMed]
- Jaibangyang, S.; Nasanit, R.; Limtong, S. Biological control of aflatoxin-producing Aspergillus flavus by volatile organic compound-producing antagonistic yeasts. BioControl 2020, 65, 377–386. [Google Scholar] [CrossRef]
- Ruiz-Moyano, S.; Hernández, A.; Galvan, A.I.; Córdoba, M.G.; Casquete, R.; Serradilla, M.J.; Martín, A. Selection and application of antifungal VOCs-producing yeasts as biocontrol agents of grey mould in fruits. Food Microbiol. 2020, 92, 103556. [Google Scholar] [CrossRef]
- Galvan, A.I.; Hernandez, A.; Cordoba, M.G.; Martin, A.; Serradilla, M.J.; Lopez-Corrales, M.; Rodriguez, A. Control of toxigenic Aspergillus spp. in dried figs by volatile organic compounds (VOCs) from antagonistic yeasts. Int. J. Food Microbiol. 2022, 376, 109772. [Google Scholar] [CrossRef]
- Delali, K.I.; Chen, O.; Wang, W.; Yi, L.; Deng, L.; Zeng, K. Evaluation of yeast isolates from kimchi with antagonistic activity against green mold in citrus and elucidating the action mechanisms of three yeast: P. kudriavzevii, K. marxianus, and Y. lipolytica. Postharvest Biol. Technol. 2021, 176, 111495. [Google Scholar] [CrossRef]
- Natarajan, S.; Balachandar, D.; Senthil, N.; Velazhahan, R.; Paranidharan, V. Volatiles of antagonistic soil yeasts inhibit growth and aflatoxin production of Aspergillus flavus. Microbiol. Res. 2022, 263, 127150. [Google Scholar] [CrossRef] [PubMed]
- Di Francesco, A.; Ugolini, L.; Lazzeri, L.; Mari, M. Production of volatile organic compounds by Aureobasidium pullulans as a potential mechanism of action against postharvest fruit pathogens. Biol. Control 2015, 81, 8–14. [Google Scholar] [CrossRef]
- Yalage Don, S.M.; Schmidtke, L.M.; Gambetta, J.M.; Steel, C.C. Volatile organic compounds produced by Aureobasidium pullulans induce electrolyte loss and oxidative stress in Botrytis cinerea and Alternaria alternata. Res. Microbiol. 2020, 172, 103788. [Google Scholar] [CrossRef] [PubMed]
- Contarino, R.; Brighina, S.; Fallico, B.; Cirvilleri, G.; Parafati, L.; Restuccia, C. Volatile organic compounds (VOCs) produced by biocontrol yeasts. Food Microbiol. 2019, 82, 70–74. [Google Scholar] [CrossRef] [PubMed]
- Ando, H.; Hatanaka, K.; Ohata, I.; Yamashita-Kitaguchi, Y.; Kurata, A.; Kishimoto, N. Antifungal activities of volatile substances generated by yeast isolated from Iranian commercial cheese. Food Control 2012, 26, 472–478. [Google Scholar] [CrossRef]
- Hua, S.S.; Beck, J.J.; Sarreal, S.B.; Gee, W. The major volatile compound 2-phenylethanol from the biocontrol yeast, Pichia anomala, inhibits growth and expression of aflatoxin biosynthetic genes of Aspergillus flavus. Mycotoxin Res. 2014, 30, 71–78. [Google Scholar] [CrossRef]
- Lu, J.; Li, J.; Li, L.; Qi, L.; Wang, Y.; Yang, S.; Xu, G.; Dou, D.; Wang, X.; Liu, J. Natural product 2-Phenylethanol inhibits ATP synthesis of P. infestans by blocking the oxidative phosphorylation pathway to prevent potato late blight. Postharvest Biol. Technol. 2023, 199, 112310. [Google Scholar] [CrossRef]
- Zou, X.; Wei, Y.; Jiang, S.; Xu, F.; Wang, H.; Zhan, P.; Shao, X. ROS stress and cell membrane disruption are the main antifungal mechanisms of 2-phenylethanol against Botrytis cinerea. J. Agric. Food Chem. 2022, 70, 14468–14479. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Volatile Compound | CAS | Molecular Formula | RT a (min) | RA b (%) | LRI(calculated) c | LRI(lit) d |
|---|---|---|---|---|---|---|
| Alcohols | ||||||
| 3-Methyl-1-butanol | 123-51-3 | C5H12O | 4.223 | 51.29 | 726 | 727 |
| 2-Methyl-1-butanol | 137-32-6 | C5H12O | 4.611 | 11.74 | 742 | 743 |
| 2-phenylethanol | 60-12-8 | C8H10O | 17.016 | 9.02 | 1110 | 1110 |
| 3-Cyclohexen-1-ol | 822-66-2 | C6H10O | 19.557 | 0.44 | 1179 | 1182 |
| Esters | ||||||
| Isoamyl acetate | 123-92-2 | C7H14O2 | 8.321 | 4.32 | 877 | 878 |
| Ethyl hexanoate | 123-66-0 | C8H16O2 | 12.796 | 1.07 | 999 | 999 |
| Octanoic acid, ethyl ester | 106-32-1 | C10H20O2 | 20.185 | 0.50 | 1196 | 1193 |
| Decanoic acid, ethyl ester | 110-38-3 | C12H24O2 | 26.941 | 0.88 | 1394 | 1391 |
| Heterocycles | ||||||
| Furan, 2-pentyl- | 3777-69-3 | C9H14O | 12.393 | 0.41 | 988 | 989 |
| Pyrazine, 3-ethyl-2,5-dimethyl- | 13360-65-1 | C8H12N2 | 15.629 | 0.63 | 1074 | 1077 |
| Pyrazine, 2, 5-dimethyl-3-(3-methylbutyl)- | 18433-98-2 | C11H18N2 | 24.177 | 0.29 | 1310 | 1310 |
| Alkanes | ||||||
| Pentane, 3-ethyl- | 617-78-7 | C7H16 | 3.598 | 0.55 | 696 | 687 |
| Dodecane | 112-40-3 | C12H26 | 20.334 | 0.37 | 1200 | 1200 |
| Acids | ||||||
| Formic acid | 64-18-6 | CH2O2 | 2.315 | 1.44 | -e | - |
| Amides | ||||||
| Formamide | 75-12-7 | CH3NO | 2.079 | 8.40 | - | - |
| Amines | ||||||
| Dimethylamine | 124-40-3 | C2H7N | 2.139 | 2.90 | - | 426 |
| Benzenes | ||||||
| o-Cymene | 527-84-4 | C10H14 | 13.722 | 0.49 | 1023 | 1027 |
| Ethers | ||||||
| Dimethyl ether | 115-10-6 | C2H6O | 1.755 | 5.26 | - | - |
| Volatile Compound | Inhibition Rate (%) | |||||
|---|---|---|---|---|---|---|
| Day 2 | Day 5 | |||||
| 50 μL/L | 100 μL/L | 150 μL/L | 50 μL/L | 100 μL/L | 150 μL/L | |
| 3-Methyl-1-butanol | 25.6 ± 12.3 c | 43.2 ± 3.7 c | 49.1 ± 9.4 cd | 4.3 ± 4.5 b | 14.4 ± 5.2 b | 21.4 ± 8.2 c |
| 2-Methyl-1-butanol | 29.2 ± 6.1 bc | 49.9 ± 3.2 bc | 56.7 ± 3.3 c | 5.7 ± 6.4 b | 19.7 ± 8.4 b | 27.4 ± 6.4 bc |
| 2-Phenylethanol | 76.6 ± 13.4 a | 87.4 ± 13.4 a | 100 ± 0.0 a | 22.5 ± 11.6 a | 83.4 ± 2.7 a | 92.9 ± 7.9 a |
| Isoamyl acetate | 40.3 ± 2.8 b | 53.9 ± 5.4 b | 90.7 ± 7.2 b | 17.2 ± 1.3 a | 22.1 ± 6.4 b | 25.5 ± 4.9 c |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zou, X.; Wei, Y.; Zhu, J.; Sun, J.; Shao, X. Volatile Organic Compounds of Scheffersomyces spartinae W9 Have Antifungal Effect against Botrytis cinerea on Strawberry Fruit. Foods 2023, 12, 3619. https://doi.org/10.3390/foods12193619
Zou X, Wei Y, Zhu J, Sun J, Shao X. Volatile Organic Compounds of Scheffersomyces spartinae W9 Have Antifungal Effect against Botrytis cinerea on Strawberry Fruit. Foods. 2023; 12(19):3619. https://doi.org/10.3390/foods12193619
Chicago/Turabian StyleZou, Xiurong, Yingying Wei, Jianhua Zhu, Jincai Sun, and Xingfeng Shao. 2023. "Volatile Organic Compounds of Scheffersomyces spartinae W9 Have Antifungal Effect against Botrytis cinerea on Strawberry Fruit" Foods 12, no. 19: 3619. https://doi.org/10.3390/foods12193619
APA StyleZou, X., Wei, Y., Zhu, J., Sun, J., & Shao, X. (2023). Volatile Organic Compounds of Scheffersomyces spartinae W9 Have Antifungal Effect against Botrytis cinerea on Strawberry Fruit. Foods, 12(19), 3619. https://doi.org/10.3390/foods12193619