

Cryoprotective Effect of NADES on Frozen-Thawed Mirror Carp Surimi in Terms of Oxidative Denaturation, Structural Properties, and Thermal Stability of Myofibrillar Proteins

Abstract
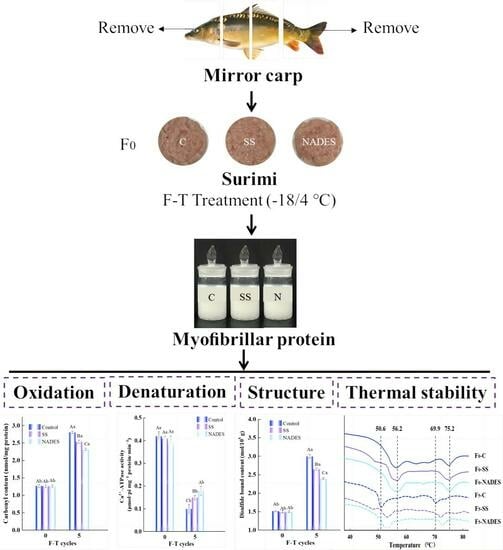
:
1. Introduction
2. Materials and Methods
2.1. Sample Preparation
2.1.1. NADES
2.1.2. F-T Surimi and MP Extraction
2.2. Determination of MP Oxidation and Denaturation
2.2.1. Schiff Base Fluorescence Intensity
2.2.2. Carbonyl Content
2.2.3. Ca2+-ATPase Activity
2.2.4. Surface Hydrophobicity
2.3. Determination of MP Structure Properties
2.3.1. Amino Acid Residue Side-Chain Groups
2.3.2. Secondary and Tertiary Structure
2.4. Thermal Stability
2.5. Statistical Analysis
3. Results and Discussion
3.1. The Inhibition of NADES on MP Oxidation and Denaturation of Frozen-Thawed Mirror Carp Surimi
3.1.1. The Inhibition of NADES on the Schiff base Fluorescence Intensity of Frozen-Thawed Mirror Carp Surimi
3.1.2. Inhibition of NADES on the Carbonyl Content of Frozen-Thawed Mirror Carp Surimi
3.1.3. Inhibition of NADES on Ca2+-ATPase Activity of Frozen-Thawed Mirror Carp Surimi
3.1.4. Inhibition of NADES on Surface Hydrophobicity of Frozen-Thawed Mirror Carp Surimi

3.2. Effect of NADES on MP Structure Properties of Frozen-Thawed Mirror Carp Surimi
3.2.1. Effect of NADES on MP Amino Acid Residue Side-Chain Groups of Frozen-Thawed Mirror Carp Surimi

3.2.2. Effect of NADES on the MP Secondary Structure of Frozen-Thawed Mirror Carp Surimi
3.2.3. Effect of NADES on the MP Tertiary Structure of Frozen-Thawed Mirror Carp Surimi

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| F-T Cycles | Samples | Secondary Structure Content (%) | Tertiary Structure | ||||
|---|---|---|---|---|---|---|---|
| α-Helix | β-Sheet | β-Turn | Random Coil | λmax (nm) | FI (A.U.) | ||
| 0 | Control | 60.5 ± 0.6 Aa | 17.0 ± 0.5 Ad | 9.30 ± 0.25 Aa | 13.2 ± 0.4 Ad | 332.4 ± 0.3 Ab | 1100 ± 5 Aa |
| SS | 60.8 ± 0.5 Aa | 17.2 ± 0.6 Ac | 9.25 ± 0.20 Aa | 12.8 ± 0.1 Ad | 332.3 ± 0.2 Aa | 1078 ± 12 Aa | |
| NADES | 60.6 ± 0.6 Aa | 17.1 ± 0.4 Ac | 9.35 ± 0.30 Aa | 13.0 ± 0.7 Ad | 332.2 ± 0.2 Ac | 1085 ± 20 Aa | |
| 1 | Control | 59.5 ± 0.5 Ab | 18.0 ± 0.3 Ac | 8.52 ± 0.17 Ab | 14.0 ± 0.4 Ac | 333.0 ± 0.2 Aa | 1050 ± 15 Ab |
| SS | 59.3 ± 0.3 Ab | 17.8 ± 0.4 Abc | 8.50 ± 0.35 Ab | 14.4 ± 0.3 Ac | 332.6 ± 0.3 Aa | 1065 ± 8 Aa | |
| NADES | 59.5 ± 0.3 Ab | 17.5 ± 0.3 Ac | 8.65 ± 0.35 Ab | 14.4 ± 0.3 Ac | 333.4 ± 0.1 Aa | 1060 ± 10 Aa | |
| 3 | Control | 56.3 ± 0.3 Bc | 19.5 ± 0.5 Ab | 7.45 ± 0.20 Ac | 16.8 ± 0.1 Ab | 331.6 ± 0.2 Ac | 903.0 ± 17.0 Bc |
| SS | 57.0 ± 0.5 ABc | 18.5 ± 0.5 Bb | 7.74 ± 0.24 Ac | 16.8 ± 0.2 Ab | 331.9 ± 0.2 Ab | 982.0 ± 12.1 Ab | |
| NADES | 57.5 ± 0.5 Ac | 18.3 ± 0.3 Bb | 7.80 ± 0.30 Ac | 16.4 ± 0.5 Ab | 332.9 ± 0.2 Bb | 1001 ± 9 Ab | |
| 5 | Control | 53.2 ± 0.5 Cd | 21.0 ± 0.4 Aa | 5.30 ± 0.15 Cd | 20.5 ± 0.3 Aa | 331.2 ± 0.2 Cc | 774.2 ± 16.3 Cd |
| SS | 54.5 ± 0.4 Bd | 20.0 ± 0.4 Ba | 6.15 ± 0.35 Bd | 19.4 ± 0.3 Ba | 331.7 ± 0.1 Bb | 880.4 ± 10.2 Bc | |
| NADES | 55.5 ± 0.3 Ad | 19.0 ± 0.4 Ca | 6.95 ± 0.20 Ad | 18.6 ± 0.2 Ca | 332.4 ± 0.1 Ac | 924.4 ± 4.6 Ac | |
3.2.4. Effect of NADES on the MP Thermal Stability of Frozen-Thawed Mirror Carp Surimi
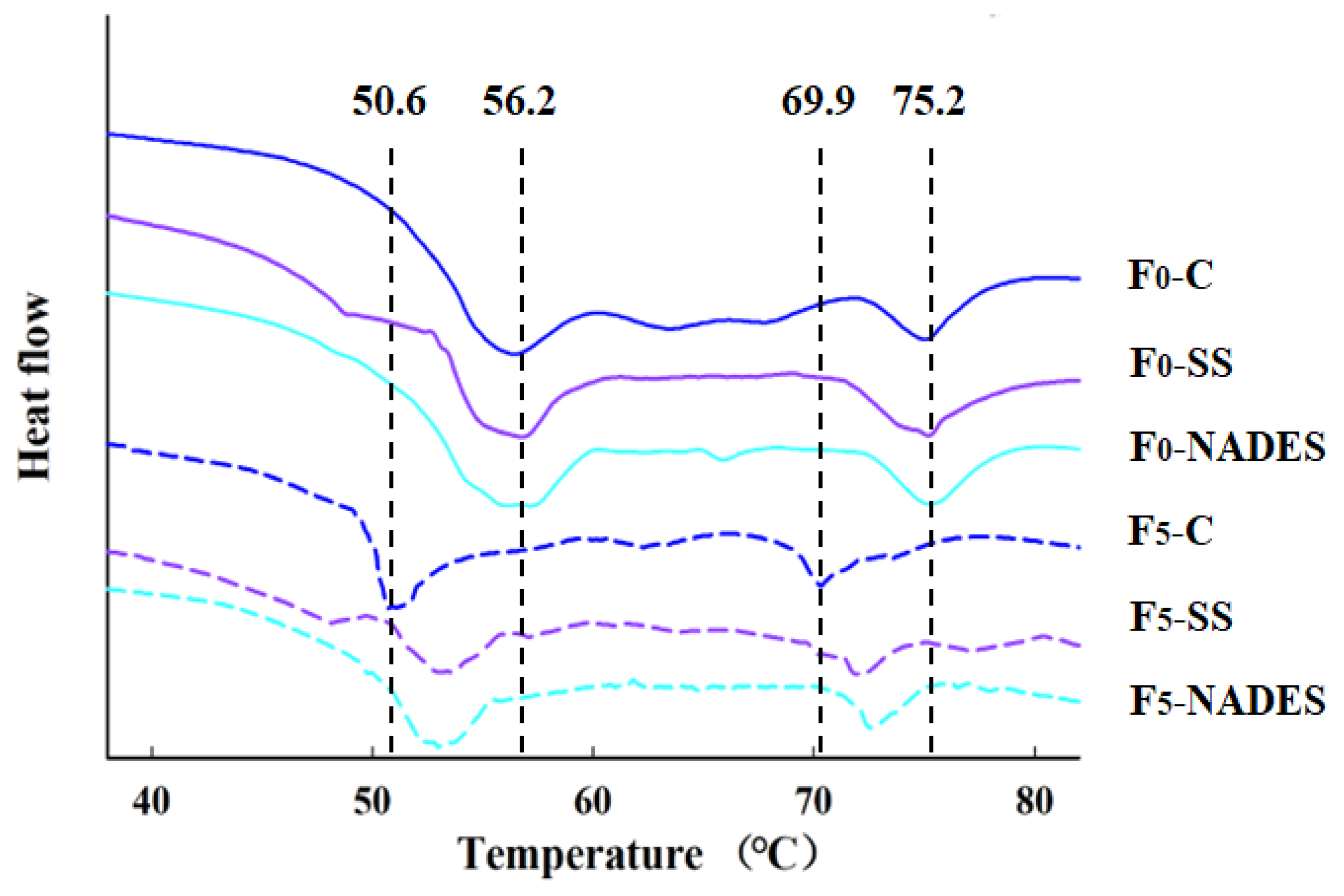
| F-T Cycles | Samples | Tmax1 (°C) | Tmax2 (°C) | ΔH1 (J/g) | ΔH2 (J/g) |
|---|---|---|---|---|---|
| 0 | Control | 56.2 ± 0.2 Aa | 75.2 ± 0.1 Aa | 1.154 ± 0.007 Aa | 0.362 ± 0.004 Aa |
| SS | 56.3 ± 0.2 Aa | 75.3 ± 0.1 Aa | 1.155 ± 0.010 Aa | 0.370 ± 0.005 Aa | |
| NADES | 56.4 ± 0.1 Aa | 75.2 ± 0.1 Aa | 1.162 ± 0.006 Aa | 0.367 ± 0.005 Aa | |
| 5 | Control | 50.6 ± 0.3 Bb | 69.9 ± 0.3 Cb | 0.680 ± 0.010 Bb | 0.227 ± 0.003 Cb |
| SS | 53.0 ± 0.3 Ab | 72.0 ± 0.2 Bb | 0.731 ± 0.005 Ab | 0.245 ± 0.003 Bb | |
| NADES | 53.1 ± 0.2 Ab | 72.9 ± 0.3 Ab | 0.738 ± 0.003 Ab | 0.255 ± 0.005 Ab |
4. Conclusions

Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zhou, A.; Benjakul, S.; Pan, K.; Gong, J.; Liu, X. Cryoprotective effects of trehalose and sodium lactate on tilapia (Sarotherodon nilotica) surimi during frozen storage. Food Chem. 2006, 96, 96–103. [Google Scholar] [CrossRef]
- Du, X.; Li, H.; Dong, C.; Ren, Y.; Pan, N.; Kong, B.; Liu, H.; Xia, X. Effect of ice structuring protein on the microstructure and myofibrillar protein structure of mirror carp (Cyprinus carpio L.) induced by freeze-thaw processes. LWT 2021, 139, 110570. [Google Scholar] [CrossRef]
- Zhang, J.; Fei, L.; Cui, P.; Walayat, N.; Ji, S.; Chen, Y.; Lyu, F.; Ding, Y. Effect of low voltage electrostatic field combined with partial freezing on the quality and microbial community of large yellow croaker. Food Res. Int. 2023, 169, 112933. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Li, S.; Yang, L.; Zhao, Z.; Xia, J.; Zhu, Y.; Li, C. Effect of multiple freeze–thaw cycles on water migration, protein conformation and quality attributes of beef longissimus dorsi muscle by real-time low field nuclear magnetic resonance and Raman spectroscopy. Food Res. Int. 2023, 166, 112644. [Google Scholar] [CrossRef]
- Bao, Y.; Ertbjerg, P.; Estévez, M.; Yuan, L.; Gao, R. Freezing of meat and aquatic food: Underlying mechanisms and implications on protein oxidation. Compr. Rev. Food Sci. Food Saf. 2021, 20, 5548–5569. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, G.; Xie, Q.; Wang, Y.; Yu, J.; Ma, X. A comprehensive review of the principles, key factors, application, and assessment of thawing technologies for muscle foods. Compr. Rev. Food Sci. Food Saf. 2023, 22, 107–134. [Google Scholar] [CrossRef]
- Bao, Y.; Wang, K.; Yang, H.; Regenstein, J.M.; Ertbjerg, P.; Zhou, P. Protein degradation of black carp (Mylopharyngodon piceus) muscle during cold storage. Food Chem. 2020, 308, 125576. [Google Scholar] [CrossRef]
- Du, X.; Wang, B.; Li, H.; Liu, H.; Shi, S.; Feng, J.; Pan, N.; Xia, X. Research progress on quality deterioration mechanism and control technology of frozen muscle foods. Compr. Rev. Food Sci. Food Saf. 2022, 21, 4812–4846. [Google Scholar] [CrossRef]
- Chen, X.; Li, X.; Yang, F.; Wu, J.; Huang, D.; Huang, J.; Wang, S. Effects and mechanism of antifreeze peptides from silver carp scales on the freeze-thaw stability of frozen surimi. Food Chem. 2022, 396, 133717. [Google Scholar] [CrossRef]
- Zhang, X.; Zhang, Y.; Dong, Y.; Ding, H.; Chen, K.; Lu, T.; Dai, Z. Study on the mechanism of protein hydrolysate delaying quality deterioration of frozen surimi. LWT 2022, 167, 113767. [Google Scholar] [CrossRef]
- EC. Commission Regulation (EU) No 1068/2013 of 30 October 2013 amending Annex II to Regulation (EC) No 1333/2008 of the European Parliament and of the Council as regards the use of diphosphates (E 450), triphosphates (E 451) and polyphosphates (E 452) in wet salted fish (L 289, 31/10/2013, pp. 58–60). 2013. Available online: https://eur-lex.europa.eu/eli/reg/2013/1068/oj/eng (accessed on 15 January 2023).
- Chen, J.; Fan, Y.; Zhang, X.; Yuan, Z.; Zhang, H.; Xu, X.; Qi, J.; Xiong, G.; Mei, L.; Zhu, Y.; et al. Effect of antifreeze protein on the quality and microstructure of frozen chicken breasts. Food Chem. 2023, 404, 134555. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Bai, G.; Jin, G.; Wang, Y.; Wang, J.; Puolanne, E.; Cao, J. Role of low molecular additives in the myofibrillar protein gelation: Underlying mechanisms and recent applications. Crit. Rev. Food Sci. 2022, xx, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Tian, Y.; Zhu, Z.; Sun, D.W. Naturally sourced biosubstances for regulating freezing points in food researches: Fundamentals, current applications and future trends. Trends Food Sci. 2020, 95, 131–140. [Google Scholar] [CrossRef]
- Liu, Y.; Friesen, J.B.; Mcalpine, J.B.; Lankin, D.C.; Chen, S.N.; Pauli, G.F. Natural deep eutectic solvents: Properties, applications, and perspectives. J. Nat. Prod. 2018, 81, 679–690. [Google Scholar] [CrossRef] [PubMed]
- Castro Vânia, I.B.; Craveiro, R.; Silva, J.M.; Reis, R.L.; Paiva, A.C.; Duarte, A.R. Natural deep eutectic systems as alternative nontoxic cryoprotective agents. Cryobiology 2018, 83, 15–26. [Google Scholar] [CrossRef]
- Jesus, A.R.; Meneses, L.; Duarte, A.R.C.; Paiva, A. Natural deep eutectic systems, an emerging class of cryoprotectant agents. Cryobiology 2021, 101, 95–104. [Google Scholar] [CrossRef]
- Tian, Y.; Sun, D.W.; Zhu, Z. Development of natural deep eutectic solvents (NADESs) as anti-freezing agents for the frozen food industry: Water-tailoring effects, anti-freezing mechanisms and applications. Food Chem. 2022, 371, 131150. [Google Scholar] [CrossRef]
- Du, X.; Kong, B.; He, J.; Zhang, Q.; An, G.; Zhang, T.; Xia, X. Cryoprotective effect of water-tailored trehalose-based natural deep eutectic solvents on frozen-thawed mirror carp (Cyprinus carpio L.) surimi. Food Chem. 2023, 426, 136633. [Google Scholar] [CrossRef]
- Antony, K.P.; Govindan, T.K. Packaging and storage studies of dried Lizard fish. Fish. Technol. 1983, 20, 34–41. Available online: http://hdl.handle.net/1834/33793 (accessed on 15 January 2023).
- Choi, Y.H.; Spronsen, J.V.; Dai, Y.; Verberne, M.; Hollmann, F.; Arends, I.W.C.E.; Witkamp, G.-J.; Verporte, R. Are natural deep eutectic solvents the missing link in understanding cellular metabolism and physiology? Plant Physiol. 2011, 156, 1701–1705. [Google Scholar] [CrossRef]
- Ferreira, I.J.; Meneses, L.; Paiva, A.; Diniz, M.; Duarte, A.R.C. Assessment of deep eutectic solvents toxicity in zebrafish (Danio rerio). Chemosphere 2022, 299, 134415. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Wang, B.; Kong, B.; Shi, S.; Xia, X. Decreased gelling properties of protein in mirror carp (Cyprinus carpio) are due to protein aggregation and structure deterioration when subjected to freeze-thaw cycles. Food Hydrocoll. 2019, 97, 105223. [Google Scholar] [CrossRef]
- Li, T.; Niu, L.; Li, X.; Wang, F.; Huang, Y.; Liu, Y. Formation of advanced glycation end-products in silver carp (Hypophthalmichthys molitrix) surimi products during heat treatment as affected by freezing-thawing cycles. Food Chem. 2022, 395, 33612. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Sun, X.; Mubango, E.; Zheng, Y.; Liu, Y.; Tan, Y.; Luo, Y.; Hong, H. Effects of protein and lipid oxidation on the water holding capacity of different parts of bighead carp: Eye, dorsal, belly and tail muscles. Food Chem. 2023, 423, 136238. [Google Scholar] [CrossRef]
- Du, X.; Chang, P.; Tian, J.; Kong, B.; Sun, F.; Xia, X. Effect of ice structuring protein on the quality, thermal stability and oxidation of mirror carp (Cyprinus carpio L.) induced by freeze-thaw cycles. LWT 2020, 124, 109140. [Google Scholar] [CrossRef]
- Chu, F.; Lin, Y.; Huang, Y.; Niu, L.; Lai, K. Effect of sucrose on the formation of advanced glycation end-products of ground pork during freeze–thaw cycles and subsequent heat treatment. Foods 2023, 12, 1024. [Google Scholar] [CrossRef]
- Utrera, M.; Estévez, M. Oxidative damage to poultry, pork, and beef during frozen storage through the analysis of novel protein oxidation markers. J. Agric. Food Chem. 2013, 61, 7987–7993. [Google Scholar] [CrossRef]
- Zhang, W.; Xiao, S.; Ahn, D.U. Protein oxidation: Basic principles and implications for meat quality. Crit. Rev. Food Sci. Nutr. 2013, 53, 1191–1201. [Google Scholar] [CrossRef]
- Li, H.; Chang, L.; Pan, N.; Du, X.; Shi, S.; Zhang, Q.; An, G.; Xia, X.; Zhang, L. Dynamic changes in postmortem quality of mirror carp (Cyprinus carpio L.): Based on oxidation reaction and mitochondrial function properties. Food Chem. 2023, 425, 136426. [Google Scholar] [CrossRef]
- Nikoo, M.; Benjakul, S. Potential application of seafood-derived peptides as bifunctional ingredients, antioxidant-cryoprotectant: A review. J. Funct. Foods 2015, 19, 753–764. [Google Scholar] [CrossRef]
- Yu, Q.; Liu, J.; Liu, Y.; Zheng, Y.; Pi, R.; Mubango, E.; Tan, Y.; Luo, Y.; Hong, H. Inhibitive effect of cryoprotectants on the oxidative and structural changes in myofibrillar proteins of unwashed mince from silver carp during frozen storage. Food Res. Int. 2022, 161, 111880. [Google Scholar] [CrossRef]
- Wang, M.; Li, Y.; Ma, Z.; Guo, L.; Huang, M.; Sun, J. Stability of native/thermally denatured myofibrillar protein particles: Improvement with decreasing pH. Food Hydrocoll. 2023, 140, 108628. [Google Scholar] [CrossRef]
- Yu, M.M.; Fan, Y.C.; Xu, S.J.; Liu, Y.X.; Wu, Z.X.; Zhou, D.Y.; Zhu, B.W. Effects of antioxidants on the texture and protein quality of ready-to-eat abalone muscles during storage. J. Food Compos. Anal. 2022, 108, 104456. [Google Scholar] [CrossRef]
- Benjakul, S.; Bauer, F. Physicochemical and enzymatic changes of cod muscle proteins subjected to different freeze-thaw cycles. J. Sci. Food Agric. 2000, 80, 1143–1150. [Google Scholar] [CrossRef]
- Wu, K.; Ren, J.; Wang, Q.; Nuerjiang, M.; Xia, X.; Bian, C. Research Progress on the Preparation and Action Mechanism of Natural Deep Eutectic Solvents and Their Application in Food. Foods 2022, 11, 3528. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Xiong, Y.L. Chlorogenic acid-mediated gel formation of oxidatively stressed myofibrillar protein. Food Chem. 2015, 180, 235–243. [Google Scholar] [CrossRef]
- Lin, J.; Hong, H.; Zhang, L.; Zhang, C.; Luo, Y. Antioxidant and cryoprotective effects of hydrolysate from gill protein of bighead carp (Hypophthalmichthys nobilis) in preventing denaturation of frozen surimi. Food Chem. 2019, 298, 124868. [Google Scholar] [CrossRef]
- Benjakul, S.; Visessanguan, W.; Tueksuban, J. Changes in physico-chemical properties and gel-forming ability of lizardfish (Saurida tumbil) during post-mortem storage in ice. Food Chem. 2003, 80, 535–544. [Google Scholar] [CrossRef]
- Sava, N.; Van der Plancken, I.; Claeys, W.; Hendrickx, M. The kinetics of heat-induced structural changes of β-lactoglobulin. J. Dairy Sci. 2005, 88, 1646–1653. [Google Scholar] [CrossRef]
- Xiong, X.; Liu, X.; Zhu, X.; Tan, Y.; Wang, Z.; Xu, J.; Tu, X.; Rao, Y.; Duan, J.; Zhao, W.; et al. A mutation in PHKG1 causes high drip loss and low meat quality in Chinese Ningdu yellow chickens. Poult. Sci. 2022, 101, 101556. [Google Scholar] [CrossRef]
- Levine, R.L.; Garland, D.; Oliver, C.N.; Amici, A.; Climent, I.; Lenz, A.G.; Ahn, B.W.; Shaltiel, S.; Stadtman, E.R. Determination of carbonyl content in oxidatively modified proteins. Methods Enzymol. 1990, 186, 464–478. [Google Scholar] [CrossRef]
- George, S. Influence of cryoprotectant levels on storage stability of surimi from Nemipterus japonicus and quality of surimi-based products. J. Food Sci. Technol. 2014, 51, 982–987. [Google Scholar] [CrossRef]
- Tian, Y.; Sun, D.W.; Xu, L.; Fan, T.H.; Zhang, S.T.; Zhu, Z. Bioinspired cryoprotectants enabled by binary natural deep eutectic solvents for sustainable and green cryopreservation. ACS Sustain. Chem. Eng. 2022, 10, 7677–7691. [Google Scholar] [CrossRef]
- Zhang, Z.; Yang, Y.; Zhou, P.; Zhang, X.; Wang, J. Effects of high pressure modification on conformation and gelation properties of myofibrillar protein. Food Chem. 2017, 217, 678–686. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Sun, D.W.; Pu, H.; Wei, Q. Principles and applications of spectroscopic techniques for evaluating food protein conformational changes: A review. Trends Food Sci. Technol. 2017, 67, 207–219. [Google Scholar] [CrossRef]
- Tan, M.; Ding, Z.; Chu, Y.; Xie, J. Potential of Good’s buffers to inhibit denaturation of myofibrillar protein upon freezing. Food Res. Int. 2023, 165, 112484. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Lin, Z.; Chen, X.; Chen, J.; Wu, J.; Chen, H.; Zeng, X. Characterization of structures and gel properties of ultra-high-pressure treated-myofibrillar protein extracted from mud carp (Cirrhinus molitorella) and quality characteristics of heat-induced sausage products. LWT 2022, 165, 113691. [Google Scholar] [CrossRef]
- Tan, M.; Ye, J.; Chu, Y.; Xie, J. The effects of ice crystal on water properties and protein stability of large yellow croaker (Pseudosciaena crocea). Int. J. Refrig. 2021, 130, 242–252. [Google Scholar] [CrossRef]
- Saeed, S.; Howell, N.K. Rheological and differential scanning calorimetry studies on structural and textural changes in frozen Atlantic mackerel (Scomber scombrus). J. Sci. Food Agric. 2004, 84, 1216–1222. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, H.; Wang, Q.; Li, W.; Xia, X. Cryoprotective Effect of NADES on Frozen-Thawed Mirror Carp Surimi in Terms of Oxidative Denaturation, Structural Properties, and Thermal Stability of Myofibrillar Proteins. Foods 2023, 12, 3530. https://doi.org/10.3390/foods12193530
Li H, Wang Q, Li W, Xia X. Cryoprotective Effect of NADES on Frozen-Thawed Mirror Carp Surimi in Terms of Oxidative Denaturation, Structural Properties, and Thermal Stability of Myofibrillar Proteins. Foods. 2023; 12(19):3530. https://doi.org/10.3390/foods12193530
Chicago/Turabian StyleLi, Haijing, Qian Wang, Wenxin Li, and Xiufang Xia. 2023. "Cryoprotective Effect of NADES on Frozen-Thawed Mirror Carp Surimi in Terms of Oxidative Denaturation, Structural Properties, and Thermal Stability of Myofibrillar Proteins" Foods 12, no. 19: 3530. https://doi.org/10.3390/foods12193530
APA StyleLi, H., Wang, Q., Li, W., & Xia, X. (2023). Cryoprotective Effect of NADES on Frozen-Thawed Mirror Carp Surimi in Terms of Oxidative Denaturation, Structural Properties, and Thermal Stability of Myofibrillar Proteins. Foods, 12(19), 3530. https://doi.org/10.3390/foods12193530