
Dynamic Change of Carbon and Nitrogen Sources in Colonized Apples by Penicillium expansum






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Fruit
2.2. Preparation of the Spore Suspension
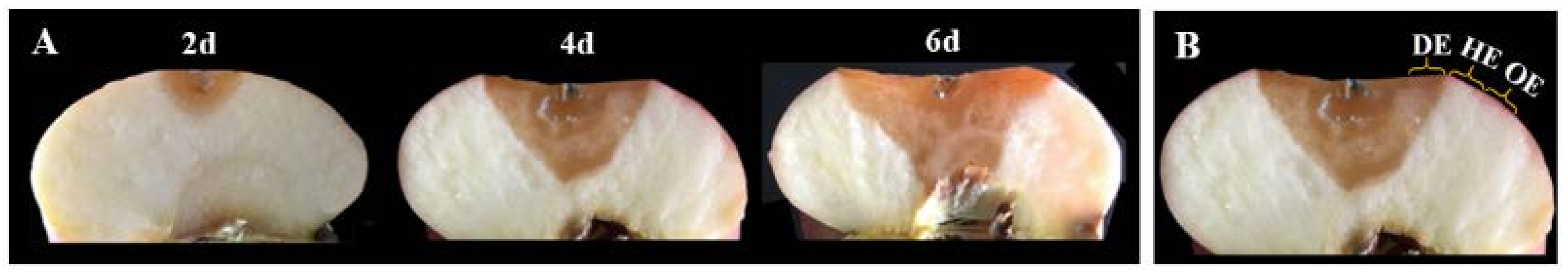
2.3. Fruit Inoculation
2.4. Sampling
2.5. Measurement of the Total Soluble Solids (TSS) and Titratable Acidity (TA) Contents
2.6. Analysis of the Sugar and Organic Acid Contents
2.7. Analysis of the Amino Acid Contents
2.8. Analysis of the Fatty Acid (FA) Contents
2.9. Statistical Analysis
3. Results and Discussion
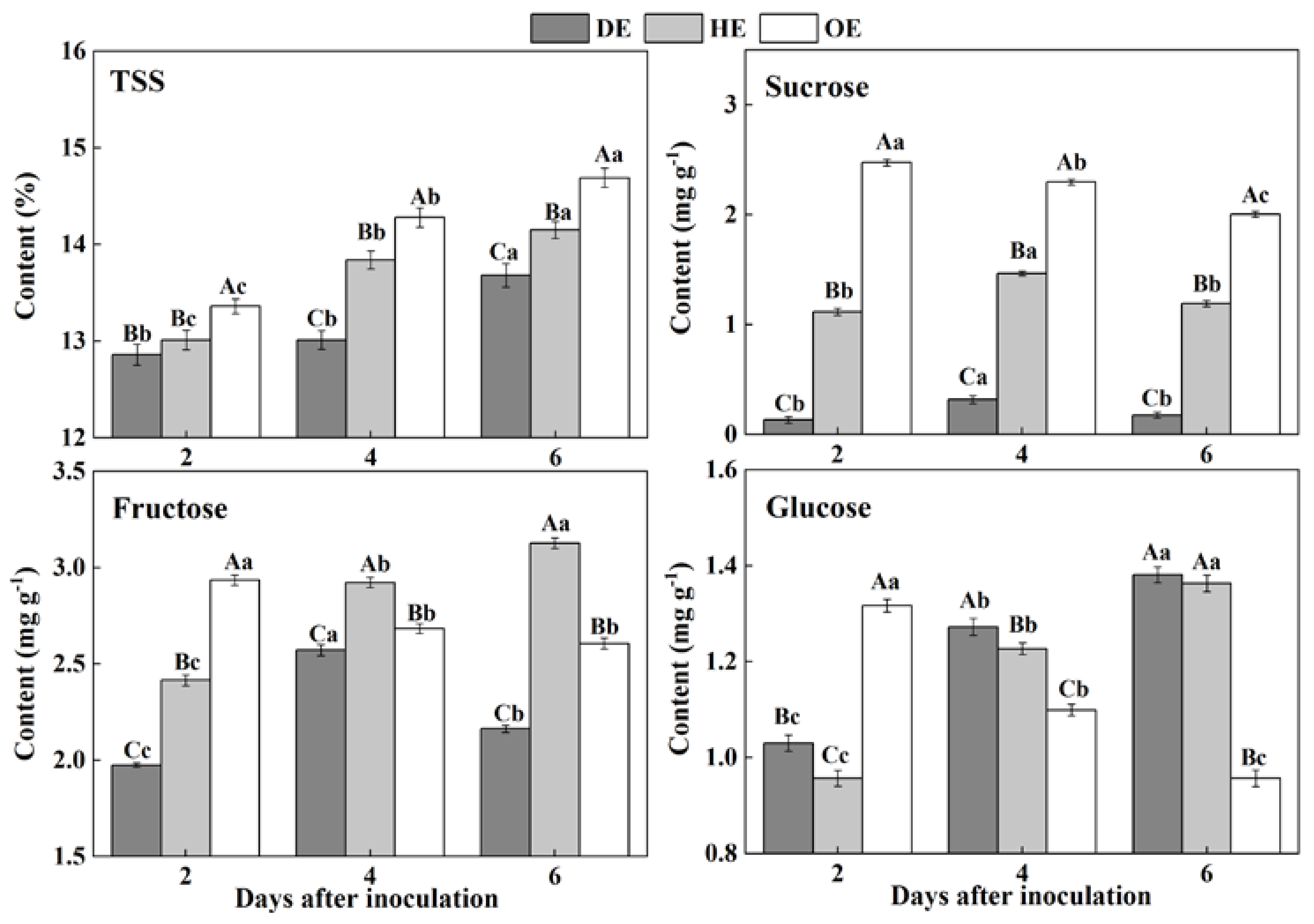
3.1. Effects of P. expansum Colonization on the Contents of TSS and Individual Sugars in Different Regions of the Apple Fruit
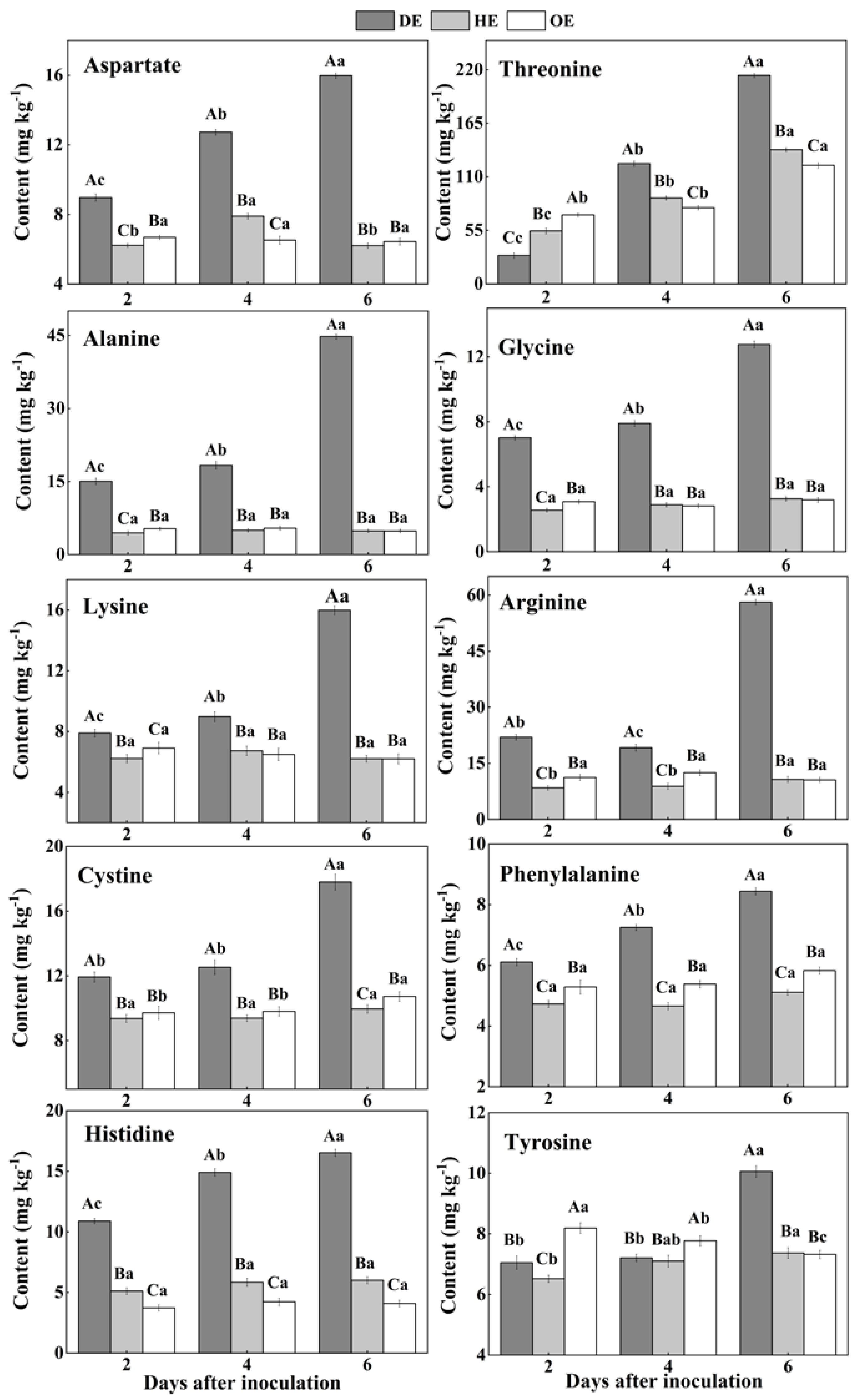
3.2. Effects of P. expansum Colonization on the Contents of Amino Acids in Different Regions of Apple Fruit
3.3. Effects of P. expansum Colonization on the Contents of TA and Individual Organic Acids in Different Regions of the Apple Fruit
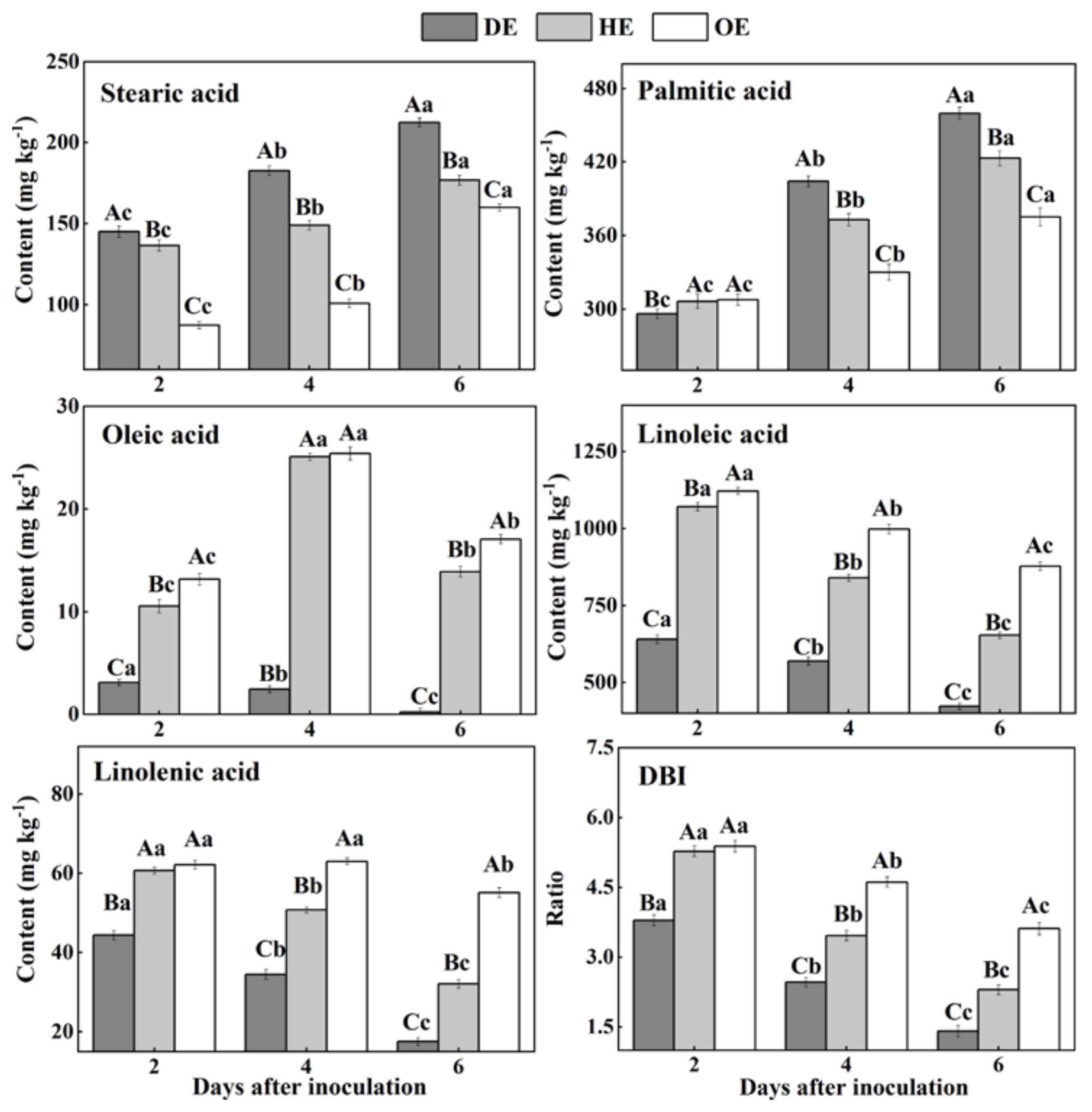
3.4. Effects of P. expansum Colonization on the Contents of Fatty Acids in the Different Regions of Apple Fruit
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Luciano-Rosario, D.; Keller, N.P.; Jurick, W.M. Penicillium expansum: Biology, omics, and management tools for a global postharvest pathogen causing blue mould of pome fruit. Mol. Plant Pathol. 2020, 21, 1391–1404. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.L.; Qiao, N.Z.; Zhao, J.X.; Zhang, H.; Tian, F.W.; Zhai, Q.X.; Chen, W. Postharvest control of Penicillium expansum in fruits: A review. Food Biosci. 2020, 36, 100633. [Google Scholar] [CrossRef]
- Prusky, D.; Alkan, N.; Mengiste, T.; Fluhr, R. Quiescent and necrotrophic lifestyle choice during postharvest disease development. Annu. Rev. Phytopathol. 2013, 51, 155–176. [Google Scholar] [CrossRef] [PubMed]
- Xylia, P.; Chrysargyris, A.; Ahmed, Z.F.R.; Tzortzakis, N. Application of rosemary and eucalyptus essential oils and their main component on the preservation of apple and pear fruits. Horticulturae 2021, 7, 479. [Google Scholar] [CrossRef]
- Xylia, P.; Chrysargyris, A.; Shahwar, D.; Ahmed, Z.F.R.; Tzortzakis, N. Application of rosemary and eucalyptus essential oils on the preservation of cucumber fruit. Horticulturae 2022, 8, 774. [Google Scholar] [CrossRef]
- García-Guzmán, G.; Heil, M. Life histories of hosts and pathogens predict patterns in tropical fungal plant diseases. New Phytol. 2014, 201, 1106–1120. [Google Scholar] [CrossRef]
- Ahmed, Z.F.; Kaur, N.; Maqsood, S.; Schmeda-Hirschmann, G. Preharvest applications of chitosan, salicylic acid, and calcium chloride have a synergistic effect on quality and storability of date palm fruit (Phoenix dactylifera L.). HortScience 2022, 57, 422–430. [Google Scholar] [CrossRef]
- Schmidt, S.M.; Panstruga, R. Pathogenomics of fungal plant parasites: What have we learnt about pathogenesis? Curr. Opin. Plant Biol. 2011, 14, 392–399. [Google Scholar] [CrossRef]
- Shen, Y.M.; Liu, M.Y.; Nie, J.Y.; Ma, N.; Xu, G.F.; Zhang, J.Y.; Li, Y.P.; Li, H.F.; Kuang, L.X.; Li, Z.Y. Metabolite changes of apple Penicillium expansum infection based on a UPLC-Q-TOF metabonomics approach. Postharvest Biol. Technol. 2021, 181, 111646. [Google Scholar] [CrossRef]
- Kou, J.; Wei, Y.Y.; He, X.X.; Xu, J.Y.; Xu, F.; Shao, X.F. Infection of post-harvest peaches by Monilinia fructicola accelerates sucrose decomposition and stimulates the Embden-Meyerhof-Parnas pathway. Hortic. Res. 2018, 5, 46. [Google Scholar] [CrossRef]
- Tang, N.; Chen, N.; Hu, N.; Deng, W.; Chen, Z.X.; Li, Z.G. Comparative metabolomics and transcriptomic profiling reveal the mechanism of fruit quality deterioration and the resistance of citrus fruit against Penicillium digitatum. Postharvest Biol. Technol. 2018, 145, 61–73. [Google Scholar] [CrossRef]
- Gong, D.; Bi, Y.; Zong, Y.Y.; Li, Y.C.; Sionov, E.; Prusky, D. Penicillium expansum-induced release of branched-chain volatile compounds in apple fruit by increasing amino acids accumulation. Postharvest Biol. Technol. 2021, 173, 111432. [Google Scholar] [CrossRef]
- Prusky, D.; McEvoy, J.L.; Saftner, R.; Conway, W.S.; Jones, R. Relationship between host acidification and virulence of Penicillium spp. on apple and citrus fruit. Phytopathology 2004, 94, 44–51. [Google Scholar] [CrossRef] [PubMed]
- Vilanova, L.; Viñas, I.; Torres, R.; Usall, J.; Buron-Moles, G.; Teixidó, N. Acidification of apple and orange hosts by Penicillium digitatum and Penicillium expansum. Int. J. Food Microbiol. 2014, 178, 39–49. [Google Scholar] [CrossRef]
- Gong, D.; Bi, Y.; Li, Y.C.; Zong, Y.Y.; Han, Y.; Prusky, D. Both Penicillium expansum and Trichothecium roseum infections promote the ripening of apples and release specific volatile compounds. Front. Plant Sci. 2019, 10, 338. [Google Scholar] [CrossRef]
- Lin, Y.F.; Chen, M.Y.; Lin, H.T.; Lin, M.S.; Hung, Y.C.; Lin, Y.X.; Chen, Y.H.; Wang, H.; Ritenourd, M.A. Phomopsis longanae-induced pericarp browning and disease development of longan fruit can be alleviated or aggravated by regulation of ATP-mediated membrane lipid metabolism. Food Chem. 2018, 269, 644–651. [Google Scholar] [CrossRef]
- Walley, J.W.; Kliebenstein, D.J.; Bostock, R.M.; Dehesh, K. Fatty acids and early detection of pathogens. Curr. Opin. Plant Biol. 2013, 16, 520–526. [Google Scholar] [CrossRef]
- Wang, H.; Chen, Y.H.; Sun, J.Z.; Lin, Y.F.; Lin, Y.X.; Lin, M.S.; Hung, Y.C.; Lin, H.T. The changes in metabolisms of membrane lipids and phenolics induced by Phomopsis longanae Chi infection in association with pericarp browning and disease occurrence of postharvest longan fruit. J. Agric. Food Chem. 2018, 66, 12794–12804. [Google Scholar] [CrossRef]
- He, Y.Z.; Li, Z.R.; Tan, F.Q.; Liu, H.; Zhu, M.; Yang, H.B.; Bi, G.L.; Wan, H.L.; Wang, J.Q.; Xu, R.W.; et al. Fatty acid metabolic flux and lipid peroxidation homeostasis maintain the biomembrane stability to improve citrus fruit storage performance. Food Chem. 2019, 292, 314–324. [Google Scholar] [CrossRef]
- Lin, Y.X.; Lin, H.T.; Chen, Y.H.; Wang, H.; Ritenour, M.A.; Lin, Y.F. Hydrogen peroxide-induced changes in activities of membrane lipids-degrading enzymes and contents of membrane lipids composition in relation to pulp breakdown of longan fruit during storage. Food Chem. 2019, 297, 124955. [Google Scholar] [CrossRef]
- Chen, Y.H.; Sun, J.Z.; Lin, H.T.; Lin, M.S.; Lin, Y.F.; Wang, H.; Hung, Y.C. Salicylic acid treatment suppresses Phomopsis longanae Chi-induced disease development of postharvest longan fruit by modulating membrane lipid metabolism. Postharvest Biol. Technol. 2020, 164, 111168. [Google Scholar] [CrossRef]
- Lee, J.; Jeong, M.C.; Ku, K.H. Chemical, physical, and sensory properties of 1-MCP-treated Fuji apple (Malus domestica Borkh.) fruits after long-term cold storage. Appl. Biol. Chem. 2017, 60, 363–374. [Google Scholar] [CrossRef]
- Zhang, W.S.; Chen, K.S.; Zhang, B.; Sun, C.D.; Cai, C.; Zhou, C.H.; Ferguson, I.B. Postharvest responses of Chinese bayberry fruit. Postharvest Biol. Technol. 2005, 37, 241–251. [Google Scholar] [CrossRef]
- Gancedo, M.C.; Luh, B. HPLC analysis of organic acids and sugars in tomato juice. J. Food Sci. 1986, 51, 571–573. [Google Scholar] [CrossRef]
- López-Hernández, J.; Oruña-Concha, M.; Simal-Lozano, J.; Vázquez-Blanco, M.; González-Castro, M. Chemical composition of Padrón peppers (Capsicum annuum L.) grown in Galicia (NW Spain). Food Chem. 1996, 57, 557–559. [Google Scholar] [CrossRef]
- Li, Y.; Qi, H.; Jin, Y.; Tian, X.; Sui, L.; Qiu, Y. Role of ethylene in biosynthetic pathway of related-aroma volatiles derived from amino acids in oriental sweet melons (Cucumis melo var. makuwa Makino). Sci. Hortic. 2016, 201, 24–35. [Google Scholar] [CrossRef]
- Valero-Garrido, D.; López-Frías, M.; Llopis, J.; López-Jurado, M. Influence of dietary fat on the lipid composition of perirenal adipose tissue in rats. Ann. Nutr. Metab. 1990, 34, 327–332. [Google Scholar] [CrossRef]
- Kim, J.Y.; Loo EP, I.; Pang, T.Y.; Lercher, M.; Frommer, W.B.; Wudick, M.M. Cellular export of sugars and amino acids: Role in feeding other cells and organisms. Plant Physiol. 2021, 187, 1893–1914. [Google Scholar] [CrossRef]
- Divon, H.H.; Fluhr, R. Nutrition acquisition strategies during fungal infection of plants. FEMS Microbiol. Lett. 2007, 266, 65–74. [Google Scholar] [CrossRef]
- Petrasch, S.; Silva, C.J.; Mesquida-Pesci, S.D.; Gallegos, K.; van den Abeele, C.; Papin, V.; Fernandez-Acero, F.J.; Knapp, S.J.; Blanco-Ulate, B. Infection strategies deployed by Botrytis cinerea, Fusarium acuminatum, and Rhizopus stolonifer as a function of tomato fruit ripening stage. Front. Plant Sci. 2019, 10, 223. [Google Scholar] [CrossRef]
- Juge, N. Plant protein inhibitors of cell wall degrading enzymes. Trends Plant Sci. 2006, 11, 359–367. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Sun, C.; Guan, X.; Lian, S.; Li, B.; Wang, C. Butylated hydroxytoluene induced resistance against Botryosphaeriadothidea in apple fruit. Front. Microbiol. 2021, 11, 599062. [Google Scholar] [CrossRef] [PubMed]
- Bezrutczyk, M.; Yang, J.; Eom, J. Sugar flux and signaling in plant-microbe interactions. Plant J. 2018, 93, 675–685. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.J.; Shi, Z.D.; Jiang, Y.P.; Zhang, X.H.; Li, X.; Li, F.J. Effects of preharvest regulation of ethylene on carbohydrate metabolism of apple (Malus domestica Borkh cv. Starkrimson) fruit at harvest and during storage. Sci. Hortic. 2021, 276, 109748. [Google Scholar] [CrossRef]
- Moghaddam, M.R.B.; Van den Ende, W. Sugars and plant innate immunity. J. Exp. Bot. 2012, 63, 3989–3998. [Google Scholar] [CrossRef]
- Luna, E.; Flandin, A.; Cassan, C.; Prigent, S.; Chevanne, C.; Kadiri, C.F.; Gibon, Y.; Pétriacq, P. Metabolomics to exploit the primed immune system of tomato fruit. Metabolites 2020, 10, 96. [Google Scholar] [CrossRef]
- Yao, S.X.; Cao, Q.; Xie, J.; Deng, L.L.; Zeng, K.F. Alteration of sugar and organic acid metabolism in postharvest granulation of Ponkan fruit revealed by transcriptome profiling. Postharvest Biol. Technol. 2018, 139, 2–11. [Google Scholar] [CrossRef]
- Jin, L.F.; Cai, Y.T.; Sun, C.; Huang, Y.N.; Yu, T. Exogenous L-glutamate treatment could induce resistance against Penicillium expansum in pear fruit by activating defense-related proteins and amino acids metabolism. Postharvest Biol. Technol. 2019, 150, 148–157. [Google Scholar] [CrossRef]
- Yang, G.; Wei, Q.; Huang, H.; Xia, J. Amino acid transporters in plant cells: A brief review. Plants 2020, 9, 967. [Google Scholar] [CrossRef]
- Xue, Q. Pathogen proteases and host protease inhibitors in molluscan infectious diseases. J. Invertebr. Pathol. 2019, 166, 107214. [Google Scholar] [CrossRef]
- Huang, H.; Nguyen Thi Thu, T.; He, X.; Gravot, A.; Bernillon, S.; Ballini, E.; Morel, J.B. Increase of fungal pathogenicity and role of plant glutamine in nitrogen-induced susceptibility (NIS) to rice blast. Front. Plant Sci. 2017, 8, 265. [Google Scholar] [CrossRef] [PubMed]
- Dinkeloo, K.; Boyd, S.; Pilot, G. Update on amino acid transporter functions and on possible amino acid sensing mechanisms in plants. Semin. Cell Dev. Biol. 2018, 74, 105–113. [Google Scholar] [CrossRef] [PubMed]
- Mäkinen, K.M.; Swarnalok, D. The significance of methionine cycle enzymes in plant virus infections. Curr. Opin. Plant Biol. 2019, 50, 67–75. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Xu, M.; Wang, G.; Galili, G. New insights into the metabolism of aspartate-family amino acids in plant seeds. Plant Reprod. 2018, 31, 203–211. [Google Scholar] [CrossRef] [PubMed]
- Benstein, R.M.; Ludewig, K.; Wulfert, S.; Wittek, S.; Gigolashvili, T.; Frerigmann, H.; Krueger, S. Arabidopsis phosphoglycerate dehydrogenase 1 of the phosphoserine pathway is essential for development and required for ammonium assimilation and tryptophan biosynthesis. Plant Cell 2013, 25, 5011–5029. [Google Scholar] [CrossRef] [PubMed]
- Binder, S. Branched-chain amino acid metabolism in Arabidopsis thaliana. Arab. Book 2010, 8, e0137. [Google Scholar] [CrossRef] [PubMed]
- Famiani, F.; Battistelli, A.; Moscatello, S.; Cruz-Castillo, J.G.; Walker, R.P. The organic acids that are accumulated in the flesh of fruits: Occurrence, metabolism and factors affecting their contents-a review. Rev. Chapingo Ser. Hortic. 2015, 21, 97–128. [Google Scholar] [CrossRef]
- Etienne, A.; Génard, M.; Lobit, P.; Mbéguié-A-Mbéguié, D.; Bugaud, C. What controls fleshy fruit acidity? A review of malate and citrate accumulation in fruit cells. J. Exp. Bot. 2013, 64, 1451–1469. [Google Scholar] [CrossRef]
- Prusky, D.; Bi, F.; Moral, J.; Barad, S. How does host carbon concentration modulate the lifestyle of postharvest pathogens during infection? Front. Plant Sci. 2016, 7, 1306. [Google Scholar] [CrossRef]
- Chakraborty, N.; Ghosh, R.; Ghosh, S.; Narula, K.; Tayal, R.; Datta, A.; Chakraborty, S. Reduction of oxalate levels in tomato fruit and consequent metabolic remodeling following overexpression of a fungal oxalate decarboxylase. Plant Physiol. 2013, 162, 364–378. [Google Scholar] [CrossRef]
- Gong, D.; Bi, Y.; Zhang, X.M.; Han, Z.H.; Zong, Y.Y.; Li, Y.C.; Sionov, E.; Prusky, D. Benzothiadiazole treatment inhibits membrane lipid metabolism and straight-chain volatile compound release in Penicillium expansum-inoculated apple fruit. Postharvest Biol. Technol. 2021, 181, 111671. [Google Scholar] [CrossRef]
- Peng, H.; Gong, D.; Wei, Y.N.; Yang, Q.; Zong, Y.Y.; Prusky, D.; Sionov, E.; Bi, Y. Changes of membrane phospholipid metabolism in apple fruit infected by Penicillium expansum. Food Sci. 2020. Available online: http://kns.cnki.net/kcms/detail/11.2206.TS.20200330.1406.048.html (accessed on 30 January 2020).
- Hasanuzzaman, M.; Bhuyan, M.H.M.B.; Zulfiqar, F.; Raza, A.; Mohsin, S.M.; Mahmud, J.A.; Fujita, M.; Fotopoulos, V. Reactive oxygen species and antioxidant defense in plants under abiotic stress: Revisiting the crucial role of a universal defense regulator. Antioxidants 2020, 9, 681. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.Q.; Greer, M.S.; Weselake, R.J. Plant phospholipase A: Advances in molecular biology, biochemistry, and cellular function. BioMolecular Concepts 2013, 4, 527–532. [Google Scholar] [CrossRef] [PubMed]
- Lim, G.H.; Singhal, R.; Kachroo, A.; Kachroo, P. Fatty acid-and lipid-mediated signaling in plant defense. Annu. Rev. Phytopathol. 2017, 55, 505–536. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gong, D.; Bi, Y.; Zong, Y.; Li, Y.; Sionov, E.; Prusky, D. Dynamic Change of Carbon and Nitrogen Sources in Colonized Apples by Penicillium expansum. Foods 2022, 11, 3367. https://doi.org/10.3390/foods11213367
Gong D, Bi Y, Zong Y, Li Y, Sionov E, Prusky D. Dynamic Change of Carbon and Nitrogen Sources in Colonized Apples by Penicillium expansum. Foods. 2022; 11(21):3367. https://doi.org/10.3390/foods11213367
Chicago/Turabian StyleGong, Di, Yang Bi, Yuanyuan Zong, Yongcai Li, Edward Sionov, and Dov Prusky. 2022. "Dynamic Change of Carbon and Nitrogen Sources in Colonized Apples by Penicillium expansum" Foods 11, no. 21: 3367. https://doi.org/10.3390/foods11213367
APA StyleGong, D., Bi, Y., Zong, Y., Li, Y., Sionov, E., & Prusky, D. (2022). Dynamic Change of Carbon and Nitrogen Sources in Colonized Apples by Penicillium expansum. Foods, 11(21), 3367. https://doi.org/10.3390/foods11213367