
Fish Hydrolysate Supplementation Prevents Stress-Induced Dysregulation of Hippocampal Proteins Relative to Mitochondrial Metabolism and the Neuronal Network in Mice

 , , and
, , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Fish Hydrolysate
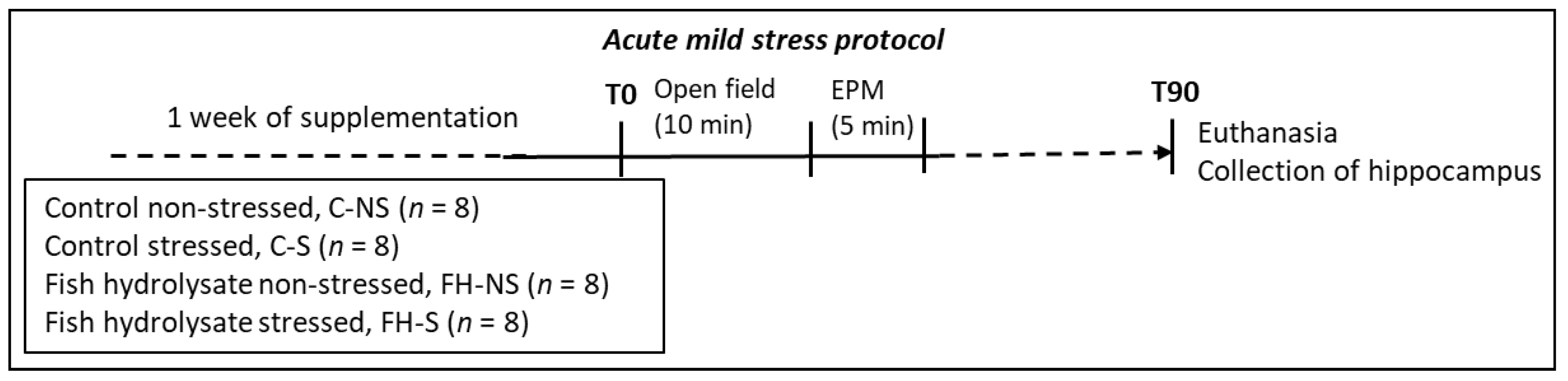
2.2. Experimental Design
2.3. Label-Free Quantitative Proteomics
2.3.1. Protein Extraction
2.3.2. Nanoliquid Chromatography Coupled with Tandem Mass Spectrometry (NanoLC-MS/MS)
2.3.3. Quantification and Statistical Analyses of Proteomics Data
3. Results
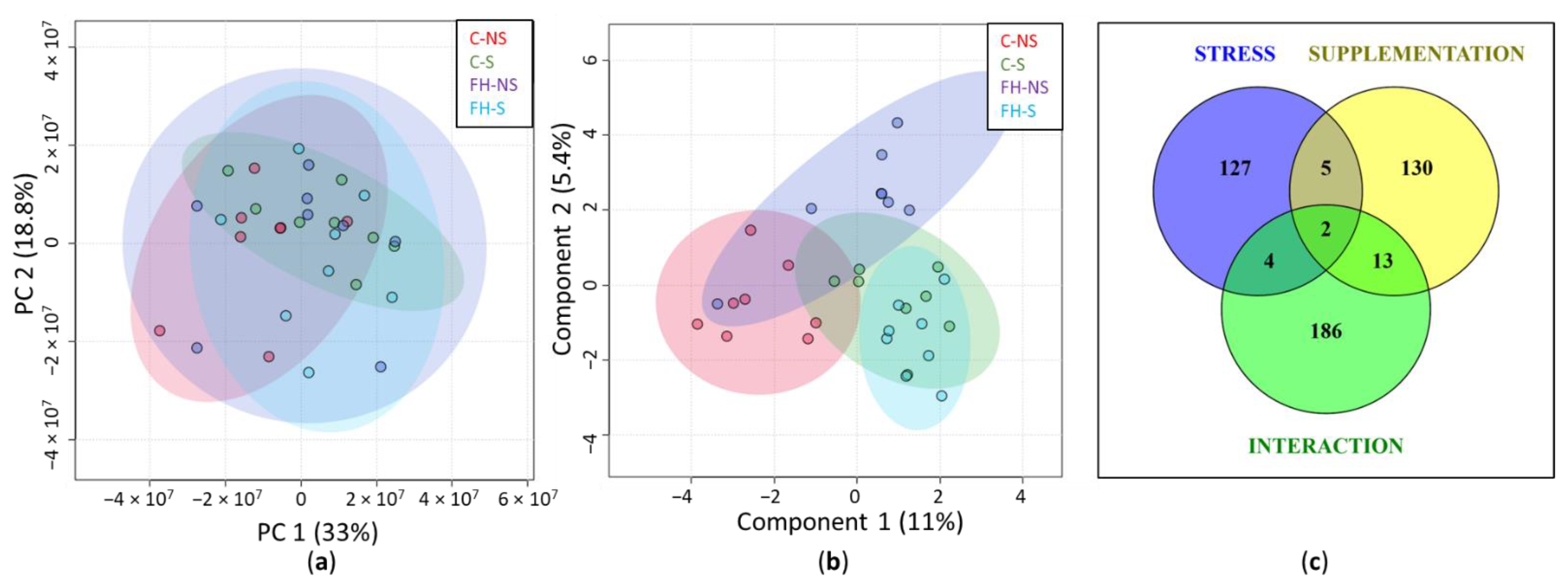
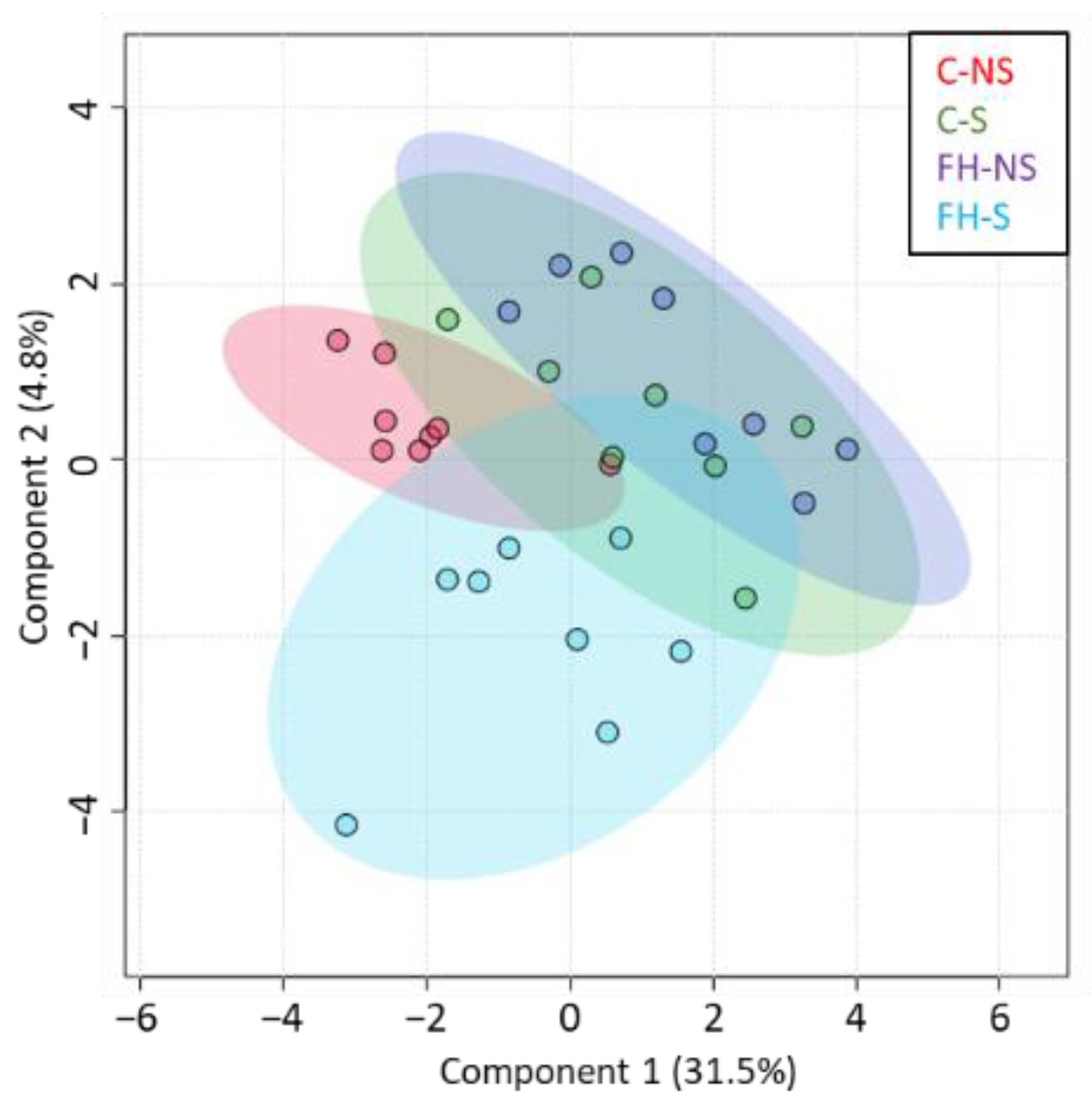
3.1. Hippocampal Protein Expression Is Modulated by Stress, Supplementation, and the Interaction Stress × Supplementation
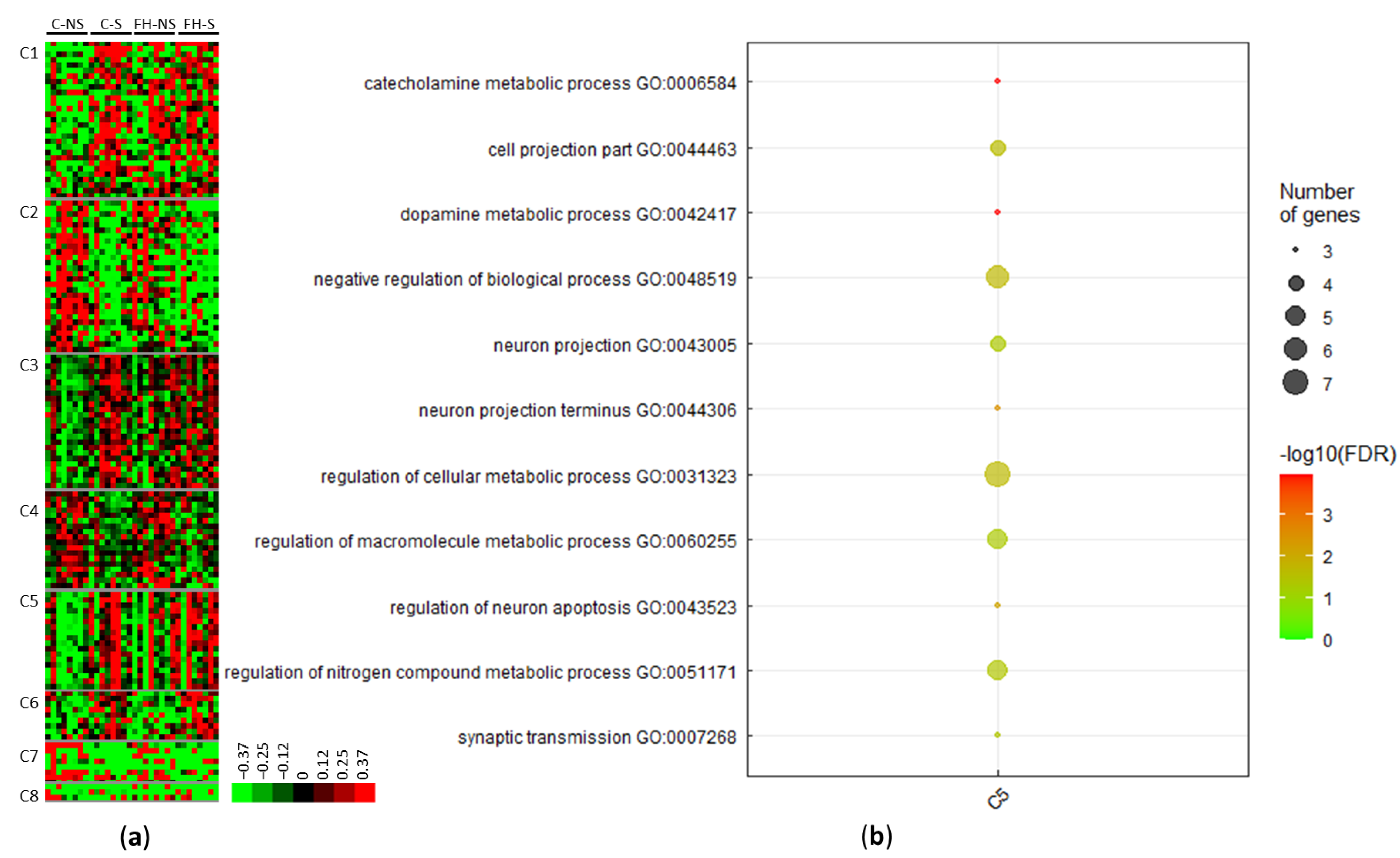
3.2. Stress Modulates the Expression of Proteins Involved in Dopamine Metabolism, Neuron Projection, and Metabolic Processes
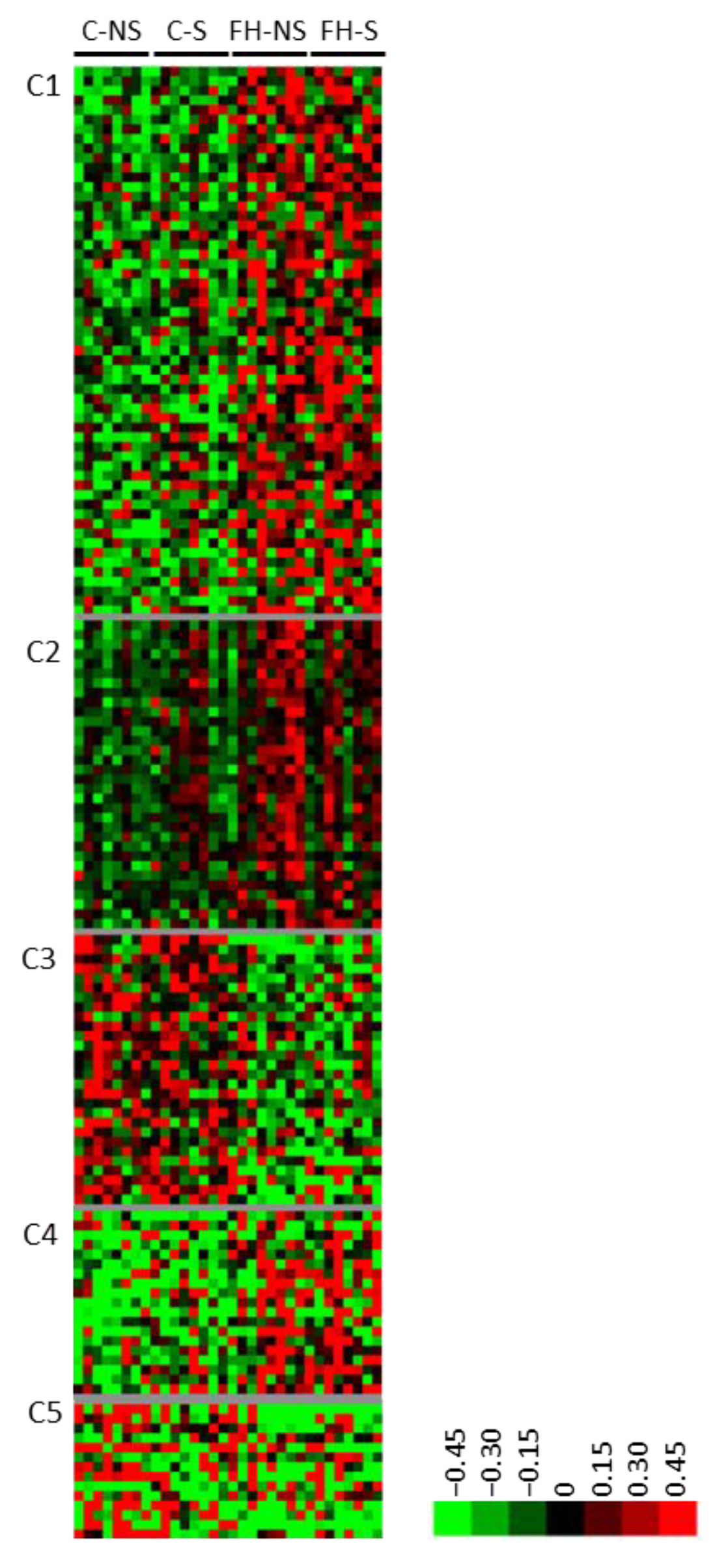
3.3. Fish Hydrolysate Supplementation Modulates the Expression of Proteins Involved in Metabolic Processes and the Neuronal Network
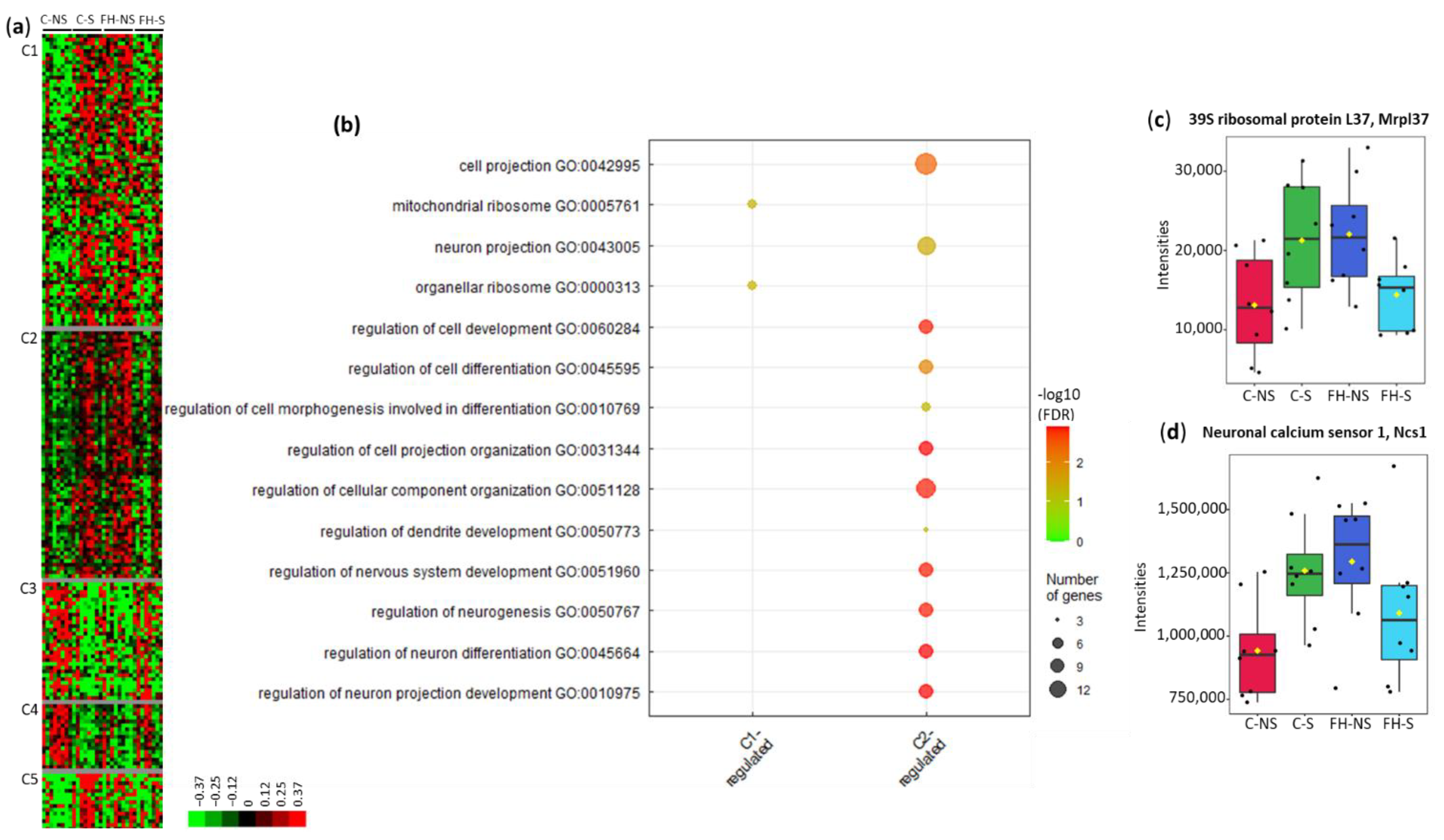
3.4. Fish Hydrolysate Supplementation Prevents Stress-Induced Dysregulation of Proteins Relative to Mitochondrial Pathways and the Neuronal Network
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- OSH in Figures: Stress at Work-Facts and Figures|Safety and Health at Work EU-OSHA. Available online: https://osha.europa.eu/en/publications/osh-figures-stress-work-facts-and-figures/view-0 (accessed on 14 February 2022).
- Pariante, C.M.; Lightman, S.L. The HPA Axis in Major Depression: Classical Theories and New Developments. Trends Neurosci. 2008, 31, 464–468. [Google Scholar] [CrossRef] [PubMed]
- Kellendonk, C.; Gass, P.; Kretz, O.; Schütz, G.; Tronche, F. Corticosteroid Receptors in the Brain: Gene Targeting Studies. Brain Res. Bull. 2002, 57, 73–83. [Google Scholar] [CrossRef]
- Coppola, D.; Lauritano, C.; Palma Esposito, F.; Riccio, G.; Rizzo, C.; de Pascale, D. Fish Waste: From Problem to Valuable Resource. Mar. Drugs 2021, 19, 116. [Google Scholar] [CrossRef] [PubMed]
- Carrera, M.; Cañas, B.; Gallardo, J.M. The Sarcoplasmic Fish Proteome: Pathways, Metabolic Networks and Potential Bioactive Peptides for Nutritional Inferences. J. Proteom. 2013, 78, 211–220. [Google Scholar] [CrossRef]
- Gevaert, B.; Veryser, L.; Verbeke, F.; Wynendaele, E.; De Spiegeleer, B. Fish Hydrolysates: A Regulatory Perspective of Bioactive Peptides. Protein Pept. Lett. 2016, 23, 1052–1060. [Google Scholar] [CrossRef]
- Zamora-Sillero, J.; Gharsallaoui, A.; Prentice, C. Peptides from Fish By-Product Protein Hydrolysates and Its Functional Properties: An Overview. Mar. Biotechnol. 2018, 20, 118–130. [Google Scholar] [CrossRef]
- Belhaj, N.; Desor, F.; Gleizes, C.; Denis, F.M.; Arab-Tehrany, E.; Soulimani, R.; Linder, M. Anxiolytic-like Effect of a Salmon Phospholipopeptidic Complex Composed of Polyunsaturated Fatty Acids and Bioactive Peptides. Mar. Drugs 2013, 11, 4294–4317. [Google Scholar] [CrossRef]
- Chataigner, M.; Mortessagne, P.; Lucas, C.; Pallet, V.; Layé, S.; Mehaignerie, A.; Bouvret, E.; Dinel, A.L.; Joffre, C. Dietary Fish Hydrolysate Supplementation Containing N-3 LC-PUFAs and Peptides Prevents Short-Term Memory and Stress Response Deficits in Aged Mice. Brain Behav. Immun. 2021, 91, 716–730. [Google Scholar] [CrossRef]
- Bernet, F.; Montel, V.; Noël, B.; Dupouy, J.P. Diazepam-like Effects of a Fish Protein Hydrolysate (Gabolysat PC60) on Stress Responsiveness of the Rat Pituitary-Adrenal System and Sympathoadrenal Activity. Psychopharmacology 2000, 149, 34–40. [Google Scholar] [CrossRef]
- Messaoudi, M.; Nejdi, A.; Bisson, J.-F.; Rozan, P.; Javelot, H.; Lalonde, R. Anxiolytic and Antidepressant-Like Effects Of Garum Armoricum® (GA), A Blue Ling Fish Protein Autolysate In Male Wistar Rats. Curr. Top. Nutraceutical Res. 2008, 6, 115–123. [Google Scholar]
- Freret, T.; Largilliere, S.; Nee, G.; Coolzaet, M.; Corvaisier, S.; Boulouard, M. Fast Anxiolytic-Like Effect Observed in the Rat Conditioned Defensive Burying Test, after a Single Oral Dose of Natural Protein Extract Products. Nutrients 2021, 13, 2445. [Google Scholar] [CrossRef] [PubMed]
- Messaoudi, M.; Lefranc–Millot, C.; Desor, D.; Demagny, B.; Bourdon, L. Effects of a Tryptic Hydrolysate Frombovine Milk AS1–Casein on Hemodynamicresponses in Healthy Human Volunteersfacing Successive Mental and Physicalstress Situations. Eur. J. Nutr. 2005, 44, 128–132. [Google Scholar] [CrossRef] [PubMed]
- Dinel, A.-L.; Lucas, C.; Le Faouder, J.; Bouvret, E.; Pallet, V.; Layé, S.; Joffre, C. Supplementation with Low Molecular Weight Peptides from Fish Protein Hydrolysate Reduces Acute Mild Stress-Induced Corticosterone Secretion and Modulates Stress Responsive Gene Expression in Mice. J. Funct. Foods 2021, 76, 104292. [Google Scholar] [CrossRef]
- Hill, A.S.; Sahay, A.; Hen, R. Increasing Adult Hippocampal Neurogenesis Is Sufficient to Reduce Anxiety and Depression-Like Behaviors. Neuropsychopharmacology 2015, 40, 2368–2378. [Google Scholar] [CrossRef]
- Simões, A.E.; Pereira, D.M.; Amaral, J.D.; Nunes, A.F.; Gomes, S.E.; Rodrigues, P.M.; Lo, A.C.; D’Hooge, R.; Steer, C.J.; Thibodeau, S.N.; et al. Efficient Recovery of Proteins from Multiple Source Samples after Trizol® or Trizol®LS RNA Extraction and Long-Term Storage. BMC Genom. 2013, 14, 181. [Google Scholar] [CrossRef]
- Méar, L.; Com, E.; Fathallah, K.; Guillot, L.; Lavigne, R.; Guével, B.; Fauconnier, A.; Vialard, F.; Pineau, C. The Eutopic Endometrium Proteome in Endometriosis Reveals Candidate Markers and Molecular Mechanisms of Physiopathology. Diagnostics 2022, 12, 419. [Google Scholar] [CrossRef]
- Achcar, F.; Camadro, J.-M.; Mestivier, D. AutoClass@IJM: A Powerful Tool for Bayesian Classification of Heterogeneous Data in Biology. Nucleic Acids Res. 2009, 37, W63–W67. [Google Scholar] [CrossRef]
- Maere, S.; Heymans, K.; Kuiper, M. BiNGO: A Cytoscape Plugin to Assess Overrepresentation of Gene Ontology Categories in Biological Networks. Bioinformatics 2005, 21, 3448–3449. [Google Scholar] [CrossRef]
- Bonnot, T.; Gillard, M.B.; Nagel, D.H. A Simple Protocol for Informative Visualization of Enriched Gene Ontology Terms. Bio-Protocol 2019, 9, e3429. [Google Scholar] [CrossRef]
- Xia, J.; Psychogios, N.; Young, N.; Wishart, D.S. MetaboAnalyst: A Web Server for Metabolomic Data Analysis and Interpretation. Nucleic Acids Res. 2009, 37, W652–W660. [Google Scholar] [CrossRef]
- Wersinger, C.; Prou, D.; Vernier, P.; Sidhu, A. Modulation of Dopamine Transporter Function by α-Synuclein Is Altered by Impairment of Cell Adhesion and by Induction of Oxidative Stress. FASEB J. 2003, 17, 1–30. [Google Scholar] [CrossRef] [PubMed]
- Chiavegatto, S.; Izidio, G.S.; Mendes-Lana, A.; Aneas, I.; Freitas, T.A.; Torrão, A.S.; Conceição, I.M.; Britto, L.R.G.; Ramos, A. Expression of α-Synuclein Is Increased in the Hippocampus of Rats with High Levels of Innate Anxiety. Mol. Psychiatry 2009, 14, 894–905. [Google Scholar] [CrossRef]
- Carboni, L.; Piubelli, C.; Pozzato, C.; Astner, H.; Arban, R.; Righetti, P.G.; Hamdan, M.; Domenici, E. Proteomic Analysis of Rat Hippocampus after Repeated Psychosocial Stress. Neuroscience 2006, 137, 1237–1246. [Google Scholar] [CrossRef]
- Foote, M.; Qiao, H.; Graham, K.; Wu, Y.; Zhou, Y. Inhibition of 14-3-3 Proteins Leads to Schizophrenia-Related Behavioral Phenotypes and Synaptic Defects in Mice. Biol. Psychiatry 2015, 78, 386–395. [Google Scholar] [CrossRef] [PubMed]
- Huang, G.; Li, H.; Zhang, H. Abnormal Expression of Mitochondrial Ribosomal Proteins and Their Encoding Genes with Cell Apoptosis and Diseases. Int. J. Mol. Sci. 2020, 21, 8879. [Google Scholar] [CrossRef] [PubMed]
- Mozhui, K.; Snively, B.M.; Rapp, S.R.; Wallace, R.B.; Williams, R.W.; Johnson, K.C. Genetic Analysis of Mitochondrial Ribosomal Proteins and Cognitive Aging in Postmenopausal Women. Front. Genet. 2017, 8, 127. [Google Scholar] [CrossRef] [PubMed]
- Picard, M.; McEwen, B.S.; Epel, E.S.; Sandi, C. An Energetic View of Stress: Focus on Mitochondria. Front. Neuroendocrinol. 2018, 49, 72–85. [Google Scholar] [CrossRef] [PubMed]
- Filiou, M.D.; Sandi, C. Anxiety and Brain Mitochondria: A Bidirectional Crosstalk. Trends Neurosci. 2019, 42, 573–588. [Google Scholar] [CrossRef]
- Tang, M.; Huang, H.; Li, S.; Zhou, M.; Liu, Z.; Huang, R.; Liao, W.; Xie, P.; Zhou, J. Hippocampal Proteomic Changes of Susceptibility and Resilience to Depression or Anxiety in a Rat Model of Chronic Mild Stress. Transl. Psychiatry 2019, 9, 1–12. [Google Scholar] [CrossRef]
- Misiewicz, Z.; Iurato, S.; Kulesskaya, N.; Salminen, L.; Rodrigues, L.; Maccarrone, G.; Martins, J.; Czamara, D.; Laine, M.A.; Sokolowska, E.; et al. Multi-Omics Analysis Identifies Mitochondrial Pathways Associated with Anxiety-Related Behavior. PLoS Genet. 2019, 15, e1008358. [Google Scholar] [CrossRef]
- Zhang, X.; Lei, K.; Yuan, X.; Wu, X.; Zhuang, Y.; Xu, T.; Xu, R.; Han, M. SUN1/2 and Syne/Nesprin-1/2 Complexes Connect Centrosome to the Nucleus during Neurogenesis and Neuronal Migration in Mice. Neuron 2009, 64, 173–187. [Google Scholar] [CrossRef] [PubMed]
- Fischer, T.T.; Nguyen, L.D.; Ehrlich, B.E. Neuronal Calcium Sensor 1 (NCS1) Dependent Modulation of Neuronal Morphology and Development. FASEB J. 2021, 35, e21873. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Jimenez, F.J.; Vilches, A.; Perez-Arago, M.A.; Clemente, E.; Roman, R.; Leal, J.; Castro, A.A.; Fustero, S.; Moreno-Manzano, V.; Jendelova, P.; et al. Activation of Neurogenesis in Multipotent Stem Cells Cultured In Vitro and in the Spinal Cord Tissue After Severe Injury by Inhibition of Glycogen Synthase Kinase-3. Neurotherapeutics 2021, 18, 515–533. [Google Scholar] [CrossRef]
- Parnell, E.; Shapiro, L.P.; Voorn, R.A.; Forrest, M.P.; Jalloul, H.A.; Loizzo, D.D.; Penzes, P. KALRN: A Central Regulator of Synaptic Function and Synaptopathies. Gene 2021, 768, 145306. [Google Scholar] [CrossRef]
- Pittenger, C.; Duman, R.S. Stress, Depression, and Neuroplasticity: A Convergence of Mechanisms. Neuropsychopharmacol 2008, 33, 88–109. [Google Scholar] [CrossRef] [PubMed]
- Bannerman, D.M.; Sprengel, R.; Sanderson, D.J.; McHugh, S.B.; Rawlins, J.N.P.; Monyer, H.; Seeburg, P.H. Hippocampal Synaptic Plasticity, Spatial Memory and Anxiety. Nat. Rev. Neurosci. 2014, 15, 181–192. [Google Scholar] [CrossRef]
- Curtiss, J.; Klemanski, D.H. Taxonicity and Network Structure of Generalized Anxiety Disorder and Major Depressive Disorder: An Admixture Analysis and Complex Network Analysis. J. Affect. Disord. 2016, 199, 99–105. [Google Scholar] [CrossRef]
- Kabbani, N.; Woll, M.P.; Nordman, J.C.; Levenson, R. Dopamine Receptor Interacting Proteins: Targeting Neuronal Calcium Sensor-1/D2 Dopamine Receptor Interaction for Antipsychotic Drug Development. Curr. Drug Targets 2012, 13, 72–79. [Google Scholar] [CrossRef]
- Chen, S.; Owens, G.C.; Edelman, D.B. Dopamine Inhibits Mitochondrial Motility in Hippocampal Neurons. PLoS ONE 2008, 3, e2804. [Google Scholar] [CrossRef]
- Qiao, H.; Yang, S.; Xu, C.; Ma, X.-M.; An, S.-C. Involvement of D2 Receptor in the NAc in Chronic Unpredictable Stress-Induced Depression-like Behaviors. Stress 2020, 23, 318–327. [Google Scholar] [CrossRef]
- Mandela, P.; Yan, Y.; LaRese, T.; Eipper, B.A.; Mains, R.E. Elimination of Kalrn Expression in POMC Cells Reduces Anxiety-like Behavior and Contextual Fear Learning. Horm. Behav. 2014, 66, 430–438. [Google Scholar] [CrossRef][Green Version]
- Schmutz, I.; Wendt, S.; Schnell, A.; Kramer, A.; Mansuy, I.M.; Albrecht, U. Protein Phosphatase 1 (PP1) Is a Post-Translational Regulator of the Mammalian Circadian Clock. PLoS ONE 2011, 6, e21325. [Google Scholar] [CrossRef] [PubMed]
- Besing, R.C.; Rogers, C.O.; Paul, J.R.; Hablitz, L.M.; Johnson, R.L.; McMahon, L.L.; Gamble, K.L. GSK3 Activity Regulates Rhythms in Hippocampal Clock Gene Expression and Synaptic Plasticity. Hippocampus 2017, 27, 890–898. [Google Scholar] [CrossRef] [PubMed]
- Crofton, E.J.; Nenov, M.N.; Zhang, Y.; Scala, F.; Page, S.A.; McCue, D.L.; Li, D.; Hommel, J.D.; Laezza, F.; Green, T.A. Glycogen Synthase Kinase 3 Beta Alters Anxiety-, Depression-, and Addiction-Related Behaviors and Neuronal Activity in the Nucleus Accumbens Shell. Neuropharmacology 2017, 117, 49–60. [Google Scholar] [CrossRef] [PubMed]
- Perez-Riverol, Y.; Bai, J.; Bandla, C.; García-Seisdedos, D.; Hewapathirana, S.; Kamatchinathan, S.; Kundu, D.J.; Prakash, A.; Frericks-Zipper, A.; Eisenacher, M.; et al. The PRIDE Database Resources in 2022: A Hub for Mass Spectrometry-Based Proteomics Evidences. Nucleic Acids Res. 2022, 50, D543–D552. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Composition | Fish Hydrolysate |
|---|---|
| Total proteins | ≥70% |
| Minerals | ≤20% |
| Lipids | ≤5% |
| Composition of Amino Acids | Fish Hydrolysate |
|---|---|
| Essential Amino Acids | 43% |
| Non-essential Amino Acids | 56% |
| Branched-Chain Amino Acids | 17% |
| Sulfur Amino Acids | 3% |
| Pathway Name | Protein IDs | Protein Name | Gene Name | Expression C-S/FH-S |
|---|---|---|---|---|
| Dopamine metabolic | P00493 | Hypoxanthine-guanine phosphoribosyltransferase | Hprt1 | up |
| Process * | Q91ZZ3 | Beta-synuclein | Sncb | up |
| O55042 | Alpha-synuclein | Snca | up | |
| Neuron projection * | Q08460 | Calcium-activated potassium channel subunit alpha-1 | Kcnma | up |
| A2A8L5 | Receptor-type tyrosine-protein phosphatase F | Ptprf | up | |
| Q91ZZ3 | Beta-synuclein | Sncb | up | |
| O55042 | Alpha-synuclein | Snca | up | |
| Regulation | Q9EQQ9 | Protein O-GlcNAcase | Oga | up |
| of macromolecule metabolic | Q8CI32 | BAG family molecular chaperone regulator 5 | Bag5 | up |
| Process * | P29595 | NEDD8 | Nedd8 | up |
| O55042 | Alpha-synuclein | Snca | up | |
| Q9Z1X4 | Interleukin enhancer-binding factor 3 | Ilf3 | up |
| Pathway Name | Protein IDs | Protein Name | Gene Name | Expression FH-S vs. C-S |
|---|---|---|---|---|
| Mitochondrial | Q9Z2Q5 | 39S ribosomal protein L40 | Mrpl40 | down |
| Ribosome * | Q9D7N6 | 39S ribosomal protein L30 | Mrpl30 | down |
| Q921S7 | 39S ribosomal protein L37 | Mrpl37 | down | |
| Q9ER88 | 28S ribosomal protein S29 | Dap3 | down | |
| Mitochondrion | P09528 | Ferritin heavy chain | Fth1 | down |
| Q8BK30 | NADH dehydrogenase [ubiquinone] flavoprotein 3 | Ndufv3 | down | |
| Q9Z2Q5 | 39S ribosomal protein L40 | Mrpl40 | down | |
| Q9CZL5 | Pterin-4-alpha-carbinolamine dehydratase 2 | Pcbd2 | down | |
| Q9CQQ7 | ATP synthase F(0) complex subunit B1 | Atp5pb | down | |
| Q9D7N6 | 39S ribosomal protein L30 | Mrpl30 | down | |
| Q921S7 | 39S ribosomal protein L37 | Mrpl37 | down | |
| Q9ER88 | 28S ribosomal protein S29 | Dap3 | down | |
| Q9CR21 | Acyl carrier protein | Ndufab1 | down | |
| Q9CQX8 | 28S ribosomal protein S36 | Mrps36 | down | |
| P62897 | Cytochrome c, somatic | Cycs | down | |
| Oxidation | P09528 | Ferritin heavy chain | Fth1 | down |
| Reduction | Q8BK30 | NADH dehydrogenase [ubiquinone] flavoprotein 3 | Ndufv3 | down |
| Q9CQF9 | Prenylcysteine oxidase | Pcyox1 | down | |
| Q8BGW1 | Alpha-ketoglutarate-dependent dioxygenase FTO | Fto | down | |
| Q9CR21 | Acyl carrier protein | Ndufab1 | down | |
| Q8K0C4 | Lanosterol 14-alpha demethylase | Cyp51a1 | down | |
| P62897 | Cytochrome c, somatic | Cycs | down | |
| Lipid metabolism | Q6P549 | Phosphatidylinositol 3,4,5-trisphosphate 5-phosphatase 2 | Inppl1 | down |
| Q80Y98 | Phospholipase DDHD2 | Ddhd2 | down | |
| Q9CR21 | Acyl carrier protein | Ndufab1 | down | |
| Q8K0C4 | Lanosterol 14-alpha demethylase | Cyp51a1 | down | |
| Q00915 | Retinol-binding protein 1 | Rbp1 | down | |
| Q9D2R0 | Acetoacetyl-CoA synthetase | Aacs | down |
| Pathway Name | Protein IDs | Protein Name | Gene Name | Expression FH-S vs. C-S |
|---|---|---|---|---|
| Regulation of | A2CG49 | Kalirin | Kalrn | down |
| Neuron | Q9WV60 | Glycogen synthase kinase-3 beta | Gsk3b | down |
| Projection | P08553 | Neurofilament medium polypeptide | Nefm | down |
| Development * | Q99P72 | Reticulon-4 | Rtn4 | down |
| Q8BNY6 | Neuronal calcium sensor 1 | Ncs1 | down | |
| Q6ZWR6 | Nesprin-1 | Syne1 | down | |
| Q5SNZ0 | Girdin | Ccdc88a | down | |
| Regulation of | Q61301 | Catenin alpha-2 | Ctnna2 | down |
| Cellular | A2CG49 | Kalirin | Kalrn | down |
| Component | Q9WV60 | Glycogen synthase kinase-3 beta | Gsk3b | down |
| Organization * | P08553 | Neurofilament medium polypeptide | Nefm | down |
| P27546 | Microtubule-associated protein 4 | Map4 | down | |
| P47757 | F-actin-capping protein subunit beta | Capzb | down | |
| Q99P72 | Reticulon-4 | Rtn4 | down | |
| P70336 | Rho-associated protein kinase 2 | Rock2 | down | |
| Q8BNY6 | Neuronal calcium sensor 1 | Ncs1 | down | |
| Q6ZWR6 | Nesprin-1 | Syne1 | down | |
| Q5SNZ0 | Girdin | Ccdc88a | down |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Le Faouder, J.; Arnaud, B.; Lavigne, R.; Lucas, C.; Com, E.; Bouvret, E.; Dinel, A.-L.; Pineau, C. Fish Hydrolysate Supplementation Prevents Stress-Induced Dysregulation of Hippocampal Proteins Relative to Mitochondrial Metabolism and the Neuronal Network in Mice. Foods 2022, 11, 1591. https://doi.org/10.3390/foods11111591
Le Faouder J, Arnaud B, Lavigne R, Lucas C, Com E, Bouvret E, Dinel A-L, Pineau C. Fish Hydrolysate Supplementation Prevents Stress-Induced Dysregulation of Hippocampal Proteins Relative to Mitochondrial Metabolism and the Neuronal Network in Mice. Foods. 2022; 11(11):1591. https://doi.org/10.3390/foods11111591
Chicago/Turabian StyleLe Faouder, Julie, Bastien Arnaud, Régis Lavigne, Céline Lucas, Emmanuelle Com, Elodie Bouvret, Anne-Laure Dinel, and Charles Pineau. 2022. "Fish Hydrolysate Supplementation Prevents Stress-Induced Dysregulation of Hippocampal Proteins Relative to Mitochondrial Metabolism and the Neuronal Network in Mice" Foods 11, no. 11: 1591. https://doi.org/10.3390/foods11111591
APA StyleLe Faouder, J., Arnaud, B., Lavigne, R., Lucas, C., Com, E., Bouvret, E., Dinel, A.-L., & Pineau, C. (2022). Fish Hydrolysate Supplementation Prevents Stress-Induced Dysregulation of Hippocampal Proteins Relative to Mitochondrial Metabolism and the Neuronal Network in Mice. Foods, 11(11), 1591. https://doi.org/10.3390/foods11111591