
Polyphenolic Characterization of Merlot, Tannat and Syrah Skin Extracts at Different Degrees of Maturity and Anti-Inflammatory Potential in RAW 264.7 Cells

 , , ,
, , ,  and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Grape Samples
2.3. Grape Samples Preparation and Polyphenolic Extraction
2.4. Determination of Total Polyphenol Content (TPC)
2.5. Determination of Radical Scavenging Activity (DPPH˙ Assay)
2.6. Analysis of Proanthocyanidins Following Acid Catalysis with Phloroglucinol
2.7. Individual Determination of Phenolic Composition
2.7.1. Momonomeric Flavan-3-ols, Procyanidins, Flavonols and Stilbenes
2.7.2. Anthocyanins
2.8. Cell Culture and Treatment
2.9. MTT Cell Viability
2.10. Intracellular NO Measurement
2.11. Intracellular Reactive Oxygen Species (ROS) Measurement
2.12. Statistical Analysis
3. Results and Discussion
3.1. TPC and Antioxidant Activity of Skins of Merlot, Tannat and Syrah Cultivars at Different Stages of Ripening
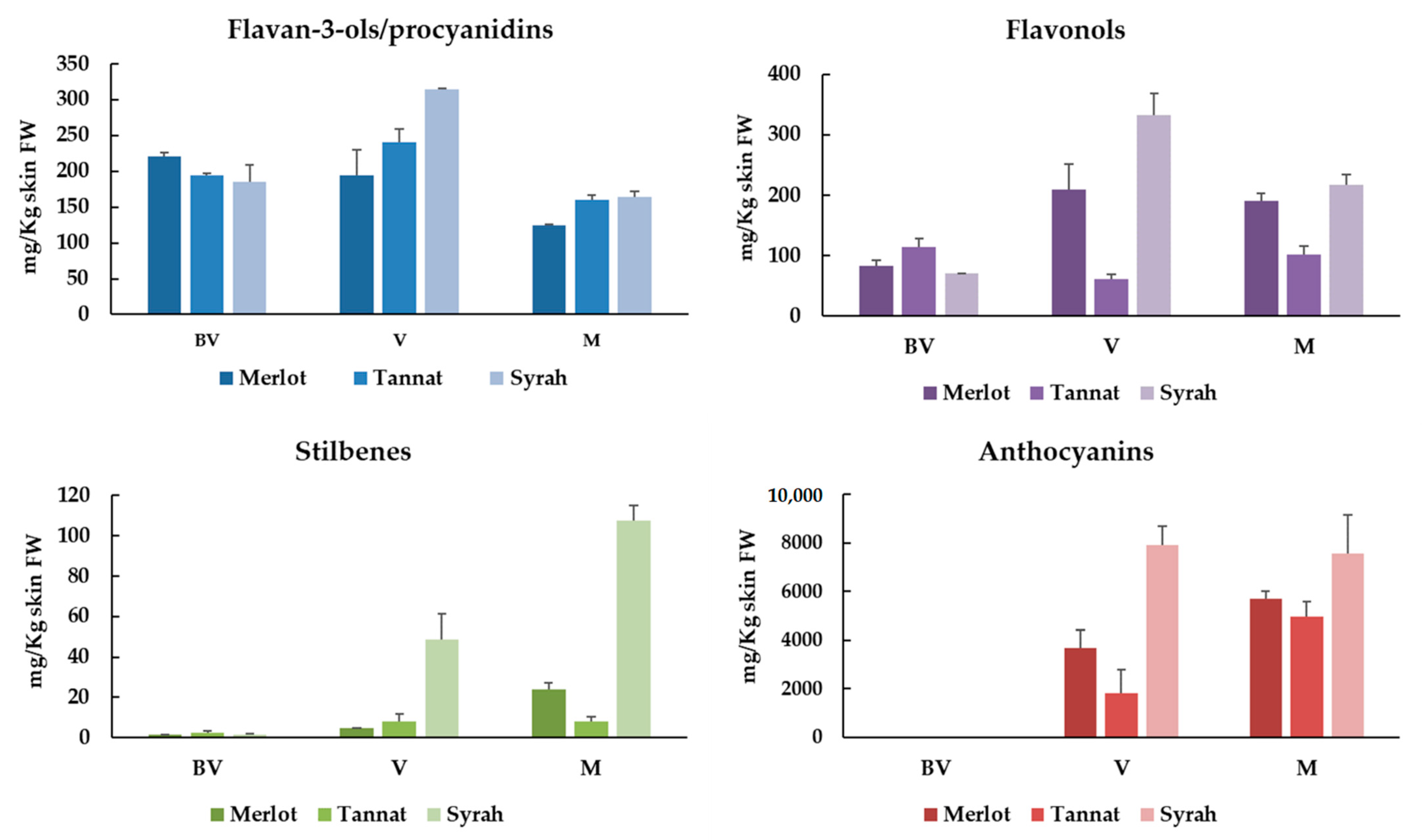
3.2. Individual Polyphenolic Characterization of Skins of Merlot, Tannat and Syrah Cultivars at Different Stages of Ripening
3.2.1. Monomeric Flavan-3-ols and Procyanidins
3.2.2. Flavonols
3.2.3. Stilbenes
3.2.4. Anthocyanins
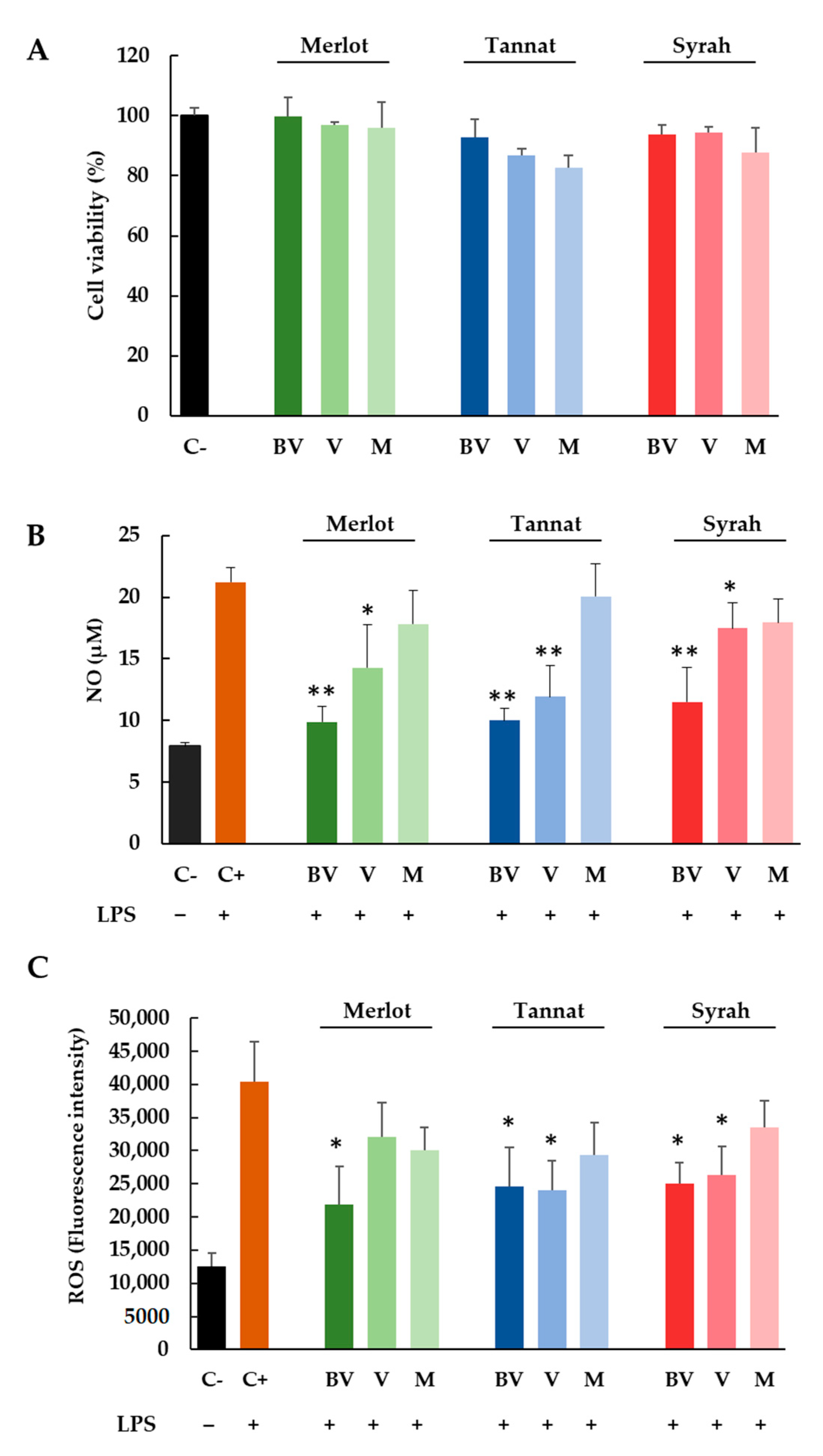
3.3. Anti-Inflammatory and Antioxidant Potential In Vitro Effects of Skins Extracts of Merlot, Tannat and Syrah Cultivars at Different Stages of Ripening
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lacombe, T. Contribution à l’étude de l’histoire Évolutive de La Vigne Cultivée (Vitis Vinifera L.) Par l’analyse de La Diversité Génétique Neutre et de Gènes d’intérêt. Ph.D. Thesis, Montpellier SupAgro, Montpellier, France, 2012. [Google Scholar]
- Han, X.; Shen, T.; Lou, H. Dietary Polyphenols and Their Biological Significance. Int. J. Mol. Sci. 2007, 8, 950–988. [Google Scholar] [CrossRef]
- Vauzour, D.; Rodriguez-Mateos, A.; Corona, G.; Oruna-Concha, M.J.; Spencer, J.P.E. Polyphenols and Human Health: Prevention of Disease and Mechanisms of Action. Nutrients 2010, 2, 1106–1131. [Google Scholar] [CrossRef]
- Saltveit, M.E. Synthesis and Metabolism of Phenolic Compounds. In Fruit and Vegetable Phytochemicals; John Wiley & Sons, Ltd: Hoboken, NJ, USA, 2009; pp. 89–100. [Google Scholar] [CrossRef]
- Yilmaz, Y.; Göksel, Z.; Erdoğan, S.S.; Öztürk, A.; Atak, A.; Özer, C. Antioxidant Activity and Phenolic Content of Seed, Skin and Pulp Parts of 22 Grape (Vitis Vinifera L.) Cultivars (4 Common and 18 Registered or Candidate for Registration). J. Food Process. Preserv. 2015, 39, 1682–1691. [Google Scholar] [CrossRef]
- Ashraf, M.A.; Iqbal, M.; Rasheed, R.; Hussain, I.; Riaz, M.; Arif, M.S. Environmental Stress and Secondary Metabolites in Plants. In Plant Metabolites and Regulation Under Environmental Stress; Elsevier: Amsterdam, The Netherlands, 2018; pp. 153–167. [Google Scholar] [CrossRef]
- Hornedo-Ortega, R.; Reyes González-Centeno, M.; Chira, K.; Jourdes, M.; Teissedre, P.-L. Phenolic Compounds of Grapes and Wines: Key Compounds and Implications in Sensory Perception. In Winemaking—Stabilization, Aging Chemistry and Biochemistry [Working Title]; IntechOpen: London, UK, 2020. [Google Scholar] [CrossRef]
- Ivanova, V.; Stefova, M.; Vojnoski, B.; Dörnyei, Á.; Márk, L.; Dimovska, V.; Stafilov, T.; Kilár, F. Identification of Polyphenolic Compounds in Red and White Grape Varieties Grown in R. Macedonia and Changes of Their Content during Ripening. Food Res. Int. 2011, 44, 2851–2860. [Google Scholar] [CrossRef]
- Obreque-Slier, E.; Peña-Neira, A.; López-Solís, R.; Zamora-Marín, F.; Ricardo-da Silva, J.M.; Laureano, O. Comparative Study of the Phenolic Composition of Seeds and Skins from Carménère and Cabernet Sauvignon Grape Varieties (Vitis Vinifera L.) during Ripening. J. Agric. Food Chem. 2010, 58, 3591–3599. [Google Scholar] [CrossRef]
- Obreque-Slier, E.; Peña-Neira, Á.; López-Solís, R.; Cáceres-Mella, A.; Toledo-Araya, H.; López-Rivera, A. Phenolic Composition of Skins from Four Carmenet Grape Varieties (Vitis Vinifera L.) during Ripening. LWT Food Sci. Technol. 2013, 54, 404–413. [Google Scholar] [CrossRef]
- di Giuseppe, R.; de Lorgeril, M.; Salen, P.; Laporte, F.; Di Castelnuovo, A.; Krogh, V.; Siani, A.; Arnout, J.; Cappuccio, F.P.; van Dongen, M.; et al. European Collaborative Group of the IMMIDIET Project. Alcohol Consumption and N-3 Polyunsaturated Fatty Acids in Healthy Men and Women from 3 European Populations. Am. J. Clin. Nutr. 2009, 89, 354–362. [Google Scholar] [CrossRef]
- Urquiaga, I.; Guasch, V.; Marshall, G.; San Martín, A.; Castillo, O.; Rozowski, J.; Leighton, F. Effect of Mediterranean and Occidental Diets, and Red Wine, on Plasma Fatty Acids in Humans. An Intervention Study. Biol. Res. 2004, 37, 253–261. [Google Scholar] [CrossRef]
- Pasinetti, G.M. Novel Role of Red Wine-Derived Polyphenols in the Prevention of Alzheimer’s Disease Dementia and Brain Pathology: Experimental Approaches and Clinical Implications. Planta Med. 2012, 78, E24. [Google Scholar] [CrossRef] [PubMed]
- Rasines-Perea, Z.; Teissedre, P.-L. Grape Polyphenols’ Effects in Human Cardiovascular Diseases and Diabetes. Molecules 2017, 22, 68. [Google Scholar] [CrossRef]
- Khansari, N.; Shakiba, Y.; Mahmoudi, M. Chronic Inflammation and Oxidative Stress as a Major Cause of Age-Related Diseases and Cancer. Recent Pat. Inflamm. Allergy Drug Discov. 2009. [Google Scholar] [CrossRef]
- Singleton, V.L.; Orthofer, R.; Lamuela-Raventós, R.M. Analysis of Total Phenols and Other Oxidation Substrates and Antioxidants by Means of Folin-Ciocalteu Reagent. In Methods in Enzymology; Oxidants and Antioxidants Part A; Academic Press: Cambridge, MA, USA, 1999; Volume 299, pp. 152–178. [Google Scholar] [CrossRef]
- Villaño, D.; Fernández-Pachón, M.S.; Moyá, M.L.; Troncoso, A.M.; García-Parrilla, M.C. Radical Scavenging Ability of Polyphenolic Compounds towards DPPH Free Radical. Talanta 2007, 71, 230–235. [Google Scholar] [CrossRef]
- Kennedy, J.A.; Jones, G.P. Analysis of Proanthocyanidin Cleavage Products Following Acid-Catalysis in the Presence of Excess Phloroglucinol. J. Agric. Food Chem. 2001, 49, 1740–1746. [Google Scholar] [CrossRef] [PubMed]
- Loupit, G.; Prigent, S.; Franc, C.; De Revel, G.; Richard, T.; Cookson, S.J.; Fonayet, J.V. Polyphenol Profiles of Just Pruned Grapevine Canes from Wild Vitis Accessions and Vitis Vinifera Cultivars. J. Agric. Food Chem. 2020. [Google Scholar] [CrossRef]
- Giuffrè, A.M. HPLC-DAD Detection of Changes in Phenol Content of Red Berry Skins during Grape Ripening. Eur. Food Res. Technol. 2013, 237, 555–564. [Google Scholar] [CrossRef]
- Mosmann, T. Rapid Colorimetric Assay for Cellular Growth and Survival: Application to Proliferation And. J. Inmunol. Methods 1983, 55, 55–63. [Google Scholar] [CrossRef]
- Boido, E.; García-Marino, M.; Dellacassa, E.; Carrau, F.; Rivas-Gonzalo, J.C.; Escribano-Bailón, M.T. Characterisation and Evolution of Grape Polyphenol Profiles of Vitis Vinifera L. Cv. Tannat during Ripening and Vinification. Aust. J. Grape Wine Res. 2011, 17, 383–393. [Google Scholar] [CrossRef]
- Lorrain, B.; Chira, K.; Teissedre, P.-L. Phenolic Composition of Merlot and Cabernet-Sauvignon Grapes from Bordeaux Vineyard for the 2009-Vintage: Comparison to 2006, 2007 and 2008 Vintages. Food Chem. 2011, 126, 1991–1999. [Google Scholar] [CrossRef]
- Chira, K.; Schmauch, G.; Saucier, C.; Fabre, S.; Teissedre, P.-L. Grape Variety Effect on Proanthocyanidin Composition and Sensory Perception of Skin and Seed Tannin Extracts from Bordeaux Wine Grapes (Cabernet Sauvignon and Merlot) for Two Consecutive Vintages (2006 and 2007). J. Agric. Food Chem. 2009, 57, 545–553. [Google Scholar] [CrossRef]
- Downey, M.O.; Harvey, J.S.; Robinson, S.P. Analysis of Tannins in Seeds and Skins of Shiraz Grapes throughout Berry Development. Aust. J. Grape Wine Res. 2003, 9, 15–27. [Google Scholar] [CrossRef]
- Ky, I.; Lorrain, B.; Kolbas, N.; Crozier, A.; Teissedre, P.-L. Wine By-Products: Phenolic Characterization and Antioxidant Activity Evaluation of Grapes and Grape Pomaces from Six Different French Grape Varieties. Molecules 2014, 19, 482–506. [Google Scholar] [CrossRef]
- Rousserie, P.; Rabot, A.; Geny-Denis, L. From Flavanols Biosynthesis to Wine Tannins: What Place for Grape Seeds? J. Agric. Food Chem. 2019, 67, 1325–1343. [Google Scholar] [CrossRef]
- Heras-Roger, J.; Alonso-Alonso, O.; Gallo-Montesdeoca, A.; Díaz-Romero, C.; Darias-Martín, J. Influence of Copigmentation and Phenolic Composition on Wine Color. J. Food Sci. Technol. 2016, 53, 2540–2547. [Google Scholar] [CrossRef]
- Ferrer-Gallego, R.; Brás, N.F.; García-Estévez, I.; Mateus, N.; Rivas-Gonzalo, J.C.; de Freitas, V.; Escribano-Bailón, M.T. Effect of Flavonols on Wine Astringency and Their Interaction with Human Saliva. Food Chem. 2016, 209, 358–364. [Google Scholar] [CrossRef] [PubMed]
- Castillo-Muñoz, N.; Gómez-Alonso, S.; García-Romero, E.; Hermosín-Gutiérrez, I. Flavonol Profiles of Vitis Vinifera Red Grapes and Their Single-Cultivar Wines. J. Agric. Food Chem. 2007, 55, 992–1002. [Google Scholar] [CrossRef] [PubMed]
- Németh, G.; Hegyi, O.; Dunai, A.; Kocsis, L. Stilbenes in the Different Organs of Vitis Vinifera Cv. Merlot Grafted on Teleki Kober 5BB Rootstock. Oeno One 2017, 51, 323–328. [Google Scholar] [CrossRef]
- Gatto, P.; Vrhovsek, U.; Muth, J.; Segala, C.; Romualdi, C.; Fontana, P.; Pruefer, D.; Stefanini, M.; Moser, C.; Mattivi, F.; et al. Ripening and Genotype Control Stilbene Accumulation in Healthy Grapes. J. Agric. Food Chem. 2008, 56, 11773–11785. [Google Scholar] [CrossRef] [PubMed]
- Dudoit, A.; Benbouguerra, N.; Richard, T.; Hornedo-Ortega, R.; Valls-Fonayet, J.; Coussot, G.; Saucier, C. α-Glucosidase Inhibitory Activity of Tannat Grape Phenolic Extracts in Relation to Their Ripening Stages. Biomolecules 2020, 10, 1088. [Google Scholar] [CrossRef] [PubMed]
- Burns, J.; Mullen, W.; Landrault, N.; Teissedre, P.-L.; Lean, M.E.J.; Crozier, A. Variations in the Profile and Content of Anthocyanins in Wines Made from Cabernet Sauvignon and Hybrid Grapes. J. Agric. Food Chem. 2002, 50, 4096–4102. [Google Scholar] [CrossRef]
- MacMicking, J.; Xie, Q.W.; Nathan, C. Nitric Oxide and Macrophage Function. Annu. Rev. Immunol. 1997, 15, 323–350. [Google Scholar] [CrossRef]
- Cíz, M.; Pavelková, M.; Gallová, L.; Králová, J.; Kubala, L.; Lojek, A. The Influence of Wine Polyphenols on Reactive Oxygen and Nitrogen Species Production by Murine Macrophages RAW 264.7. Physiol. Res. 2008, 57, 393–402. [Google Scholar] [PubMed]
- Park, Y.C.; Rimbach, G.; Saliou, C.; Valacchi, G.; Packer, L. Activity of Monomeric, Dimeric, and Trimeric Flavonoids on NO Production, TNF-α Secretion, and NF-ΚB-Dependent Gene Expression in RAW 264.7 Macrophages. FEBS Lett. 2000, 465, 93–97. [Google Scholar] [CrossRef]
- Bognar, E.; Sarszegi, Z.; Szabo, A.; Debreceni, B.; Kalman, N.; Tucsek, Z.; Sumegi, B.; Gallyas, F. Antioxidant and Anti-Inflammatory Effects in RAW264.7 Macrophages of Malvidin, a Major Red Wine Polyphenol. PLoS ONE 2013, 8. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Grapes. | Merlot | Tannat | Syrah | |||
|---|---|---|---|---|---|---|
| °Brix | pH | °Brix | pH | °Brix | pH | |
| BV | 4.37 | 2.56 | 4.3 | 2.57 | 4.37 | 2.58 |
| V | 8.57 | 3.34 | 11.64 | 2.75 | 16.97 | 3.42 |
| M | 25.97 | 3.78 | 25.84 | 3.87 | 25.37 | 4 |
| TPC (g GAE/kg of Skins FW) * | |||
| Skins | Merlot | Tannat | Syrah |
| BV | 18.5 ± 2.6 a | 19.2 ± 7.9 a | 18.3 ± 1.2 a |
| V | 17.0 ± 2.1 b | 16.7 ± 2.9 b | 16.9 ± 2.5 b |
| M | 13.1 ± 2.6 c | 16.5 ± 1.7 b | 16.2 ± 0.8 b |
| DPPH (g TE/kg of skins FW) ** | |||
| BV | 4.6 ± 0.4 b | 6.0 ± 0.6 b | 6.5 ± 0.4 b |
| V | 3.8 ± 1.3 c | 4.1 ± 0.9 ab | 5.9 ± 1.1 b |
| M | 3.3 ± 0.1 a | 3.9 ± 0.9 a | 2.9 ± 0.4 a |
| Merlot | Tannat | Syrah | |||||||
|---|---|---|---|---|---|---|---|---|---|
| BV | V | M | BV | V | M | BV | V | M | |
| Flavan-3-ols/procyanidins * | |||||||||
| (+)-Catechin | 107.04 ± 1.48 | 53.36 ± 0.51 | 23.95 ± 0.95 | 114.45 ± 4.66 | 90.11 ± 5.18 | 29.43 ± 6.56 | 97.72 ± 16.66 | 111.11 ± 0.02 | 39.82 ± 5.84 |
| (−)-Epicatechin | 1.38 ± 0.44 | 5.30 ± 0.37 | 5.90 ± 2.47 | 2.32 ± 0.78 | 9.85 ± 0.96 | 7.31 ± 1.50 | 0.86 ± 0.08 | 30.94 ± 5.54 | 8.75 ± 0.30 |
| Procyanidin B1 | 75.53 ± 3.90 | 128.23 ± 3.56 | 71.43 ± 2.14 | 50.77 ± 0.39 | 108.18 ± 5.16 | 105.36 ± 6.09 | 52.66 ± 4.24 | 138.63 ± 8.90 | 89.92 ± 9.03 |
| Procyanidin B2 | 2.09 ± 0.36 | 3.83 ± 0.50 | 9.21 ± 0.76 | 3.07 ± 0.11 | 4.27 ± 0.03 | 4.50 ± 0.17 | 1.16 ± 0.10 | 6.82 ± 2.44 | 9.57 ± 0.20 |
| Procyanidin B3 | 29.45 ± 2.48 | 22.99 ± 3.85 | 12.04 ± 1.74 | 17.35 ± 0.04 | 24.27 ± 2.34 | 6.99 ± 1.71 | 26.14 ± 2.95 | 30.26 ± 0.64 | 16.51 ± 3.56 |
| Procyanidin B4 | 5.61 ± 0.33 | 5.14 ± 0.62 | 7.42 ± 2.07 | 7.35 ± 0.57 | 8.22 ± 0.87 | 4.16 ± 1.04 | 6.42 ± 1.17 | 7.78 ± 1.92 | 6.73 ± 1.20 |
| Flavonols * | |||||||||
| Quercetin | 0.13 ± 0.04 | 0.33 ± 0.06 | 0.28 ± 0.16 | 0.08 ± 0.04 | 0.25 ± 0.06 | 0.34 ± 0.09 | 0.07 ± 0.00 | 2.02 ± 0.30 | 1.03 ± 0.45 |
| Quercetin-3-O-hexoside | 2.78 ± 0.49 | 28.35 ± 5.83 | 38.07 ± 3.01 | 3.21 ± 0.65 | 6.09 ± 2.28 | 8.56 ± 0.11 | 2.61 ± 1.03 | 75.88 ± 1.34 | 148.13 ± 25.29 |
| Quercetin-3-O-galactoside | 0.62 ± 0.08 | 4.51 ± 1.06 | 3.99 ± 0.92 | 1.04 ± 0.10 | 0.87 ± 0.13 | 0.75 ± 0.05 | 0.30 ± 0.04 | 7.26 ± 0.66 | 9.66 ± 1.79 |
| Quercetin-3-O-glucuronide | 75.14 ± 9.01 | 154.63 ± 15.09 | 122.17 ± 6.80 | 98.93 ± 13.71 | 44.78 ± 4.22 | 58.10 ± 21.21 | 63.90 ± 0.75 | 227.90 ± 18.01 | 117.70 ± 12.62 |
| Quercetin-3-O-rhamnoside | 0.03 ± 0.02 | 1.06 ± 0.43 | 0.45 ± 0.18 | 0.01 ± 0.00 | 0.02 ± 0.00 | 0.02 ± 0.00 | 0.02 ± 0.00 | 1.16 ± 0.12 | 1.10 ± 0.84 |
| Quercetin-3-O-rutinoside | 1.33 ± 0.23 | 1.75 ± 0.11 | 0.49 ± 0.05 | 5.46 ± 0.78 | 3.34 ± 0.28 | 0.80 ± 0.46 | 0.38 ± 0.21 | 0.58 ± 0.50 | 1.22 ± 1.10 |
| Kaempferol-3-O-glucoside | 0.29 ± 0.10 | 6.78 ± 1.75 | 4.28 ± 0.10 | 0.40 ± 0.15 | 1.67 ± 0.52 | 0.32 ± 0.12 | 0.08 ± 0.05 | 3.87 ± 1.38 | 9.06 ± 2.37 |
| Stilbenes * | |||||||||
| Cis-resveratrol | 0.03 ± 0.02 | 0.04 ± 0.01 | 0.14 ± 0.02 | 0.03 ± 0.01 | 0.45 ± 0.26 | 0.09 ± 0.02 | 0.03 ± 0.00 | 1.94 ± 0.05 | 1.43 ± 0.65 |
| Trans-resveratrol | 0.11 ± 0.04 | 0.13 ± 0.02 | 0.25 ± 0.11 | 0.04 ± 0.01 | 0.44 ± 0.38 | 0.17 ± 0.10 | 0.13 ± 0.02 | 4.07 ± 1.21 | 5.34 ± 0.18 |
| ∑ resveratrol | 0.15 | 0.17 | 0.39 | 0.07 | 0.89 | 0.26 | 0.16 | 6.01 | 6.77 |
| Cis-piceid | 0.30 ± 0.06 | 1.39 ± 0.44 | 9.63 ± 0.93 | 1.04 ± 0.48 | 2.36 ± 1.31 | 3.00 ± 1.12 | 0.73 ± 0.21 | 15.74 ± 6.84 | 41.99 ± 1.41 |
| Trans-piceid | 0.10 ± 0.02 | 0.36 ± 0.01 | 1.93 ± 0.37 | 0.68 ± 0.32 | 0.52 ± 0.25 | 1.41 ± 0.28 | 0.20 ± 0.08 | 2.12 ± 1.14 | 7.65 ± 1.53 |
| ∑ piceid | 0.39 | 1.75 | 11.56 | 1.72 | 2.88 | 4.41 | 0.93 | 17.86 | 49.64 |
| Cis-piceatannol | 0.03 ± 0.01 | 0.38 ± 0.19 | 0.26 ± 0.07 | 0.05 ± 0.02 | 0.03 ± 0.00 | 0.09 ± 0.03 | 0.03 ± 0.00 | 9.18 ± 1.52 | 1.52 ± 0.53 |
| Trans-piceatannol | 0.02 ± 0.01 | 0.23 ± 0.11 | 1.38 ± 0.80 | 0.02 ± 0.01 | 0.02 ± 0.01 | 0.32 ± 0.15 | 0.02 ± 0.00 | 5.48 ± 0.91 | 10.49 ± 1.93 |
| ∑ piceatannol | 0.04 | 0.61 | 1.64 | 0.06 | 0.23 | 0.41 | 0.05 | 14.66 | 12.01 |
| Cis-astringin | 0.03 ± 0.00 | 0.24 ± 0.00 | 2.93 ± 0.49 | 0.04 ± 0.02 | 0.13 ± 0.08 | 0.28 ± 0.01 | 0.04 ± 0.02 | 0.83 ± 0.33 | 3.77 ± 0.98 |
| Trans-astringin | 0.04 ± 0.01 | 0.72 ± 0.10 | 5.91 ± 1.14 | 0.09 ± 0.06 | 0.29 ± 0.16 | 1.70 ± 0.81 | 0.04 ± 0.02 | 1.97 ± 0.33 | 13.27 ± 1.03 |
| ∑ astringin | 0.07 | 0.96 | 8.84 | 0.13 | 0.42 | 1.98 | 0.08 | 2.79 | 17.05 |
| ε-viniferin | 0.73 ± 0.19 | 0.93 ± 0.37 | 1.61 ± 0.26 | 0.12 ± 0.04 | 1.70 ± 0.70 | 0.63 ± 0.16 | 0.27 ± 0.11 | 1.38 ± 0.37 | 4.77 ± 0.21 |
| ω-viniferin | 0.03 ± 0.01 | 0.04 ± 0.01 | 0.10 ± 0.04 | 0.01 ± 0.000 | 0.07 ± 0.03 | 0.06 ± 0.01 | 0.01 ± 0.00 | 0.27 ± 0.05 | 0.95 ± 0.04 |
| σ-viniferin | 0.01 ± 0.00 | 0.01 ± 0.00 | 0.95 ± 0.14 | 0.01 ± 0.01 | 0.07 ± 0.02 | 0.08 ± 0.02 | 0.01 ± 0.00 | 0.84 ± 0.13 | 2.97 ± 0.70 |
| Pallidol | 0.02 ± 0.01 | 0.03 ± 0.01 | 1.88 ± 1.11 | 0.03 ± 0.01 | 0.48 ± 0.30 | 0.28 ± 0.05 | 0.05 ± 0.02 | 1.06 ± 0.46 | 6.73 ± 2.57 |
| Parthenocissin A | 0.06 ± 0.04 | 0.08 ± 0.07 | 2.37 ± 1.11 | 0.06 ± 0.04 | 0.53 ± 0.37 | 0.39 ± 0.14 | 0.05 ± 0.02 | 1.47 ± 0.18 | 4.81 ± 0.91 |
| Miyabenol C | 0.06 ± 0.01 | 0.01 ± 0.00 | 0.22 ± 0.02 | 0.08 ± 0.06 | 0.27 ± 0.15 | 0.16 ± 0.07 | 0.34 ± 0.02 | 0.57 ± 0.09 | 3.05 ± 0.85 |
| Hopeaphenol | 0.03 ± 0.01 | 0.06 ± 0.01 | 0.26 ± 0.04 | 0.01 ± 0.00 | 0.02 ± 0.00 | 0.03 ± 0.01 | 0.02 ± 0.00 | 0.05 ± 0.01 | 0.22 ± 0.08 |
| Isohopeaphenol | 0.03 ± 0.00 | 0.04 ± 0.01 | 0.44 ± 0.19 | 0.04 ± 0.01 | 0.04 ± 0.00 | 0.61 ± 0.07 | 0.03 ± 0.01 | 1.07 ± 0.31 | 2.95 ± 0.22 |
| Merlot | Tannat | Syrah | |||||||
|---|---|---|---|---|---|---|---|---|---|
| BV | V | M | BV | V | M | BV | V | M | |
| Anthocyanins | |||||||||
| Delphinidin-3-O-glucoside | nd | 269.52 ± 18.26 | 352.42 ± 27.39 | nd | 145.86 ± 72.87 | 395.07 ± 134.98 | nd | 526.08 ± 204.63 | 396.80 ± 151.81 |
| Cyanidin-3-O-glucoside | nd | 265.65 ± 64.55 | 346.87 ± 26.86 | nd | 100.56 ± 54.84 | 156.56 ± 41.43 | nd | 195.24 ± 105.11 | 143.69 ± 23.56 |
| Petunidin-3-O-glucoside | nd | 86.89 ± 69.48 | 348.83 ± 17.31 | nd | 218.86 ± 11.05 | 624.99 ± 136.52 | nd | 1090.05 ± 660.16 | 543.70 ± 184.11 |
| Peonidin-3-O-glucoside | nd | 801.77 ± 173.48 | 1185.78 ± 94.42 | nd | 176.48 ± 117.35 | 324.17 ± 33.72 | nd | 742.67 ± 312.21 | 709.13 ± 109.47 |
| Malvidin-3-O-glucoside | nd | 993.74 ± 238.54 | 1378.45 ± 69.64 | nd | 635.40 ± 389.99 | 1948.62 ± 204.63 | nd | 2362.31 ± 339.03 | 2321.65 ± 682.10 |
| ∑ 3-O-glucosides | - | 2617.58 | 3612.86 | - | 1277.18 | 3449.42 | - | 4916.36 | 4114.97 |
| Petunidin-3-O-(6-O-acetyl)-glucoside | nd | 107.48 + 19.38 | 125.87 + 3.82 | nd | 58.61 + 24.12 | 160.74 + 42.68 | nd | 185.95 + 37.20 | 172.08 + 34.93 |
| Peonidin-3-O-(6-O-acetyl)-glucoside | nd | 200.81 ± 32.21 | 300.34 ± 36.02 | nd | 57.46 ± 20.84 | 94.67 ± 12.26 | nd | 279.77 ± 60.58 | 287.06 ± 18.66 |
| Malvidin-3-O-(6-O-acetyl)-glucoside | nd | 385.37 ± 78.51 | 572.86 ± 14.73 | nd | 123.42 ± 67.78 | 427.06 ± 60.19 | nd | 785.32 ± 70.90 | 978.46 ± 203.50 |
| ∑ 3-O-acetylglucosides | - | 693.66 | 999.07 | - | 239.49 | 682.46 | - | 1251.04 | 1437.6 |
| Delphinidin-3-O-(6-O-p-coumaroyl)-glucoside | nd | 83.91 + 15.15 | 134.52 + 8.02 | nd | 53.52 + 20.15 | 92.14 + 9.06 | nd | 151.28 + 22.97 | 164.11 + 8.47 |
| Cyanidin-3-O-(6-O-p-coumaroyl)-glucoside | nd | 105.93 + 15.5 | 126 + 7.63 | nd | 66.31 + 26.66 | 160.24 + 24.22 | nd | 237.91 + 16.67 | 226.65 + 66.26 |
| Peonidin-3-O-(6-O-p-coumaroyl)-glucoside | nd | 153.37 ± 22.60 | 250.27 ± 31.35 | nd | 59.60 ± 24.76 | 118.29 ± 10.51 | nd | 444.01 ± 83.32 | 451.89 ± 70.38 |
| Malvidin-3-O-(6-O-p-coumaroyl)-glucoside | nd | 342.07 ± 59.97 | 585.13 ± 48.62 | nd | 143.01 ± 80.71 | 470.23 ± 63.55 | nd | 905.34 ± 126.90 | 1183.21 ± 385.23 |
| ∑ 3-O-coumaroylglucosides | - | 685.29 | 1095.91 | - | 322.44 | 840.9 | - | 1738.19 | 2025.86 |
| mDP | |||
|---|---|---|---|
| Skins | Merlot | Tannat | Syrah |
| BV | 18.05 ± 0.07 | 13.88 ± 0.48 | 18.11 ± 0.40 |
| V | 32.21 ± 0.94 | 15.52 ± 0.01 | 17.62 ± 0.07 |
| M | 25.00 ± 0.13 | 16.08 ± 0.44 | 25.82 ± 0.90 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Benbouguerra, N.; Valls-Fonayet, J.; Krisa, S.; Garcia, F.; Saucier, C.; Richard, T.; Hornedo-Ortega, R. Polyphenolic Characterization of Merlot, Tannat and Syrah Skin Extracts at Different Degrees of Maturity and Anti-Inflammatory Potential in RAW 264.7 Cells. Foods 2021, 10, 541. https://doi.org/10.3390/foods10030541
Benbouguerra N, Valls-Fonayet J, Krisa S, Garcia F, Saucier C, Richard T, Hornedo-Ortega R. Polyphenolic Characterization of Merlot, Tannat and Syrah Skin Extracts at Different Degrees of Maturity and Anti-Inflammatory Potential in RAW 264.7 Cells. Foods. 2021; 10(3):541. https://doi.org/10.3390/foods10030541
Chicago/Turabian StyleBenbouguerra, Nawel, Josep Valls-Fonayet, Stephanie Krisa, François Garcia, Cédric Saucier, Tristan Richard, and Ruth Hornedo-Ortega. 2021. "Polyphenolic Characterization of Merlot, Tannat and Syrah Skin Extracts at Different Degrees of Maturity and Anti-Inflammatory Potential in RAW 264.7 Cells" Foods 10, no. 3: 541. https://doi.org/10.3390/foods10030541
APA StyleBenbouguerra, N., Valls-Fonayet, J., Krisa, S., Garcia, F., Saucier, C., Richard, T., & Hornedo-Ortega, R. (2021). Polyphenolic Characterization of Merlot, Tannat and Syrah Skin Extracts at Different Degrees of Maturity and Anti-Inflammatory Potential in RAW 264.7 Cells. Foods, 10(3), 541. https://doi.org/10.3390/foods10030541