
Modulation of Gut Microbiota by Lactobacillus casei Fermented Raspberry Juice In Vitro and In Vivo


Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Preparation and Fermentation of Raspberry Juice
2.3. Determination of Antioxidant Activity
2.3.1. The 2,2′-Azino-bis (3-ethylbenzothiazoline-6-sulfonic acid) Radical Cation Scavenging Activity Assay (ABTS)
2.3.2. The 2,2-Diphenyl-1-picrylhydrazyl Radical Scavenging Activity (DPPH)
2.4. The Total Phenolic Content (TPC) Analysis
2.5. In Vitro Digestion and Colonic Fermentation
2.6. Determination of Phenolic Compounds in FRJ
2.7. Animal Study
2.8. In Vivo Redox Status Analysis
2.9. DNA/RNA Extraction and qPCR
2.10. Analysis of 16S rRNA Illumina Sequencing
2.11. SCFAs Analysis
2.12. Statistical Analysis
3. Results
3.1. Growth of L. casei and pH of FRJ
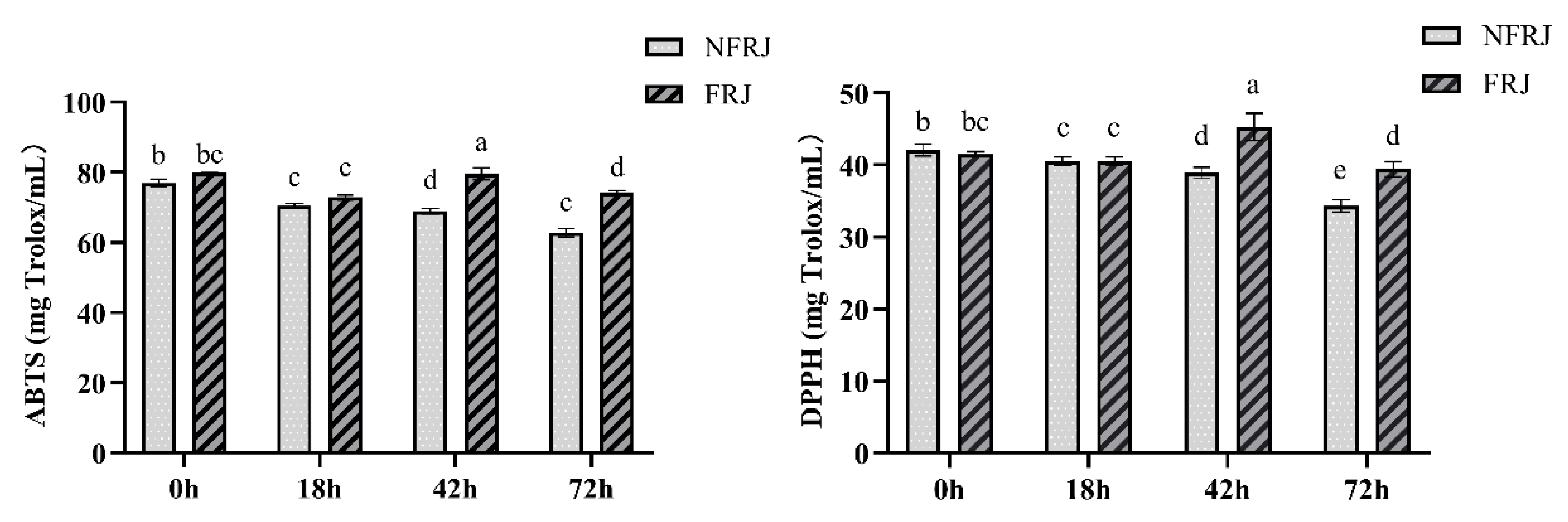
3.2. Antioxidant Activities of FRJ and NFRJ
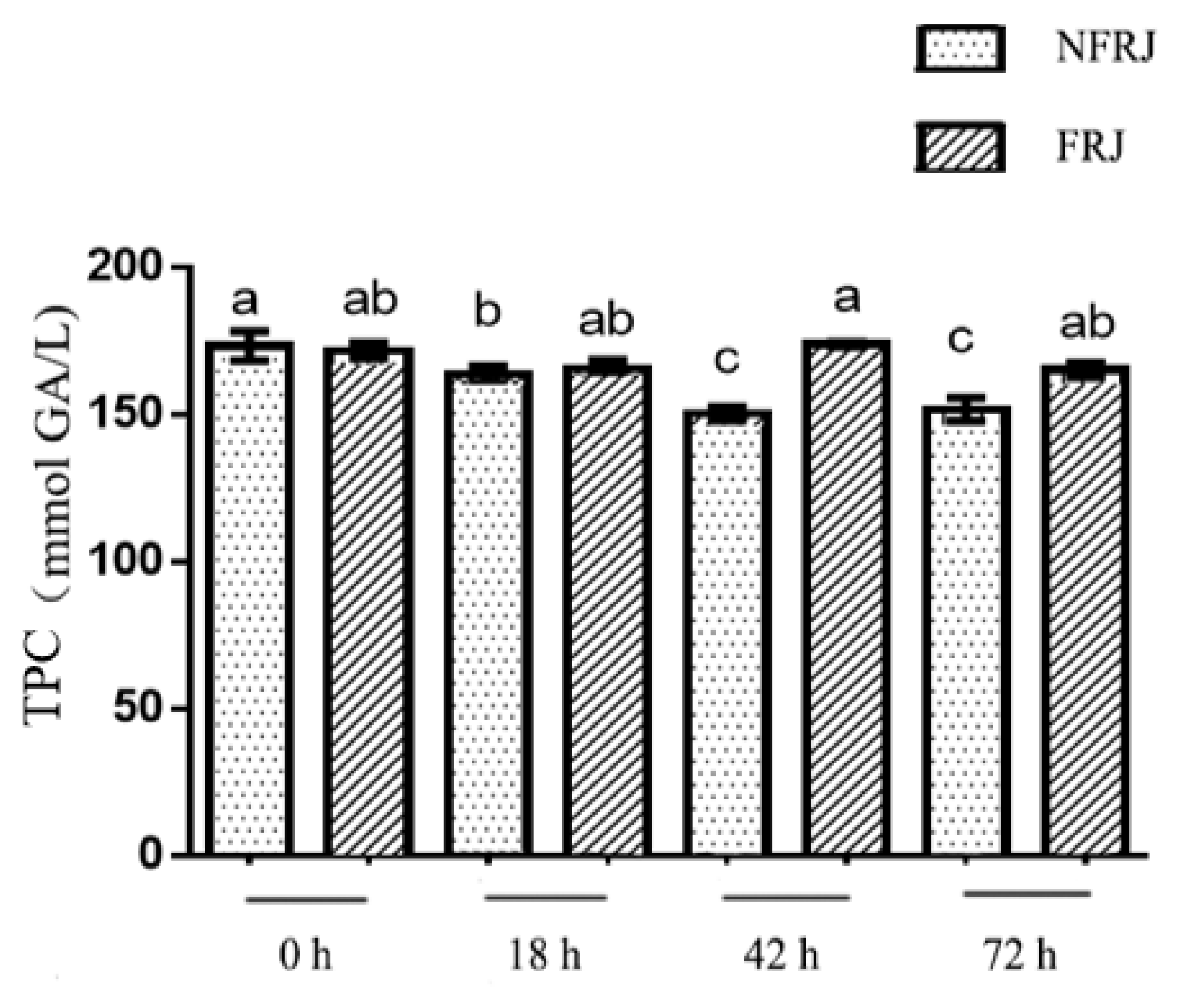
3.3. The Total Phenolic Content (TPC) of FRJ and NFRJ
3.4. Determination of Phenolic Compounds in FRJ
3.5. Effects of FRJ on Fecal Microbiota In Vitro
3.6. Effects of FRJ and NFRJ on In Vitro Production of Short Chain Fatty Acids (SCFAs)
3.7. Effects of FRJ on Growth Performance of Mice
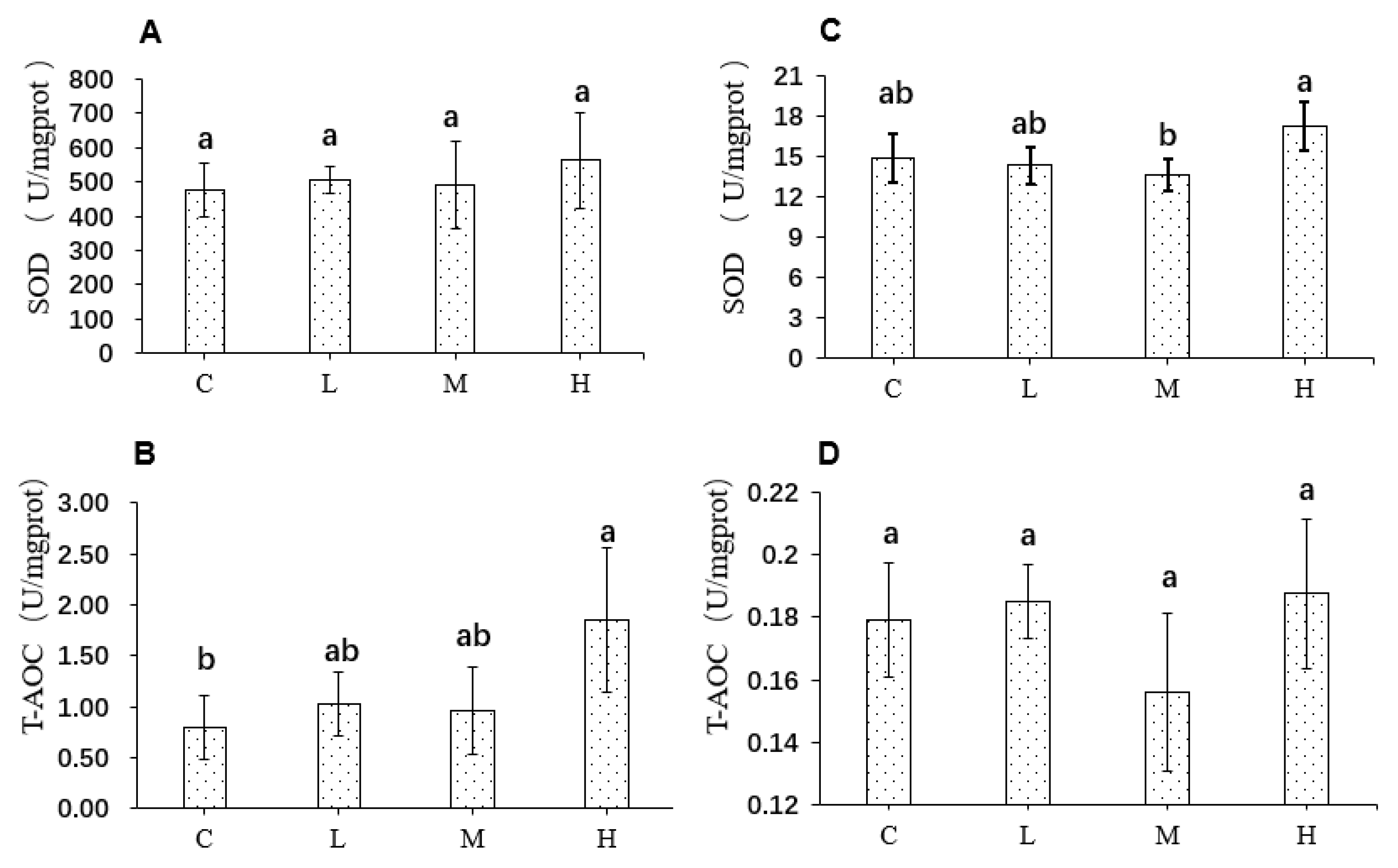
3.8. Effects of FRJ on Redox Status in the Liver and Colon of Mice
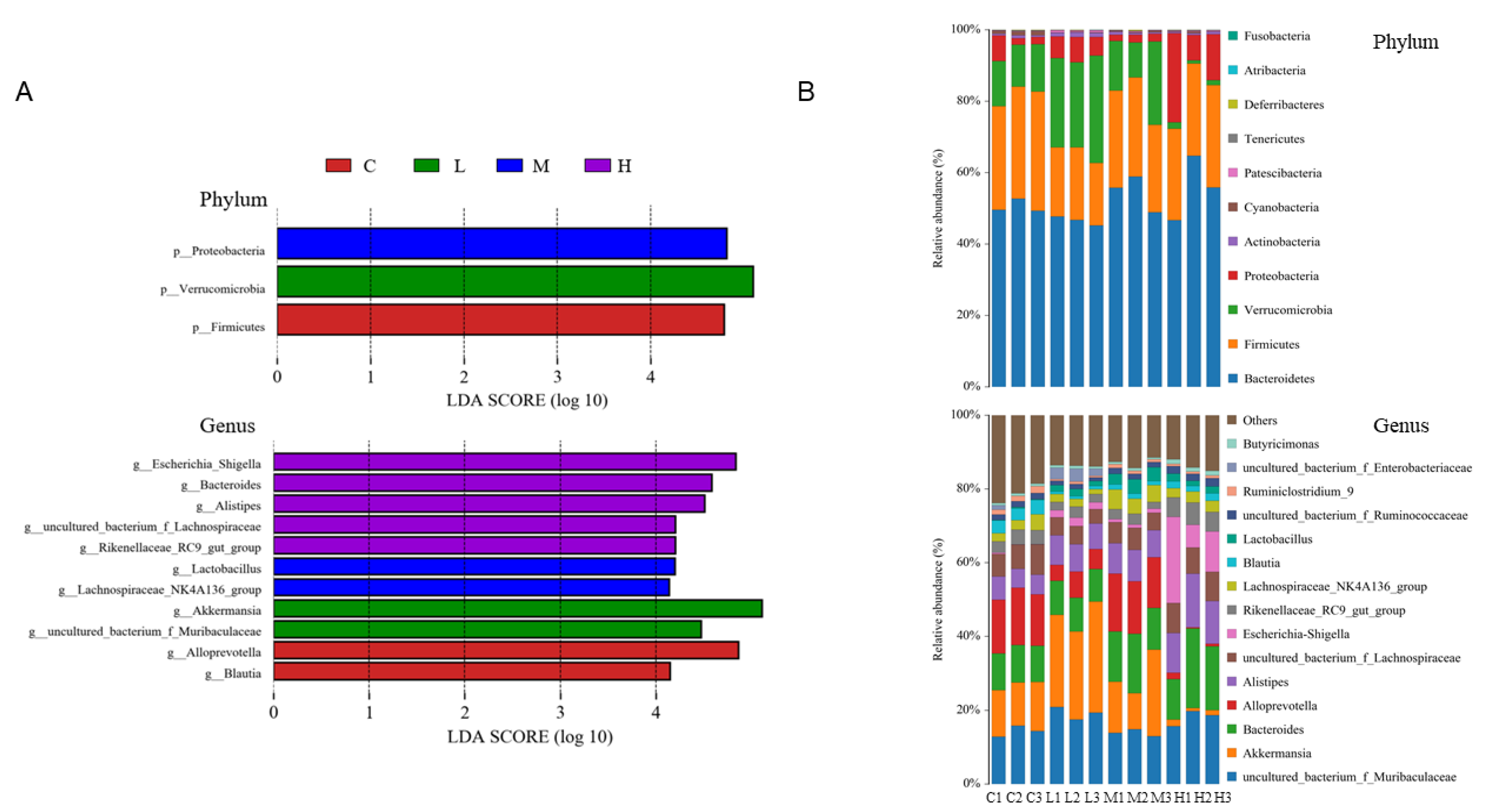
3.9. Effects of FRJ on Fecal Microbiota In Vivo
3.10. Effects of FRJ on Production of SCFAs in Mice
3.11. Effects of FRJ on the Gene Expression of Colon Mechanical Barrier
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Yang, J.; Cui, J.; Chen, J.; Yao, J.; Hao, Y.; Fan, Y.; Liu, Y. Evaluation of physicochemical properties in three raspberries (Rubus idaeus) at five ripening stages in northern China. Sci. Hortic. 2020, 263, 109146. [Google Scholar] [CrossRef]
- Borges, G.; Degeneve, A.; Mullen, W.; Crozier, A. Identification of Flavonoid and Phenolic Antioxidants in Black Currants, Blueberries, Raspberries, Red Currants, and Cranberries. J. Agric. Food Chem. 2010, 58, 3901–3909. [Google Scholar] [CrossRef]
- García-Niño, W.R.; Zazueta, C. Ellagic acid: Pharmacological activities and molecular mechanisms involved in liver protection. Pharmacol. Res. 2015, 97, 84–103. [Google Scholar] [CrossRef]
- Gomes-Neto, J.C.; Mantz, S.; Held, K.; Sinha, R.; Munoz, R.R.S.; Schmaltz, R.; Benson, A.K.; Walter, J.; Ramer-Tait, A.E. A real-time PCR assay for accurate quantification of the individual members of the Altered Schaedler Flora microbiota in gnotobiotic mice. J. Microbiol. Methods 2017, 135, 52–62. [Google Scholar] [CrossRef] [Green Version]
- Siracusa, L.; Giuseppe, R. Not Only What Is Food Is Good—Polyphenols From Edible and Nonedible Vegetable Waste- ScienceDirect. Polyphen. Plants 2019, 3–21. [Google Scholar] [CrossRef]
- Piccolo, E.L.; Garcìa, L.M.; Landi, M.; Guidi, L.; Remorini, D. Influences of Postharvest Storage and Processing Techniques on Antioxidant and Nutraceutical Properties of Rubus idaeus L.: A Mini-Review. Horticulturae 2020, 6, 105. [Google Scholar] [CrossRef]
- Panghal, A.; Janghu, S.; Virkar, K.; Gat, Y.; Kumar, V.; Chhikara, N. Potential non-dairy probiotic products–A healthy approach. Food Biosci. 2018, 21, 80–89. [Google Scholar] [CrossRef]
- Bancalari, E.; Castellone, V.; Bottari, B.; Gatti, M. Wild Lactobacillus casei Group Strains: Potentiality to Ferment Plant Derived Juices. Foods 2020, 9, 314. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.Y.; Lee, M.Y.; Ji, G.E.; Lee, Y.S.; Hwang, K.T. Production of γ-aminobutyric acid in black raspberry juice during fermentation by Lactobacillus brevis GABA100. Int. J. Food Microbiol. 2009, 130, 12–16. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Zhang, M.; Wang, W.; Lv, H.; Ta, N.Z. The in vitro Effects of the Probiotic Strain, Lactobacillus casei ZX633 on Gut Microbiota Composition in Infants With Diarrhea. Front. Cell. Infect. Microbiol. 2020, 10, 576185. [Google Scholar] [CrossRef]
- Flint, H.J.; Duncan, S.H.; Scott, K.P.; Louis, P. Links between diet, gut microbiota composition and gut metabolism. Proc. Nutr. Soc. 2015, 74, 13–22. [Google Scholar] [CrossRef] [Green Version]
- Lavefve, L.; Howard, L.R.; Carbonero, F. Berry polyphenols metabolism and impact on human gut microbiota and health. Food Funct. 2020, 11, 45–65. [Google Scholar] [CrossRef]
- Cheng, Y.; Wu, T.; Chu, X.; Tang, S.; Xu, X. Fermented blueberry pomace with antioxidant properties improves fecal microbiota community structure and short chain fatty acids production in an in vitro mode. LWT 2020, 125, 109260. [Google Scholar] [CrossRef]
- Kwaw, E.; Ma, Y.; Tchabo, W.; Apaliya, M.T.; Wu, M.; Sackey, A.S.; Xiao, L.; Tahir, H.E. Effect of lactobacillus strains on phenolic profile, color attributes and antioxidant activities of lactic-acid-fermented mulberry juice. Food Chem. 2018, 250, 148–154. [Google Scholar] [CrossRef]
- Ramirez, J.E.; Zambrano, R.; Sepulveda, B.; Kennelly, E.J.; Simirgiotis, M.J. Anthocyanins and antioxidant capacities of six Chilean berries by HPLC–HR-ESI-ToF-MS. Food Chem. 2015, 176, 106–114. [Google Scholar] [CrossRef] [PubMed]
- Koehnlein, E.A.; Koehnlein, É.M.; Corrêa, R.; Nishida, V.S.; Correa, V.G.; Bracht, A.; Peralta, R.M. Analysis of a whole diet in terms of phenolic content and antioxidant capacity: Effects of a simulated gastrointestinal digestion. Int. J. Food Sci. Nutr. 2016, 67, 614–623. [Google Scholar] [CrossRef]
- Corrêa, R.; Haminiuk, C.; Barros, L.; Dias, M.I.; Calhelha, R.C.; Kato, C.G.; Correa, V.G.; Peralta, R.M.; Ferreira, I. Stability and biological activity of Merlot (Vitis vinifera) grape pomace phytochemicals after simulated in vitro gastrointestinal digestion and colonic fermentation. J. Funct. Foods 2017, 36, 410–417. [Google Scholar] [CrossRef] [Green Version]
- Tang, S.; Cheng, Y.; Wu, T.; Hu, F.; Pan, S.; Xu, X. Effect of Lactobacillus plantarum-fermented mulberry pomace on antioxidant properties and fecal microbial community. LWT 2021, 147, 111651. [Google Scholar] [CrossRef]
- Cheng, Y.; Wu, T.; Tang, S.; Liang, F.; Fang, Y.; Cao, W.; Pan, S.; Xu, X. Fermented blueberry pomace ameliorates intestinal barrier function through the NF-κB-MLCK signaling pathway in high-fat diet mice. Food Funct. 2020, 11, 3167–3179. [Google Scholar] [CrossRef]
- Kong, C.; Gao, R.; Yan, X.; Huang, L.; Qin, H. Probiotics improve gut microbiota dysbiosis in obese mice fed a high-fat or high-sucrose diet. Nutrition 2019, 60, 175–184. [Google Scholar] [CrossRef] [PubMed]
- Mousavi, Z.E.; Mousavi, S.M.; Razavi, S.H.; Emam-Djomeh, Z.; Kiani, H. Fermentation of pomegranate juice by probiotic lactic acid bacteria. World J. Microbiol. Biotechnol. 2011, 27, 123–128. [Google Scholar] [CrossRef]
- Cirlini, M.; Ricci, A.; Galaverna, G.; Lazzi, C. Application of lactic acid fermentation to elderberry juice: Changes in acidic and glucidic fractions. LWT 2019, 118, 108779. [Google Scholar] [CrossRef]
- Clark, C.J.; Cooney, J.M.; Hopkins, W.A.; Currie, A. Global Mid-Infrared Prediction Models Facilitate Simultaneous Analysis of Juice Composition from Berries of Actinidia, Ribes, Rubus and Vaccinium Species. Food Anal. Methods 2018, 11, 3147–3160. [Google Scholar] [CrossRef]
- Ryu, J.; Kang, H.R.; Cho, S.K. Changes Over the Fermentation Period in Phenolic Compounds and Antioxidant and Anticancer Activities of Blueberries Fermented by Lactobacillus plantarum. J. Food Sci. 2019, 84, 2347–2356. [Google Scholar] [CrossRef]
- Filannino, P.; Bai, Y.; Cagno, R.D.; Gobbetti, M.; Gaenzle, M.G. Metabolism of phenolic compounds by Lactobacillus spp. during fermentation of cherry juice and broccoli puree. Food Microbiol. 2015, 46, 272–279. [Google Scholar] [CrossRef]
- Chait, Y.A.; Gunenc, A.; Hosseinian, F.; Bendali, F. Antipathogenic and probiotic potential of Lactobacillus brevis strains newly isolated from Algerian artisanal cheeses. Folia Microbiol. 2021, 66, 429–440. [Google Scholar] [CrossRef]
- Yang, S.-J.; Lee, J.-E.; Lim, S.-M.; Kim, Y.-J.; Lee, N.-K.; Paik, H.-D. Antioxidant and immune-enhancing effects of probiotic Lactobacillus plantarum 200655 isolated from kimchi. Food Sci. Biotechnol. 2019, 28, 491–499. [Google Scholar] [CrossRef]
- Yxa, B.; Rong, F.C.; Jing, S.; Amt, C.; Scc, D.; Rta, B. Polyphenolic fractions isolated from red raspberry whole fruit, pulp, and seed differentially alter the gut microbiota of mice with diet-induced obesity -ScienceDirect. J. Funct. Foods 2020, 76, 104288. [Google Scholar]
- Reid, G.; Abrahamsson, T.; Bailey, M.; Bindels, L.B.; Bubnov, R.; Ganguli, K.; Martoni, C.; O’Neill, C.; Savignac, H.M.; Stanton, C.; et al. How do probiotics and prebiotics function at distant sites? Benef. Microbes 2017, 8, 521–533. [Google Scholar] [CrossRef]
- Linares, D.M.; Carolina, G.; Erica, R.; Fresno, J.M.; Tornadijo, M.E.; Ross, R.P.; Catherine, S. Lactic Acid Bacteria and Bifidobacteria with Potential to Design Natural Biofunctional Health-Promoting Dairy Foods. Front. Microbiol. 2017, 8, 846. [Google Scholar] [CrossRef] [PubMed]
- Jayachandran, M.; Chung, S.; Xu, B. A critical review of the relationship between dietary components, the gut microbe Akkermansia muciniphila, and human health. Crit. Rev. Food Sci. Nutr. 2019, 60, 1–12. [Google Scholar] [CrossRef]
- Xing, H.; Goedert, J.J.; Pu, A.; Yu, G.; Shi, J. Allergy associations with the adult fecal microbiota: Analysis of the American Gut Project. EBioMedicine 2016, 3, 172–179. [Google Scholar]
- Ren, X.; Zhu, Y.; Gamallat, Y.; Ma, S.; Chiwala, G.; Meyiah, A.; Xin, Y.E. Coli O124 K72 alters the intestinal barrier and the tight junctions proteins of guinea pig intestine. Biomed. Pharmacother. 2017, 94, 468–473. [Google Scholar] [CrossRef] [PubMed]
- Wen, J.; Ma, L.; Xu, Y.; Wu, J.; Li, L. Effects of probiotic litchi juice on immunomodulatory function and gut microbiota in mice. Food Res. Int. 2020, 137, 109433. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.V.; Frassetto, A.; Kowalik, E.J., Jr.; Nawrocki, A.R.; Lu, M.M.; Kosinski, J.R.; Hubert, J.A.; Szeto, D.; Yao, X.; Forrest, G.; et al. Butyrate and Propionate Protect against Diet-Induced Obesity and Regulate Gut Hormones via Free Fatty Acid Receptor 3-Independent Mechanisms. PLoS ONE 2012, 7, e35240. [Google Scholar] [CrossRef]
- Ríos-Covián, D.; Ruas-Madiedo, P.; Margolles, A.; Gueimonde, M.; Reyes-Gavilán, C.; Salazar, N. Intestinal Short Chain Fatty Acids and their Link with Diet and Human Health. Front. Microbiol. 2016, 7, 185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, M.G.; Fei, Y.Q.; Wang, Y.; Wang, W.Y.; Wang, Z. Chlorogenic Acid Alleviates Colon Mucosal Damage Induced by a High-Fat Diet via Gut Microflora Adjustment to Increase Short-Chain Fatty Acid Accumulation in Rats. Oxidative Med. Cell. Longev. 2021, 2021, 1–18. [Google Scholar] [CrossRef]
- Ley, R.E.; Backhed, F.; Turnbaugh, P.; Lozupone, C.A.; Knight, R.D.; Gordon, J.I. Obesity alters gut microbial ecology. Proc. Natl. Acad. Sci. USA 2005, 102, 11070–11075. [Google Scholar] [CrossRef] [Green Version]
- Shin, N.R.; Whon, T.W.; Bae, J.W. Proteobacteria: Microbial signature of dysbiosis in gut microbiota. Trends Biotechnol. 2015, 33, 496–503. [Google Scholar] [CrossRef] [PubMed]
- Tx, A.; Wd, A.; Zz, A.; Sl, A.; Yz, A.; Bg, A.; Yu, Z.A.; Jyb, C.; Min, W.A. Polyphenol-rich vinegar extract regulates intestinal microbiota and immunity and prevents alcohol-induced inflammation in mice. Food Res. Int. 2021, 140, 110064. [Google Scholar]
- Henning, S.M.; Summanen, P.H.; Lee, R.-P.; Yang, J.; Finegold, S.M.; Heber, D.; Li, Z. Pomegranate ellagitannins stimulate the growth of Akkermansia muciniphila in vivo. Anaerobe 2017, 43, 56–60. [Google Scholar] [CrossRef]
- Reunanen, J.; Kainulainen, V.; Huuskonen, L.; Ottman, N.; Belzer, C.; Huhtinen, H.; De Vos, W.M.; Satokari, R.; Goodrich-Blair, H. Akkermansia muciniphila Adheres to Enterocytes and Strengthens the Integrity of the Epithelial Cell Layer. Appl. Environ. Microbiol. 2015, 81, 3655–3662. [Google Scholar] [CrossRef] [Green Version]
- Othaim, A.A.; Marasini, D.; Carbonero, F. Impact of increasing concentration of tart and sweet cherries juices concentrates on healthy mice gut microbiota. Food Front. 2020, 1, 224–233. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Rt (min) | [M−H]+ | Error (ppm) | MS2 | [M−H]− | Error (ppm) | MS2 | Formula | Identification | Concentration (μg/g) |
|---|---|---|---|---|---|---|---|---|---|---|
| Anthocyanins | ||||||||||
| 1 | 5.49 | 449.10703 | −3.020 | 287 | C21H20O11 | cyanidin-3-O-glucoside | 298.34 ± 2.58 | |||
| 2 | 5.69 | 595.16431 | −3.335 | 287 | C27H30O15 | cyanidin-3-O-rutinoside | 10.06 ± 0.05 | |||
| 3 | 6.30 | 463.12244 | −3.447 | 301 | C22H22O11 | peonidin-3-O-glucoside/ peonidin-3-O-galactcoside | 2.20 ± 0.01 | |||
| 4 | 7.38 | 287.05429 | −4.434 | 287 | C15H10O6 | cyanidin | 58.22 ± 1.44 | |||
| 5 | 9.31 | 465.10172 | −3.399 | 303 | C21H20O12 | delphinidin-3-O-glucoside/ delphinidin-3-O-galactcoside | 19.11 ± 0.25 | |||
| Total Anthocyanins | 387.93 | |||||||||
| Flavanols | ||||||||||
| 6 | 6.07 | 289.07187 | 2.273 | C15H14O6 | catechin | 0.04 ± 0.00 | ||||
| 7 | 6.47 | 577.13519 | 1.021 | C30H26O12 | procyanidin B | 32.17 ± 1.03 | ||||
| Total Flavanols | 32.21 | |||||||||
| Flavonols | ||||||||||
| 8 | 5.50 | 447.09314 | 0.903 | 300 | C21H20O11 | quercetin-3-O-rhamnoside | 10.03 ± 0.15 | |||
| 9 | 9.30 | 463.08835 | 1.510 | 300 | C21H20O12 | quercetin-3-O-glucoside/ quercetin-3-O-galactcoside | 5.07 ± 0.04 | |||
| 10 | 13.06 | 301.03534 | 1.703 | C15H10O7 | quercetin | 16.42 ± 0.20 | ||||
| 11 | 14.97 | 285.04041 | 1.744 | C15H10O6 | kaempferol | 0.06 ± 0.00 | ||||
| Total Flavonols | 31.58 | |||||||||
| Phenolic acids | ||||||||||
| 12 | 2.55 | 169.01335 | −2.060 | C7H6O5 | gallic acid | 0.12 ± 0.00 | ||||
| 13 | 5.94 | 137.02328 | −4.298 | C7H6O3 | p -hydroxybenzoic acid | 0.36 ± 0.02 | ||||
| 14 | 8.53 | 153.01830 | −3.160 | C7H6O4 | protocatechuic acid | 0.27 ± 0.00 | ||||
| 15 | 6.85 | 179.03415 | −1.584 | C9H8O4 | caffeic acid | 0.46 ± 0.01 | ||||
| 16 | 8.81 | 300.99899 | 1.821 | C14H6O8 | ellagic acid | 496.64 ± 2.91 | ||||
| 17 | 13.38 | 147.04410 | −3.430 | C9H8O2 | cinnamic acid | 0.29 ± 0.05 | ||||
| Total phenolic acids | 498.14 | |||||||||
| Bacteroides | Bifidobacterium | Ruminococcus | Escherichia coli | Butyrate- Producing Bacteria | Lactobacillus | Enterococcus | Akkermansia | |
|---|---|---|---|---|---|---|---|---|
| NFRJ | 1.00 ± 0.00 a | 1.00 ± 0.00 b | 1.00 ± 0.00 a | 1.00 ± 0.00 c | 1.00 ± 0.00 b | 1.00 ± 0.00 c | 1.00 ± 0.00 a | 1.00 ± 0.00 b |
| FRJ | 0.16 ± 0.02 b | 1.14 ± 0.12 b | 0.82 ± 0.14 b | 1.28 ± 0.42 b | 1.37 ± 0.22 a | 5.56 ± 0.19 b | 1.06 ± 0.14 a | 1.74 ± 0.14 a |
| Group | Acetic Acid (mmol/L) | Propionic Acid (mmol/L) | Butyric Acid (mmol/L) | Valeric Acid (mmol/L) | Isovaleric Acid (mmol/L) |
|---|---|---|---|---|---|
| NFRJ | 275.07 ± 24.38 a | 5.37 ± 1.86 a | 16.69 ± 2.06 a | 1.23 ± 0.23 b | 3.94 ± 0.24 b |
| FRJ | 266.83 ± 12.92 a | 4.54 ± 0.23 ab | 17.75 ± 0.62 a | 1.63 ± 0.05 a | 8.03 ± 0.29 a |
| Group | Acetic Acid (mmol/L) | Propionic Acid (mmol/L) | Butyric Acid (mmol/L) | Isovaleric Acid (mmol/L) |
|---|---|---|---|---|
| C | 281.42 ± 8.29 c | 331.57 ± 60.75 a | 308.10 ± 42.75 b | 162.46 ± 12.59 b |
| L | 358.65 ± 14.44 b | 366.74 ± 57.29 a | 350.27 ± 17.38 ab | 169.08 ± 31.04 b |
| M | 378.63 ± 40.78 a | 377.61 ± 19.52 a | 395.48 ± 20.66 a | 207.00 ± 11.03 a |
| H | 342.17± 8.40 b | 362.43 ± 30.35 a | 374.40 ± 23.03 a | 202.75 ± 11.96 a |
| Group | ZO-1 | Claudin-1 | Claudin-4 | Occludin | E-cadherin | Muc-2 |
|---|---|---|---|---|---|---|
| C | 1.00 ± 0.00 a | 1.00 ± 0.00 a | 1.00 ± 0.00 a | 1.00 ± 0.00 a | 1.00 ± 0.00 a | 1.00 ± 0.00 a |
| L | 1.41 ± 0.13 b | 1.31 ± 0.13 b | 1.30 ± 0.06 b | 1.13 ± 0.08 b | 1.19 ± 0.09 b | 1.49 ± 0.21 b |
| M | 1.82 ± 0.06 c | 1.48 ± 0.13 c | 1.43 ± 0.08 c | 1.33 ± 0.11 c | 1.35 ± 0.24 b | 1.82 ± 0.33 c |
| H | 1.58 ± 0.10 bc | 1.61 ± 0.04 d | 1.42 ± 0.12 c | 1.32 ± 0.05 c | 1.30 ± 0.07 b | 1.67 ± 0.18 bc |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, T.; Chu, X.; Cheng, Y.; Tang, S.; Zogona, D.; Pan, S.; Xu, X. Modulation of Gut Microbiota by Lactobacillus casei Fermented Raspberry Juice In Vitro and In Vivo. Foods 2021, 10, 3055. https://doi.org/10.3390/foods10123055
Wu T, Chu X, Cheng Y, Tang S, Zogona D, Pan S, Xu X. Modulation of Gut Microbiota by Lactobacillus casei Fermented Raspberry Juice In Vitro and In Vivo. Foods. 2021; 10(12):3055. https://doi.org/10.3390/foods10123055
Chicago/Turabian StyleWu, Ting, Xueqi Chu, Yuxin Cheng, Shuxin Tang, Daniel Zogona, Siyi Pan, and Xiaoyun Xu. 2021. "Modulation of Gut Microbiota by Lactobacillus casei Fermented Raspberry Juice In Vitro and In Vivo" Foods 10, no. 12: 3055. https://doi.org/10.3390/foods10123055
APA StyleWu, T., Chu, X., Cheng, Y., Tang, S., Zogona, D., Pan, S., & Xu, X. (2021). Modulation of Gut Microbiota by Lactobacillus casei Fermented Raspberry Juice In Vitro and In Vivo. Foods, 10(12), 3055. https://doi.org/10.3390/foods10123055