Abstract
We consider a delayed prey–predator model incorporating a refuge with a non-monotone functional response. It is supposed that prey can live in the predatory region and prey refuge, respectively. Based on Mawhin’s coincidence degree and nontrivial estimation techniques for a priori bounds of unknown solutions to the operator equation , we prove the existence of multiple periodic solutions. Finally, an example demonstrates the feasibility of our main results.
1. Introduction
The predator–prey model [1,2] generally takes the form of
To this day, the Lotka–Volterra type system is one of the important themes in mathematical biology. Many scholars have made contributions to it (see, e.g., [3,4,5,6,7,8,9,10,11,12,13,14,15,16,17,18,19,20,21]). In the interaction between prey and predator, there always exists a phenomenon of prey refuge. In general, the entire prey population lives in two areas: the predatory region and the prey refuge. From a biological view, the prey refuge can exist without predators; therefore, it can help improve the population density of the prey. In addition, a prey refuge is an effective strategy to reduce predation in the evolution of prey population. For this reason, it was proposed by Gause et al. [22,23]. Moreover, Magalhes [24] considered the refuge effect on the dynamics of thrips prey and mite predators. Ghosh et al. [25] studied the influence on a predator–prey system of adding extra food for predators and incorporating a prey refuge. Xie [26] investigated a prey–predator model incorporating fractional-order factors and a prey refuge. When in a high-prey refuge ecological system, Sahoo et al. [27,28] observed that the possibility of predator extinction could be eliminated by providing additional food to the predator population. Motivated by these works, Jana et al. [29] considered the following prey–predator model with prey refuge:
where u is the density of the prey in the refuge and v is the density the prey in the predatory region; w denotes the density of the predator in the predatory region; the intrinsic growth rate for u and v is denoted by and , respectively; the prey migrating in the refuge to the predatory region is given by and migrating from the predatory region to the refuge is denoted by ; and are the environment carrying capacity for u and v, respectively; and are the natural death rate and the density dependent mortality rate of predator, respectively; the rate of the predator consuming prey is denoted by (suppose that ); is the delay; and a and are two parameters in Holling type II functional response.
On the other hand, functional response is also an important factor affecting the predator–prey model. In [30], we proved that there exists at least one positive periodic solution for Jana’s model with a Holling type II functional response. However, there exists a functional response unlike the Holling functional responses. Some experiments have indicated that it may occur at the microbial level: when nutrient concentrations reach a high level, they may inhibit specific growth rates; see [31]. Hence, the non-monotone functional response was considered and used to model the inhibitory effect at high concentrations [32,33,34]. In addition, in real-world applications, some researchers believe that predators living in the predatory region are classified by two fixed ages: one is mature predators, and the other is immature predators—the immature predator have no ability to attack prey; see [35,36,37,38,39].
Motivated by the works of Jana et al. [29] and Chen [34], in the present paper, we establish the following delayed stage-structured prey–predator model with a prey refuge and non-monotone functional response:
where and are the density of the immature predator and mature predator at time t, respectively. , , , , , , , , , and are all continuously positive periodic solutions with period . and are the death rate of the predator. Moreover, the non-monotone functional response is . The term
indicates the number of immature predators that were born at time that still survive at time t and then become mature predators. Moreover, the term denotes the stage-structured degree of the immature predator; one can refer to Liu et al. (see [11] pp. 670–671).
In terms of the number of creatures, the initial conditions are associated with
It is well known that the global existence of periodic solutions is a very basic and important problem in the study of periodic population dynamics. Because periodic environments such as the seasonable effect are important factors, the existence of periodic solutions plays a similar role to a global equilibrium in an autonomous model. Thus, the aim of the present paper is to find some suitable conditions of the existence of positive periodic solutions for system (2). Based on Mawhin’s coincidence degree theory and other nontrivial techniques, we prove the existence of multiple positive periodic solutions for system (2) in Section 2. In Section 3, we propose some examples to demonstrate the feasibility of our main results.
2. The Existence of Multiple Positive Periodic Solutions
Firstly, we separate the third equation of system (2) from the whole system and obtain the following subsystem:
The initial values for system (3) are
To obtain the multiple positive periodic solutions of system (3), we summarize the following lemmas.
Lemma 1.
Let be an open bounded set. Assume that L is a Fredholm operator of index zero and N is L-compact on . Furthermore, if the following conditions are fulfilled
- (a)
- for each fixed ;
- (b)
- for each fixed , and ;then the operator equation has at least one solution in .
Note that operator L is said to be a Fredholm operator of index zero if and is closed in V. If L is a Fredholm mapping of index zero, then there exist continuous projectors and such that and , where are Banach spaces. For further symbolic meaning of the concepts in Lemma 2.1, one can refer to [40,41,42] for details.
Lemma 2
([32,41]). Assume that is an ω-periodic function that is continuously differentiable. Then, there exists a such that
For the sake of convenience, we use notations as follows.
Furthermore, suppose that
As mentioned in the last paragraph of the introduction, periodic solutions are of great importance. It is reasonable to seek conditions (that is, , ) under which the resulting periodic system would have positive periodic solutions. From a biological viewpoint, the assumption implies that the rate of predator feeding on prey is affected by the death rate of predators, the time delay, and the migration of prey, while the assumption implies that the intrinsic growth rate of prey in a predator region is influenced by prey migration, environmental carrying capacity, and predator predation.
Now, we are in a position to state our main theorem.
Theorem 1.
If and hold, then system (2) has at least two positive periodic solutions.
Proof.
We prove this theorem into two steps.
- Step 1
We claim that there are at least two periodic solutions of subsystem (3). In fact, by the variables transformation
then subsystem (3) reads
where
Define
then are both Banach Spaces with the norm as follows:
For any , it is easy to see that
and
are all -periodic functions. Indeed,
Clearly, are also periodic functions in a similar way. Set
where and is given by
Define
It follows form the above definitions that and , It is easy to see that the inverse Therefore,
and
Obviously, and are continuous. It follows from the Arzela–Ascoli theorem [34,40] that N is L-compact on with any open-bounded set
The next work is to find an appropriate open-bounded subset for the application of the continuation theorem. Corresponding to for some , we obtain
Similarly, from the second equation of (4), (5) and from the third equation of (4), (5), the following inequalities hold:
Combining Lemma 2 with these inequalities, we next construct the upper and lower bounds of subsystem (3). Since , there exist such that
This implies that
therefore, we have
or
In particular,
or
Therefore, in view of , we obtain
Now, let Equation (11) equal 0; we see that
Hence,
This can be rewritten as
In view of , we obtain
Let Equation (13) equal 0; we see that
Therefore,
Clearly,
Hence, we take
Therefore, we know that
Therefore, we take
In view of , we see that
In a similar way, from the second equation of (5), we have
Then we take
Now, we consider with . Note that
where .
From and , we know that the equation has two different solutions: . Thus, we have the following formula:
- (1)
- Since , we have
- (2)
- Since , we have
- (3)
- Since , we have
Define
We choose , such that
Let
Then, both and are bounded open subsets of V, and . It is easily noticed that and satisfies the requirement in Lemma 1 for . In addition, for . A direct computation gives
Therefore, system (3) has at least two -periodic solutions . Let , , and , , . Then, by Equation (4), and are two different positive -periodic solutions of (3).
- Step 2
We now claim that the third equation of (2) has two -periodic solutions associated with the obtained solutions. Let
Then, the third equation of (2) is given by
In fact, we see that
Similarly,
Since is negative and , the linear system admits exponential dichotomy. Hence,
Consequently, and are two different periodic solutions of system(2). □
3. Example
Corresponding to system (2), we give an example as follows:
where and , and , , , and . here is the non-monotone functional response, which reflects the capture ability of the mature predator at time t. The term
stands for the number of immature predators born at time that still survive at time t and become mature predators.
Taking the initial values , , , and , the periodic solution is shown in Figure 1.
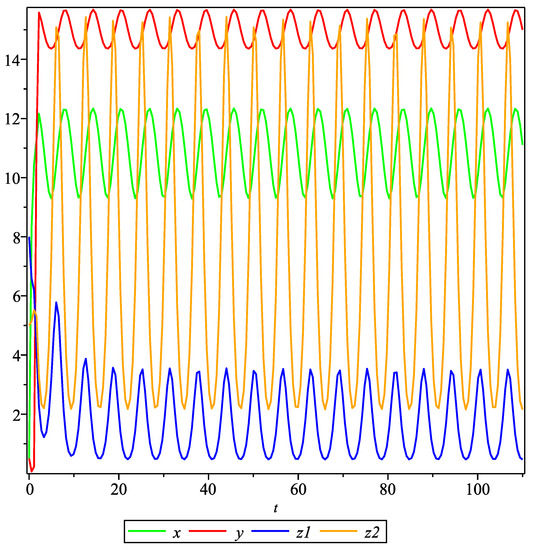
Figure 1.
The periodic solution.
4. Conclusions
In this paper, a delayed prey–predator model incorporating a refuge with a non-monotone functional response is considered. It is assumed that prey can live in the predatory region and the prey refuge, respectively. From the biological point, a prey refuge can help to improve the population density of the prey, and it is an effective strategy to reduce predation in the evolution of prey population. Based on the method of Mawhin’s coincidence degree theory and non-trivial estimation techniques for a priori bounds of unknown solutions to the operator equation , we obtain some interesting and novel sufficient conditions for the existence of multiple periodic solutions of the prey–predator model. However, the limitation of this method is that we cannot determine the specific number of periodic solutions. However, predictably, for different biological models, we can give a lower bound on the number of periodic solutions. In addition, we believe that, in this paper, the occurrence of two periodic solutions is influenced by non-monotonic functional response. These two periodic solutions are generated by the system for parameters in different parameter ranges, but the periods are the same. Namely, in the bounded open subset , system (2) presentws a positive periodic solution; in another bounded open subset , system (2) also has a positive periodic solution with the same period.
Author Contributions
Methodology, W.L. and Y.X.; software and figure, W.L.; validation, W.L. and Y.X.; formal analysis, W.L. and Y.X.; investigation, W.L. and Y.X.; resources, W.L. and Y.X.; writing—original draft preparation, W.L. and Y.X.; writing—review and editing, W.L. and Y.X.; supervision, Y.X. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the National Natural Science Foundation of China under Grant (No. 11931016).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data used to support the findings of this study are available from the corresponding author upon request.
Conflicts of Interest
The authors declare that there is no conflict of interest regarding the publication of this article.
References
- Lotka, A. Elements of Physical Biology; Williams Wilkins Co.: Balitmore, MD, USA, 1925. [Google Scholar]
- Volterra, V. Variazioni e fluttuazioni del numero dindividui in specie animali conviventi. Mem. Acad Lincei Roma 1926, 2, 31–113. [Google Scholar]
- Chen, X.; Du, Z. Existence of positive periodic solutions for a neutral delay predator-prey model with Hassell-Varley type functional response and impulse. Qual. Theory Dyn. Syst. 2018, 17, 67–80. [Google Scholar] [CrossRef]
- Lv, Y.; Du, Z. Existence and global attractivity of a positive periodic solution to a Lotka–Volterra model with mutual interference and Holling III type functional response. Nonlinear Anal. RWA 2011, 12, 3654–3664. [Google Scholar] [CrossRef]
- Wang, D. Positive periodic solutions for a nonautonomous neutral delay prey-predator model with impulse and Hassell-Varley type functional response. Proc. Am. Math. Soc. 2014, 142, 623–638. [Google Scholar] [CrossRef]
- Wang, D. Four positive periodic solutions of a delayed plankton allelopathy system on time scales with multiple exploited (or harvesting) terms. IMA J. Appl. Math. 2013, 78, 449–473. [Google Scholar] [CrossRef]
- Yu, S.; Liu, J.; Lin, X. Multiple positive periodic solutions of a Gause-type predator-prey model with Allee effect and functional responses. AIMS Math. 2020, 5, 6135–6148. [Google Scholar] [CrossRef]
- Liu, Y.; Zhang, X.; Zhou, T. Multiple periodic solutions of a delayed predator-prey model with non-monotonic functional response and stage structure. J. Biol. Dynam. 2014, 1, 145–160. [Google Scholar] [CrossRef][Green Version]
- Zhang, Z.; Luo, Z. Multiple periodic solutions of a delayed predator–prey system with stage structure for the predator. Nonlinear Anal. RWA 2010, 1, 4109–4120. [Google Scholar] [CrossRef]
- Kant, S.; Kumar, V. Stability analysis of predator-prey system with migrating prey and disease infection in both species. Appl. Math. Model. 2017, 42, 509–539. [Google Scholar] [CrossRef]
- Liu, S.; Chen, L.; Liu, Z. Extinction and permanence in nonautonomous competitive system with stage structure. J. Math. Anal. Appl. 2002, 274, 667–684. [Google Scholar] [CrossRef]
- Zhang, T.; Li, H.; Xi, N.; Fu, W.; Wang, K.; Ding, X. Mathematical analysis and simulation of a Hepatitis B model with time delay: A case study for Xinjiang, China. Math. Biosci. Eng. 2020, 17, 1757–1775. [Google Scholar] [CrossRef] [PubMed]
- Lu, S. On the existence of positive periodic solutions to a Lotka-Volterra cooperative population model with multiple delays. Nonlinear Anal. 2008, 68, 1746–1753. [Google Scholar] [CrossRef]
- Song, Y.; Tang, X. Stability, steady-state bifurcations and turing patterns in a predator-prey model with herd behavior and prey-taxis. Stud. Appl. Math. 2017, 139, 371–404. [Google Scholar] [CrossRef]
- Lv, Y.; Chen, L.; Chen, F.; Li, Z. Stability and bifurcation in an SI epidemic model with additive Allee effect and time delay. Int. J. Bifurcat. Chaos 2021, 31, 2150060. [Google Scholar] [CrossRef]
- Chen, L.; Liu, T.; Chen, F. Stability and bifurcation in a two-patch model with additive Allee effect. AIMS Math. 2022, 7, 536–551. [Google Scholar] [CrossRef]
- Wei, Z.; Xia, Y.; Zhang, T. Stability and bifurcation analysis of a amensalism model with weak Allee effect. Qual. Theor. Dyn. Syst. 2020, 19, 23. [Google Scholar] [CrossRef]
- Wei, Z.; Xia, Y.; Zhang, T. Stability and bifurcation analysis of a commensal model with additive Allee effect and nonlinear growth rate. Int. J. Bifurcat. Chaos 2021, 31, 2150204. [Google Scholar] [CrossRef]
- Xu, R.; Ma, Z. Stability and Hopf bifurcation in a ratio-dependent predator prey system with stage structure. Chaos Solitons Fractals 2008, 38, 669–684. [Google Scholar] [CrossRef]
- Xu, J.; Zhang, T.; Song, K. A stochastic model of bacterial infection associated with neutrophils. Appl. Math. Comput. 2020, 373, 125025. [Google Scholar] [CrossRef]
- Chen, L.; Chen, F. Qualitative analysis of a predator-prey model with Holling type II functional response incorporating a constant prey refuge. Nonlinear Anal. 2010, 11, 246–252. [Google Scholar] [CrossRef]
- Gause, G.; Smaragdova, N.; Witt, A. Further studies of interaction between predators and prey. J. Anim. Ecol. 1936, 5, 1–18. [Google Scholar] [CrossRef]
- Gause, G. The Struggle for Existence; Williams Wilkins Co.: Balitmore, MD, USA, 1934. [Google Scholar]
- Magalhᾶes, S.; Rijn, P.; Montserrat, M.; Pallini, A.; Sabelis, M. Population dynamics of thrips prey and their mite predators in a refuge. Oecologia 2007, 150, 557–568. [Google Scholar]
- Ghosh, J.; Sahoo, B.; Poria, S. Prey-predator dynamics with prey refuge providing additional food to predator. Chaos Solitons Fractals 2017, 96, 110–119. [Google Scholar] [CrossRef]
- Xie, Y.; Lu, J.; Wang, Z. Stability analysis of a fractional-order diffused prey-predator model with prey refuges. Physica A 2019, 526, 120773. [Google Scholar] [CrossRef]
- Sahoo, B.; Poria, S. Effects of additional food in a delayed predator-prey model. Math. Biosci. 2015, 261, 62–73. [Google Scholar] [CrossRef] [PubMed]
- Sahoo, B.; Poria, S. Dynamics of predator-prey system with fading memory. Appl. Math. Comput. 2019, 347, 319–333. [Google Scholar] [CrossRef]
- Jana, S.; Chakraborty, M.; Chakraborty, K.; Kar, T. Global stability and bifurcation of time delayed prey-predator system incorporating prey refuge. Math. Comput. Simulat. 2012, 85, 57–77. [Google Scholar] [CrossRef]
- Lu, W.; Xia, Y.; Bai, Y. Periodic solution of a stage-structured predator-prey model incorporating prey refuge. Math. Biosci. Eng. 2020, 4, 3160–3174. [Google Scholar] [CrossRef]
- Bush, A.; Cook, A. The effect of time delay and growth rate inhibition in the bacterial treatment of wastewater. J. Theor. Biol. 1976, 63, 385–395. [Google Scholar] [CrossRef]
- Chen, L. Mathematical Models and Methods in Ecology; Science Press: Beijing, China, 1998. (In Chinese) [Google Scholar]
- Xia, Y.; Cao, J.; Cheng, S. Multiple periodic solutions of a delayed stage-structured predator-prey model with nonmonotone functional responses. Appl. Math. Model. 2007, 9, 1947–1959. [Google Scholar] [CrossRef]
- Chen, Y. Multiple periodic solution of delayed predator-prey systems with type IV functional responses. Nonlinear Anal. 2004, 5, 45–53. [Google Scholar] [CrossRef]
- Aiello, W.; Freedman, H.; Wu, J. Analysis of a model representing stage-structured population growth with state-dependent time delay. SIAM J. Appl. Math. 1992, 52, 885–889. [Google Scholar] [CrossRef]
- Brauer, F.; Ma, Z. Stability of stage-structured population models. J. Math. Anal. Appl. 1987, 126, 301–315. [Google Scholar] [CrossRef]
- Freedman, H.; Wu, J. Persistence and global asymptotic stability of single species dispersal models with stage-structure. Q. Appl. Math. 1991, 49, 351–371. [Google Scholar] [CrossRef]
- Wang, W.; Chen, L. A predator-prey system with stage-structure for predator. Comput. Math. Appl. 1997, 33, 83–91. [Google Scholar] [CrossRef]
- Wang, W.; Mulone, G.; Salemi, F.; Salone, V. Permanence and stability of a stage-structured predator prey model. J. Math. Anal. Appl. 2001, 262, 499–528. [Google Scholar] [CrossRef]
- Fan, M.; Wang, Q.; Zou, X. Dynamics of a nonautonomous ratio-dependent predator-prey system. Proc. R. Soc. Edinb. Sect. A Math. 2003, 133, 97–118. [Google Scholar] [CrossRef]
- Fan, M.; Wong, P.; Agarwal, R. Periodicity and stability in periodic n-species Lotka-Volterra competition system with feedback controls and deviating arguments. Acta. Math. Sin. 2003, 19, 801–822. [Google Scholar] [CrossRef]
- Gaines, R.; Mawhin, J. Coincidence Degree and Nonlinear Differential Equations; Lecture Notes in Mathematics; Springer: Berlin/Heidelberg, Germany, 2006; Volume 568. [Google Scholar]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).