The Future of Semi-Arid Regions: A Weak Fabric Unravels


{kind=link}
Abstract
1. Introduction: Regions with Semi-Arid Climates as a Distinct Global Entity
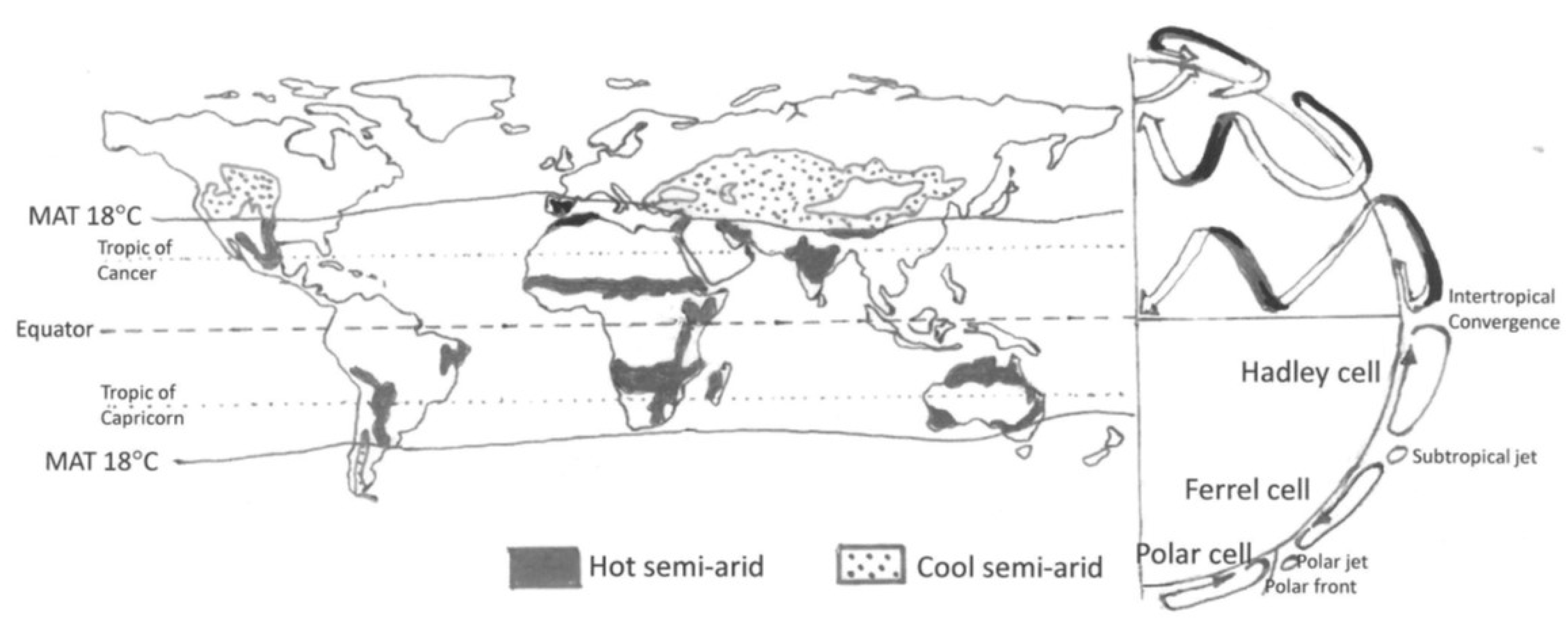
2. The Climatic Determinants of Semi-Aridity and How They are Changing
3. Plant Production Dynamics in Semi-Arid Ecosystems
4. Plant and Animal and Coping Strategies and Their Limits
5. The Failure of Coupled Socio-Ecological Systems in Semiarid Regions
6. Interventions in Semi-Arid Lands
6.1. Actions That are Unlikely to Help
6.2. Interventions with Greater Promise of Success
Funding
Acknowledgments
Conflicts of Interest
References
- Higgins, G.M.; Kassam, A.H. The FAO agro-ecological zone approach to determination of land potential. Pedologie 1981, 31, 147–168. [Google Scholar]
- Black, J.N. The distribution of solar radiation over the earth’s surface. Arch. Meteorol. Geophys. Bioklimatol. Ser. B 1956, 7, 165–189. [Google Scholar] [CrossRef]
- Dima, I.M.; Wallace, J.M. On the seasonality of the Hadley cell. J. Atmos. Sci. 2003, 60, 1522–1527. [Google Scholar] [CrossRef]
- Walsh, R.P.D.; Lawler, D.M. Rainfall seasonality: Description, spatial patterns and change through time. Weather 1981, 36, 201–208. [Google Scholar] [CrossRef]
- Camberlin, P.; Janicot, S.; Poccard, I. Seasonality and atmospheric dynamics of the teleconnection between African rainfall and tropical sea-surface temperature: Atlantic vs. ENSO. Int. J. Climatol. A J. R. Meteorol. Soc. 2001, 21, 973–1005. [Google Scholar] [CrossRef]
- Ellis, J.E.; Swift, D.M. Stability of African pastoral ecosystems: Alternate paradigms and implications for development. J. Rangel. Manag. 1988, 41, 450–459. [Google Scholar] [CrossRef]
- Von Wehrden, H.; Hanspach, J.; Kaczensky, P.; Fischer, J.; Wesche, K. Global assessment of the non-equilibrium concept in rangelands. Ecol. Appl. 2012, 22, 393–399. [Google Scholar] [CrossRef] [PubMed]
- Aguiar, M.R.; Sala, O.E. Patch structure, dynamics and implications for the functioning of arid ecosystems. Trends Ecol. Evol. 1999, 14, 273–277. [Google Scholar] [CrossRef]
- Taylor, C.M.; Birch, C.E.; Parker, D.J.; Dixon, N.; Guichard, F.; Nikulin, G.; Lister, G.M. Modeling soil moisture-precipitation feedback in the Sahel: Importance of spatial scale versus convective parameterization. Geophys. Res. Lett. 2013, 40, 6213–6218. [Google Scholar] [CrossRef]
- Milne, G.A. A provisional soil map of East Africa. East Afr. Agric. Res. Stn. 1936, 88, 465. [Google Scholar]
- Scholes, R.J.; Bond, W.J.; Eckhardt, H.C. Vegetation dynamics in the Kruger ecosystem. In The Kruger Experience: Ecology and Management of Savanna Heterogeneity; Island Press: Washington, DC, USA, 2003; pp. 242–262. [Google Scholar]
- Mueller, E.N.; Wainwright, J.; Parsons, A.J. Spatial variability of soil and nutrient characteristics of semi-arid grasslands and shrublands, Jornada Basin, New Mexico. Ecohydrol. Ecosyst. Land Water Process Interact. Ecohydrogeomorphol. 2008, 1, 3–12. [Google Scholar] [CrossRef]
- Hallegatte, S.; Rozenberg, J. Climate change through a poverty lens. Nat. Clim. Chang. 2017, 7, 250–256. [Google Scholar] [CrossRef]
- Cook, B.I.; Miller, R.L.; Seager, R. Amplification of the North American “Dust Bowl” drought through human-induced land degradation. Proc. Natl. Acad. Sci. USA 2009, 106, 4997–5001. [Google Scholar] [CrossRef] [PubMed]
- Fu, B.; Wang, S.; Liu, Y.; Liu, J.; Liang, W.; Miao, C. Hydrogeomorphic ecosystem responses to natural and anthropogenic changes in the Loess Plateau of China. Annu. Rev. Earth Planet. Sci. 2017, 45, 223–243. [Google Scholar] [CrossRef]
- Chartres, C. Australia's Land Resources at Risk; Cambridge University Press: Cambridge, UK, 1987; pp. 7–26. [Google Scholar]
- Hoffmann, T.; Todd, S.; Ntshona, Z.; Turner, S. Land Degradation in South Africa; University of Cape Town: Cape Town, South Africa, 2014. [Google Scholar]
- Nel, E.; Hill, T. Marginalisation and demographic change in the semi-arid Karoo, South Africa. J. Arid Environ. 2008, 72, 2264–2274. [Google Scholar] [CrossRef]
- Pavanello, S. Pastoralists’ Vulnerability in the Horn of Africa: Exploring Political Marginalisation, Donors’ Policies and Cross-Border Issues–Literature Review; Humanitarian Policy Group (HPG) Overseas Development Institute: London, UK, 2009. [Google Scholar]
- Blaikie, P. The Political Economy of Soil Erosion in Developing Countries; Routledge: London, UK, 2016. [Google Scholar]
- Schneider, T.; Bischoff, T.; Haug, G.H. Migrations and dynamics of the Intertropical Convergence Zone. Nature 2014, 513, 45–53. [Google Scholar] [CrossRef]
- Nicholson, S.E. The ITCZ and the Seasonal Cycle over Equatorial Africa. Bull. Am. Meteorol. Soc. 2018, 99, 337–348. [Google Scholar] [CrossRef]
- Gill, A.E. Some simple solutions for heat-induced tropical circulation. Q. J. R. Meteorol. Soc. 1980, 106, 447–462. [Google Scholar] [CrossRef]
- Intergovernmental Panel on Climate Chang. The Physical Science Basis-Summary for Policymakers, Observed Changes in the Climate System; IPCC: Geneva, Switzerland, 2013; p. 15. [Google Scholar]
- Fernandez, J.P.R.; Franchito, S.H.; Rao, V.B. Future changes in the aridity of South America from regional climate model projections. Pure Appl. Geophys. 2019, 176, 2719–2728. [Google Scholar] [CrossRef]
- Chen, C.; Cane, M.A.; Wittenberg, A.T.; Chen, D. ENSO in the CMIP5 simulations: Life cycles, diversity, and responses to climate change. J. Clim. 2017, 30, 775–801. [Google Scholar] [CrossRef]
- McNaughton, S.J. Ecology of a grazing ecosystem: The Serengeti. Ecol. Monogr. 1985, 55, 259–294. [Google Scholar] [CrossRef]
- Sala, O.E.; Parton, W.J.; Joyce, L.A.; Lauenroth, W.K. Primary production of the central grassland region of the United States. Ecology 1988, 69, 40–45. [Google Scholar] [CrossRef]
- Sala, O.E.; Gherardi, L.A.; Reichmann, L.; Jobbagy, E.; Peters, D. Legacies of precipitation fluctuations on primary production: Theory and data synthesis. Philos. Trans. R. Soc. B Biol. Sci. 2012, 367, 3135–3144. [Google Scholar] [CrossRef] [PubMed]
- Scholes, R.J. Convex relationships in ecosystems containing mixtures of trees and grass. Environ. Resour. Econ. 2003, 26, 559–574. [Google Scholar] [CrossRef]
- Van den Hoof, C.; Verstraete, M.; Scholes, R.J. Differing Responses to Rainfall Suggest More Than One Functional Type of Grassland in South Africa. Remote Sens. 2018, 10, 2055. [Google Scholar] [CrossRef]
- Scholes, R.J. Syndromes of dryland degradation in southern Africa. Afr. J. Range Forage Sci. 2009, 26, 113–125. [Google Scholar] [CrossRef]
- Montanarella, L.; Scholes, R.; Brainich, A. (Eds.) The IPBES Assessment Report on Land Degradation and Restoration; Secretariat of the Intergovernmental Science-Policy Platform on Biodiversity and Ecosystem Services: Bonn, Germany, 2018. [Google Scholar]
- Vicente-Serrano, S.M.; Cabello, D.; Tomás-Burguera, M.; Martín-Hernández, N.; Beguería, S.; Azorin-Molina, C.; Kenawy, A.E. Drought variability and land degradation in semiarid regions: Assessment using remote sensing data and drought indices (1982–2011). Remote Sens. 2015, 7, 4391–4423. [Google Scholar] [CrossRef]
- Herrmann, S.M.; Hutchinson, C.F. The scientific basis: Links between land degradation, drought and desertification. In Governing Global Desertification; Routledge: London, UK, 2016; pp. 31–46. [Google Scholar]
- Williams, C.A.; Hanan, N.; Scholes, R.J.; Kutsch, W. Complexity in water and carbon dioxide fluxes following rain pulses in an African savanna. Oecologia 2009, 161, 469–480. [Google Scholar] [CrossRef]
- Osakabe, Y.; Osakabe, K.; Shinozaki, K.; Tran, L.S.P. Response of plants to water stress. Front. Plant Sci. 2014, 5, 86. [Google Scholar] [CrossRef]
- Mirfenderesgi, G.; Matheny, A.M.; Bohrer, G. Hydrodynamic trait coordination and cost–benefit trade-offs throughout the isohydric–anisohydric continuum in trees. Ecohydrology 2019, 12, e2041. [Google Scholar] [CrossRef]
- Tilman, D.; El Haddi, A. Drought and biodiversity in grasslands. Oecologia 1992, 89, 257–264. [Google Scholar] [CrossRef] [PubMed]
- Symstad, A.J.; Chapin, F.S.; Wall, D.H.; Gross, K.L.; Huenneke, L.F.; Mittelbach, G.G.; Peters, D.P.; Tilman, D. Long-term and large-scale perspectives on the relationship between biodiversity and ecosystem functioning. Bioscience 2003, 53, 89–98. [Google Scholar] [CrossRef]
- Petrie, M.D.; Bradford, J.B.; Hubbard, R.M.; Lauenroth, W.K.; Andrews, C.M.; Schlaepfer, D.R. Climate change may restrict dryland forest regeneration in the 21st century. Ecology 2017, 98, 1548–1559. [Google Scholar] [CrossRef]
- Walker, B.H.; Emslie, R.H.; Owen-Smith, R.N.; Scholes, R.J. To cull or not to cull: Lessons from a southern African drought. J. Appl. Ecol. 1987, 24, 381–401. [Google Scholar] [CrossRef]
- Hitchcock, R.K. Coping with Uncertainty: Adaptive Responses to Drought. In Sustainable Livelihoods in Kalahari Environments: A Contribution to Global Debates; Oxford Geographical and Environmental Studies; Oxford University Press: Oxford, UK, 2002; pp. 161–171. [Google Scholar]
- Conradie, S.R.; Woodborne, S.M.; Cunningham, S.J.; McKechnie, A.E. Chronic, sublethal effects of high temperatures will cause severe declines in southern African arid-zone birds during the 21st century. Proc. Natl. Acad. Sci. USA 2019, 116, 14065–14070. [Google Scholar] [CrossRef]
- Suzman, J. Affluence without Abundance: The Disappearing World of the Bushmen; Bloomsbury Publishing: New York, NY, USA, 2017. [Google Scholar]
- Hermans-Neumann, K.; Priess, J.; Herold, M. Human migration, climate variability, and land degradation: Hotspots of socio-ecological pressure in Ethiopia. Reg. Environ. Chang. 2017, 17, 1479–1492. [Google Scholar] [CrossRef]
- Hummel, D. Climate change, land degradation and migration in Mali and Senegal–some policy implications. Migr. Dev. 2015, 5, 211–233. [Google Scholar] [CrossRef]
- Buhaug, H.; Nordkvelle, J.; Bernauer, T. One effect to rule them all? A comment on climate and conflict. Clim. Chang. 2014, 127, 391–397. [Google Scholar] [CrossRef]
- Xu, J. Sand-dust storms in and around the Ordos Plateau of China as influenced by land use change and desertification. Catena 2006, 65, 279–284. [Google Scholar] [CrossRef]
- Al-Awadhi, J.M.; Al-Dousari, A.M.; Khalaf, F.I. Influence of land degradation on the local rate of dust fallout in Kuwait. Atmos. Clim. Sci. 2014, 4, 437–446. [Google Scholar] [CrossRef]
- De la Paix, M.J.; Lanhai, L.; Xi, C.; Ahmed, S.; Varenyam, A. Soil degradation and altered flood risk as a consequence of deforestation. Land Degrad. Dev. 2013, 24, 478–485. [Google Scholar] [CrossRef]
- Paeth, H.; Born, K.; Girmes, R.; Podzun, R.; Jacob, D. Regional climate change in tropical and northern Africa due to greenhouse forcing and land use changes. J. Clim. 2009, 22, 114–132. [Google Scholar] [CrossRef]
- Arribas, A.; Gallardo, C.; Gaertner, M.; Castro, M. Sensitivity of the Iberian Peninsula climate to a land degradation. Clim. Dyn. 2003, 20, 477–489. [Google Scholar] [CrossRef]
- Bonn Challenge. Available online: www.bonnchallenge.org (accessed on 12 February 2020).
- Sankaran, M.; Hanan, N.P.; Scholes, R.J.; Ratnam, J.; Augustine, D.J.; Cade, B.S.; Gignoux, J.; Higgins, S.I.; Le Roux, X.; Ludwig, F.; et al. Determinants of woody cover in African savannas. Nature 2005, 438, 846–849. [Google Scholar] [CrossRef]
- Bond, W.J.; Stevens, N.; Midgley, G.F.; Lehmann, C.E. The trouble with trees: Afforestation plans for Africa. Trends Ecol. Evol. 2019, 34, 963–965. [Google Scholar] [CrossRef]
- Allison, G.B.; Gee, G.W.; Tyler, S.W. Vadose-zone techniques for estimating groundwater recharge in arid and semiarid regions. Soil Sci. Soc. Am. J. 1994, 58, 6–14. [Google Scholar] [CrossRef]
- Sternberg, T. Piospheres and pastoralists: Vegetation and degradation in steppe grasslands. Hum. Ecol. 2012, 40, 811–820. [Google Scholar] [CrossRef]
- Briske, D.D.; Derner, J.D.; Brown, J.R.; Fuhlendorf, S.D.; Teague, W.R.; Havstad, K.M.; Gillen, R.L.; Ash, A.J.; Willms, W.D. Rotational grazing on rangelands: Reconciliation of perception and experimental evidence. Rangel. Ecol. Manag. 2008, 61, 3–17. [Google Scholar] [CrossRef]
- Farmer, H. Understanding Impacts of Water Supplementation in a Heterogeneous Landscape. Ph.D. Thesis, University of the Witwatersrand, Johannesburg, South Africa, 2010. [Google Scholar]
- Cornelis, W.; Waweru, G.; Araya, T. Building Resilience against Drought and Floods: The Soil-Water Management Perspective. In Sustainable Agriculture Reviews; Springer: Cham, Switzerland, 2019; Volume 29, pp. 125–142. [Google Scholar]
- O’Farrell, P.J.; Anderson, P.M.L.; Milton, S.J.; Dean, W.R.J. Human response and adaptation to drought in the arid zone: Lessons from southern Africa. S. Afr. J. Sci. 2009, 105, 34–39. [Google Scholar]
- Hobbs, R.J. Landscapes, ecology and wildlife management in highly modified environments—An Australian perspective. Wildl. Res. 2005, 32, 389–398. [Google Scholar] [CrossRef]
- Wallander, S.; Aillery, M.; Hellerstein, D.; Hand, M. The role of conservation programs in drought risk adaptation. Econ. Res. Serv. Econ. Res. Rep. 2013, 148, 1–68. [Google Scholar]
- Young, R. Beyond the Year of the Outback: What now for rural Australia? Impact 2003, 8. [Google Scholar]
- Carruthers, J. Wilding the farm or farming the wild? The evolution of scientific game ranching in South Africa from the 1960s to the present. Trans. R. Soc. S. Afr. 2008, 63, 160–181. [Google Scholar]

© 2020 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Scholes, R.J. The Future of Semi-Arid Regions: A Weak Fabric Unravels. Climate 2020, 8, 43. https://doi.org/10.3390/cli8030043
Scholes RJ. The Future of Semi-Arid Regions: A Weak Fabric Unravels. Climate. 2020; 8(3):43. https://doi.org/10.3390/cli8030043
Chicago/Turabian StyleScholes, Robert J. 2020. "The Future of Semi-Arid Regions: A Weak Fabric Unravels" Climate 8, no. 3: 43. https://doi.org/10.3390/cli8030043
APA StyleScholes, R. J. (2020). The Future of Semi-Arid Regions: A Weak Fabric Unravels. Climate, 8(3), 43. https://doi.org/10.3390/cli8030043