Essential Oils of Zingiber Species from Vietnam: Chemical Compositions and Biological Activities

 , , , , ,
, , , , , 
Abstract
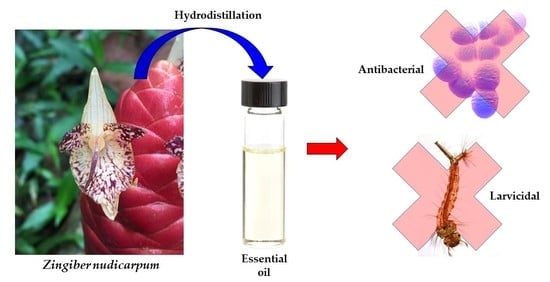
1. Introduction
2. Results and Discussion
2.1. Essential Oil Compositions
2.2. Mosquito Larvicidal Activity
2.3. Antimicrobial Activity
3. Materials and Methods
3.1. Plant Material
3.2. Gas Chromatographic Analysis
3.3. Mosquito Larvicidal Screening
3.4. Antimicrobial Screening
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Nealon, J.; Taurel, A.-F.; Yoksan, S.; Moureau, A.; Bonaparte, M.; Quang, L.C.; Capeding, M.R.; Prayitno, A.; Hadinegoro, S.R.; Chansinghakul, D.; et al. Serological evidence of Japanese encephalitis virus circulation in Asian children from dengue-endemic countries. J. Infect. Dis. 2019, 219, 375–381. [Google Scholar] [CrossRef] [PubMed]
- Thao, T.T.N.; de Bruin, E.; Phuong, H.T.; Vy, N.H.T.; van den Ham, H.-J.; Wills, B.A.; Tien, N.T.H.; Le Duyen, H.T.; Trung, D.T.; Whitehead, S.S.; et al. Using NS1 flavivirus protein microarray to infer past infecting dengue virus serotype and number of past dengue virus infections in Vietnamese individuals. J. Infect. Dis. 2020, in press. [Google Scholar] [CrossRef] [PubMed]
- Dinh, T.C.; Bac, N.D.; Minh, L.B.; Ngoc, V.T.N.; Pham, V.-H.; Vo, H.-L.; Tien, N.L.B.; Van Thanh, V.; Tao, Y.; Show, P.L.; et al. Zika virus in Vietnam, Laos, and Cambodia: Are there health risks for travelers? Eur. J. Clin. Microbiol. Infect. Dis. 2019, 38, 1585–1590. [Google Scholar] [CrossRef] [PubMed]
- Karna, A.K.; Bowen, R.A. Experimental evaluation of the role of ecologically-relevant hosts and vectors in Japanese encephalitis virus genotype displacement. Viruses 2019, 11, 32. [Google Scholar] [CrossRef]
- de Wispelaere, M.; Desprès, P.; Choumet, V. European Aedes albopictus and Culex pipiens are competent vectors for Japanese encephalitis virus. PLoS Negl. Trop. Dis. 2017, 11, e0005294. [Google Scholar] [CrossRef]
- Oliveira, A.R.S.; Cohnstaedt, L.W.; Cernicchiaro, N. Japanese encephalitis virus: Placing disease ectors in the epidemiologic triad. Ann. Entomol. Soc. Am. 2018, 111, 295–303. [Google Scholar] [CrossRef]
- Van Ta, T.; Tran, H.T.; Ha, Q.N.T.; Nguyen, X.T.; Tran, V.K.; Pham, H.T.; Simmons, C. The correlation of clinical and subclinical presentations with dengue serotypes and plasma viral load: The case of children with dengue hemorrhagic fever in Vietnam. Int. J. Res. Pharm. Sci. 2019, 10, 2578–2585. [Google Scholar] [CrossRef]
- Tun, M.M.N.; Nguyen, T.T.T.; Ando, T.; Dumre, S.P.; Soe, A.M.; Buerano, C.C.; Nguyen, M.T.; Le, N.T.N.; Pham, V.Q.; Nguyen, T.H.; et al. Clinical, virological, and cytokine profiles of children infected with dengue virus during the outbreak in southern Vietnam in 2017. Am. J. Trop. Med. Hyg. 2020, 102, 1217–1225. [Google Scholar]
- Higa, Y.; Yen, N.T.; Kawada, H.; Son, T.H.; Hoa, N.T.; Takagi, M. Geographic distribution of Aedes aegypti and Aedes albopictus collected from used tires in Vietnam. J. Am. Mosq. Control. Assoc. 2010, 26, 1–9. [Google Scholar] [CrossRef]
- Chu, D.-T.; Ngoc, V.T.N.; Tao, Y. Zika virus infection in Vietnam: Current epidemic, strain origin, spreading risk, and perspective. Eur. J. Clin. Microbiol. Infect. Dis. 2017, 36, 2041–2042. [Google Scholar] [CrossRef]
- Duong, T.T.; Van Dung, N.; Chinh, V.D.; Trung, H.D. Mapping insecticide resistance in dengue vectors in the northern Viet Nam, 2010–2013. Vector Biol. J. 2016, 1, 1–6. [Google Scholar] [CrossRef]
- Thi, K.P.; Viet, H.H.; Nguyen, H.M. Major resistant mechanism to insecticides of Aedes aegypti mosquito: A vector of dengue and Zika virus in Vietnam. SM Trop. Med. J. 2016, 1, 1010. [Google Scholar]
- Kasai, S.; Caputo, B.; Tsunoda, T.; Cuong, T.C.; Maekaw, Y.; Lam-Phua, S.G.; Pichler, V.; Itokawa, K.; Murota, K.; Komagata, O.; et al. First detection of a Vssc allele V1016G conferring a high level of insecticide resistance in Aedes albopictus collected from Europe (Italy) and Asia (Vietnam), 2016: A new emerging threat to controlling arboviral diseases. Euro Surveill. 2019, 24, 1700847. [Google Scholar] [CrossRef] [PubMed]
- Pasteur, N.; Marquine, M.; Hoang, T.H.; Nam, V.S.; Failloux, A.B. Overproduced esterases in Culex pipiens quinquefasciatus (Diptera: Culicidae) from Vietnam. J. Med. Entomol. 2001, 38, 740–745. [Google Scholar] [CrossRef] [PubMed]
- Itokawa, K.; Komagata, O.; Kasai, S.; Kawada, H.; Mwatele, C.; Dida, G.O.; Njenga, S.M.; Mwandawiro, C.; Tomita, T. Global spread and genetic variants of the two CYP9M10 haplotype forms associated with insecticide resistance in Culex quinquefasciatus Say. Heredity 2013, 111, 216–226. [Google Scholar] [CrossRef] [PubMed]
- Lien, L.T.Q.; Lan, P.T.; Chuc, N.T.K.; Hoa, N.Q.; Nhung, P.H.; Thoa, N.T.M.; Diwan, V.; Tamhankar, A.J.; Lundborg, C.S. Antibiotic resistance and antibiotic resistance genes in Escherichia coli isolates from hospital wastewater in Vietnam. Int. J. Environ. Res. Public Health 2017, 14, 699. [Google Scholar] [CrossRef] [PubMed]
- Tran, G.M.; Ho-Le, T.P.; Ha, D.T.; Tran-Nguyen, C.H.; Nguyen, T.S.M.; Pham, T.T.N.; Nguyen, T.A.; Nguyen, D.A.; Hoang, H.Q.; Tran, N.V.; et al. Patterns of antimicrobial resistance in intensive care unit patients: A study in Vietnam. BMC Infect. Dis. 2017, 17, 429. [Google Scholar] [CrossRef]
- Van, P.H.; Binh, P.T.; Minh, N.H.L.; Morrissey, I.; Torumkuney, D. Results from the Survey of Antibiotic Resistance (SOAR) 2009-11 in Vietnam. J. Antimicrob. Chemother. 2016, 71, i93–i102. [Google Scholar] [CrossRef]
- Dat, V.Q.; Vu, H.N.; Nguyen The, H.; Nguyen, H.T.; Hoang, L.B.; Vu Tien Viet, D.; Bui, C.L.; Van Nguyen, K.; Nguyen, T.V.; Trinh, D.T.; et al. Bacterial bloodstream infections in a tertiary infectious diseases hospital in northern Vietnam: Aetiology, drug resistance, and treatment outcome. BMC Infect. Dis. 2017, 17, 493. [Google Scholar] [CrossRef]
- Mabberley, D.J. Mabberley’s Plant-Book, 3rd ed.; Cambridge University Press: Cambridge, UK, 2008; ISBN 978-0-521-82071-4. [Google Scholar]
- Kew Royal Botanic Gardens. The Plant List. Available online: http://www.theplantlist.org/tpl1.1/search?q=Zingiber (accessed on 21 July 2020).
- Sharifi-Rad, M.; Varoni, E.M.; Salehi, B.; Sharifi-Rad, J.; Matthews, K.R.; Ayatollahi, S.A.; Kobarfard, F.; Ibrahim, S.A.; Mnayer, D.; Zakaria, Z.A.; et al. Plants of the genus Zingiber as a source of bioactive phytochemicals: From tradition to pharmacy. Molecules 2017, 22, 2145. [Google Scholar] [CrossRef]
- Leong-Skornikova, J.; Newman, M. Gingers of Cambodia, Laos & Vietnam.; Oxford Graphic Printers Pte Ltd.: Singapore, 2015. [Google Scholar]
- Ly, N.-S. Zingiber skornickovae, a new species of Zingiberaceae from Central Vietnam. Phytotaxa 2016, 265, 139–144. [Google Scholar]
- Le, T.H.; Trinh, T.H.; Ly, N.S. Zingiber cornubracteatum Triboun & K. Larsen (Zingiberaceae), a new record for flora in Vietnam (in Vietnamese). J. Agric. Rural Dev. 2019, 21, 111–114. [Google Scholar]
- Triboun, P.; Larsen, K.; Chantaranothai, P. A key to the genus Zingiber (Zingiberaceae) in Thailand with descriptions of 10 new taxa. Thai J. Bot. 2014, 6, 53–77. [Google Scholar]
- Flora of China. Volume 24. Available online: http://www.efloras.org/flora_page.aspx?flora_id=2 (accessed on 14 July 2020).
- Nguyen, D.H.; Tran, M.H.; Ly, N.S.; Le, T.H.; Do, N.D. Zingiber neotruncatum T.L. Wu, K. Larsen & Turland, new record for flora in Vietnam (in Vietnamese). J. Sci. Nat. Sci. Technol. 2020, 36. in press. [Google Scholar]
- Newman, M.F. A new species of Zingiber (Zingiberaceae) from Lao P.D.R. Gard. Bull. Singap. 2015, 67, 123–127. [Google Scholar]
- Nguyen, V.H.; Le, T.H.; Do, N.D.; Ly, N.S.; Nguyen, T.T. A new record Zingiber nitens M. F. Newman (Zingiberaceae) for flora of Vietnam (in Vietnamese). J. Sci. Nat. Sci. Technol. 2017, 33, 46–50. [Google Scholar]
- Hung, N.V.; Dai, D.N.; Thai, T.H.; San, N.D.; Ogunwande, I.A. Zingiber nitens M.F. Newman: A new species and its essential oil constituent. J. Essent. Oil Bear. Plants 2017, 20, 69–75. [Google Scholar]
- Souvannakhoummane, K.; Leong-Skorničková, J. Eight new records of Zingiber Mill. (Zingiberaceae) for the flora of Lao P.D.R. Edinb. J. Bot. 2018, 75, 3–18. [Google Scholar]
- Ly, N.-S.; Dang, V.-S.; Do, D.-G.; Tran, T.-T.; Do, N.-D.; Nguyen, D.-H. Zingiber nudicarpum D. Fang (Zingiberaceae), a newly recorded species for Vietnam. Biosci. Discov. 2017, 8, 1–5. [Google Scholar]
- Hung, N.D.; Huong, L.T.; Sam, L.N.; Hoi, T.M.; Ogunwande, I.A. Constituents of essential oil of Zingiber nudicarpum from Vietnam. Chem. Nat. Compd. 2019, 55, 361–363. [Google Scholar]
- Ly, N.-S.; Truong, B.-V.; Le, T.H. Zingiber ottensii Valeton (Zingiberaceae)—A newly recorded species for Vietnam. Biosci. Discov. 2016, 7, 93–96. [Google Scholar]
- Aung, M.M.; Tanaka, N. Seven taxa of Zingiber (Zingiberaceae) newly recorded for the flora of Myanmar. Bull. Natl. Museum Nat. Sci. Ser. B Bot. 2019, 45, 1–8. [Google Scholar]
- Sirat, H.M.; Nordin, A.B. Essential oil of Zingiber ottensii Valeton. J. Essent. Oil Res. 1994, 6, 635–636. [Google Scholar]
- Malek, S.N.A.; Ibrahim, H.; Lai, H.S.; Serm, L.G.; Seng, C.K.; Yussoff, M.M.; Ali, N.A.M. Essential oils of Zingiber ottensii Valet. and Zingiber zerumbet (L.) Sm. from Sabah, Malaysia. Malays. J. Sci. 2005, 24, 49–58. [Google Scholar]
- Thubthimthed, S.; Limsiriwong, P.; Rerk-Am, U.; Suntorntanasat, T. Chemical composition and cytotoxic activity of the essential oil of Zingiber ottensii. Acta Hortic. 2005, 675, 107–109. [Google Scholar] [CrossRef]
- Theanphong, O.; Jenjittikul, T.; Mingvanish, W. Chemotaxonomic study of volatile oils from rhizomes of 9 Zingiber species (Zingiberaceae). Thai J. Bot. 2016, 8, 127–139. [Google Scholar]
- Marliani, L.; Subarnas, A.; Moelyono, M.W.; Halimah, E.; Pratiwi, F.W.; Suhardiman, A. Essential oil components of leaves and rhizome of Zingiber ottensii Val. from Bandung, Indonesia. Res. J. Chem. Environ. 2018, 22, 54–57. [Google Scholar]
- Tong, S.; Xia, Y. New taxa of Zingiberaceae from southern Yunnan. Acta Phytotaxon. Sin. 1987, 25, 460–471. [Google Scholar]
- Averyanov, L.V.; Nguyen, T.H.; Nguyen, S.K.; Nguyen, T.S.; Ngan, C.Q.; Maisak, T.V. Conservation Assessment of Engandered Lao-Vietnamese Stenoendemic—Pinus Cernua (Pinaceae); Mohamed bin Zayed Species Conservation Fund: Abu Dhabi, UAE, 2016. [Google Scholar]
- Nguyen, T.C.; Ly, N.S.; Le, T.H. Zingiber recurvatum, new record for flora in Vietnam (in Vietnamese). J. Agric. Rural Dev. 2020, in press. [Google Scholar]
- Le, T.-H.; Trinh, T.-H.; Do, N.-D.; Nguyen, V.-H.; Ly, N.-S. Zingiber vuquangense (Sect. Cryptanthium: Zingiberaceae): A new species from north central coast region, Vietnam. Phytotaxa 2019, 388, 295–300. [Google Scholar]
- Huong, L.T.; Huong, T.T.; Huong, N.T.T.; Chau, D.T.M.; Sam, L.N.; Ogunwande, I.A. Zingiber vuquangensis and Z. castaneum: Two newly discovered species from Vietnam and their essential oil constituents. Nat. Prod. Commun. 2018, 13, 763–766. [Google Scholar] [CrossRef]
- Ito, M.; Toyoda, M.; Honda, G. the essential oil of Perilla frutescens. Nat. Med. 1999, 53, 32–36. [Google Scholar]
- Mathela, C.S.; Melkani, A.B.; Bisht, J.C.; Pant, A.K.; Bestmann, H.J.; Erler, J.; Kobold, U.; Rauscher, J.; Vostrowsky, O. Chemical varieties of essential oils from Elsholtzia polystachya from two different locations in India. Planta Med. 1992, 58, 376–379. [Google Scholar]
- Gupta, S.; Pandotra, P.; Ram, G.; Anand, R.; Gupta, A.P.; Husain, M.K.; Bedia, Y.S.; Mallavarapu, G.R. Composition of a monoterpenoid-rich essential oil from the rhizome of Zingiber officinale from north western Himalayas. Nat. Prod. Commun. 2011, 6, 93–96. [Google Scholar] [CrossRef] [PubMed]
- Höferl, M.; Stoilova, I.; Wanner, J.; Schmidt, E.; Jirovetz, L.; Trifonova, D.; Stanchev, V.; Krastanov, A. Composition and comprehensive antioxidant activity of ginger (Zingiber officinale) essential oil from Ecuador. Nat. Prod. Commun. 2015, 10, 1085–1090. [Google Scholar] [CrossRef] [PubMed]
- Chane-Ming, J.; Vera, R.; Chalchat, J.-C. Chemical composition of the essential oil from rhizomes, leaves and flowers of Zingiber zerumbet Smith from Reunion Island. J. Essent. Oil Res. 2003, 15, 202–205. [Google Scholar]
- Dias, C.N.; Moraes, D.F.C. Essential oils and their compounds as Aedes aegypti L. (Diptera: Culicidae) larvicide: Review. Parasitol. Res. 2014, 113, 565–592. [Google Scholar] [CrossRef]
- Giatropoulos, A.; Papachristos, D.P.; Kimbaris, A.; Koliopoulos, G.; Polissiou, M.G.; Emmanouel, N.; Michaelakis, A. Evaluation of bioefficacy of three Citrus essential oils against the dengue vector Aedes albopictus (Diptera: Culicidae) in correlation to their components enantiomeric distribution. Parasitol. Res. 2012, 111, 2253–2263. [Google Scholar] [CrossRef]
- Andrade-Ochoa, S.; Sánchez-Aldana, D.; Chacón-Vargas, K.F.; Rivera-Chavira, B.E.; Sánchez-Torres, L.E.; Camacho, A.D.; Nogueda-Torres, B.; Nevárez-Moorillón, G.V. Oviposition deterrent and larvicidal and pupaecidal activity of seven essential oils and their major components against Culex quinquefasciatus Say (Diptera: Culicidae): Synergism—Antagonism effects. Insects 2018, 9, 25. [Google Scholar] [CrossRef]
- Govindarajan, M.; Rajeswary, M.; Hoti, S.L.; Bhattacharyya, A.; Benelli, G. Eugenol, α-pinene and β-caryophyllene from Plectranthus barbatus essential oil as eco-friendly larvicides against malaria, dengue and Japanese encephalitis mosquito vectors. Parasitol. Res. 2016, 115, 807–815. [Google Scholar] [CrossRef]
- Andrade-Ochoa, S.; Sánchez-Torres, L.E.; Nevárez-Moorillón, G.V.; Camacho, A.D.; Nogueda-Torres, B. Aceites esenciales y sus constituyentes como una alternativa en el control de mosquitos vectores de enfermedades. Biomedica 2017, 37, 224–243. [Google Scholar]
- Govindarajan, M. Chemical composition and larvicidal activity of leaf essential oil from Clausena anisata (Willd.) Hook. f. ex Benth (Rutaceae) against three mosquito species. Asian Pac. J. Trop. Med. 2010, 3, 874–877. [Google Scholar] [CrossRef]
- Andrade-Ochoa, S.; Correa-Basurto, J.; Rodríguez-Valdez, L.M.; Sánchez-Torres, L.E.; Nogueda-Torres, B. In vitro and in silico studies of terpenes, terpenoids and related compounds with larvicidal and pupaecidal activity against Culex quinquefasciatus Say (Diptera: Culicidae). Chem. Cent. J. 2018, 12, 53. [Google Scholar] [CrossRef] [PubMed]
- Govindarajan, M.; Rajeswary, M.; Arivoli, S.; Tennyson, S.; Benelli, G. Larvicidal and repellent potential of Zingiber nimmonii (J. Graham) Dalzell (Zingiberaceae) essential oil: An eco-friendly tool against malaria, dengue, and lymphatic filariasis mosquito vectors? Parasitol. Res. 2016, 115, 1807–1816. [Google Scholar] [CrossRef]
- Rajeswary, M.; Govindarajan, M.; Alharbi, N.S.; Kadaikunnan, S.; Khaled, J.M.; Benelli, G. Zingiber cernuum (Zingiberaceae) essential oil as effective larvicide and oviposition deterrent on six mosquito vectors, with little non-target toxicity on four aquatic mosquito predators. Environ. Sci. Pollut. Res. 2018, 25, 10307–10316. [Google Scholar] [CrossRef]
- Huong, L.T.; Chinh, H.V.; An, N.T.G.; Viet, N.T.; Hung, N.H.; Thuong, N.T.H.; Giwa-Ajeniya, A.O.; Ogunwande, I.A. Zingiber zerumbet rhizome essential oil: Chemical composition, antimicrobial and mosquito larvicidal activities. Eur. J. Med. Plants 2019, 30, 53652. [Google Scholar] [CrossRef]
- Pushpanathan, T.; Jebanesan, A.; Govindarajan, M. The essential oil of Zingiber officinalis Linn (Zingiberaceae) as a mosquito larvicidal and repellent agent against the filarial vector Culex quinquefasciatus Say (Diptera: Culicidae). Parasitol. Res. 2008, 102, 1289–1291. [Google Scholar] [CrossRef]
- Huong, L.T.; Huong, T.T.; Huong, N.T.T.; Hung, N.H.; Dat, P.T.T.; Luong, N.X.; Ogunwande, I.A. Mosquito larvicidal activity of the essential oil of Zingiber collinsii against Aedes albopictus and Culex quinquefasciatus. J. Oleo Sci. 2020, 69, 153–160. [Google Scholar]
- Smith-Palmer, A.; Stewart, J.; Fyfe, L. Antimicrobial properties of plant essential oils and essences against five important food-borne pathogens. Lett. Appl. Microbiol. 1998, 26, 118–122. [Google Scholar] [CrossRef]
- Izadi, Z.; Esna-Ashari, M.; Piri, K.; Davoodi, P. Chemical composition and antimicrobial activity of feverfew (Tanacetum parthenium) essential oil. Int. J. Agric. Biol. 2010, 12, 759–763. [Google Scholar]
- Djihane, B.; Wafa, N.; Elkhamssa, S.; Pedro, D.H.J.; Maria, A.E.; Mohamed Mihoub, Z. Chemical constituents of Helichrysum italicum (Roth) G. Don essential oil and their antimicrobial activity against Gram-positive and Gram-negative bacteria, filamentous fungi and Candida albicans. Saudi Pharm. J. 2017, 25, 780–787. [Google Scholar] [CrossRef] [PubMed]
- Inouye, S.; Takizawa, T.; Yamaguchi, H. Antibacterial activity of essential oils and their major constituents against respiratory tract pathogens by gaseous contact. J. Antimicrob. Chemother. 2001, 47, 565–573. [Google Scholar] [CrossRef] [PubMed]
- Nazzaro, F.; Fratianni, F.; De Martino, L.; Coppola, R.; De Feo, V. Effect of essential oils on pathogenic bacteria. Pharmaceuticals 2013, 6, 1451–1474. [Google Scholar] [CrossRef] [PubMed]
- van Zyl, R.L.; Seatlholo, S.T.; van Vuuren, S.F.; Viljoen, A.M. The biological activities of 20 nature identical essential oil constituents. J. Essent. Oil Res. 2006, 18, 129–133. [Google Scholar] [CrossRef]
- An, N.T.G.; Huong, L.T.; Satyal, P.; Tai, T.A.; Dai, D.N.; Hung, N.H.; Ngoc, N.T.B.; Setzer, W.N. Mosquito larvicidal activity, antimicrobial activity, and chemical compositions of essential oils from four species of Myrtaceae from central Vietnam. Plants 2020, 9, 544. [Google Scholar]
- De Carlo, A.; Zeng, T.; Dosoky, N.S.; Satyal, P.; Setzer, W.N. The essential oil composition and antimicrobial activity of Liquidambar formosana oleoresin. Plants 2020, 9, 822. [Google Scholar]
- Chung, N.T.; Huong, L.T.; Hung, N.H.; Hoi, T.M.; Dai, D.N. Chemical composition of Actinodaphne pilosa essential oil From Vietnam, mosquito larvicidal activity, and antimicrobial activity. Nat. Prod. Commun. 2020, 15, 1934578X20917792. [Google Scholar] [CrossRef]
- Adams, R.P. Identification of Essential Oil Components by Gas. Chromatography/Mass Spectrometry, 4th ed.; Allured Publishing: Carol Stream, IL, USA, 2007; ISBN 978-1-932633-21-4. [Google Scholar]
- Hung, N.H.; Satyal, P.; Do, N.D.; Tai, T.A.; Huong, L.T.; Chuong, N.T.H.; Hieu, H.V.; Tuan, P.A.; Van Vuong, P.; Setzer, W.N. Chemical compositions of Crassocephalum crepidioides essential oils and larvicidal activities against Aedes aegypti, Aedes albopictus, and Culex quinquefasciatus. Nat. Prod. Commun. 2019, 14, 1934578X19850033. [Google Scholar] [CrossRef]
- Finney, D. Probit Analysis, Reissue ed.; Cambridge University Press: Cambridge, UK, 2009; ISBN 978-0521135900. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Zingiber Species | Vietnamese Name | Collection Site Coordinates; Elevation | Collection Month Year | Voucher Number | Plant Part | % Yield (v/w) |
|---|---|---|---|---|---|---|
| Zingiber cornubraceatum Triboun & K. Larsen | Gừng lá bắc cựa | Khe Kèm water fall, Pù Mát National Park 19°58′14″ N, 104°48′20″ E; 280 m | September 2019 | 830 | Leaf Rhizome | 0.12 0.15 |
| Xuân Lý commune, Như Thanh District, Bến En National Park 19°39′09″ N, 105°30′24″ E; 80 m | October 2019 | 832 | Leaf Stems Rhizome | 0.14 0.10 0.17 | ||
| Tri Lễ commune, Quế Phong District, Pù Hoạt Nature Reserve 19°38′57″ N, 104°40′55″ E; 660 m | September 2018 | 735 | Leaf Rhizome | 0.10 0.12 | ||
| Zingiber neotruncatum T.L. Wu, K. Larsen & Turland | Gừng lá mới | Đồng Văn commune, Quế Phong district, Pù Hoạt Nature Reserve 19°53′58″ N, 105°07′45″ E; 315 m | October 2018 | 746 | Rhizome | 0.26 |
| Zingiber nitens M.F. Newman | Gừng lá sáng bóng | Vũ Quang National Park 18°17′13″ N, 105°26′12″ E; 180 m | September 2018 | 738 | Leaf Stems | 0.21 0.45 |
| Zingiber nudicarpum D.Fang | Gừng quả trần | Thông Thụ commune, Quế Phong district, Pù Hoạt Nature Reserve 19°49′5″ N, 104°55′35″ E; 460 m | April 2019 | 760 | Leaf Stems Rhizome | 0.12 0.08 0.15 |
| Bạch Mã National Park 16°13′44″ N, 107°53′54″ E; 350 m | April 2018 | 778 | Leaf Stems | 0.15 0.10 | ||
| Nam Đông District, Thừa Thiên Huế province Nam Đông District, Thừa Thiên Huế province 16°13′9″ N, 107°43′28″ E; 110 m | July 2019 | 777 | Leaf Stems Rhizome | 0.14 0.09 0.18 | ||
| Zingiber ottensii Valeton | Gừng ottensi | Tri Lễ commune, Quế Phong district, Pù Hoạt Nature Reserve 19°39′05″ N, 105°40′59″ E; 640 m | April 2019 | 772 | Leaf Roots | 0.18 0.25 |
| Zingiber recurvatum S.Q. Tong & Y.M. Xia | Gừng lá bắc cong | Tri Lễ commune, Quế Phong district, Pù Hoạt Nature Reserve 19°39′37″ N, 104°40′29″ E; 640 m | July 2019 | 792 | Leaf Rhizome | 0.14 0.19 |
| Zingiber vuquangensis Ly N.S., Le T.H., Trinh T.H., Nguyen V.H., Do N.D. | Gừng vũ quang | Nậm Nhong commune, Quế Phong district, Pù Hoạt Nature Reserve 19°30′31″ N, 104°43′13″ E; 590 m | April 2019 | 771 | Leaf Stems Rhizome | 0.15 0.10 0.14 |
| RIcalc | RIdb | Compounds | Z. cornubracteatum (Pù Hoạt) Leaf | Z. cornubracteatum (Pù Hoạt) Rhizome | Z. cornubracteatum (Pù Mát) Leaf | Z. cornubracteatum (Pù Mát) Rhizome | Z. cornubracteatum (Bến En) leaf | Z. Cornubracteatum (Bến En) Stem | Z. cornubracteatum (Bến En) Rhizome | Z. neotruncatum Rhizome | Z. nitens Leaf | Z. nitens Rhizome | Z. ottensii Leaf | Z. ottensii Rhizome | |
| 899 | 894 | 2-Heptanol | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | |
| 921 | 923 | Tricyclene | --- | --- | --- | --- | --- | --- | --- | --- | --- | 0.9 | --- | --- | |
| 925 | 927 | α-Thujene | --- | --- | --- | 2.1 | --- | --- | 2.2 | --- | --- | --- | --- | 0.9 | |
| 934 | 933 | α-Pinene | 5.1 | 14.5 | 2.7 | 8.2 | 10.1 | 9.9 | 9.8 | 1.9 | 11.2 | 10.5 | 4.0 | 3.4 | |
| 943 | 945 | α-Fenchene | --- | --- | --- | 0.3 | --- | --- | 0.3 | --- | --- | 0.9 | --- | --- | |
| 951 | 953 | Camphene | --- | 1.2 | --- | 3.2 | 0.3 | 0.3 | 1.3 | 0.2 | 0.3 | 40.4 | --- | 0.3 | |
| 952 | 953 | Thuja-2,4(10)diene | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | |
| 971 | 972 | Sabinene | 0.8 | 0.5 | 0.9 | 0.7 | 1.4 | 1.4 | 11.9 | 0.4 | 1.7 | 0.2 | 0.4 | 21.6 | |
| 978 | 978 | β-Pinene | 20.1 | 14.8 | 18.8 | 8.8 | 67.3 | 66.8 | 33.1 | 3.1 | 6.0 | 1.1 | 17.1 | 11.7 | |
| 981 | 985 | 6-Methylhept-5-en-2-one | --- | --- | --- | --- | --- | --- | --- | 0.3 | --- | --- | --- | --- | |
| 988 | 991 | Myrcene | 0.4 | 0.4 | 1.8 | 7.7 | 1.0 | 1.0 | 4.2 | 0.3 | 0.4 | 2.0 | 0.4 | 1.3 | |
| 989 | 988 | Dehydro-1,8-cineole | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | |
| 989 | 984 | 2-Pentylfuran | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | |
| 1002 | 998 | Octanal | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | |
| 1004 | 1108 | 1,3,8-p-Menthatriene | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | |
| 1006 | 1007 | α-Phellandrene | 0.8 | 0.4 | 1.1 | 0.4 | --- | --- | 1.6 | --- | --- | 0.2 | --- | 0.6 | |
| 1008 | 1009 | δ-3-Carene | 0.2 | 0.1 | 0.2 | 5.3 | --- | --- | 2.1 | --- | --- | --- | --- | --- | |
| 1016 | 1018 | α-Terpinene | --- | --- | 0.2 | 0.2 | --- | --- | 0.8 | --- | --- | --- | --- | 3.5 | |
| 1018 | 1022 | m-Cymene | --- | --- | --- | 0.1 | --- | --- | --- | --- | --- | --- | --- | --- | |
| 1023 | 1025 | p-Cymene | 0.4 | 0.5 | 0.3 | 3.9 | --- | --- | 2.7 | --- | --- | --- | --- | 2.1 | |
| 1034 | 1028 | Limonene | 0.6 | 1.0 | 1.2 | 5.1 | 1.2 | 1.2 | 3.4 | 0.2 | 0.9 | 4.6 | 0.8 | 0.9 | |
| 1035 | 1031 | β-Phellandrene | 0.5 | 0.9 | 0.8 | 2.2 | 0.3 | 0.3 | 0.5 | --- | 0.2 | --- | 0.2 | 0.3 | |
| 1035 | 1030 | 1,8-Cineole | --- | 10.4 | --- | 6.1 | 0.2 | 0.2 | 2.5 | 0.7 | --- | --- | --- | 4.3 | |
| 1036 | 1034 | (Z)-β-Ocimene | 0.3 | --- | --- | --- | --- | --- | --- | --- | 0.2 | 0.9 | 0.3 | --- | |
| 1046 | 1045 | (E)-β-Ocimene | 0.6 | 0.4 | 0.4 | 0.4 | 0.1 | 0.1 | 0.8 | 1.2 | 2.1 | 12.7 | 2.1 | 0.9 | |
| 1057 | 1057 | γ-Terpinene | 0.3 | 0.4 | 0.4 | 0.7 | 0.2 | 0.2 | 2.4 | --- | --- | 0.1 | --- | 5.5 | |
| 1067 | 1063 | 1-Octanol | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | |
| 1068 | 1067 | cis-Linalool oxide (furanoid) | --- | 0.3 | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | |
| 1072 | 1065 | cis-Sabinene hydrate | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | 0.3 | |
| 1084 | 1086 | Terpinolene | 0.1 | --- | 0.3 | --- | --- | --- | 0.7 | --- | --- | 0.8 | --- | 1.1 | |
| 1084 | 1084 | trans-Linalool oxide (furanoid) | --- | 0.3 | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | |
| 1087 | 1083 | Fenchone | --- | --- | --- | --- | --- | --- | --- | --- | --- | 0.2 | --- | --- | |
| 1088 | 1089 | p-Cymenene | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | |
| 1099 | 1102 | Perillene | --- | --- | --- | 0.3 | 0.1 | 0.1 | --- | 51.3 | --- | --- | --- | --- | |
| 1099 | 1097 | 2-Nonanol | --- | --- | --- | 0.3 | --- | --- | --- | --- | --- | --- | --- | --- | |
| 1100 | 1101 | Linalool | 0.5 | 31.0 | - | 11.6 | 0.2 | 0.2 | 0.4 | --- | 0.4 | --- | --- | 0.2 | |
| 1100 | 1104 | Rosefuran | --- | --- | --- | --- | --- | --- | --- | 0.2 | --- | --- | --- | --- | |
| 1104 | 1098 | trans-Sabinene hydrate | --- | 0.3 | --- | --- | --- | --- | --- | --- | --- | --- | --- | 0.2 | |
| 1105 | 1107 | Nonanal | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | |
| 1112 | 1116 | (E)-4,8-Dimethylnona-1,3,7-triene | --- | --- | 0.1 | 0.2 | --- | --- | --- | --- | --- | --- | --- | --- | |
| 1118 | 1114 | endo-Fenchol | --- | --- | --- | 0.3 | --- | --- | 0.2 | --- | --- | --- | --- | --- | |
| 1118 | 1118 | cis-p-Menth-2-en-1-ol | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | 0.4 | |
| 1120 | 1128 | allo-Ocimene | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | |
| 1124 | 1118 | exo-Fenchol | --- | 0.3 | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | |
| 1128 | 1122 | α-Campholenal | --- | --- | --- | 0.1 | --- | --- | --- | --- | --- | --- | --- | --- | |
| 1135 | 1136 | trans-p-Menth-2-en-1-ol | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | 0.3 | |
| 1137 | 1135 | Nopinone | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | |
| 1141 | 1135 | trans-Pinocarveol | --- | --- | --- | --- | --- | --- | 0.2 | --- | --- | --- | --- | --- | |
| 1144 | 1140 | trans-Verbenol | --- | 0.8 | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | |
| 1146 | 1149 | Camphor | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | |
| 1148 | 1165 | Lavandulol | --- | --- | --- | --- | --- | --- | --- | 0.2 | --- | --- | --- | --- | |
| 1159 | 1167 | Benzyl acetate | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | |
| 1162 | 1160 | Pinocarvone | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | |
| 1167 | 1165 | iso-Neral | --- | --- | --- | --- | --- | --- | --- | 0.7 | --- | --- | --- | --- | |
| 1169 | 1162 | δ-Terpineol | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | |
| 1170 | 1166 | p-Mentha-1,5-dien-8-ol | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | |
| 1171 | 1165 | Borneol | --- | 3.2 | --- | 0.7 | --- | --- | 0.1 | --- | --- | 0.8 | --- | 0.5 | |
| 1180 | 1180 | Terpinen-4-ol | 0.1 | 0.7 | --- | 0.5 | --- | --- | 1.4 | 0.1 | --- | --- | --- | 17.1 | |
| 1185 | 1179 | p-Cymen-8-ol | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | |
| 1185 | 1185 | iso-Geranial | --- | --- | --- | --- | --- | --- | --- | 1.1 | --- | --- | --- | --- | |
| 1194 | 1195 | α-Terpineol | --- | 0.6 | --- | 0.2 | --- | --- | 0.1 | --- | --- | --- | --- | 0.9 | |
| 1194 | 1195 | Myrtenal | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | |
| 1194 | 1193 | (4Z)-Decenal | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | |
| 1195 | 1194 | Myrtenol | 1.0 | 0.2 | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | |
| 1198 | 1195 | cis-Piperitol | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | 0.1 | |
| 1205 | 1204 | Verbenone | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | |
| 1206 | 1206 | Decanal | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | |
| 1209 | 1207 | trans-Piperitol | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | 0.2 | |
| 1216 | 1218 | endo-Fenchyl acetate | --- | --- | --- | 0.6 | --- | --- | 0.5 | --- | --- | 3.3 | --- | --- | |
| 1217 | 1215 | trans-Carveol | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | |
| 1220 | 1217 | β-Cyclocitral | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | |
| 1222 | 1227 | Nerol | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | |
| 1224 | 1223 | Citronellol | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | |
| 1227 | 1232 | Thymyl methyl ether | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | |
| 1236 | 1235 | Neral | --- | --- | --- | --- | --- | --- | --- | 12.3 | --- | --- | --- | --- | |
| 1241 | 1238 | Cuminal | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | |
| 1242 | 1239 | Carvone | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | |
| 1248 | 1249 | Geraniol | --- | --- | --- | 0.2 | --- | --- | --- | 0.3 | --- | --- | --- | --- | |
| 1265 | 1264 | Geranial | --- | --- | --- | --- | --- | --- | --- | 17.0 | --- | --- | --- | --- | |
| 1271 | 1266 | 1-Decanol | --- | --- | --- | 0.2 | --- | --- | --- | --- | --- | --- | --- | --- | |
| 1281 | 1287 | iso-Bornyl acetate | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | |
| 1284 | 1285 | Bornyl acetate | 0.2 | 2.8 | --- | 7.9 | --- | --- | 2.5 | --- | --- | 14.5 | --- | 0.3 | |
| 1285 | 1291 | Safrole | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | |
| 1293 | 1290 | Dihydroedulan IIA | --- | --- | --- | --- | --- | 0.2 | --- | --- | --- | --- | 0.3 | --- | |
| 1295 | 1298 | Carvacrol | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | |
| 1295 | 1293 | Methyl myrtenate | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | |
| 1307 | 1295 | trans-Sabinyl acetate | --- | --- | --- | --- | --- | --- | 0.2 | --- | --- | --- | --- | --- | |
| 1330 | 1324 | Methyl geranate | --- | --- | --- | 0.3 | --- | --- | --- | 0.2 | --- | --- | --- | --- | |
| 1330 | 1332 | Bicycloelemene | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | |
| 1330 | 1335 | δ-Elemene | 1.4 | --- | 1.5 | --- | 0.4 | 0.5 | --- | --- | --- | --- | 0.2 | --- | |
| 1333 | 1325 | Myrtenyl acetate | 0.1 | 0.5 | --- | 2.0 | --- | --- | 0.7 | --- | --- | --- | --- | --- | |
| 1342 | 1349 | α-Cubebene | 0.1 | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | |
| 1345 | 1346 | α-Terpinyl acetate | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | |
| 1364 | 1367 | Cyclosativene | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | |
| 1364 | 1372 | iso-Ledene | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | |
| 1368 | 1373 | α-Ylangene | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | |
| 1374 | 1375 | α-Copaene | 0.2 | --- | 0.4 | --- | --- | --- | --- | --- | --- | --- | 3.2 | --- | |
| 1375 | 1380 | Geranyl acetate | --- | --- | --- | --- | --- | --- | --- | 0.6 | --- | --- | --- | --- | |
| 1377 | 1383 | cis-β-Elemene | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | |
| 1377 | 1365 | Eugenol | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | |
| 1382 | 1382 | β-Bourbonene | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | |
| 1386 | 1387 | β-Cubebene | 0.4 | --- | 0.5 | --- | --- | --- | --- | --- | --- | --- | --- | --- | |
| 1387 | 1390 | trans-β-Elemene | 0.2 | --- | 0.5 | --- | 3.9 | 4.4 | 0.7 | --- | 0.8 | 0.2 | 17.0 | 0.3 | |
| 1400 | 1403 | Methyl eugenol | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | |
| 1403 | 1405 | Sesquithujene | --- | --- | --- | --- | --- | --- | --- | --- | 0.1 | --- | --- | --- | |
| 1403 | 1408 | (Z)-Caryophyllene | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | |
| 1405 | 1406 | α-Gurjunene | --- | --- | 0.3 | --- | --- | --- | 0.2 | --- | --- | --- | 0.6 | --- | |
| 1405 | 1415 | β-Maaliene | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | |
| 1415 | 1416 | cis-α-Bergamotene | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | |
| 1419 | 1417 | (E)-Caryophyllene | 8.9 | 0.4 | 13.9 | 1.7 | 1.8 | 2.1 | 0.6 | 1.5 | --- | --- | 28.0 | 0.7 | |
| 1427 | 1432 | γ-Elemene | 0.3 | --- | --- | --- | 0.2 | 0.3 | 0.1 | --- | --- | --- | --- | --- | |
| 1428 | 1424 | 2,5-Dimethoxy-p-cymene | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | |
| 1428 | 1430 | β-Copaene | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | |
| 1428 | 1438 | α-Maaliene | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | |
| 1431 | 1416 | α-Santalene | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | |
| 1431 | 1432 | trans-α-Bergamotene | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | |
| 1433 | 1437 | α-Guaiene | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | |
| 1437 | 1438 | Aromadendrene | 0.3 | --- | 0.3 | --- | 0.2 | --- | --- | --- | --- | --- | 0.1 | --- | |
| 1440 | 1440 | (Z)-β-Farnesene | --- | --- | --- | --- | --- | --- | --- | --- | 0.3 | --- | --- | --- | |
| 1444 | 1431 | β-Gurjunene | 0.3 | --- | 0.6 | --- | --- | --- | --- | --- | --- | --- | --- | --- | |
| 1445 | 1445 | Myltayl-4(12)-ene | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | |
| 1446 | 1453 | Geranyl acetone | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | |
| 1447 | 1453 | trans-Muurola-3,5-diene | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | |
| 1450 | 1452 | (E)-β-Farnesene | --- | --- | --- | --- | --- | --- | --- | --- | 0.2 | --- | --- | --- | |
| 1452 | 1457 | Prezizaene | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | |
| 1454 | 1454 | α-Humulene | 0.7 | --- | 1.0 | 0.3 | 0.2 | 0.3 | --- | --- | --- | --- | 3.0 | 3.8 | |
| 1458 | 1457 | allo-Aromadendrene | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | |
| 1460 | 1461 | cis-Cadina-1(6),4-diene | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | |
| 1470 | 1472 | trans-Cadina-1(6),4-diene | --- | --- | 0.7 | --- | --- | --- | --- | --- | --- | --- | --- | --- | |
| 1471 | 1476 | β-Chamigrene | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | |
| 1471 | 1475 | γ-Gurjunene | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | |
| 1472 | 1482 | γ-Curcumene | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | |
| 1473 | 1478 | γ-Muurolene | 0.3 | --- | 0.5 | 0.1 | --- | --- | --- | --- | --- | --- | --- | --- | |
| 1474 | 1476 | Selina-4,11-diene | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | |
| 1477 | 1482 | α-Amorphene | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | |
| 1477 | 1479 | ar-Curcumene | --- | --- | --- | --- | --- | --- | --- | --- | 5.2 | 0.2 | --- | --- | |
| 1477 | 1487 | (E)-β-Ionone | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | |
| 1478 | 1483 | trans-β-Bergamotene | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | |
| 1479 | 1464 | 9-epi-(E)-Caryophyllene | 0.4 | --- | 0.8 | --- | 0.1 | 0.2 | --- | --- | 0.2 | --- | 0.2 | --- | |
| 1480 | 1480 | Germacrene D | 1.6 | --- | 13.7 | 0.8 | 0.7 | 0.8 | 0.2 | --- | 0.7 | --- | 2.5 | --- | |
| 1481 | 1488 | δ-Selinene | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | |
| 1482 | 1492 | cis-β-Guaiene | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | |
| 1483 | 1487 | Aristolochene | --- | --- | --- | --- | --- | --- | --- | --- | --- | 0.3 | --- | --- | |
| 1488 | 1492 | β-Selinene | 0.3 | --- | 0.4 | --- | 0.5 | 0.6 | --- | --- | --- | --- | 0.8 | --- | |
| 1489 | 1496 | Valencene | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | 0.4 | --- | |
| 1490 | 1490 | γ-Amorphene | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | |
| 1491 | 1493 | α-Zingiberene | --- | --- | --- | --- | --- | --- | --- | --- | 17.4 | 1.6 | --- | --- | |
| 1494 | 1498 | α-Selinene | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | |
| 1494 | 1497 | Bicyclogermacrene | 7.4 | --- | 18.9 | 0.5 | 2.7 | 3.0 | 0.4 | 0.9 | 1.5 | --- | 1.2 | --- | |
| 1496 | 1500 | α-Muurolene | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | |
| 1496 | 1506 | α-Bulnesene | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | |
| 1497 | 1502 | trans-β-Guiaene | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | |
| 1500 | 1500 | n-Pentadecane | --- | --- | --- | --- | --- | --- | --- | 0.2 | --- | --- | --- | --- | |
| 1501 | 1505 | (E,E)-α-Farnesene | --- | --- | 0.9 | 0.2 | 0.5 | 0.6 | 0.2 | --- | --- | --- | 1.1 | --- | |
| 1501 | 1495 | γ-Amorphene | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | |
| 1502 | 1496 | Viridiflorene | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | |
| 1503 | 1509 | β-Curcumene | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | |
| 1506 | 1508 | β-Bisabolene | --- | --- | --- | 0.2 | --- | --- | 0.2 | 1.2 | 3.7 | 0.2 | --- | --- | |
| 1511 | 1512 | γ-Cadinene | 0.3 | --- | 0.5 | --- | --- | 0.2 | --- | --- | --- | --- | --- | --- | |
| 1511 | 1511 | δ-Amorphene | --- | --- | 0.4 | --- | 0.2 | 0.2 | --- | --- | --- | --- | --- | --- | |
| 1513 | 1519 | Cubebol | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | |
| 1516 | 1518 | δ-Cadinene | 0.6 | - | 0.9 | 0.2 | 0.2 | 0.2 | --- | --- | --- | --- | 2.2 | --- | |
| 1517 | 1520 | 7-epi-α-Selinene | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | |
| 1517 | 1521 | Zonarene | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | |
| 1519 | 1521 | trans-Calamenene | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | |
| 1520 | 1528 | (E)-γ-Bisabolene | --- | --- | --- | --- | --- | --- | --- | --- | 0.2 | --- | --- | --- | |
| 1523 | 1521 | Eugenyl acetate | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | 0.2 | 0.1 | |
| 1524 | 1521 | β-Sesquiphellandrene | --- | --- | --- | --- | --- | --- | --- | --- | 10.1 | 0.6 | --- | --- | |
| 1529 | 1528 | cis-Calamene | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | |
| 1530 | 1536 | trans-Cadina-1,4-diene | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | |
| 1535 | 1537 | α-Cadinene | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | |
| 1539 | 1544 | α-Calacorene | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | |
| 1546 | 1549 | α-Elemol | 0.2 | --- | 0.2 | --- | --- | --- | 0.2 | --- | --- | --- | 0.2 | 0.4 | |
| 1557 | 1559 | Germacrene B | 0.3 | --- | 0.5 | --- | 0.2 | 0.3 | 0.3 | --- | --- | --- | --- | --- | |
| 1558 | 1561 | (E)-Nerolidol | 23.0 | --- | 2.7 | 8.4 | 0.9 | 1.1 | 0.3 | 0.1 | 10.0 | 1.1 | 0.5 | --- | |
| 1560 | 1564 | β-Calacorene | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | |
| 1564 | 1568 | Palustrol | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | |
| 1568 | 1571 | Maaliol | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | |
| 1570 | 1573 | (3E,7E)-4,8,12-Trimethyltrideca-1,3,7,11-tetraene | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | |
| 1575 | 1576 | Spathulenol | 2.5 | --- | 1.4 | --- | 0.1 | 0.2 | --- | 0.4 | 0.6 | --- | 0.2 | --- | |
| 1575 | 1582 | Neryl isovalerate | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | |
| 1575 | 1574 | Germacrene D-4-ol | 0.2 | --- | --- | --- | --- | --- | --- | --- | 0.9 | --- | --- | --- | |
| 1582 | 1577 | Caryophyllene oxide | 2.2 | --- | 2.0 | 0.6 | --- | 0.2 | 0.3 | 0.3 | --- | 0.1 | 1.2 | 0.4 | |
| 1582 | 1582 | ar-Turmerol | --- | --- | --- | --- | --- | --- | --- | --- | 0.2 | --- | --- | --- | |
| 1583 | 1570 | Dendrolasin | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | |
| 1583 | 1585 | epi-Globulol | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | |
| 1587 | 1590 | Globulol | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | |
| 1590 | 1581 | Clovenol | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | |
| 1592 | 1594 | Salvial-4(14)en-1-one | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | |
| 1593 | 1592 | Viridiflorol | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | |
| 1593 | 1593 | Scapanol g | --- | --- | 0.3 | --- | --- | --- | --- | --- | --- | --- | --- | --- | |
| 1594 | 1600 | Guaiol | 0.5 | --- | 0.2 | --- | 0.1 | 0.2 | 0.3 | --- | --- | --- | --- | --- | |
| 1594 | 1601 | trans-β-Elemenone | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | |
| 1596 | 1599 | Cubeban-11-ol | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | |
| 1599 | 1592 | Humulene epoxide I | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | 1.2 | |
| 1600 | 1609 | Rosifoliol | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | |
| 1604 | 1607 | 5,7-di-epi-Eudesmol | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | |
| 1604 | 1602 | Ledol | 0.3 | --- | 0.3 | --- | --- | --- | --- | --- | --- | --- | --- | --- | |
| 1613 | 1618 | 1,10-di-epi-Cubenol | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | |
| 1616 | 1613 | Humulene epoxide II | --- | --- | --- | 0.2 | --- | --- | --- | --- | --- | --- | --- | 0.6 | |
| 1616 | 1611 | Zingiberenol | --- | --- | --- | --- | --- | --- | --- | --- | 7.2 | 0.2 | --- | --- | |
| 1618 | 1623 | Humulane-1,6-dien-3-ol | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | |
| 1620 | 1622 | 10-epi-γ-Eudesmol | --- | --- | 0.3 | --- | --- | --- | --- | --- | --- | --- | --- | --- | |
| 1625 | 1631 | 1-epi-Cubenol | --- | --- | 0.2 | --- | --- | --- | --- | --- | --- | --- | --- | --- | |
| 1627 | 1618 | epi-Cedrol | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | |
| 1628 | 1622 | Alismol | 0.4 | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | |
| 1630 | 1630 | γ-Eudesmol | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | 0.2 | |
| 1632 | 1631 | Caryophylla-4(12),8(13)-dien-5α-ol | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | |
| 1634 | 1632 | α-Acorenol | --- | --- | --- | --- | --- | --- | --- | --- | 3.3 | 0.2 | --- | --- | |
| 1636 | 1639 | Caryophylla-4(12),8(13)-dien-5β-ol | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | |
| 1640 | 1638 | τ-Cadinol | 0.9 | --- | 0.2 | --- | --- | --- | --- | --- | --- | --- | --- | --- | |
| 1640 | 1643 | Cubenol | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | |
| 1642 | 1644 | τ-Murrolol | --- | --- | 0.4 | --- | --- | --- | --- | --- | --- | --- | --- | --- | |
| 1645 | 1644 | α-Muurolol (=δ-Cadinol) | --- | --- | 0.2 | --- | --- | --- | --- | --- | --- | --- | --- | --- | |
| 1651 | 1649 | β-Eudesmol | --- | --- | 0.3 | 0.3 | --- | --- | --- | --- | --- | --- | --- | 0.3 | |
| 1652 | 1652 | α-Cadinol | 0.5 | --- | 1.0 | --- | --- | 0.1 | --- | --- | 0.4 | --- | 0.3 | --- | |
| 1652 | 1647 | 1,2-Diacetoxy-4-allylbenzene | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | |
| 1653 | 1652 | α-Eudesmol | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | 0.3 | |
| 1657 | 1658 | Selin-11-en-4α-ol | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | |
| 1662 | 1658 | neo-Intermedeol | 0.5 | --- | --- | --- | --- | 0.1 | - | --- | 0.7 | 0.2 | 0.2 | --- | |
| 1668 | 1668 | 14-Hydroxy-9-epi-(E)-caryophyllene | --- | --- | 0.2 | --- | --- | --- | --- | --- | --- | --- | --- | --- | |
| 1670 | 1675 | Cadalene | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | |
| 1671 | 1676 | Mustakone | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | |
| 1679 | 1681 | γ-Bicyclofarnesal | --- | --- | --- | 0.5 | --- | --- | 0.5 | --- | --- | --- | --- | --- | |
| 1681 | 1685 | α-Bisabolol | --- | --- | --- | --- | --- | --- | --- | --- | 4.1 | --- | --- | --- | |
| 1684 | 1685 | Germacra-4(15),5,10(14)-trien-1α-ol | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | |
| 1684 | 1683 | epi-α-Bisabolol | --- | --- | --- | 0.1 | --- | --- | --- | --- | --- | --- | --- | --- | |
| 1690 | 1693 | Germacrone | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | |
| 1691 | 1694 | Germacra-4(15),5,10(14)-trien-1β-ol | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | |
| 1696 | 1689 | 6α-Hydroxygermacra-1(10),4-diene | --- | --- | --- | --- | --- | --- | --- | --- | 0.7 | --- | --- | --- | |
| 1701 | 1698 | (2Z,6Z)-Farnesol | --- | --- | --- | --- | --- | --- | --- | --- | 0.9 | --- | --- | --- | |
| 1702 | 1690 | (Z)-trans-α-Bergamotol | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | |
| 1707 | 1714 | Nootkatol | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | |
| 1713 | 1713 | Pentadecanal | 0.2 | --- | --- | --- | --- | --- | 0.2 | --- | --- | --- | --- | --- | |
| 1738 | 1740 | Mint sulfide | 1.5 | --- | 1.9 | --- | --- | 0.1 | --- | --- | --- | --- | --- | --- | |
| 1739 | 1732 | Zerumbone | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | 12.5 | |
| 1761 | 1759 | Benzyl benzoate | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | |
| 1774 | 1760 | (Z)-Lanceol | --- | --- | --- | --- | --- | --- | --- | 0.3 | --- | --- | --- | --- | |
| 1799 | 1809 | Ambrial | 0.1 | --- | --- | 0.9 | --- | --- | 3.6 | --- | --- | --- | 0.3 | --- | |
| 1838 | 1838 | Phytone | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | |
| 1848 | 1845 | 6,10,14-Trimethylpentadecan-2-one | 1.1 | --- | --- | --- | 0.1 | 0.2 | --- | --- | --- | --- | --- | --- | |
| 1862 | 1864 | Benzyl salicylate | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | |
| 2002 | --- | (E)-15,16-Bisnorlabda-8(17),11-dien-13-one | --- | --- | --- | 0.3 | --- | --- | 0.2 | --- | --- | --- | --- | --- | |
| 2104 | 2105 | (E)-Phytol | --- | --- | 0.3 | --- | 0.3 | 0.4 | --- | --- | --- | --- | 0.2 | --- | |
| Monoterpene hydrocarbons | 30.2 | 35.1 | 29.1 | 49.3 | 81.9 | 81.2 | 77.8 | 7.3 | 23.0 | 75.3 | 25.3 | 54.1 | |||
| Oxygenated monoterpenoids | 1.9 | 51.4 | 0.0 | 30.8 | 0.5 | 0.5 | 8.8 | 84.7 | 0.4 | 18.8 | 0.0 | 24.8 | |||
| Sesquiterpene hydrocarbons | 24.0 | 0.4 | 57.2 | 4.0 | 11.8 | 13.7 | 2.9 | 3.6 | 40.4 | 3.1 | 60.5 | 4.8 | |||
| Oxygenated sesquiterpenoids | 31.2 | 0.0 | 9.9 | 10.1 | 1.1 | 1.9 | 1.6 | 1.1 | 29.0 | 1.8 | 2.6 | 15.9 | |||
| Others | 2.9 | 0.0 | 2.3 | 1.9 | 0.4 | 0.9 | 4.0 | 0.5 | 0.0 | 0.0 | 1.0 | 0.1 | |||
| Total identified | 90.2 | 86.9 | 98.5 | 96.1 | 95.7 | 98.2 | 95.1 | 97.2 | 92.8 | 99.0 | 89.4 | 99.7 | |||
| RIcalc | RIdb | Compounds | Z. nudicarpum (Pù Hoạt) leaf | Z. nudicarpum (Pù Hoạt) stem | Z. nudicarpum (Pù Hoạt) rhizome | Z. nudicarpum (Nam Đông) leaf | Z. nudicarpum (Nam Đông) stem | Z. nudicarpum (Nam Đông) rhizome | Z. nudicarpum (Bạch Mã) leaf | Z. nudicarpum (Bạch Mã) stem | Z. recurvatumleaf | Z. recurvatum rhizome | Z. vuquangensis leaf | Z. vuquangensis stem | Z. vuquangensis rhizome |
| 899 | 894 | 2-Heptanol | --- | --- | --- | --- | 0.2 | --- | --- | --- | --- | --- | --- | --- | --- |
| 921 | 923 | Tricyclene | --- | --- | --- | --- | --- | tr | tr | tr | --- | --- | --- | --- | --- |
| 925 | 927 | α-Thujene | 0.1 | --- | 0.1 | 0.8 | 0.1 | tr | 0.1 | tr | 0.3 | 0.3 | --- | --- | --- |
| 934 | 933 | α-Pinene | 10.9 | --- | 18.7 | 5.0 | 10.6 | 4.0 | 6.5 | 6.1 | 16.3 | 1.9 | 11.3 | 0.5 | 9.8 |
| 943 | 945 | α-Fenchene | --- | --- | --- | --- | --- | --- | --- | --- | 0.2 | --- | --- | --- | --- |
| 951 | 953 | Camphene | 1.6 | --- | 1.3 | 0.1 | 0.3 | 2.1 | 0.6 | 0.8 | 0.4 | 1.9 | 0.2 | --- | 0.3 |
| 952 | 953 | Thuja-2,4(10)diene | --- | --- | --- | tr | 0.2 | --- | --- | --- | --- | --- | --- | --- | --- |
| 971 | 972 | Sabinene | --- | --- | 1.4 | 2.3 | 0.1 | tr | 0.2 | 0.1 | 1.2 | 0.3 | 1.0 | --- | 0.7 |
| 978 | 978 | β-Pinene | 34.0 | 0.5 | 58.3 | 26.6 | 9.0 | 9.8 | 0.7 | 5.6 | 71.6 | 4.7 | 38.5 | 3.1 | 45.0 |
| 981 | 985 | 6-Methylhept-5-en-2-one | --- | --- | --- | --- | 0.1 | --- | --- | --- | --- | --- | --- | --- | --- |
| 988 | 991 | Myrcene | 0.8 | --- | 0.9 | 0.8 | 0.4 | 0.6 | 0.7 | 1.2 | 1.0 | 2.1 | 0.5 | --- | 0.7 |
| 989 | 988 | Dehydro-1,8-cineole | --- | --- | --- | --- | 0.1 | --- | --- | --- | --- | --- | --- | --- | --- |
| 989 | 984 | 2-Pentylfuran | --- | --- | --- | tr | --- | --- | --- | --- | --- | --- | --- | --- | --- |
| 1002 | 998 | Octanal | --- | --- | --- | --- | 0.1 | --- | --- | --- | --- | --- | --- | --- | --- |
| 1004 | 1108 | 1,3,8-p-Menthatriene | --- | --- | --- | --- | 0.1 | --- | --- | --- | --- | --- | --- | --- | --- |
| 1006 | 1007 | α-Phellandrene | --- | --- | --- | tr | 0.1 | 0.2 | 0.9 | 0.6 | 0.2 | 0.6 | --- | --- | --- |
| 1008 | 1009 | δ-3-Carene | --- | --- | 0.2 | --- | 0.2 | 3.1 | 3.7 | 1.8 | --- | 1.5 | --- | --- | --- |
| 1016 | 1018 | α-Terpinene | 0.1 | --- | --- | 0.2 | tr | tr | 0.1 | tr | --- | --- | --- | --- | 0.1 |
| 1018 | 1022 | m-Cymene | --- | --- | --- | --- | tr | tr | --- | --- | --- | --- | --- | --- | --- |
| 1023 | 1025 | p-Cymene | 0.2 | --- | 0.4 | 0.4 | 6.0 | 0.9 | 0.2 | 0.1 | --- | 0.6 | --- | --- | 0.2 |
| 1034 | 1028 | Limonene | 1.5 | --- | 1.8 | 2.0 | 2.1 | 1.8 | 1.7 | 6.0 | 1.8 | 2.4 | 0.9 | --- | 1.9 |
| 1035 | 1031 | β-Phellandrene | 0.2 | --- | 0.4 | 0.2 | --- | 0.7 | 0.3 | 0.3 | 0.3 | 0.6 | 0.3 | --- | 2.3 |
| 1035 | 1030 | 1,8-Cineole | 0.9 | --- | 0.3 | 0.9 | 21.2 | 6.8 | tr | tr | --- | 0.6 | --- | --- | --- |
| 1036 | 1034 | (Z)-β-Ocimene | --- | --- | --- | tr | --- | --- | 0.1 | 0.2 | 0.2 | --- | --- | --- | --- |
| 1046 | 1045 | (E)-β-Ocimene | --- | --- | 0.7 | tr | 0.1 | --- | 0.1 | 0.2 | 0.6 | 0.4 | --- | --- | --- |
| 1057 | 1057 | γ-Terpinene | 0.5 | --- | 0.3 | 0.8 | 0.1 | 0.4 | 0.1 | 0.1 | 0.1 | 0.6 | --- | --- | 0.4 |
| 1067 | 1063 | 1-Octanol | --- | --- | --- | --- | 0.3 | --- | --- | --- | --- | --- | --- | --- | --- |
| 1068 | 1067 | cis-Linalool oxide (furanoid) | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- |
| 1072 | 1065 | cis-Sabinene hydrate | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- |
| 1084 | 1086 | Terpinolene | 0.1 | --- | 0.3 | 0.2 | 0.1 | 0.3 | 0.1 | 0.1 | --- | 0.4 | --- | --- | 0.4 |
| 1084 | 1084 | trans-Linalool oxide (furanoid) | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- |
| 1087 | 1083 | Fenchone | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- |
| 1088 | 1089 | p-Cymenene | --- | --- | --- | --- | 0.1 | --- | --- | --- | --- | --- | --- | --- | --- |
| 1099 | 1102 | Perillene | --- | --- | --- | --- | 0.1 | --- | --- | --- | --- | --- | --- | --- | --- |
| 1099 | 1097 | 2-Nonanol | --- | --- | --- | --- | 0.1 | --- | --- | --- | --- | --- | --- | --- | --- |
| 1100 | 1101 | Linalool | 2.4 | 11.0 | 0.5 | 0.6 | 0.3 | 0.5 | 0.7 | 1.0 | --- | 3.7 | --- | --- | --- |
| 1100 | 1104 | Rosefuran | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- |
| 1104 | 1098 | trans-Sabinene hydrate | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- |
| 1105 | 1107 | Nonanal | --- | --- | --- | 0.1 | --- | --- | tr | tr | --- | --- | --- | --- | --- |
| 1112 | 1116 | (E)-4,8-Dimethylnona-1,3,7-triene | 0.1 | --- | --- | tr | --- | --- | --- | --- | --- | 0.4 | --- | --- | --- |
| 1118 | 1114 | endo-Fenchol | --- | --- | --- | --- | 0.1 | 0.7 | --- | --- | --- | 0.2 | --- | --- | --- |
| 1118 | 1118 | cis-p-Menth-2-en-1-ol | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- |
| 1120 | 1128 | allo-Ocimene | 0.3 | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- |
| 1124 | 1118 | exo-Fenchol | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- |
| 1128 | 1122 | α-Campholenal | --- | --- | --- | tr | 0.2 | --- | --- | --- | --- | --- | --- | --- | --- |
| 1135 | 1136 | trans-p-Menth-2-en-1-ol | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- |
| 1137 | 1135 | Nopinone | --- | --- | --- | --- | 0.1 | --- | --- | --- | --- | --- | --- | --- | --- |
| 1141 | 1135 | trans-Pinocarveol | --- | --- | --- | tr | 2.0 | 0.2 | --- | --- | --- | --- | --- | --- | --- |
| 1144 | 1140 | trans-Verbenol | --- | --- | --- | --- | 0.4 | --- | --- | --- | --- | --- | --- | --- | --- |
| 1146 | 1149 | Camphor | --- | --- | --- | --- | 0.1 | 0.3 | 0.1 | tr | --- | --- | --- | --- | --- |
| 1148 | 1165 | Lavandulol | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- |
| 1159 | 1167 | Benzyl acetate | --- | --- | --- | --- | --- | --- | 0.1 | tr | --- | --- | --- | --- | --- |
| 1162 | 1160 | Pinocarvone | --- | --- | 0.2 | tr | 1.5 | 0.2 | --- | --- | --- | --- | 0.2 | --- | 0.3 |
| 1167 | 1165 | iso-Neral | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- |
| 1169 | 1162 | δ-Terpineol | --- | --- | --- | --- | 0.1 | --- | --- | --- | --- | --- | --- | --- | --- |
| 1170 | 1166 | p-Mentha-1,5-dien-8-ol | --- | --- | --- | --- | 0.6 | --- | --- | --- | --- | --- | --- | --- | --- |
| 1171 | 1165 | Borneol | 0.2 | 0.6 | --- | tr | 0.2 | 4.0 | --- | --- | --- | --- | --- | --- | --- |
| 1180 | 1180 | Terpinen-4-ol | --- | 0.2 | --- | 0.1 | 0.4 | 0.4 | 0.1 | 0.1 | 0.1 | 0.1 | --- | --- | --- |
| 1185 | 1179 | p-Cymen-8-ol | --- | --- | --- | tr | 0.3 | --- | --- | --- | --- | --- | --- | --- | --- |
| 1185 | 1185 | iso-Geranial | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- |
| 1194 | 1195 | α-Terpineol | --- | 0.2 | --- | 0.1 | --- | 1.6 | tr | 0.1 | --- | --- | --- | --- | 0.2 |
| 1194 | 1195 | Myrtenal | 0.2 | --- | 0.3 | 0.1 | 2.6 | --- | --- | --- | --- | --- | 0.2 | --- | 0.3 |
| 1194 | 1193 | (4Z)-Decenal | --- | --- | --- | 0.1 | --- | --- | --- | --- | --- | --- | --- | --- | --- |
| 1195 | 1194 | Myrtenol | --- | --- | --- | --- | 0.9 | --- | --- | --- | --- | --- | --- | --- | --- |
| 1198 | 1195 | cis-Piperitol | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- |
| 1205 | 1204 | Verbenone | --- | --- | --- | --- | 0.3 | --- | --- | --- | --- | --- | --- | --- | --- |
| 1206 | 1206 | Decanal | --- | --- | --- | 0.1 | --- | --- | --- | 0.1 | --- | 0.2 | --- | --- | --- |
| 1209 | 1207 | trans-Piperitol | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- |
| 1216 | 1218 | endo-Fenchyl acetate | --- | --- | 0.1 | tr | 0.5 | 6.5 | --- | --- | --- | 0.2 | --- | --- | --- |
| 1217 | 1215 | trans-Carveol | --- | --- | --- | --- | 0.2 | --- | --- | --- | --- | --- | --- | --- | --- |
| 1220 | 1217 | β-Cyclocitral | --- | --- | --- | tr | --- | --- | --- | --- | --- | --- | --- | --- | --- |
| 1222 | 1227 | Nerol | --- | --- | --- | --- | 0.1 | --- | --- | --- | --- | --- | --- | --- | --- |
| 1224 | 1223 | Citronellol | --- | --- | --- | --- | 0.3 | 0.4 | --- | --- | --- | --- | --- | --- | --- |
| 1227 | 1232 | Thymyl methyl ether | --- | --- | --- | --- | --- | 0.4 | --- | --- | --- | --- | --- | --- | --- |
| 1236 | 1235 | Neral | --- | --- | --- | --- | 0.1 | --- | --- | --- | --- | --- | --- | --- | --- |
| 1241 | 1238 | Cuminal | --- | --- | --- | --- | tr | --- | --- | --- | --- | --- | --- | --- | --- |
| 1242 | 1239 | Carvone | --- | --- | --- | --- | 0.2 | --- | --- | --- | --- | --- | --- | --- | --- |
| 1248 | 1249 | Geraniol | --- | --- | --- | --- | 1.0 | 0.8 | --- | --- | --- | 0.3 | --- | --- | --- |
| 1265 | 1264 | Geranial | --- | --- | --- | --- | 0.2 | tr | --- | --- | --- | --- | --- | --- | --- |
| 1271 | 1266 | 1-Decanol | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- |
| 1281 | 1287 | iso-Bornyl acetate | --- | --- | --- | --- | --- | --- | tr | 0.4 | --- | 0.2 | --- | --- | --- |
| 1284 | 1285 | Bornyl acetate | --- | --- | 0.3 | --- | 0.4 | 1.2 | 0.2 | 1.1 | --- | 10.4 | --- | --- | --- |
| 1285 | 1291 | Safrole | --- | --- | --- | --- | --- | --- | tr | 0.2 | --- | --- | --- | --- | --- |
| 1293 | 1290 | Dihydroedulan IIA | --- | --- | --- | 1.1 | --- | --- | --- | --- | --- | --- | 2.6 | 0.8 | 0.1 |
| 1295 | 1298 | Carvacrol | --- | --- | --- | --- | 0.3 | --- | --- | --- | --- | --- | --- | --- | --- |
| 1295 | 1293 | Methyl myrtenate | --- | --- | --- | 0.1 | --- | --- | --- | --- | --- | --- | --- | --- | --- |
| 1307 | 1295 | trans-Sabinyl acetate | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- |
| 1330 | 1324 | Methyl geranate | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- |
| 1330 | 1332 | Bicycloelemene | --- | --- | --- | 0.2 | --- | --- | 1.1 | 0.5 | --- | --- | --- | --- | --- |
| 1330 | 1335 | δ-Elemene | 0.2 | 1.1 | --- | --- | --- | --- | 0.9 | 0.4 | --- | 0.2 | 1.0 | 1.4 | 0.5 |
| 1333 | 1325 | Myrtenyl acetate | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- |
| 1342 | 1349 | α-Cubebene | --- | --- | --- | --- | --- | --- | 0.7 | 0.4 | --- | --- | --- | --- | --- |
| 1345 | 1346 | α-Terpinyl acetate | --- | --- | --- | 0.1 | --- | 1.3 | --- | --- | --- | --- | --- | --- | --- |
| 1364 | 1367 | Cyclosativene | --- | --- | --- | --- | --- | --- | 0.3 | 0.1 | --- | --- | --- | --- | --- |
| 1364 | 1372 | iso-Ledene | --- | --- | --- | --- | --- | --- | --- | tr | --- | --- | --- | --- | --- |
| 1368 | 1373 | α-Ylangene | --- | --- | --- | tr | 0.2 | 0.1 | --- | --- | --- | --- | --- | --- | --- |
| 1374 | 1375 | α-Copaene | --- | --- | --- | 0.1 | 3.1 | 0.7 | 3.2 | 1.5 | --- | 0.2 | --- | 0.3 | 0.2 |
| 1375 | 1380 | Geranyl acetate | --- | --- | --- | --- | --- | --- | --- | 0.2 | --- | 0.5 | --- | --- | --- |
| 1377 | 1383 | cis-β-Elemene | --- | --- | --- | --- | --- | --- | 0.2 | 0.2 | --- | --- | --- | --- | --- |
| 1377 | 1365 | Eugenol | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | 0.9 | --- |
| 1382 | 1382 | β-Bourbonene | --- | --- | --- | tr | --- | --- | 0.4 | 0.1 | --- | --- | --- | --- | --- |
| 1386 | 1387 | β-Cubebene | --- | --- | --- | --- | 0.1 | --- | 1.7 | 0.6 | --- | --- | --- | --- | --- |
| 1387 | 1390 | trans-β-Elemene | 1.1 | 1.1 | 0.4 | 2.3 | 0.4 | 0.8 | 4.7 | 3.7 | --- | 0.8 | 5.9 | 10.0 | 2.5 |
| 1400 | 1403 | Methyl eugenol | --- | --- | --- | 0.1 | --- | --- | --- | --- | --- | --- | --- | --- | --- |
| 1403 | 1405 | Sesquithujene | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- |
| 1403 | 1408 | (Z)-Caryophyllene | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | 0.4 | --- |
| 1405 | 1406 | α-Gurjunene | --- | --- | --- | 0.1 | --- | --- | 0.1 | 0.1 | --- | 0.3 | --- | --- | --- |
| 1405 | 1415 | β-Maaliene | --- | --- | --- | --- | --- | --- | 0.1 | --- | --- | --- | --- | --- | --- |
| 1415 | 1416 | cis-α-Bergamotene | --- | 0.4 | --- | 0.2 | --- | --- | 0.1 | 0.1 | --- | --- | 0.2 | 0.3 | --- |
| 1419 | 1417 | (E)-Caryophyllene | 13.9 | 52.6 | 1.9 | 24.3 | 1.6 | 4.9 | 6.4 | 6.3 | 1.7 | 11.3 | 12.2 | 14.4 | 2.3 |
| 1427 | 1432 | γ-Elemene | --- | --- | --- | --- | --- | 0.3 | 0.7 | --- | --- | --- | --- | --- | --- |
| 1428 | 1424 | 2,5-Dimethoxy-p-cymene | 0.1 | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- |
| 1428 | 1430 | β-Copaene | --- | --- | --- | 0.2 | --- | --- | --- | 0.5 | --- | --- | --- | --- | --- |
| 1428 | 1438 | α-Maaliene | --- | --- | --- | --- | --- | --- | --- | 0.1 | --- | --- | --- | --- | --- |
| 1431 | 1416 | α-Santalene | --- | --- | 0.1 | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- |
| 1431 | 1432 | trans-α-Bergamotene | 0.2 | 0.1 | --- | tr | --- | --- | 0.3 | 0.1 | --- | --- | --- | --- | --- |
| 1433 | 1437 | α-Guaiene | --- | --- | --- | tr | --- | 0.2 | --- | --- | --- | --- | --- | --- | --- |
| 1437 | 1438 | Aromadendrene | 0.2 | 0.2 | --- | 0.1 | --- | --- | 0.6 | 0.5 | --- | --- | 0.2 | 0.4 | 0.4 |
| 1440 | 1440 | (Z)-β-Farnesene | 0.2 | 0.2 | --- | --- | --- | --- | --- | --- | --- | 1.1 | --- | 0.5 | --- |
| 1444 | 1431 | β-Gurjunene | --- | 0.1 | --- | --- | --- | --- | --- | --- | --- | --- | --- | 0.9 | 0.2 |
| 1445 | 1445 | Myltayl-4(12)-ene | --- | --- | --- | 0.1 | --- | --- | --- | --- | --- | --- | --- | --- | --- |
| 1446 | 1453 | Geranyl acetone | --- | --- | --- | 0.3 | --- | --- | --- | --- | --- | --- | --- | --- | --- |
| 1447 | 1453 | trans-Muurola-3,5-diene | --- | --- | --- | --- | 0.1 | 0.4 | 0.2 | 0.1 | --- | --- | --- | --- | --- |
| 1450 | 1452 | (E)-β-Farnesene | --- | --- | 0.3 | 0.2 | --- | 0.1 | 0.3 | 0.2 | --- | --- | --- | --- | --- |
| 1452 | 1457 | Prezizaene | --- | --- | --- | --- | --- | --- | --- | 0.4 | --- | --- | --- | --- | --- |
| 1454 | 1454 | α-Humulene | 2.1 | 5.9 | 0.3 | 3.2 | 1.1 | 4.8 | 6.4 | 8.8 | 0.2 | 6.9 | 1.1 | 1.6 | 0.5 |
| 1458 | 1457 | allo-Aromadendrene | --- | --- | --- | 0.9 | 0.7 | 0.3 | 1.9 | 0.1 | --- | --- | --- | --- | --- |
| 1460 | 1461 | cis-Cadina-1(6),4-diene | --- | --- | --- | --- | 0.1 | 0.1 | --- | --- | --- | --- | --- | --- | --- |
| 1470 | 1472 | trans-Cadina-1(6),4-diene | --- | --- | --- | --- | 0.2 | --- | 0.4 | 0.2 | --- | --- | --- | --- | --- |
| 1471 | 1476 | β-Chamigrene | --- | --- | --- | 0.1 | --- | --- | --- | --- | --- | --- | 0.6 | 2.6 | 0.7 |
| 1471 | 1475 | γ-Gurjunene | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | 1.5 | --- |
| 1472 | 1482 | γ-Curcumene | --- | --- | --- | --- | --- | --- | 0.3 | 0.3 | --- | --- | --- | --- | --- |
| 1473 | 1478 | γ-Muurolene | 0.2 | --- | --- | 0.1 | 0.3 | 0.1 | 0.3 | 0.3 | --- | 0.4 | --- | --- | --- |
| 1474 | 1476 | Selina-4,11-diene | --- | --- | --- | --- | --- | 1.8 | --- | 0.1 | --- | --- | --- | --- | --- |
| 1477 | 1482 | α-Amorphene | --- | --- | --- | --- | tr | 0.2 | --- | 0.1 | --- | --- | --- | --- | --- |
| 1477 | 1479 | ar-Curcumene | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | 1.4 | 2.8 | 0.8 |
| 1477 | 1487 | (E)-β-Ionone | --- | --- | --- | 0.1 | --- | --- | --- | --- | --- | --- | --- | --- | --- |
| 1478 | 1483 | trans-β-Bergamotene | --- | --- | --- | --- | --- | --- | 0.2 | --- | --- | --- | --- | --- | --- |
| 1479 | 1464 | 9-epi-(E)-Caryophyllene | 1.7 | 0.5 | --- | --- | --- | --- | --- | --- | --- | 0.3 | --- | 0.2 | - |
| 1480 | 1480 | Germacrene D | 0.6 | 0.4 | --- | 3.6 | tr | 0.3 | 6.5 | 4.2 | 0.8 | 2.7 | 2.0 | 1.4 | 2.2 |
| 1481 | 1488 | δ-Selinene | --- | --- | --- | --- | --- | --- | --- | 0.2 | --- | --- | --- | --- | --- |
| 1482 | 1492 | cis-β-Guaiene | --- | --- | --- | --- | --- | --- | 0.3 | --- | --- | --- | --- | --- | --- |
| 1483 | 1487 | Aristolochene | --- | --- | --- | 0.5 | 0.8 | --- | --- | --- | --- | --- | --- | --- | --- |
| 1488 | 1492 | β-Selinene | 0.6 | 0.3 | 0.3 | 0.8 | 5.4 | 2.4 | 1.4 | 1.8 | --- | 0.3 | --- | 2.0 | 0.6 |
| 1489 | 1496 | Valencene | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- |
| 1490 | 1490 | γ-Amorphene | --- | --- | --- | 0.1 | 0.1 | 0.4 | 1.3 | 0.8 | --- | --- | 0.8 | 1.6 | --- |
| 1491 | 1493 | α-Zingiberene | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | 3.6 | 1.7 | 0.6 |
| 1494 | 1498 | α-Selinene | --- | --- | --- | --- | 1.4 | 2.2 | --- | --- | --- | --- | 1.0 | --- | --- |
| 1494 | 1497 | Bicyclogermacrene | 9.6 | 1.1 | 0.3 | 3.3 | --- | --- | 16.1 | 9.8 | 2.0 | 5.1 | 0.7 | --- | 1.3 |
| 1496 | 1500 | α-Muurolene | --- | --- | --- | 0.3 | 0.2 | 0.2 | --- | --- | --- | --- | --- | --- | --- |
| 1496 | 1506 | α-Bulnesene | --- | --- | --- | --- | --- | --- | 0.5 | 0.3 | --- | --- | --- | --- | --- |
| 1497 | 1502 | trans-β-Guiaene | --- | --- | --- | --- | --- | --- | 0.1 | --- | --- | --- | --- | --- | --- |
| 1500 | 1500 | n-Pentadecane | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- |
| 1501 | 1505 | (E,E)-α-Farnesene | 1.6 | 0.7 | 0.4 | --- | 0.4 | 0.3 | --- | --- | --- | 0.7 | --- | --- | --- |
| 1501 | 1495 | γ-Amorphene | --- | 0.2 | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- |
| 1502 | 1496 | Viridiflorene | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | 0.4 |
| 1503 | 1509 | β-Curcumene | --- | --- | --- | --- | --- | --- | 0.1 | 0.1 | --- | --- | --- | --- | --- |
| 1506 | 1508 | β-Bisabolene | --- | --- | --- | 0.1 | 0.1 | 0.6 | 0.7 | 0.3 | --- | --- | 1.4 | 2.0 | 0.4 |
| 1511 | 1512 | γ-Cadinene | 0.1 | --- | --- | 0.2 | 1.7 | 3.1 | 0.2 | 0.2 | --- | 0.2 | --- | 0.3 | 0.2 |
| 1511 | 1511 | δ-Amorphene | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- |
| 1513 | 1519 | Cubebol | --- | --- | --- | --- | 0.1 | --- | 0.4 | 0.2 | --- | --- | --- | --- | --- |
| 1516 | 1518 | δ-Cadinene | 0.6 | 0.2 | --- | 0.4 | 1.0 | 1.7 | 2.9 | 2.4 | --- | 0.5 | --- | 0.9 | 1.4 |
| 1517 | 1520 | 7-epi-α-Selinene | --- | --- | --- | --- | tr | 1.1 | --- | --- | --- | --- | 1.0 | 1.1 | --- |
| 1517 | 1521 | Zonarene | --- | --- | --- | --- | --- | --- | 0.1 | 0.3 | --- | --- | --- | --- | --- |
| 1519 | 1521 | trans-Calamenene | --- | --- | --- | --- | 0.1 | --- | --- | --- | --- | --- | --- | --- | --- |
| 1520 | 1528 | (E)-γ-Bisabolene | --- | --- | --- | --- | --- | --- | 0.5 | 0.2 | --- | --- | --- | --- | --- |
| 1523 | 1521 | Eugenyl acetate | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | 1.0 | 0.6 |
| 1524 | 1521 | β-Sesquiphellandrene | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | 0.7 | 0.5 | --- |
| 1529 | 1528 | cis-Calamene | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | 0.3 | --- |
| 1530 | 1536 | trans-Cadina-1,4-diene | --- | --- | --- | --- | 0.1 | --- | 0.3 | 0.2 | --- | --- | --- | --- | --- |
| 1535 | 1537 | α-Cadinene | --- | --- | --- | --- | tr | 0.2 | --- | --- | --- | --- | --- | --- | --- |
| 1539 | 1544 | α-Calacorene | --- | --- | --- | --- | 0.5 | 0.4 | --- | --- | --- | --- | --- | --- | --- |
| 1546 | 1549 | α-Elemol | --- | 0.2 | --- | --- | 0.1 | 0.4 | 0.8 | 0.9 | --- | 0.2 | --- | 0.6 | --- |
| 1557 | 1559 | Germacrene B | --- | --- | --- | --- | --- | 0.7 | 0.6 | 0.2 | --- | --- | 0.3 | --- | 0.2 |
| 1558 | 1561 | (E)-Nerolidol | 1.1 | 1.2 | 0.2 | 0.2 | 1.1 | --- | --- | --- | 0.3 | 3.3 | 0.8 | 3.7 | 0.9 |
| 1560 | 1564 | β-Calacorene | --- | --- | --- | --- | 0.2 | 0.1 | --- | --- | --- | --- | --- | --- | --- |
| 1564 | 1568 | Palustrol | --- | --- | --- | --- | --- | --- | tr | 0.1 | --- | --- | --- | --- | --- |
| 1568 | 1571 | Maaliol | --- | --- | --- | --- | --- | 0.2 | tr | 0.3 | --- | --- | --- | --- | --- |
| 1570 | 1573 | (3E,7E)-4,8,12-Trimethyltrideca-1,3,7,11-tetraene | --- | --- | --- | 0.1 | --- | --- | --- | --- | --- | --- | --- | --- | --- |
| 1575 | 1576 | Spathulenol | 0.9 | 0.7 | 0.1 | 2.1 | tr | 0.2 | 1.2 | 0.9 | 0.2 | 1.3 | --- | 1.2 | 0.2 |
| 1575 | 1582 | Neryl isovalerate | --- | --- | --- | --- | --- | 0.2 | --- | --- | --- | --- | --- | --- | --- |
| 1575 | 1574 | Germacrene D-4-ol | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- |
| 1582 | 1577 | Caryophyllene oxide | 1.3 | 4.4 | 0.4 | 3.6 | 8.0 | 2.4 | tr | 0.3 | 0.2 | 4.1 | 2.0 | 4.5 | 1.6 |
| 1582 | 1582 | ar-Turmerol | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- |
| 1583 | 1570 | Dendrolasin | 0.2 | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- |
| 1583 | 1585 | epi-Globulol | --- | --- | --- | 0.8 | tr | 0.9 | --- | --- | --- | --- | --- | --- | --- |
| 1587 | 1590 | Globulol | --- | --- | --- | --- | tr | 0.4 | 1.0 | 1.6 | --- | --- | --- | --- | --- |
| 1590 | 1581 | Clovenol | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | 0.2 | --- |
| 1592 | 1594 | Salvial-4(14)en-1-one | --- | --- | --- | 0.1 | --- | --- | --- | --- | --- | --- | --- | --- | --- |
| 1593 | 1592 | Viridiflorol | --- | --- | --- | 0.2 | --- | 0.5 | 0.7 | 1.2 | --- | --- | --- | --- | --- |
| 1593 | 1593 | Scapanol g | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- |
| 1594 | 1600 | Guaiol | 0.4 | 0.6 | 0.1 | --- | 0.2 | 0.5 | --- | --- | --- | 0.2 | --- | 0.7 | --- |
| 1594 | 1601 | trans-β-Elemenone | --- | --- | --- | --- | tr | 0.4 | --- | --- | --- | --- | --- | --- | --- |
| 1596 | 1599 | Cubeban-11-ol | --- | --- | --- | 0.1 | --- | --- | tr | 0.3 | --- | --- | --- | --- | --- |
| 1599 | 1592 | Humulene epoxide I | --- | --- | --- | --- | --- | --- | --- | --- | --- | 0.4 | --- | 0.9 | --- |
| 1600 | 1609 | Rosifoliol | --- | --- | --- | --- | --- | --- | 0.2 | 0.4 | --- | --- | --- | --- | --- |
| 1604 | 1607 | 5,7-di-epi-Eudesmol | --- | --- | --- | --- | --- | 0.2 | --- | --- | --- | --- | --- | --- | --- |
| 1604 | 1602 | Ledol | --- | --- | --- | 0.3 | --- | --- | --- | --- | --- | 0.2 | --- | --- | --- |
| 1613 | 1618 | 1,10-di-epi-Cubenol | --- | --- | --- | --- | 0.2 | 0.7 | --- | --- | --- | --- | --- | --- | --- |
| 1616 | 1613 | Humulene epoxide II | 0.2 | 0.8 | --- | 0.4 | 1.8 | 1.3 | tr | 0.2 | --- | 2.8 | --- | 0.6 | 0.6 |
| 1616 | 1611 | Zingiberenol | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | 0.3 | --- |
| 1618 | 1623 | Humulane-1,6-dien-3-ol | --- | --- | --- | --- | --- | --- | 8.2 | 9.9 | --- | --- | --- | --- | --- |
| 1620 | 1622 | 10-epi-γ-Eudesmol | --- | --- | --- | --- | tr | 0.4 | --- | --- | --- | 2.9 | --- | --- | --- |
| 1625 | 1631 | 1-epi-Cubenol | 0.2 | 0.3 | --- | --- | 0.2 | 0.3 | --- | 1.3 | --- | --- | --- | --- | --- |
| 1627 | 1618 | epi-Cedrol | --- | --- | 0.2 | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- |
| 1628 | 1622 | Alismol | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | 0.4 |
| 1630 | 1630 | γ-Eudesmol | --- | --- | --- | --- | 0.3 | 1.7 | 0.5 | 0.7 | --- | --- | --- | --- | --- |
| 1632 | 1631 | Caryophylla-4(12),8(13)-dien-5α-ol | --- | --- | --- | 0.7 | 0.1 | --- | --- | --- | --- | --- | --- | --- | --- |
| 1634 | 1632 | α-Acorenol | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- |
| 1636 | 1639 | Caryophylla-4(12),8(13)-dien-5β-ol | --- | --- | --- | 0.9 | 0.3 | 0.4 | --- | --- | --- | --- | --- | --- | --- |
| 1640 | 1638 | τ-Cadinol | 0.9 | 2.2 | --- | 0.1 | --- | 0.3 | --- | --- | --- | --- | --- | 1.7 | 0.6 |
| 1640 | 1643 | Cubenol | --- | --- | --- | --- | 0.1 | --- | 0.6 | 0.7 | --- | --- | --- | --- | --- |
| 1642 | 1644 | τ-Murrolol | --- | --- | --- | 0.2 | 0.1 | 0.4 | --- | 0.4 | --- | 0.2 | --- | 0.6 | --- |
| 1645 | 1644 | α-Muurolol (= δ-Cadinol) | 0.2 | --- | --- | 0.1 | tr | 0.1 | 0.2 | 0.7 | --- | --- | --- | --- | --- |
| 1651 | 1649 | β-Eudesmol | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- |
| 1652 | 1652 | α-Cadinol | 0.5 | 0.5 | --- | 0.7 | tr | 1.3 | 1.0 | 1.7 | --- | 0.3 | 0.2 | 2.0 | 0.5 |
| 1652 | 1647 | 1,2-Diacetoxy-4-allylbenzene | 0.1 | 1.9 | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- |
| 1653 | 1652 | α-Eudesmol | --- | --- | --- | --- | 0.6 | 1.1 | --- | --- | --- | --- | --- | 1.0 | --- |
| 1657 | 1658 | Selin-11-en-4α-ol | --- | --- | --- | 0.2 | tr | 0.5 | --- | --- | --- | --- | --- | --- | --- |
| 1662 | 1658 | neo-Intermedeol | 0.7 | 1.2 | --- | --- | 0.2 | 4.4 | 0.2 | 0.6 | --- | 0.2 | 0.4 | 2.6 | --- |
| 1668 | 1668 | 14-Hydroxy-9-epi-(E)-caryophyllene | --- | 0.6 | --- | 0.6 | 0.3 | 0.5 | --- | --- | --- | 0.4 | 0.2 | 1.0 | --- |
| 1670 | 1675 | Cadalene | --- | --- | --- | --- | 0.1 | --- | --- | --- | --- | --- | --- | --- | --- |
| 1671 | 1676 | Mustakone | --- | --- | --- | --- | 0.1 | --- | --- | --- | --- | --- | --- | --- | --- |
| 1679 | 1681 | γ-Bicyclofarnesal | --- | 0.9 | 0.6 | --- | --- | --- | --- | --- | --- | 1.2 | --- | 0.6 | 0.4 |
| 1681 | 1685 | α-Bisabolol | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- |
| 1684 | 1685 | Germacra-4(15),5,10(14)-trien-1α-ol | --- | --- | --- | 0.1 | --- | --- | --- | --- | --- | --- | --- | --- | --- |
| 1684 | 1683 | epi-α-Bisabolol | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- |
| 1690 | 1693 | Germacrone | --- | --- | --- | --- | 0.2 | 1.6 | --- | --- | --- | --- | --- | --- | --- |
| 1691 | 1694 | Germacra-4(15),5,10(14)-trien-1β-ol | --- | --- | --- | 0.1 | --- | --- | --- | --- | --- | --- | --- | --- | --- |
| 1696 | 1689 | 6α-Hydroxygermacra-1(10),4-diene | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- |
| 1701 | 1698 | (2Z,6Z)-Farnesol | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- |
| 1702 | 1690 | (Z)-trans-α-Bergamotol | --- | 0.3 | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- |
| 1707 | 1714 | Nootkatol | --- | --- | --- | 0.1 | --- | --- | --- | --- | --- | --- | --- | --- | --- |
| 1713 | 1713 | Pentadecanal | --- | --- | --- | 0.1 | --- | --- | --- | --- | --- | 0.3 | --- | --- | 0.9 |
| 1738 | 1740 | Mint sulfide | --- | --- | --- | 0.1 | --- | --- | --- | --- | 0.2 | 0.9 | 0.6 | 2.6 | --- |
| 1739 | 1732 | Zerumbone | --- | --- | --- | --- | --- | --- | --- | --- | 0.2 | 3.9 | --- | --- | --- |
| 1761 | 1759 | Benzyl benzoate | --- | --- | --- | --- | --- | --- | 2.5 | 3.0 | --- | 0.3 | --- | 0.3 | --- |
| 1774 | 1760 | (Z)-Lanceol | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- |
| 1799 | 1809 | Ambrial | --- | --- | --- | 0.2 | tr | 0.5 | --- | --- | --- | --- | 0.4 | 2.8 | 2.4 |
| 1838 | 1838 | Phytone | --- | --- | --- | 0.7 | --- | --- | --- | --- | --- | --- | 0.2 | 0.2 | --- |
| 1848 | 1845 | 6,10,14-Trimethylpentadecan-2-one | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- | --- |
| 1862 | 1864 | Benzyl salicylate | --- | --- | --- | --- | --- | --- | 0.5 | 0.8 | --- | --- | --- | --- | --- |
| 2002 | --- | (E)-15,16-Bisnorlabda-8(17),11-dien-13-one | --- | 0.8 | --- | --- | --- | --- | --- | --- | --- | --- | --- | 0.3 | --- |
| 2104 | 2105 | (E)-Phytol | --- | 0.2 | --- | 0.9 | --- | --- | --- | --- | --- | 0.2 | --- | 0.6 | --- |
| Monoterpene hydrocarbons | 50.3 | 0.5 | 84.8 | 39.4 | 29.6 | 23.9 | 16.0 | 23.4 | 94.2 | 18.3 | 52.7 | 3.6 | 61.8 | ||
| Oxygenated monoterpenoids | 3.8 | 12.0 | 1.7 | 2.0 | 34.8 | 25.3 | 1.1 | 2.9 | 0.1 | 16.2 | 0.4 | 0.0 | 0.8 | ||
| Sesquiterpene hydrocarbons | 32.9 | 65.1 | 4.0 | 41.4 | 20.0 | 28.5 | 63.3 | 46.6 | 4.7 | 31.0 | 34.1 | 49.1 | 15.4 | ||
| Oxygenated sesquiterpenoids | 6.6 | 13.9 | 1.6 | 11.6 | 14.0 | 21.3 | 14.8 | 22.2 | 0.9 | 21.6 | 3.6 | 22.2 | 5.2 | ||
| Others | 0.2 | 2.9 | 0.0 | 4.0 | 0.8 | 0.5 | 3.1 | 4.1 | 0.2 | 2.3 | 3.8 | 9.5 | 4.0 | ||
| Total identified | 93.8 | 94.4 | 92.1 | 98.4 | 99.2 | 99.5 | 98.3 | 99.2 | 100.0 | 89.4 | 94.6 | 84.4 | 87.2 | ||
| Major Components | Collection Location | |||||
|---|---|---|---|---|---|---|
| Johor, Malaysia [37] | Petchaburi, Thailand [39] | Sabah, Malaysia [38] | Chiang Mai, Thailand [40] | Bandung, Indonesia [41] | Pù Hoạt Nature Reserve, Vietnam (This Work) | |
| Sabinene | 7.2 | 6.5 | 4.1 | 21.1 | 8.6 | 21.6 |
| β-Pinene | 5.1 | 4.3 | 1.3 | 6.8 | 3.5 | 11.7 |
| p-Cymene | 0.0 | 6.9 | 0.0 | 0.0 | 0.0 | 2.1 |
| 1,8-Cineole | 3.3 | 1.2 | 0.6 | 3.9 | 5.8 | 4.3 |
| γ-Terpinene | 5.1 | 0.2 | 0.4 | 4.7 | 4.8 | 5.5 |
| Terpinen-4-ol | 16.8 | 11.2 | 3.2 | 19.7 | 16.6 | 17.1 |
| α-Humulene | 10.9 | 5.6 | 18.3 | 15.6 | 3.3 | 3.8 |
| Zerumbone | 25.6 | 40.1 | 36.7 | 0.0 | 14.2 | 12.5 |
| Major Components | Vu Quang National Park [46] | Pù Hoạt Nature Reserve | ||||
|---|---|---|---|---|---|---|
| Leaf | Stem | Rhizome | Leaf | Stem | Rhizome | |
| α-Pinene | 6.9 | 5.2 | 2.2 | 11.3 | 0.5 | 9.8 |
| β-Pinene | 24.7 | 26.1 | 8.0 | 38.5 | 3.1 | 45.0 |
| Bornyl acetate | 2.5 | 0.0 | 20.9 | 0.0 | 0.0 | 0.0 |
| trans-β-Elemene | 0.8 | 0.5 | 0.6 | 5.9 | 10.0 | 2.5 |
| (E)-Caryophyllene | 12.3 | 13.9 | 4.5 | 12.2 | 14.4 | 2.3 |
| α-Humulene | 7.7 | 8.2 | 9.6 | 1.1 | 1.6 | 0.5 |
| Zerumbone | 3.0 | 3.6 | 14.1 | 0.0 | 0.0 | 0.0 |
| Zingiber Essential Oil | LC50 (95% Confidence Levels) | LC90 (95% Confidence Levels) | χ2 | p |
|---|---|---|---|---|
| Twenty-four-hour | ||||
| Aedes aegypti | ||||
| Z. cornubracteatum (BE) rhizome | 16.97 (15.49–18.42) | 24.56 (22.65–27.31) | 1.7 × 10−6 | 1.000 |
| Z. neotruncatum rhizome | 34.95 (32.49–37.37) | 51.49 (48.32–55.56) | 6.625 | 0.036 |
| Z. nudicarpum (PH) leaf | 19.30 (17.95–20.56) | 23.74 (22.40–25.40) | 0.0 | 1.000 |
| Z. nudicarpum (PH) rhizome | 23.44 (21.94–25.17) | 31.81 (29.07–36.31) | 1.1 × 10−3 | 0.999 |
| Z. ottensii rhizome | 38.16 (35.50–40.89) | 57.87 (53.60–63.62) | 3.286 | 0.193 |
| Z. recurvatum rhizome | 20.90 (18.45–23.30) | 36.35 (32.79–41.63) | 3.697 | 0.157 |
| Permethrin (control) | 0.0094 (0.0082–0.0107) | 0.0211 (0.0185–0.0249) | 57.6 | 0.000 |
| Aedes albopictus | ||||
| Z. cornubracteatum (BE) rhizome | 12.72 (10.43–14.41) | 21.56 (19.43–25.10) | 4.6 × 10−6 | 1.000 |
| Z. neotruncatum rhizome | 21.50 (19.71–23.52) | 31.99 (28.97–36.94) | 0.0402 | 0.980 |
| Z. nudicarpum (PH) leaf | 22.33 (20.93–23.86) | 31.12 (27.52–36.51) | 0.2454 | 0.970 |
| Z. nudicarpum (PH) rhizome | 28.05 (25.80–30.41) | 42.09 (38.92–46.29) | 3.205 | 0.201 |
| Z. ottensii rhizome | 19.79 (17.91–21.75) | 30.81 (27.81–35.76) | 0.03531 | 0.983 |
| Z. recurvatum rhizome | 45.58 (43.24–47.49) | 58.58 (55.18–63.22) | 0.01253 | 0.994 |
| Permethrin (control) | 0.0024 (0.0021–0.0026) | 0.0042 (0.0038–0.0049) | 4.64 | 0.031 |
| Culex quinquefasciatus | ||||
| Z. cornubracteatum (BE) rhizome | 24.31 (21.79–26.84) | 41.34 (37.72–46.34) | 25.99 | 0.000 |
| Z. neotruncatum rhizome | 33.58 (31.39–36.00) | 42.76 (39.94–46.48) | 6.675 | 0.036 |
| Z. nudicarpum (PH) leaf | 12.44 (8.78–15.67) | 44.29 (39.05–51.64) | 34.20 | 0.000 |
| Z. nudicarpum (PH) rhizome | 11.50 (9.65–13.36) | 27.15 (23.92–31.87) | 30.33 | 0.000 |
| Z. ottensii rhizome | 27.19 (25.04–29.58) | 40.09 (36.82–44.64) | 8.377 | 0.015 |
| Z. recurvatum rhizome | 31.67 (29.30–34.05) | 47.02 (43.94–50.94) | 1.300 | 0.522 |
| Permethrin (control) | 0.0188 (0.0173–0.0206) | 0.0294 (0.0270–0.0326) | 24.1 | 0.000 |
| Forty-eight-hour | ||||
| Aedes aegypti | ||||
| Z. cornubracteatum (BE) rhizome | 16.32 (14.72–17.84) | 24.52 (22.50–27.54) | 9.8 × 10−6 | 1.000 |
| Z. neotruncatum rhizome | 33.91 (31.45–36.33) | 50.42 (47.26–54.46) | 8.156 | 0.017 |
| Z. nudicarpum (PH) leaf | 16.97 (15.76–18.32) | 22.22 (20.59–24.53) | 0 | 1.000 |
| Z. nudicarpum (PH) rhizome | 22.73 (21.09–24.63) | 31.31 (28.88–35.35) | 8.8 × 10−3 | 0.996 |
| Z. ottensii rhizome | 36.59 (33.77–39.48) | 57.61 (53.69–62.84) | 7.113 | 0.029 |
| Z. recurvatum rhizome | 17.16 (14.00–19.79) | 34.57 (30.87–40.26) | 3.687 | 0.158 |
| Aedes albopictus | ||||
| Z. cornubracteatum (BE) rhizome | 12.32 (10.36–13.78) | 19.57 (17.61–23.23) | 0 | 1.000 |
| Z. neotruncatum rhizome | 19.97 (18.07–21.98) | 31.24 (28.16–36.33) | 0.0528 | 0.974 |
| Z. nudicarpum (PH) leaf | 14.31 (12.03–16.52) | 29.76 (27.71–32.79) | 2.548 | 0.467 |
| Z. nudicarpum (PH) rhizome | 23.80 (21.40–26.25) | 39.59 (36.06–44.53) | 12.33 | 0.002 |
| Z. ottensii rhizome | 17.30 (15.15–19.24) | 28.94 (25.97–33.94) | 0.02453 | 0.988 |
| Z. recurvatum rhizome | 42.47 (40.04–44.88) | 57.289 (54.63–61.53) | 1.031 | 0.597 |
| Culex quinquefasciatus | ||||
| Z. cornubracteatum (BE) rhizome | 11.48 (5.78–15.28) | 33.79 (29.71–40.16) | 9.085 | 0.011 |
| Z. neotruncatum rhizome | 29.91 (27.77–32.33) | 41.42 (38.31–45.66) | 13.46 | 0.001 |
| Z. nudicarpum (PH) leaf | 4.901 (3.982–5.817) | 18.31 (14.98–23.84) | 83.59 | 0.000 |
| Z. nudicarpum (PH) rhizome | 6.687 (4.942–8.197) | 18.85 (16.52–22.30) | 49.77 | 0.000 |
| Z. ottensii rhizome | 24.64 (22.58–26.98) | 36.94 (33.58–41.98) | 1.512 | 0.470 |
| Z. recurvatum rhizome | 7.965 (4.368–10.991) | 33.52 (26.87–46.73) | 2.905 | 0.234 |
| Zingiber Essential Oil (EO) | Gram-Positive Bacteria | Gram-Negative Bacteria | Yeast | ||||
|---|---|---|---|---|---|---|---|
| Enterococcus faecalis ATCC 299212 | Staphylococcus aureus ATCC 25923 | Bacillus cereus ATCC 14579 | Escherichia coli ATCC 25922 | Pseudomonas aeruginosa ATCC 27853 | Salmonella enterica ATCC 13076 | Candida albicans ATCC 10231 | |
| MIC (μg/mL) | |||||||
| Z. cornubracteatum (BE) leaf EO | 128 | 128 | na | 64 | na | na | na |
| Z. cornubracteatum (BE) rhizome EO | 32 | 32 | 64 | 256 | na | na | 256 |
| Z. cornubracteatum (BE) stem EO | 128 | 128 | na | 64 | na | na | na |
| Z. cornubracteatum (PM) leaf EO | 64 | 64 | 128 | 128 | na | na | 16 |
| Z. cornubracteatum (PM) rhizome EO | 32 | 64 | 128 | 64 | 256 | na | 16 |
| Z. nudicarpum (PH) leaf EO | 32 | 16 | 16 | na | 128 | na | 16 |
| Z. nudicarpum (PH) rhizome EO | 2 | 8 | 1 | 64 | 16 | na | 16 |
| Z. ottensii leaf EO | 64 | 64 | 64 | na | 16 | na | 64 |
| Z. ottensii rhizome EO | 8 | 64 | 32 | na | na | na | 8 |
| Z. recurvatum leaf EO | 16 | 64 | 128 | na | na | na | 128 |
| Z. recurvatum rhizome EO | 16 | 128 | 64 | na | 256 | 256 | 64 |
| Z. vuquangensis leaf EO | 16 | 16 | 16 | na | na | na | 32 |
| Streptomycin | 256 | 256 | 128 | 32 | 256 | 128 | nt |
| Nistatin | nt | nt | nt | nt | nt | nt | 8 |
| Cyclohexamide | nt | nt | nt | nt | nt | nt | 32 |
| IC50 (μg/mL) | |||||||
| Z. cornubracteatum (BE) leaf EO | 45.67 | 45.67 | na | 18.78 | na | na | na |
| Z. cornubracteatum (BE) rhizome EO | 10.45 | 9.34 | 18.79 | 68.99 | na | na | 99.34 |
| Z. cornubracteatum (BE) stem EO | 46.78 | 19.78 | na | 67.89 | na | na | na |
| Z. cornubracteatum (PM) leaf EO | 20.78 | 9.34 | 45.67 | 45.67 | na | na | 5.67 |
| Z. cornubracteatum (PM) rhizome EO | 9.56 | 13.56 | 43.67 | 20.45 | 100.34 | na | 7.89 |
| Z. nudicarpum leaf EO | 16.33 | 8.54 | 8.57 | na | 65.44 | na | 8.67 |
| Z. nudicarpum rhizome EO | 1.33 | 4.35 | 0.567 | 33.22 | 8.66 | na | 8.99 |
| Z. ottensii leaf EO | 32.66 | 32.33 | 33.77 | na | 7.99 | na | 32.33 |
| Z. ottensii rhizome EO | 2.57 | 17.89 | 15.56 | na | na | na | 3.56 |
| Z. recurvatum leaf EO | 3.99 | 13.67 | 55.89 | na | na | na | 67.74 |
| Z. recurvatum rhizome EO | 6.46 | 36.87 | 32.33 | na | 108.99 | 112.67 | 25.67 |
| Z. vuquangensis leaf EO | 15.66 | 8.56 | 7.33 | na | na | na | 16.33 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huong, L.T.; Chung, N.T.; Huong, T.T.; Sam, L.N.; Hung, N.H.; Ogunwande, I.A.; Dai, D.N.; Linh, L.D.; Setzer, W.N. Essential Oils of Zingiber Species from Vietnam: Chemical Compositions and Biological Activities. Plants 2020, 9, 1269. https://doi.org/10.3390/plants9101269
Huong LT, Chung NT, Huong TT, Sam LN, Hung NH, Ogunwande IA, Dai DN, Linh LD, Setzer WN. Essential Oils of Zingiber Species from Vietnam: Chemical Compositions and Biological Activities. Plants. 2020; 9(10):1269. https://doi.org/10.3390/plants9101269
Chicago/Turabian StyleHuong, Le Thi, Nguyen Thanh Chung, Trinh Thi Huong, Ly Ngoc Sam, Nguyen Huy Hung, Isiaka Ajani Ogunwande, Do Ngoc Dai, Le Duy Linh, and William N. Setzer. 2020. "Essential Oils of Zingiber Species from Vietnam: Chemical Compositions and Biological Activities" Plants 9, no. 10: 1269. https://doi.org/10.3390/plants9101269
APA StyleHuong, L. T., Chung, N. T., Huong, T. T., Sam, L. N., Hung, N. H., Ogunwande, I. A., Dai, D. N., Linh, L. D., & Setzer, W. N. (2020). Essential Oils of Zingiber Species from Vietnam: Chemical Compositions and Biological Activities. Plants, 9(10), 1269. https://doi.org/10.3390/plants9101269