

Entomopathogenic Bacteria Species and Toxins Targeting Aphids (Hemiptera: Aphididae): A Review

 , ,
, ,  ,
,  , ,
, ,  and
and
Abstract
1. Introduction
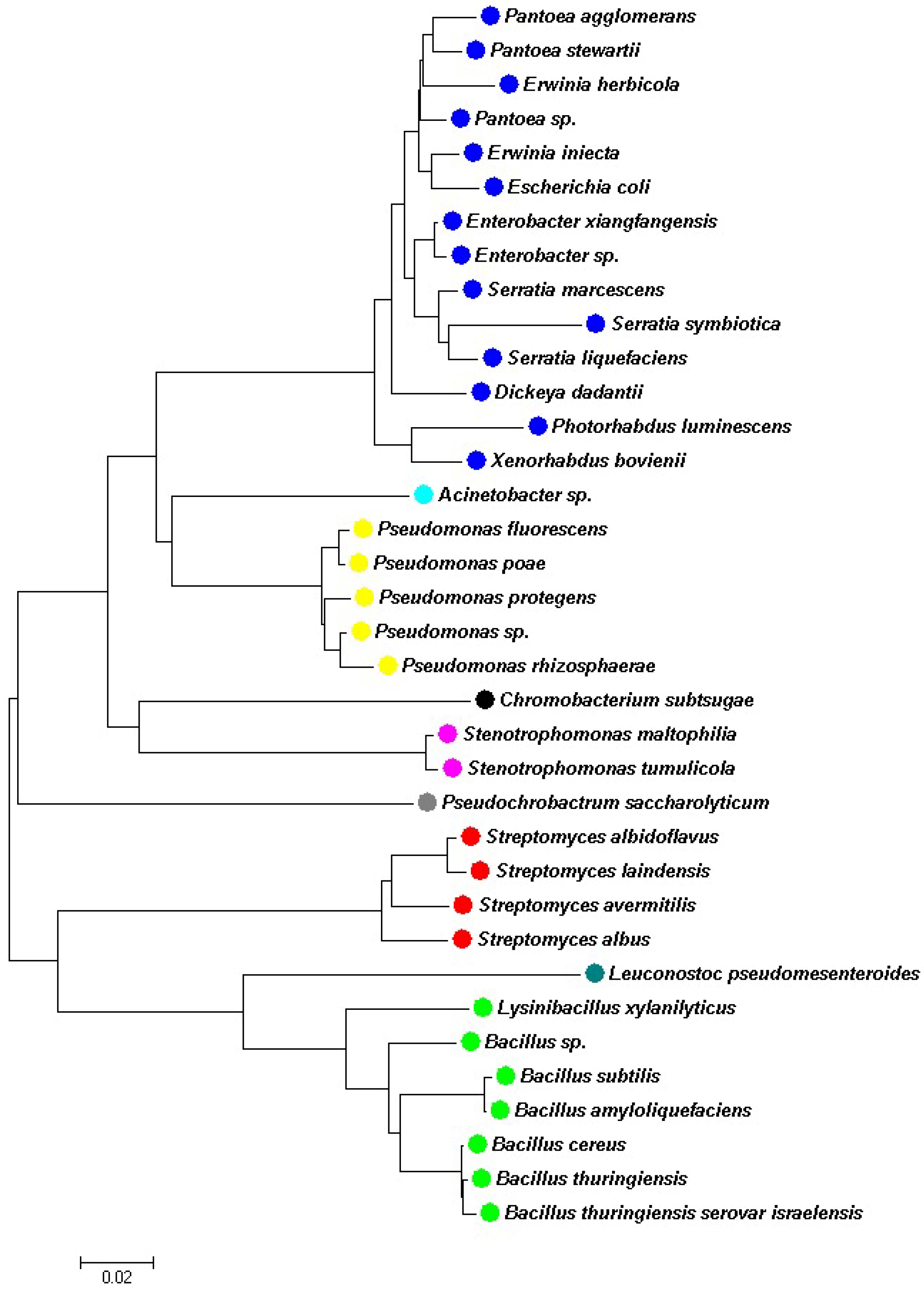
2. Entomopathogenic Bacterial Families That Affect Aphids
2.1. Bacillaceae
2.2. Enterobacteriaceae
2.3. Moraxellaceae
2.4. Xanthomonadaceae
2.5. Pseudomonadaceae
2.6. Streptomycetaceae
2.7. Neisseriaceae
2.8. Brucellaceae
2.9. Leuconostocaceae
3. Possible Reasons for the Limited Toxicity of BPP Against Aphids
4. Mode of Action of Bacterial BPPs in Aphids
5. Conclusions and Future Perspectives
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Remaudière, G.; Remaudière, M. Catalogue of the Worlds Aphididae (Homoptera Aphidoidea); INRA: París, Francia, 1997; p. 473. [Google Scholar]
- Emden, H.F.; Harrington, R. Aphids as Crop Pests; CAB International: Wallingford, UK, 2017; p. 714. [Google Scholar]
- Singh, B.U.; Padmaia, P.G.; Seetharama, N. Biology and management of the sugarcane aphid, Melanaphis sacchari (Zehntner) (Homoptera: Aphididae), in sorghum: A review. Crop Prot. 2004, 23, 739–755. [Google Scholar] [CrossRef]
- Bowling, R.D.; Brewer, M.J.; Kerns, D.L.; Gordy, J.; Seiter, N.; Elliott, N.; Buntin, G.D.; Way, M.O.; Royer, T.A.; Biles, S.; et al. Sugarcane aphid (Hemiptera: Aphididae): A new pest on sorghum in North America. J. Integr. Pest Manag. 2016, 7, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, L.; Terán, A. Melanaphis sacchari (Hemiptera: Aphididae): A new sorghum insect pest in Mexico. Southwest. Entomol. 2015, 40, 433–434. [Google Scholar] [CrossRef]
- Field, L.M.; Devonshire, A.L.; Forde, B.G. Molecular evidence that insecticide resistance in peach-potato aphids (Myzus persicae Sulz.) results from amplification of an esterase gene. Biochem. J. 1988, 251, 309–312. [Google Scholar] [CrossRef]
- Sarwar, M. The killer chemicals as controller of agriculture insect pests: The conventional insecticides. Int. J. Chem. Sci. 2015, 1, 141–147. [Google Scholar]
- Vaňková, J.; Purrini, K. Natural epizooties caused by bacilli of the species Bacillus thuringiensis and Bacillus cereus. J. Appl. Entomol. 1979, 88, 216–221. [Google Scholar] [CrossRef]
- Porcar, M.; Grenier, A.; Federici, B.; Rahbe, Y. Effects of Bacillus thuringiensis endotoxins on the Pea Aphid (Acyrthosiphon pisum). Appl. Environ. Microbiol. 2009, 75, 4897–4900. [Google Scholar] [CrossRef]
- Palma, L.; Muñoz, D.; Berry, C.; Murillo, J.; de Escudero, I.R.; Caballero, P. Molecular and insecticidal characterization of a novel Cry-related protein from Bacillus thuringiensis toxic against Myzus persicae. Toxins 2014, 6, 3144–3156. [Google Scholar] [CrossRef]
- de Maagd, R.A.; Bravo, A.; Berry, C.; Crickmore, N.; Schnepf, H.E. Structure, diversity, and evolution of protein toxins from spore-forming entomopathogenic bacteria. Annu. Rev. Genet. 2003, 37, 409–433. [Google Scholar] [CrossRef]
- Alquisira, E.V.; Paredes, J.R.; Hernández, V.M.; Ramírez, J.A.; Peña, G. In vitro susceptibility of Varroa destructor and Apis mellifera to native strains of Bacillus thuringiensis. Apidologie 2014, 45, 707–718. [Google Scholar] [CrossRef]
- Ruiu, L.; Satta, A.; Floris, I. Emerging entomopathogenic bacteria for insect pest management. Bull. Insectology 2013, 66, 181–186. [Google Scholar]
- Shi, Y.; Zhang, X.; Lou, K. Isolation, characterization, and insecticidal activity of an endophyte of drunken horse grass, Achnatherum inebrians. J. Insect Sci. 2013, 13, 151. [Google Scholar] [CrossRef] [PubMed]
- Fisher, T.W.; Garczynski, S.F. Isolation, culture, preservation, and identification of entomopathogenic bacteria of the Bacilli. In Manual of Techniques in Invertebrate Pathology, 2nd ed.; Academic Press: New York, NY, USA, 2012; pp. 75–79. [Google Scholar]
- Tsagou, V.; Lianou, A.; Lazarakis, D.; Emmanouel, N.; Aggelis, G. Newly isolated bacterial strains belonging to Bacillaceae (Bacillus sp.) and Micrococcaceae accelerate death of the honey bee mite, Varroa destructor (V. jacobsoni), in laboratory assays. Biotechnol. Lett. 2004, 26, 529–532. [Google Scholar] [CrossRef] [PubMed]
- Bulla, L.A.; Rhodes, R.A.; Julian, G.S. Bacteria as insect pathogens. Annu. Rev. Microbiol. 1975, 29, 163–190. [Google Scholar] [CrossRef]
- Payne, J.R.; Cannon, R.J.C. Use of Bacillus thuringiensis isolates for controlling pests in the family Aphididae. US Patent 5,262,159, 16 November 1993. [Google Scholar]
- Lacey, L.A.; Grzywacz, D.; Shapiro, D.I.; Frutos, R.; Brownbridge, M.; Goettel, M.S. Insect pathogens as biological control agents: Back to the future. J. Invertebr. Pathol. 2015, 132, 1–41. [Google Scholar] [CrossRef]
- Li, H.; Chougule, N.P.; Bonning, B.C. Interaction of the Bacillus thuringiensis delta endotoxins Cry1Ac and Cry3Aa with the gut of the pea aphid, Acyrthosiphon pisum (Harris). J. Invertebr. Pathol. 2011, 107, 69–78. [Google Scholar] [CrossRef]
- Yao, Z.; Shuai, Z.; Jun-Yu, L.; Chun-Yi, W.; Li-Min, L.; Xiao-Ping, W.; Jin-Jie, C.; Chao-Liang, L. Bt proteins Cry1Ah and Cry2Ab do not affect cotton aphid Aphis gossypii and ladybeetle Propylea japonica. Sci. Rep. 2016, 6, 20368. [Google Scholar] [CrossRef]
- Jin, L.; Zhang, B.W.; Lu, J.W.; Liao, J.A.; Zhu, Q.J.; Lin, Y.; Yu, X.Q. The mechanism of Cry41-related toxin against Myzus persicae based on its interaction with Buchnera-derived ATP-dependent 6-phosphofructokinase. Pest Manag. Sci. 2023, 79, 1684–1691. [Google Scholar] [CrossRef]
- Chougule, N.P.; Bonning, B.C. Toxins for transgenic resistance to hemipteran pests. Toxins 2012, 4, 405–429. [Google Scholar] [CrossRef]
- Mishra, R.; Arora, A.K.; Jiménez, J.; Dos Santos, C.; Banerjee, R.; Panneerselvam, S.; Bonning, B.C. Bacteria-derived pesticidal proteins active against hemipteran pests. J. Invertebr. Pathol. 2022, 195, 107834. [Google Scholar] [CrossRef]
- Walters, F.S.; English, L.H. Toxicity of Bacillus thuringiensis δ-endotoxins toward the potato aphid in an artificial diet bioassay. Entomol. Exp. Appl. 1995, 77, 211–216. [Google Scholar] [CrossRef]
- Gerardo, N.M.; Altincicek, B.; Anselme, C.; Atamian, H.; Barribeau, S.M.; de Vos, M.; Duncan, E.J.; Evans, J.D.; Gabaldón, T.; Ghanim, M.; et al. Immunity and other defenses in pea aphids, Acyrthosiphon pisum. Genome Biol. 2010, 11, R21. [Google Scholar] [CrossRef]
- Oliver, K.M.; Degnan, P.H.; Burke, G.R.; Moran, N.A. Facultative symbionts in aphids and the horizontal transfer of ecologically important traits. Annu. Rev. Entomol. 2010, 55, 247–266. [Google Scholar] [CrossRef]
- Logan, N.A.; Vos, P.D. Bacillus. Bergey’s Manual of Systematics of Archaea and Bacteria; Whitman, W.B., Ed.; Wiley: Hoboken, NJ, USA, 2015; pp. 1–163. [Google Scholar] [CrossRef]
- Mandic, I.; Stefanic, P.; van Elsas, J.D. Ecology of Bacillaceae. Microbiol. Spectr. 2015, 3, 2. [Google Scholar] [CrossRef]
- Lambert, B.; Peferoen, M. Insecticidal promise of Bacillus thuringiensis. Facts and mysteries about a successful biopesticide. BioScience 1992, 42, 112–122. [Google Scholar] [CrossRef]
- Pigott, C.R.; Ellar, D.J. Role of receptors in Bacillus thuringiensis crystal toxin activity. Microbiol. Mol. Biol. Rev. 2007, 71, 255–281. [Google Scholar] [CrossRef] [PubMed]
- Frankenhuyzen, K.V. Insecticidal activity of Bacillus thuringiensis crystal proteins. J. Invertebr. Pathol. 2009, 101, 1–16. [Google Scholar] [CrossRef]
- Malik, K.; Riazuddin, S. Immunoassay-based approach for detection of novel Bacillus thuringiensis-endotoxins, entomocidal to cotton aphids (Aphis gossypii) and whiteflies (Bemisia tabaci). Pak. J. Bot. 2006, 38, 757–765. [Google Scholar]
- Monnerat, R.G.; Melatti, V.; Praça, L.; Martins, É.; Sujii, E.; Berry, C. Selection of Bacillus thuringiensis strains toxic against cotton aphid, Aphis gossypii Glover (Hemiptera: Aphididae). BioAssay 2010, 5, 2. [Google Scholar] [CrossRef]
- Alahyane, H.; Ouknin, M.; Alahyane, A.; Aboussaid, H.; Oufdou, K.; El Messoussi, S.; Mounir, A.; Majidi, L. Aphicidal activities of Moroccan Bacillus thuringiensis strains against cotton aphid (Aphis gossypii). Biointerface Res. Appl. Chem. 2021, 12, 3348–3356. [Google Scholar] [CrossRef]
- Rajashekhar, M.; Kalia, V.K. Native Bt strains efficacy against cotton aphid Aphis gossypii Glover. J. Pharmacogn. Phytochem. 2017, 6, 938–940. [Google Scholar]
- Ramasamy, A.; Suresh, M.; Mallesh, M.S.H. Toxicity evaluation of Aphidicidal crystalliferous toxins from Bacillus strains: A molecular study. Ann. Microbiol. 2020, 70, 214. [Google Scholar] [CrossRef]
- Torres, M.C.; Arenas, I.; Hernández, V.M.; Suárez, R.; Peña, G. Characterization of Bacillus thuringiensis (Bacillaceae) strains pathogenic to Myzus persicae (Hemiptera: Aphididae). Fla. Entomol. 2016, 99, 639–643. [Google Scholar] [CrossRef]
- López, G.; Alvarez, A.E.; Petroselli, G.; Erra, R.; Audisio, M.C. Aphicidal activity of Bacillus amyloliquefaciens strains in the peach-potato aphid (Myzus persicae). Microbiol. Res. 2019, 226, 41–47. [Google Scholar] [CrossRef]
- Sattar, S.; Biswas, P.K.; Hossain, M.A.; Maiti, M.K.; Sen, S.K.; Basu, A. Search for vegetative insecticidal proteins (VIPs) from local isolates of Bacillus thuringiensis effective against lepidopteran and homopteran insect pests. Biopestic. Int. 2008, 1, 216–222. [Google Scholar] [CrossRef]
- Baazeem, A.; Alotaibi, S.S.; Khalaf, L.K.; Kumar, U.; Zaynab, M.; Alharthi, S.; Darwish, H.; Alghamdi, A.; Jat, S.K.; Al-Barty, A.; et al. Identification and environment-friendly biocontrol potential of five different bacteria against Aphis punicae and Aphis illinoisensis (Hemiptera: Aphididae). Front. Microbiol. 2022, 13, 961349. [Google Scholar] [CrossRef]
- Yu, X.; Liu, T.; Liang, X.; Tang, C.; Zhu, J.; Wang, S.; Li, P. Rapid detection of vip1-type genes from Bacillus cereus and characterization of a novel vip binary toxin gene. FEMS Microbiol. Lett. 2011, 325, 30–36. [Google Scholar] [CrossRef]
- Sattar, S.; Maiti, M.K. Molecular characterization of a novel vegetative insecticidal protein from Bacillus thuringiensis effective against sap-sucking insect pest. Microbiol. Biotechn. 2011, 21, 937–946. [Google Scholar] [CrossRef]
- Loth, K.; Costechareyre, D.; Effantin, G.; Rahbé, Y.; Condemine, G.; Landon, C.; da Silva, P. New Cyt-like δ-endotoxins from Dickeya dadantii: Structure and aphicidal activity. Sci. Rep. 2015, 5, 8791. [Google Scholar] [CrossRef]
- Paula, D.P.; Andow, D.A. Differential Cry toxin detection and effect on Brevicoryne brassicae and Myzus persicae feeding on artificial diet. Entomol. Exp. Appl. 2016, 159, 54–60. [Google Scholar] [CrossRef]
- Rausch, M.A.; Chougule, N.P.; Deist, B.R.; Bonning, B.C. Modification of Cry4Aa toward improved toxin processing in the gut of the pea aphid, Acyrthosiphon pisum. PLoS ONE 2016, 11, e0155466. [Google Scholar] [CrossRef]
- Borman, E.K.; Stuart, C.A.; Wheeler, K.M. Taxonomy of the family Enterobacteriaceae. J. Bacteriol. 1944, 48, 351–367. [Google Scholar] [CrossRef]
- Jenkins, C.; Rentenaar, R.J.; Landraud, L.; Brisse, S. Enterobacteriaceae A2. In Infectious Diseases, 4th ed.; Elsevier: New York, NY, USA, 2017; Volume 2, pp. 1565–1578.e2. [Google Scholar]
- Kang, E.; Crouse, A.; Chevallier, L.; Pontier, S.M.; Alzahrani, A.; Silué, N.; Campbell-Valois, F.X.; Montagutelli, X.; Gruenheid, S.; Malo, D. Enterobacteria and host resitence to infection. Mamm. Gaenome 2018, 29, 558–576. [Google Scholar] [CrossRef]
- Harada, H.; Ishikawa, H. Experimental pathogenicity of Erwinia aphidicola to pea aphid, Acyrthosiphon pisum. J. Gen. Appl. Microbiol. 1997, 43, 363–367. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, Y. Study of the bacteria pathogenic for aphids, isolation of bacteria and identification of insecticidal compound. In Report of Hokkaido Prefectural Agricultural Experiment Stations; Kamikawa Agricultural Experiment Station: Asahikawa, Japan, 2002; Volume 102, pp. 1–48. [Google Scholar]
- Paliwal, D.; Hamilton, A.J.; Barrett, G.A.; Alberti, F.; van Emden, H.; Monteil, C.L.; Mauchline, T.H.; Nauen, R.; Wagstaff, C.; Bass, C.; et al. Identification of novel aphid-killing bacteria to protect plants. Microb. Biotechnol. 2022, 4, 1203–1220. [Google Scholar] [CrossRef]
- Stavrinides, J.; No, A.; Ochman, H. A single genetic locus in the phytopathogen Pantoea stewartii enables gut colonization and pathogenicity in an insect host. Environ. Microbiol. 2010, 12, 147–155. [Google Scholar] [CrossRef] [PubMed]
- Campillo, T.; Luna, E.; Portier, P.; Fischer, L.; Saux, M.; Lapitan, N.; Tisserat, N.A.; Leach, J.E. Erwinia iniecta sp. nov., isolated from Russian wheat aphid (Diuraphis noxia). Int. J. Syst. Evol. Microbiol. 2015, 65, 3625–3633. [Google Scholar] [CrossRef] [PubMed]
- Grenier, A.M.; Duport, G.; Pagès, S.; Condemine, G.; Rahbé, Y. The phytopathogen Dickeya dadantii (Erwinia chrysanthemi 3937) is a pathogen of the pea aphid. Appl. Environ. Microbiol. 2006, 72, 1956–1965. [Google Scholar] [CrossRef]
- Costechareyre, D.; Dridi, B.; Rahbé, Y.; Condemine, G. Cyt toxin expression reveals an inverse regulation of insect and plant virulence factors of Dickeya dadantii. Environ. Microbiol. 2010, 12, 3290–3301. [Google Scholar] [CrossRef]
- Costechareyre, D.; Balmand, S.; Condemine, G.; Rahbé, Y. Dickeya dadantii, a plant pathogenic bacterium producing cyt-like entomotoxins, causes septicemia in the pea aphid Acyrthosiphon pisum. PLoS ONE 2012, 7, e30702. [Google Scholar] [CrossRef]
- Costechareyre, D.; Chich, J.F.; Strub, J.M.; Rahbé, Y.; Condemine, G. Transcriptome of Dickeya dadantii infecting Acyrthosiphon pisum reveals a strong defense against antimicrobial peptides. PLoS ONE 2013, 8, e54118. [Google Scholar] [CrossRef]
- Renoz, F.; Noël, C.; Errachid, A.; Foray, V.; Hance, T. Infection dynamic of symbiotic bacteria in the pea aphid Acyrthosiphon pisum gut and host immune response at the early steps in the infection process. PLoS ONE 2015, 10, e0122099. [Google Scholar] [CrossRef]
- Wu, S.; Toews, M.D.; Cottrell, T.E. Toxicity of Photorhabdus luminescens and Xenorhabdus bovienii bacterial metabolites to pecan aphids (Hemiptera: Aphididae) and the lady beetle Harmonia axyridis (Coleoptera:Coccinellidae). J. Invertebr. Pathol. 2022, 194, 107806. [Google Scholar] [CrossRef] [PubMed]
- Altincicek, B.; Ter, B.; Laughton, A.M.; Udekwu, K.I.; Gerardo, N.M. Escherichia coli K-12 pathogenicity in the pea aphid, Acyrthosiphon pisum, reveals reduced antibacterial defense in aphids. Dev. Comp. Immunol. 2011, 35, 1091–1097. [Google Scholar] [CrossRef]
- Teixeira, L.M.; Merquior, V.L.C. The Family Moraxellaceae. In The Prokaryotes; Rosenberg, E., DeLong, E.F., Lory, S., Stackebrandt, E., Thompson, F., Eds.; Springer: Berlin/Heidelberg, Germany, 2014; pp. 254–263. [Google Scholar] [CrossRef]
- LaSala, P.R.; Segal, J.; Han, F.S.; Tarrand, J.J.; Han, X.Y. First reported infections caused by three newly described genera in the family Xanthomonadaceae. J. Clin. Microbiol. 2007, 45, 641–644. [Google Scholar] [CrossRef] [PubMed]
- An, S.Q.; Potnis, N.; Dow, M.; Vorhölter, F.J.; He, Y.Q.; Becker, A.; Teper, D.; Li, Y.; Wang, N.; Bleris, L.; et al. Mechanistic insights into host adaptation, virulence and epidemiology of the phytopathogen Xanthomonas. FEMS Microbiol. Lett. 2020, 44, 1–32. [Google Scholar] [CrossRef] [PubMed]
- Palleroni, N.J. Introduction to the Family Pseudomonadaceae. In The Prokaryotes; Starr, M.P., Stolp, H., Trüper, H.G., Balows, A., Schlegel, H.G., Eds.; Springer: Berlin/Heidelberg, Germany, 1981; pp. 655–665. [Google Scholar]
- Mnif, I.; Ghribi, D. Potential of bacterial derived biopesticides in pest management. Crop Prot. 2015, 77, 52–64. [Google Scholar] [CrossRef]
- Barahona, E.; Navazo, A.; Martínez, F.; Zea, T.; Pérez, R.M.; Martín, M.; Rivilla, R. Pseudomonas fluorescens F113 mutant with enhanced competitive colonization ability and improved biocontrol activity against fungal root pathogens. Appl. Environ. Microbiol. 2011, 77, 5412–5419. [Google Scholar] [CrossRef]
- Cronin, D.; Moenne, Y.; Fenton, A.; Dunne, C.; Dowling, D.N.; O’gara, F. Role of 2, 4-diacetylphloroglucinol in the interactions of the biocontrol Pseudomonad strain F113 with the potato cyst nematode Globodera rostochiensis. Appl. Environ. Microbiol. 1997, 63, 1357–1361. [Google Scholar] [CrossRef]
- Villacieros, M.; Power, B.; Sánchez, M.; Lloret, J.; Oruezabal, R.I.; Martín, M.; Rivilla, R. Colonization behaviour of Pseudomonas fluorescens and Sinorhizobium meliloti in the alfalfa (Medicago sativa) rhizosphere. Plant Soil 2003, 251, 47–54. [Google Scholar] [CrossRef]
- Manjula, T.R.; Kannan, G.S.; Sivasubramanian, P. Field efficacy of Pseudomonas fluorescens against the cotton aphid, Aphis gossypii Glover (Hemiptera: Aphididae) in Bt and non Bt cotton. Int. J. Curr. Microbiol. Appl. Sci. 2018, 6, 11–24. [Google Scholar]
- Paliwal, D.; Rabiey, M.; Mauchline, T.H.; Hassani-Pak, K.; Nauen, R.; Wagstaff, C.; Andrews, S.; Bass, C.; Jackson, R.W. Multiple toxins and a protease contribute to the aphid-killing ability of Pseudomonas fluorescens PpR24. Environ. Microbiol. 2024, 4, e16604. [Google Scholar] [CrossRef]
- Jansson, R.K.; Dybas, R.A. Avermectins: Biochemical mode of action, biological activity and agricultural importance. In Insecticides with Novel Modes of Action-Mechanisms and Application; Ishaaya, I., Degheele, D., Eds.; Springer: Berlin/Heidelberg, Germany, 1998; pp. 153–170. [Google Scholar]
- Fisher, M.H.; Mrozik, H. The chemistry and pharmacology of avermectins. Annu. Rev. Pharmacol. Toxicol. 1992, 32, 537–553. [Google Scholar] [CrossRef] [PubMed]
- Campbell, W.C. Ivermectin and Abamectin, 1st ed.; Springer: New York, NY, USA, 1989; p. 363. [Google Scholar]
- Kim, S.K.; Kim, Y.C.; Lee, S.; Kim, J.C.; Yun, M.Y.; Kim, I.S. Insecticidal activity of rhamnolipid isolated from Pseudomonas sp. EP-3 against green peach aphid (Myzus persicae). J. Agric. Food Chem. 2011, 59, 934–938. [Google Scholar] [CrossRef] [PubMed]
- Jang, J.Y.; Yang, S.Y.; Kim, Y.C.; Lee, C.W.; Park, M.S.; Kim, J.C.; Kim, I.S. Identification of orfamide A as an insecticidal metabolite produced by Pseudomonas protegens F6. J. Agric. Food Chem. 2013, 61, 6786–6791. [Google Scholar] [CrossRef]
- Xu, L.; Liang, K.; Duan, B.; Yu, M.; Meng, W.; Wang, Q.; Yu, Q. A novel insecticidal peptide SLP1 produced by Streptomyces laindensis H008 against Lipaphis erysimi. Molecules 2016, 21, 1101. [Google Scholar] [CrossRef]
- Yang, S.Y.; Lim, D.J.; Noh, M.Y.; Kim, J.C.; Kim, Y.C.; Kim, I.S. Characterization of biosurfactants as insecticidal metabolites produced by Bacillus subtilis Y9. Entomol. Res. 2016, 47, 55–59. [Google Scholar] [CrossRef]
- Lim, D.J.; Yang, S.Y.; Noh, M.Y.; Lee, C.W.; Kim, J.C.; Kim, I.S. Identification of lipopeptide xantholysins from Pseudomonas sp. DJ15 and their insecticidal activity against Myzus persicae. Entomol. Res. 2017, 47, 337–343. [Google Scholar] [CrossRef]
- Rumyantsev, S.D.; Alekseev, V.Y.; Sorokan, A.V.; Burkhanova, G.F.; Cherepanova, E.A.; Garafutdinov, R.R.; Maksimov, I.V.; Veselova, S.V. Additive Effect of the Composition of Endophytic Bacteria Bacillus subtilis on Systemic Resistance of Wheat against Greenbug Aphid Schizaphis graminum Due to Lipopeptides. Life 2023, 13, 214. [Google Scholar] [CrossRef]
- Kämpfer, P.; Glaeser, S.P.; Parkes, L.; van Keulen, G.; Dyson, P. The Family Streptomycetaceae. In The Prokaryotes; Rosenberg, E., DeLong, E.F., Lory, S., Stackebrandt, E., Thompson, F., Eds.; Springer: Berlin/Heidelberg, Germany, 2014; pp. 889–1010. [Google Scholar]
- Procópio, R.E.; Silva, I.R.; Martins, M.K.; Azevedo, J.L.; Araújo, J.M. Antibiotics produced by Streptomyces. Braz. J. Infect. Dis. 2012, 16, 466–471. [Google Scholar] [CrossRef]
- Aggarwal, N.; Thind, S.K.; Sharma, S. Role of secondary metabolites of actinomycetes in crop protection. In Plant Growth Promoting Actinobacteria; Subramaniam, G., Arumugam, S., Rajendran, V., Eds.; Springer: Singapore, 2016; pp. 99–121. [Google Scholar]
- Bérdy, J. Thoughts and facts about antibiotics: Where we are now and where we are heading. J. Antibiot. 2012, 65, 385–395. [Google Scholar] [CrossRef]
- Chen, Y.; Shafi, J.; Li, M.; Fu, D.; Ji, M. Insecticidal activity of endophytic actinomycetes isolated from Azadirachta indica against Myzus persicae. Arch. Biol. Sci. 2017, 70, 349–357. [Google Scholar] [CrossRef]
- Dewhirst, F.E.; Paster, B.J.; Bright, P.L. Chromobacterium, Eikenella, Kingella, Neisseria, Simonsiella, and Vitreoscilla Species Comprise a Major Branch of the beta Group Proteobacteria by 16S ribosomal ribonucleic acid sequence comparison: Transfer of Eikenella and Simonsiella to the family Neisseriaceae (emend.). Int. J. Syst. Evol. Microbiol. 1989, 39, 258–266. [Google Scholar] [CrossRef]
- Martin, P.A.W.; Gundersen, D.; Blackburn, M.; Buyer, J. Chromobacterium subtsugae sp. a betaproteobacterium toxic to Colorado potato beetle and other insect pests. Int. J. Syst. Evol. Microbiol. 2007, 57, 993–999. [Google Scholar] [CrossRef] [PubMed]
- Shapiro, D.I.; Cottrell, T.E.; Jackson, M.A.; Wood, B.W. Control of key pecan insect pests using biorational pesticides. J. Econ. Entomol. 2013, 106, 257–266. [Google Scholar] [CrossRef] [PubMed]
- Blackburn, M.B.; Sparks, M.E.; Gundersen, D.E. The genome of the insecticidal Chromobacterium subtsugae PRAA4-1 and its comparison with that of Chromobacterium violaceum ATCC 12472. Genom. Data 2016, 10, 1–3. [Google Scholar] [CrossRef]
- Rajendhran, J. Genomic insights into Brucella. Infect. Genet. Evol. 2021, 87, 104635. [Google Scholar] [CrossRef]
- Hemme, D.; Foucaud-Scheunemann, C. Leuconostoc, characteristics, use in dairy technology and prospects in functional foods. Int. Dairy J. 2004, 14, 467–494. [Google Scholar] [CrossRef]
- Hiebert, N.; Kessel, T.; Skaljac, M.; Spohn, M.; Vilcinskas, A.; Lee, K.Z. The Gram-Positive Bacterium Leuconostoc pseudomesenteroides Shows Insecticidal Activity against Drosophilid and Aphid Pests. Insects 2020, 11, 471. [Google Scholar] [CrossRef]
- Rosell, R.C.; Davidson, E.W.; Jancovich, J.K.; Hendrix, D.L.; Brown, J.K. Size Limitations in the Filter Chamber and Digestive Tract of Nymphal and Adult Bemisia tabaci Whiteflies (Hemiptera: Aleyrodidae). Ann. Entomol. Soc. Am. 2003, 96, 544–552. [Google Scholar] [CrossRef]
- Shakesby, A.J.; Wallace, I.S.; Isaacs, H.V.; Pritchard, J.; Roberts, D.M.; Douglas, A.E. A water-specific aquaporin involved in aphid osmoregulation. Insect Biochem. Mol. Biol. 2009, 39, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Hayakawa, T.; Yasuyuki, S.; Kazuhisa, M.; Hidetaka, H. GalNAc pretreatment inhibits trapping of Bacillus thuringiensis Cry1Ac on the peritrophic membrane of Bombyx mori. FEBS Lett. 2004, 576, 331–335. [Google Scholar] [CrossRef]
- Terra, W.R.; Barroso, I.G.; Dias, R.O.; Ferreira, C. Molecular physiology of insect midgut. In Advances in Insect Physiology; Academic Press: Cambridge, MA, USA, 2019; Volume 56, pp. 117–163. [Google Scholar] [CrossRef]
- Terra, W.R.; Ferreira, C. Evolutionary trends of digestion and absorption in the major insect orders. Arthropod Struct. Dev. 2020, 56, 100931. [Google Scholar] [CrossRef]
- Brandt, S.L.; Coudron, T.A.; Habibi, J.; Brown, G.R.; Ilagan, O.M.; Wagner, R.M.; Wright, M.K.; Backus, E.A.; Huesing, J.E. Interaction of two Bacillus thuringiensis δ-endotoxins with the digestive system of Lygus hesperus. Curr. Microbiol. 2004, 48, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Cristofoletti, P.T.; Ribeiro, A.F.; Deraison, C.; Rahbé, Y.; Terra, W.R. Midgut adaptation and digestive enzyme distribution in a phloem feeding insect, the pea aphid Acyrthosiphon pisum. Insect Physiol. 2003, 49, 11–24. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.D.; Zhang, B.W.; Fu, L.J.; Li, Q.L.; Lin, Y.; Yu, X.Q. Possible Insecticidal Mechanism of Cry41-Related Toxin against Myzus persicae by Enhancing Cathepsin B Activity. J. Agric. Food Chem. 2020, 68, 4607–4615. [Google Scholar] [CrossRef]
- Feng, H.; Edwards, N.; Anderson, C.; Althaus, M.; Duncan, R.; Hsu, Y. Trading amino acids at the aphid-Buchnera symbiotic interface. Proc. Natl. Acad. Sci. USA 2019, 116, 16003–16011. [Google Scholar] [CrossRef]
- Wilson, A.C.C.; Ashton, P.D.; Calevro, F.; Charles, H.; Colella, S.; Febvay, G. Genomic insight into the amino acid relations of the pea aphid, Acyrthosiphon pisum, with its symbiotic bacterium Buchnera aphidicola. Insect Mol. Biol. 2010, 19, 249–258. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
| Target Aphid | Protein | % Mortality | ng/mL (Time) | Origin | Reference |
|---|---|---|---|---|---|
| Potato aphid Macrosiphum euphorbiae | Mixture CryI; CryIA(a), CryIA(b), CryIC and CryIF CryIIA CryIIIA CryIVD | 100 ± 0 93 ± 0 98 ± 4 93 ± 10 | 100 each Cry (4 days) 200 (4 days) 360 (4 days) 350 (4 days) | Recombinant strains of Bt | [25] |
| Pea aphid Acyrthosiphon pisum | CryIAb Cry3A Cry4A and Cry4B Cry11A CytIA | 35 60 100 100 Growth inhibition | 500 (5 days) 500 (6 days) 500 (4 days) 500 (3 days) 125 (7 days) | Recombinant strains Bt subsp. israelensis | [9] |
| Cotton aphid Aphis gossypii | VipIAcI and Vip2Ae3 | LC50 | 0.0875 (NR days) | Bacillus cereus | [42] |
| Pea aphid Acyrthosiphon pisum | CryIAc Cry3Aa | 71 71 | 500 (7 days) | Bacillus thuringiensis | [20] |
| Cotton aphid Aphis gossypii | VipIAe and Vip2Ae | LC50 | 0.576 (2 days) | Bacillus thuringiensis | [43] |
| Pea aphid Acyrthosiphon pisum | Cyt2Aa CGAL1 CGAL3 CGAL4 CGSL1 CGSL4 | LC50 LC50 LC50 LC50 LC50 LC50 | 150 ± 0.00 19.71 ± 5.74 9.55 ± 2.54 11.92 ± 1.99 28.74 ± 2.92 15.13 ± 0.23 | Recombinant proteins | [23] |
| Green peach aphid Myzus persicae | Cyt2Aa CGAL1 CGAL3 CGAL4 CGSL1 CGSL4 | LC50 LC50 LC50 LC50 LC50 LC50 | 150 ± 0.00 58.04 ± 2.08 42.68 ± 0.49 92.75 ± 2.54 ND ND | Bacillus thuringiensis Recombinant proteins | [23] |
| Green peach aphid Myzus persicae | Cry-Related | LC50 | 32.7 (3 days) | Bacillus thuringiensis | [10] |
| Pea aphid Acyrthosiphon pisum | CytIA CytC CytB CytA | TL50 TL50 TL50 TL50 | 1000 (3.24 days) 1000 (10.1 days) 500 (5.1 days) 1000 (2.28 days) | Dickeya dadantii | [44] |
| Cotton aphid Aphis gossypii | Cry1Ah Cry2Ab | No mortality | >1000 | Bacillus thuringiensis | [21] |
| Cabbage aphid Brevicoryne brassicae Green peach aphid Myzus persicae | Cry1Ac Cry1F Cry1Ac and Cry1F Cry1Ac Cry1F Cry1Ac and Cry1F | Decreased the net population growth rate Decreased the net population growth rate | 20 (3 days) 20 (3 days) 20 (3 days) 20 (3 days) 20 (3 days) 20 (3 days) | Bacillus sp. | [45] |
| Pea aphid Acyrthosiphon pisum | Cry4Aa (trypsin activated) Cry4Aa 2A | 63.3 ± 24.5 51.1 ± 2.2 | 120 (2 days) 120 (2 days) | Bacillus thuringiensis Modified toxin | [46] |
| Green peach aphid Myzus persicae | Cry1Cb2 | LC50 | 6.58 (3 days) | Bacillus sp. | [38] |
| Target Aphid | Molecule | % Mortality, or LC50-90 | μg/mL (Time) | Origin | Reference |
| Pea aphid Acyrthosiphon pisum | Avermectin B1 | LC90 | 0.4 ppm (72–96 h) | Streptomyces avermitilis | [72,73] |
| Cotton aphid Aphis gossypii Black bean aphid Aphis fabae | Avermectin B1 | 50% | 450 ppm (NM) | Streptomyces avermitilis | [74] |
| Cotton aphid Aphis gossypii | Viscosin | 90–99% | 200 ppm (6 days) | Pseudomonas fluorescens | [51] |
| Green peach aphid Myzus persicae | Dirhamnolipid | 100% | 100 μg/mL (24 h) | Pseudomonas sp. | [75] |
| Green peach aphid Myzus persicae | Orfamide A | LC50 | 34.5 μg/mL (24 h) | Pseudomonas protegens | [76] |
| Green peach aphid Myzus persicae | Surfactin | LC50 | 35.82 μg/mL (24 h) | Bacillus amyloliquefaciens | [42] |
| Mustard aphid Lipaphis erysimi | Peptide | 100% | 700 μg/mL (48 h) | Streptomyces laindensis | [77] |
| Green peach aphid Myzus persicae | Surfactin isomers | LC50 | 20.4 μg/mL 22.2 μg/mL 54.5 μg/mL (24 h) | Bacillus subtilis | [78] |
| Green peach aphid Myzus persicae | Xantholysins A and B | LC50 | 13.4 μg/mL 24.6 μg/mL (24 h) | Pseudomonas sp. | [79] |
| Green peach aphid Myzus persicae | Kurstakins, Surfactins, Iturins, Fngycins | No mortality | ND | Bacillus amyloliquefaciens strains CBMDDrag3, PGPBacCA2 | [39] |
| Greenbug Aphid Schizaphis graminum | Surfactin Iturin lipopeptides | 100% Using a mix of both strains | 112 μg/mL (5 days) | Bacillus subtilis strains 26D and 11VM | [80,81] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Toledo-Hernández, E.; Torres-Quíntero, M.C.; Mancilla-Dorantes, I.; Sotelo-Leyva, C.; Delgado-Núñez, E.J.; Hernández-Velázquez, V.M.; Dunstand-Guzmán, E.; Salinas-Sánchez, D.O.; Peña-Chora, G. Entomopathogenic Bacteria Species and Toxins Targeting Aphids (Hemiptera: Aphididae): A Review. Plants 2025, 14, 943. https://doi.org/10.3390/plants14060943
Toledo-Hernández E, Torres-Quíntero MC, Mancilla-Dorantes I, Sotelo-Leyva C, Delgado-Núñez EJ, Hernández-Velázquez VM, Dunstand-Guzmán E, Salinas-Sánchez DO, Peña-Chora G. Entomopathogenic Bacteria Species and Toxins Targeting Aphids (Hemiptera: Aphididae): A Review. Plants. 2025; 14(6):943. https://doi.org/10.3390/plants14060943
Chicago/Turabian StyleToledo-Hernández, Erubiel, Mary Carmen Torres-Quíntero, Ilse Mancilla-Dorantes, César Sotelo-Leyva, Edgar Jesús Delgado-Núñez, Víctor Manuel Hernández-Velázquez, Emmanuel Dunstand-Guzmán, David Osvaldo Salinas-Sánchez, and Guadalupe Peña-Chora. 2025. "Entomopathogenic Bacteria Species and Toxins Targeting Aphids (Hemiptera: Aphididae): A Review" Plants 14, no. 6: 943. https://doi.org/10.3390/plants14060943
APA StyleToledo-Hernández, E., Torres-Quíntero, M. C., Mancilla-Dorantes, I., Sotelo-Leyva, C., Delgado-Núñez, E. J., Hernández-Velázquez, V. M., Dunstand-Guzmán, E., Salinas-Sánchez, D. O., & Peña-Chora, G. (2025). Entomopathogenic Bacteria Species and Toxins Targeting Aphids (Hemiptera: Aphididae): A Review. Plants, 14(6), 943. https://doi.org/10.3390/plants14060943