
The Kinetics of Mesophyll Conductance and Photorespiration During Light Induction


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
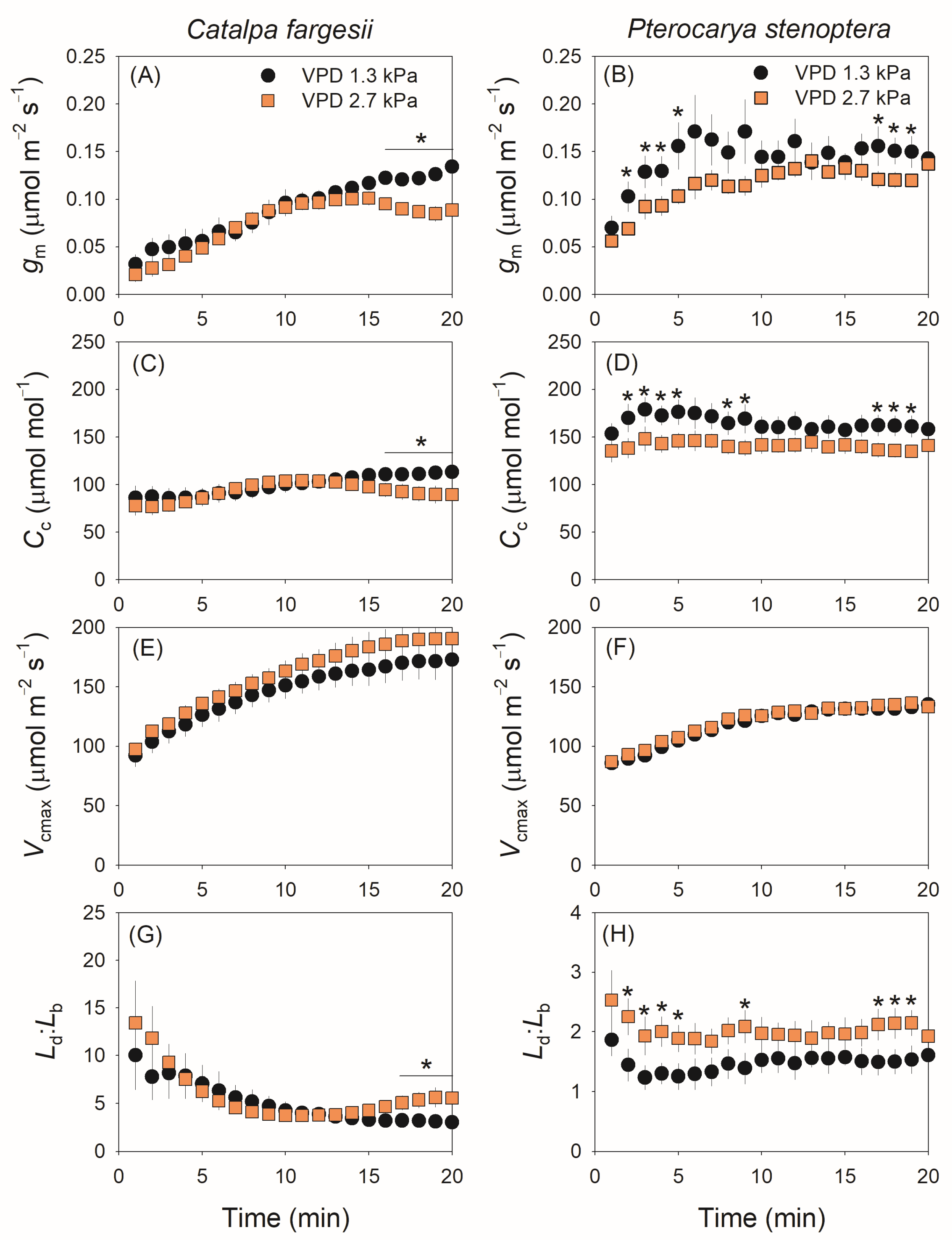
2.1. The Response of The Induction Kinetics of CO2 Assimilation, gs and gm to VPD
2.2. Correlation Between gs and gm During Light Induction
2.3. The Time-Integrated Limitations of AN During Light Induction
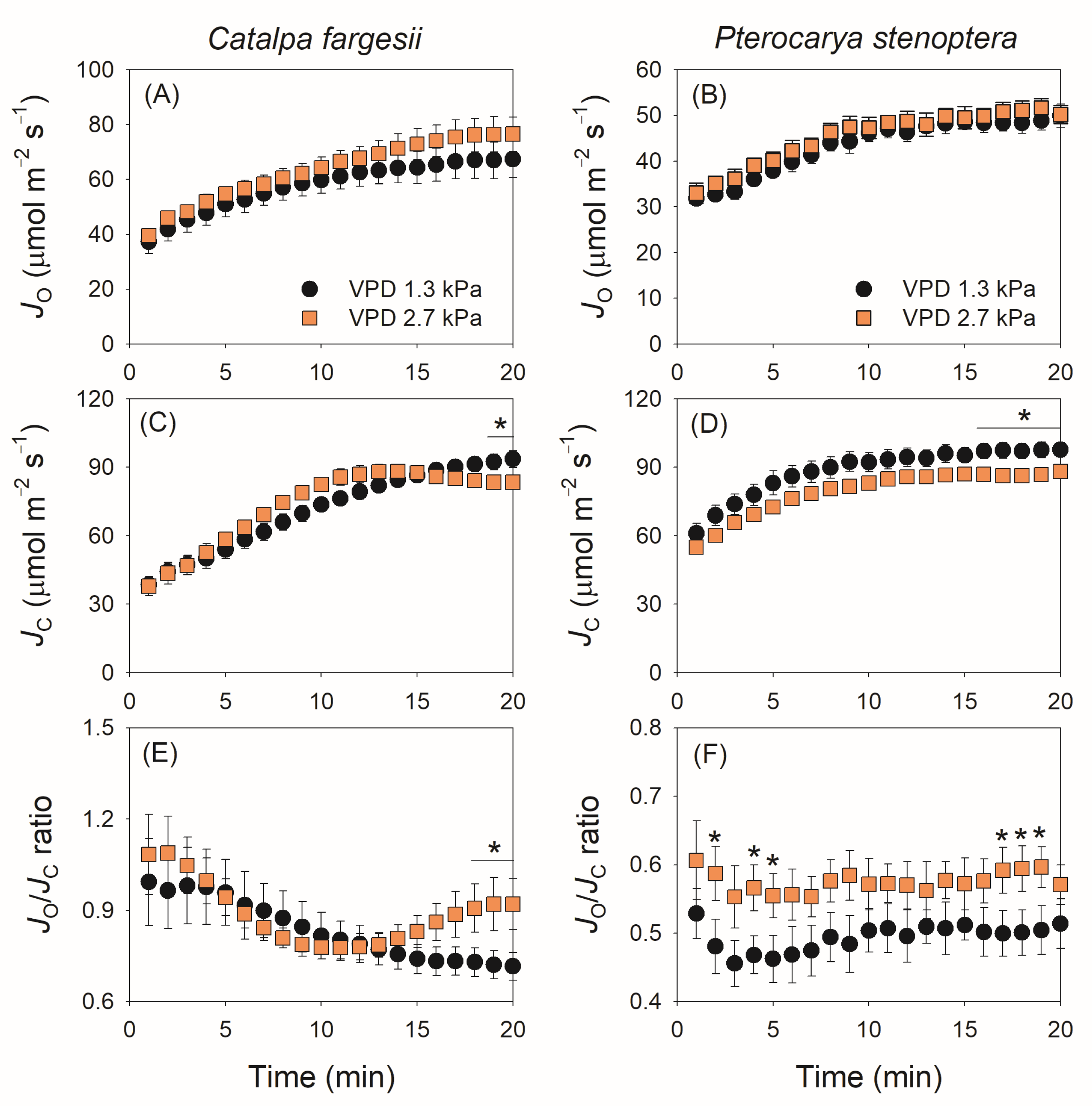
2.4. Induction Kinetics of Photorespiration
3. Discussion
3.1. Relationship Between gs and gm During Light Induction Is Species-Dependent
3.2. Differential Effects of VPD on gm Kinetics During Light Induction
3.3. Modulation of Photorespiration in Response to Photosynthetic Limitation
4. Materials and Methods
4.1. Plant Materials and Growing Conditions
4.2. Gas Exchange and Chlorophyll Fluorescence Measurements
4.3. Calculation of the Mesophyll Conductance
4.4. Quantitative Calculation of Photosynthetic Limitation
4.5. Calculation of Electron Flow for Photorespiration
4.6. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Slattery, R.A.; Walker, B.J.; Weber, A.P.M.; Ort, D.R. The Impacts of Fluctuating Light on Crop Performance. Plant Physiol. 2018, 176, 990–1003. [Google Scholar] [CrossRef] [PubMed]
- Roden, J.S.; Pearcy, R.W. Photosynthetic Gas Exchange Response of Poplars to Steady-State and Dynamic Light Environments. Oecologia 1993, 93, 208–214. [Google Scholar] [CrossRef]
- Pearcy, R.W. Sunflecks and Photosynthesis in Plant Canopies. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1990, 41, 421–453. [Google Scholar] [CrossRef]
- Liu, T.; Barbour, M.M.; Yu, D.; Rao, S.; Song, X. Mesophyll Conductance Exerts a Significant Limitation on Photosynthesis during Light Induction. New Phytol. 2022, 233, 360–372. [Google Scholar] [CrossRef]
- Kaiser, E.; Kromdijk, J.; Harbinson, J.; Heuvelink, E.; Marcelis, L.F.M. Photosynthetic Induction and Its Diffusional, Carboxylation and Electron Transport Processes as Affected by CO2 Partial Pressure, Temperature, Air Humidity and Blue Irradiance. Ann. Bot. 2017, 119, 191–205. [Google Scholar] [CrossRef]
- Kaiser, E.; Morales, A.; Harbinson, J. Fluctuating Light Takes Crop Photosynthesis on a Rollercoaster Ride. Plant Physiol. 2018, 176, 977–989. [Google Scholar] [CrossRef]
- Way, D.A.; Pearcy, R.W. Sunflecks in Trees and Forests: From Photosynthetic Physiology to Global Change Biology. Tree Physiol. 2012, 32, 1066–1081. [Google Scholar] [CrossRef] [PubMed]
- Kimura, H.; Hashimoto-Sugimoto, M.; Iba, K.; Terashima, I.; Yamori, W. Improved Stomatal Opening Enhances Photosynthetic Rate and Biomass Production in Fluctuating Light. J. Exp. Bot. 2020, 71, 2339–2350. [Google Scholar] [CrossRef]
- Kromdijk, J.; Głowacka, K.; Leonelli, L.; Gabilly, S.T.; Iwai, M.; Niyogi, K.K.; Long, S.P. Improving Photosynthesis and Crop Productivity by Accelerating Recovery from Photoprotection. Science 2016, 354, 857–861. [Google Scholar] [CrossRef]
- De Souza, A.P.; Burgess, S.J.; Doran, L.; Hansen, J.; Manukyan, L.; Maryn, N.; Gotarkar, D.; Leonelli, L.; Niyogi, K.K.; Long, S.P. Soybean Photosynthesis and Crop Yield Are Improved by Accelerating Recovery from Photoprotection. Science 2022, 377, 851–854. [Google Scholar] [CrossRef]
- Basso, L.; Sakoda, K.; Kobayashi, R.; Yamori, W.; Shikanai, T. Flavodiiron Proteins Enhance the Rate of CO2 Assimilation in Arabidopsis under Fluctuating Light Intensity. Plant Physiol. 2022, 189, 375–387. [Google Scholar] [CrossRef] [PubMed]
- Long, S.P.; Taylor, S.H.; Burgess, S.J.; Carmo-Silva, E.; Lawson, T.; De Souza, A.P.; Leonelli, L.; Wang, Y. Into the Shadows and Back into Sunlight: Photosynthesis in Fluctuating Light. Annu. Rev. Plant Biol. 2022, 73, 617–648. [Google Scholar] [CrossRef]
- Yamori, W.; Kusumi, K.; Iba, K.; Terashima, I. Increased Stomatal Conductance Induces Rapid Changes to Photosynthetic Rate in Response to Naturally Fluctuating Light Conditions in Rice. Plant. Cell Environ. 2020, 43, 1230–1240. [Google Scholar] [CrossRef]
- Acevedo-Siaca, L.G.; Coe, R.; Wang, Y.; Kromdijk, J.; Quick, W.P.; Long, S.P. Variation in Photosynthetic Induction between Rice Accessions and Its Potential for Improving Productivity. New Phytol. 2020, 227, 1097–1108. [Google Scholar] [CrossRef]
- Salter, W.T.; Merchant, A.M.; Richards, R.A.; Trethowan, R.; Buckley, T.N. Rate of Photosynthetic Induction in Fluctuating Light Varies Widely among Genotypes of Wheat. J. Exp. Bot. 2019, 70, 2787–2796. [Google Scholar] [CrossRef]
- Soleh, M.A.; Tanaka, Y.; Nomoto, Y.; Iwahashi, Y.; Nakashima, K.; Fukuda, Y.; Long, S.P.; Shiraiwa, T. Factors Underlying Genotypic Differences in the Induction of Photosynthesis in Soybean [Glycine max (L.) Merr.]. Plant Cell Environ. 2016, 39, 685–693. [Google Scholar] [CrossRef]
- De Souza, A.P.; Wang, Y.; Orr, D.J.; Carmo-Silva, E.; Long, S.P. Photosynthesis across African Cassava Germplasm Is Limited by Rubisco and Mesophyll Conductance at Steady State, but by Stomatal Conductance in Fluctuating Light. New Phytol. 2020, 225, 2498–2512. [Google Scholar] [CrossRef] [PubMed]
- Yoshiyama, Y.; Wakabayashi, Y.; Mercer, K.L.; Kawabata, S.; Kobayashi, T.; Tabuchi, T.; Yamori, W. Natural Genetic Variation in Dynamic Photosynthesis Is Correlated with Stomatal Anatomical Traits in Diverse Tomato Species across Geographical Habitats. J. Exp. Bot. 2024, 75, 6762–6777. [Google Scholar] [CrossRef] [PubMed]
- Yamori, W.; Makino, A.; Shikanai, T. A Physiological Role of Cyclic Electron Transport around Photosystem I in Sustaining Photosynthesis under Fluctuating Light in Rice. Sci. Rep. 2016, 6, 20147. [Google Scholar] [CrossRef]
- Sakoda, K.; Yamori, W.; Groszmann, M.; Evans, J.R. Stomatal, Mesophyll Conductance, and Biochemical Limitations to Photosynthesis during Induction. Plant Physiol. 2021, 185, 146–160. [Google Scholar] [CrossRef]
- Yamori, W.; Masumoto, C.; Fukayama, H.; Makino, A. Rubisco Activase Is a Key Regulator of Non-Steady-State Photosynthesis at Any Leaf Temperature and, to a Lesser Extent, of Steady-State Photosynthesis at High Temperature. Plant J. 2012, 71, 871–880. [Google Scholar] [CrossRef] [PubMed]
- Evans, J.R.; Kaldenhoff, R.; Genty, B.; Terashima, I. Resistances along the CO2 Diffusion Pathway inside Leaves. J. Exp. Bot. 2009, 60, 2235–2248. [Google Scholar] [CrossRef] [PubMed]
- Ögren, E.; Sundin, U. Photosynthetic Responses to Variable Light: A Comparison of Species from Contrasting Habitats. Oecologia 1996, 106, 18–27. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Peng, S.; Li, Y. Increase Rate of Light-Induced Stomatal Conductance Is Related to Stomatal Size in the Genus Oryza. J. Exp. Bot. 2019, 70, 5259–5269. [Google Scholar] [CrossRef]
- Xiong, D.; Douthe, C.; Flexas, J. Differential Coordination of Stomatal Conductance, Mesophyll Conductance, and Leaf Hydraulic Conductance in Response to Changing Light across Species. Plant. Cell Environ. 2018, 41, 436–450. [Google Scholar] [CrossRef]
- Lawson, T.; Blatt, M.R. Stomatal Size, Speed, and Responsiveness Impact on Photosynthesis and Water Use Efficiency. Plant Physiol. 2014, 164, 1556–1570. [Google Scholar] [CrossRef]
- Faralli, M.; Matthews, J.; Lawson, T. Exploiting Natural Variation and Genetic Manipulation of Stomatal Conductance for Crop Improvement. Curr. Opin. Plant Biol. 2019, 49, 1–7. [Google Scholar] [CrossRef]
- McAusland, L.; Vialet-Chabrand, S.; Davey, P.; Baker, N.R.; Brendel, O.; Lawson, T. Effects of Kinetics of Light-induced Stomatal Responses on Photosynthesis and Water-use Efficiency. New Phytol. 2016, 211, 1209–1220. [Google Scholar] [CrossRef]
- Eyland, D.; van Wesemael, J.; Lawson, T.; Carpentier, S. The Impact of Slow Stomatal Kinetics on Photosynthesis and Water Use Efficiency under Fluctuating Light. Plant Physiol. 2021, 186, 998–1012. [Google Scholar] [CrossRef]
- Flexas, J.; Diaz-Espejo, A.; Galmés, J.; Kaldenhoff, R.; Medrano, H.; Ribas-Carbo, M. Rapid Variations of Mesophyll Conductance in Response to Changes in CO2 Concentration around Leaves. Plant Cell Environ. 2007, 30, 1284–1298. [Google Scholar] [CrossRef]
- Douthe, C.; Dreyer, E.; Epron, D.; Warren, C.R. Mesophyll Conductance to CO2, Assessed from Online TDL-AS Records of 13CO2 Discrimination, Displays Small but Significant Short-Term Responses to CO2 and Irradiance in Eucalyptus Seedlings. J. Exp. Bot. 2011, 62, 5335–5346. [Google Scholar] [CrossRef]
- Douthe, C.; Dreyer, E.; Brendel, O.; Warren, C.R. Is Mesophyll Conductance to CO2 in Leaves of Three Eucalyptus Species Sensitive to Short-Term Changes of Irradiance under Ambient as Well as Low O2? Funct. Plant Biol. 2012, 39, 435–448. [Google Scholar] [CrossRef]
- Xiong, D.; Liu, X.; Liu, L.; Douthe, C.; Li, Y.; Peng, S.; Huang, J. Rapid Responses of Mesophyll Conductance to Changes of CO2 Concentration, Temperature and Irradiance Are Affected by N Supplements in Rice. Plant Cell Environ. 2015, 38, 2541–2550. [Google Scholar] [CrossRef]
- Campany, C.E.; Tjoelker, M.G.; von Caemmerer, S.; Duursma, R.A. Coupled Response of Stomatal and Mesophyll Conductance to Light Enhances Photosynthesis of Shade Leaves under Sunflecks. Plant Cell Environ. 2016, 39, 2762–2773. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Shi, Q.; Liu, N.-Y.; Zhang, S.-B.; Huang, W. Drought Stress Delays Photosynthetic Induction and Accelerates Photoinhibition under Short-Term Fluctuating Light in Tomato. Plant Physiol. Biochem. 2023, 196, 152–161. [Google Scholar] [CrossRef]
- Zeng, Z.-L.; Wang, X.-Q.; Zhang, S.-B.; Huang, W. Mesophyll Conductance Limits Photosynthesis in Fluctuating Light under Combined Drought and Heat Stresses. Plant Physiol. 2024, 194, 1498–1511. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Zhang, Y.-Q.; Zhang, S.; Huang, W. Photosynthetic Induction Under Fluctuating Light Is Affected by Leaf Nitrogen Content in Tomato. Front. Plant Sci. 2022, 13, 835571. [Google Scholar] [CrossRef]
- Merilo, E.; Yarmolinsky, D.; Jalakas, P.; Parik, H.; Tulva, I.; Rasulov, B.; Kilk, K.; Kollist, H. Stomatal VPD Response: There Is More to the Story Than ABA. Plant Physiol. 2018, 176, 851–864. [Google Scholar] [CrossRef] [PubMed]
- Sancho-Knapik, D.; Mendoza-Herrer, Ó.; Alonso-Forn, D.; Saz, M.Á.; Martín-Sánchez, R.; dos Santos Silva, J.V.; Ogee, J.; Peguero-Pina, J.J.; Gil-Pelegrín, E.; Ferrio, J.P. Vapor Pressure Deficit Constrains Transpiration and Photosynthesis in Holm Oak: A Comparison of Three Methods during Summer Drought. Agric. For. Meteorol. 2022, 327, 109218. [Google Scholar] [CrossRef]
- Shi, Q.; Wang, X.; He, B.; Yang, Y.; Huang, W. Differential Impact of Decreasing Relative Humidity on Photosynthesis under Fluctuating Light between Maize and Tomato. Physiol. Plant 2024, 176, e14179. [Google Scholar] [CrossRef]
- Liu, N.-Y.; Yang, Q.-Y.; Wang, J.-H.; Zhang, S.-B.; Yang, Y.-J.; Huang, W. Differential Effects of Increasing Vapor Pressure Deficit on Photosynthesis at Steady State and Fluctuating Light. J. Plant Growth Regul. 2024, 43, 2329–2339. [Google Scholar] [CrossRef]
- Kanazawa, A.; Kramer, D.M. In Vivo Modulation of Nonphotochemical Exciton Quenching (NPQ) by Regulation of the Chloroplast ATP Synthase. Proc. Natl. Acad. Sci. USA 2002, 99, 12789–12794. [Google Scholar] [CrossRef] [PubMed]
- Smith, K.; Strand, D.D.; Kramer, D.M.; Walker, B.J. The Role of Photorespiration in Preventing Feedback Regulation via ATP Synthase in Nicotiana Tabacum. Plant Cell Environ. 2024, 47, 416–428. [Google Scholar] [CrossRef] [PubMed]
- Timm, S.; Sun, H.; Huang, W. Photorespiration—Emerging Insights into Photoprotection Mechanisms. Trends Plant Sci. 2024, xx, 1–4. [Google Scholar] [CrossRef]
- Shi, Q.; Sun, H.; Timm, S.; Zhang, S.; Huang, W. Photorespiration Alleviates Photoinhibition of Photosystem I under Fluctuating Light in Tomato. Plants 2022, 11, 195. [Google Scholar] [CrossRef]
- Huang, W.; Hu, H.; Zhang, S.-B. Photorespiration Plays an Important Role in the Regulation of Photosynthetic Electron Flow under Fluctuating Light in Tobacco Plants Grown under Full Sunlight. Front. Plant Sci. 2015, 6, 621. [Google Scholar] [CrossRef]
- Walker, B.J.; VanLoocke, A.; Bernacchi, C.J.; Ort, D.R. The Costs of Photorespiration to Food Production Now and in the Future. Annu. Rev. Plant Biol. 2016, 67, 107–129. [Google Scholar] [CrossRef]
- Loriaux, S.D.; Avenson, T.J.; Welles, J.M.; Mcdermitt, D.K.; Eckles, R.D.; Riensche, B.; Genty, B. Closing in on Maximum Yield of Chlorophyll Fluorescence Using a Single Multiphase Flash of Sub-Saturating Intensity. Plant Cell Environ. 2013, 36, 1755–1770. [Google Scholar] [CrossRef]
- Krall, J.P.; Edwards, G.E. Relationship between Photosystem II Activity and CO2 Fixation in Leaves. Physiol. Plant. 1992, 86, 180–187. [Google Scholar] [CrossRef]
- Yin, X.; Struik, P.C.; Romero, P.; Harbinson, J.; Evers, J.B.; VAN DER Putten, P.E.L.; Vos, J. Using Combined Measurements of Gas Exchange and Chlorophyll Fluorescence to Estimate Parameters of a Biochemical C Photosynthesis Model: A Critical Appraisal and a New Integrated Approach Applied to Leaves in a Wheat (Triticum aestivum) Canopy. Plant Cell Environ. 2009, 32, 448–464. [Google Scholar] [CrossRef]
- Harley, P.C.; Loreto, F.; Di Marco, G.; Sharkey, T.D. Theoretical Considerations When Estimating the Mesophyll Conductance to CO2 Flux by Analysis of the Response of Photosynthesis to CO2. Plant Physiol. 1992, 98, 1429–1436. [Google Scholar] [CrossRef] [PubMed]
- Yamori, W.; Evans, J.R.; Von Caemmerer, S. Effects of Growth and Measurement Light Intensities on Temperature Dependence of CO2 Assimilation Rate in Tobacco Leaves. Plant Cell Environ. 2010, 33, 332–343. [Google Scholar] [CrossRef]
- Warren, C.R.; Dreyer, E. Temperature Response of Photosynthesis and Internal Conductance to CO2: Results from Two Independent Approaches. J. Exp. Bot. 2006, 57, 3057–3067. [Google Scholar] [CrossRef] [PubMed]
- Moualeu-Ngangue, D.P.; Chen, T.; Stützel, H. A New Method to Estimate Photosynthetic Parameters through Net Assimilation Rate-Intercellular Space CO2 Concentration (A-Ci ) Curve and Chlorophyll Fluorescence Measurements. New Phytol. 2017, 213, 1543–1554. [Google Scholar] [CrossRef] [PubMed]
- Grassi, G.; Magnani, F. Stomatal, Mesophyll Conductance and Biochemical Limitations to Photosynthesis as Affected by Drought and Leaf Ontogeny in Ash and Oak Trees. Plant. Cell Environ. 2005, 28, 834–849. [Google Scholar] [CrossRef]
- Valentini, R.; Epron, D.; Angelis, P.D.E.; Matteucci, G.; Dreyer, E. In Situ Estimation of Net CO2 Assimilation, Photosynthetic Electron Flow and Photorespiration in Turkey Oak (Q. cerris L.) Leaves: Diurnal Cycles under Different Levels of Water Supply. Plant Cell Environ. 1995, 18, 631–640. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, N.; Cao, J.; Yang, M.; Li, Y.; Huang, W. The Kinetics of Mesophyll Conductance and Photorespiration During Light Induction. Plants 2025, 14, 850. https://doi.org/10.3390/plants14060850
Liu N, Cao J, Yang M, Li Y, Huang W. The Kinetics of Mesophyll Conductance and Photorespiration During Light Induction. Plants. 2025; 14(6):850. https://doi.org/10.3390/plants14060850
Chicago/Turabian StyleLiu, Ningyu, Jianxin Cao, Mingying Yang, Yiyun Li, and Wei Huang. 2025. "The Kinetics of Mesophyll Conductance and Photorespiration During Light Induction" Plants 14, no. 6: 850. https://doi.org/10.3390/plants14060850
APA StyleLiu, N., Cao, J., Yang, M., Li, Y., & Huang, W. (2025). The Kinetics of Mesophyll Conductance and Photorespiration During Light Induction. Plants, 14(6), 850. https://doi.org/10.3390/plants14060850