

Heat Priming and Heat Stress Enhance Transgenerational Heat Tolerance in the Early Growth Stages of Oryza sativa L. Progeny



Abstract
1. Introduction
2. Results
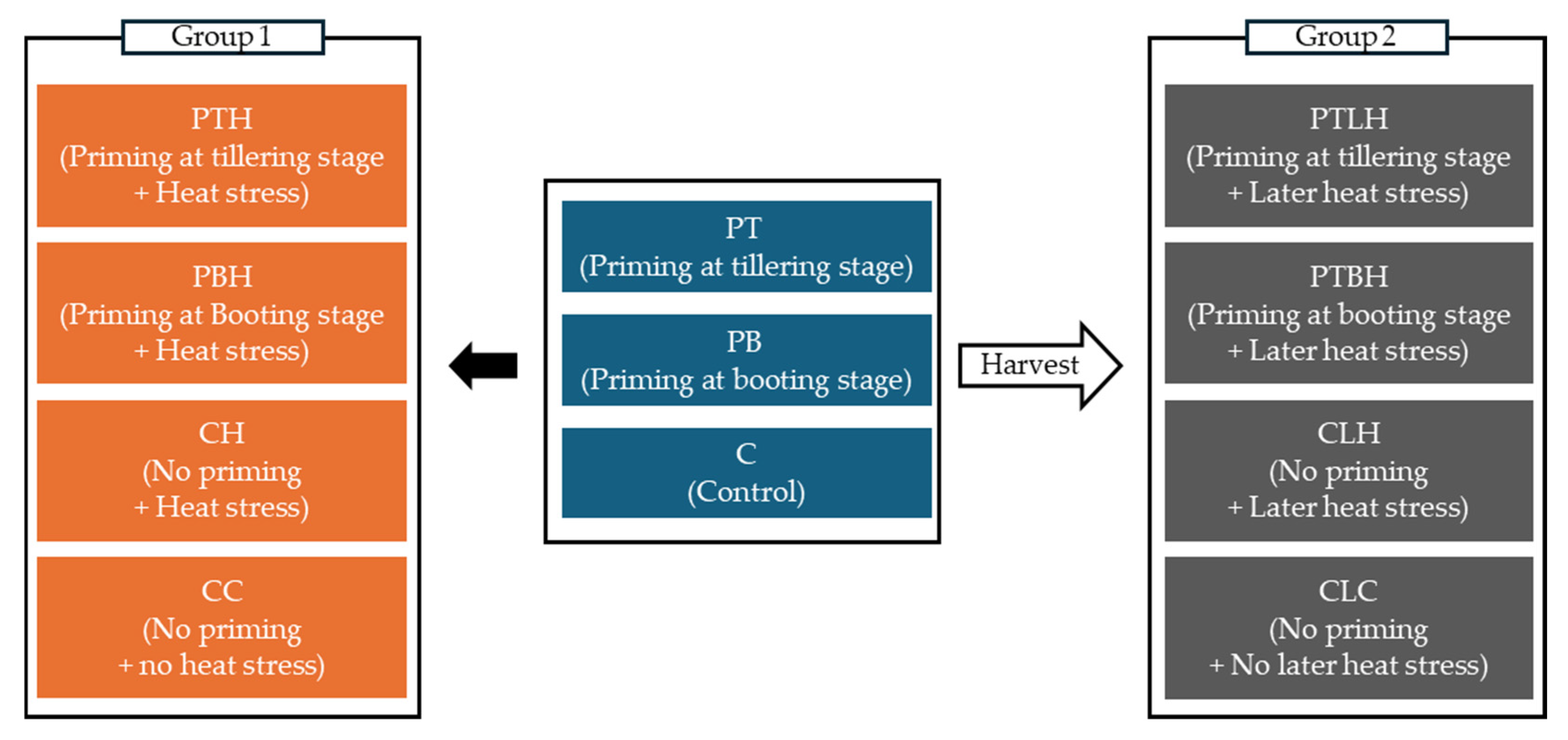
2.1. Experimental Design Overview
2.2. Changes in Germination Parameters
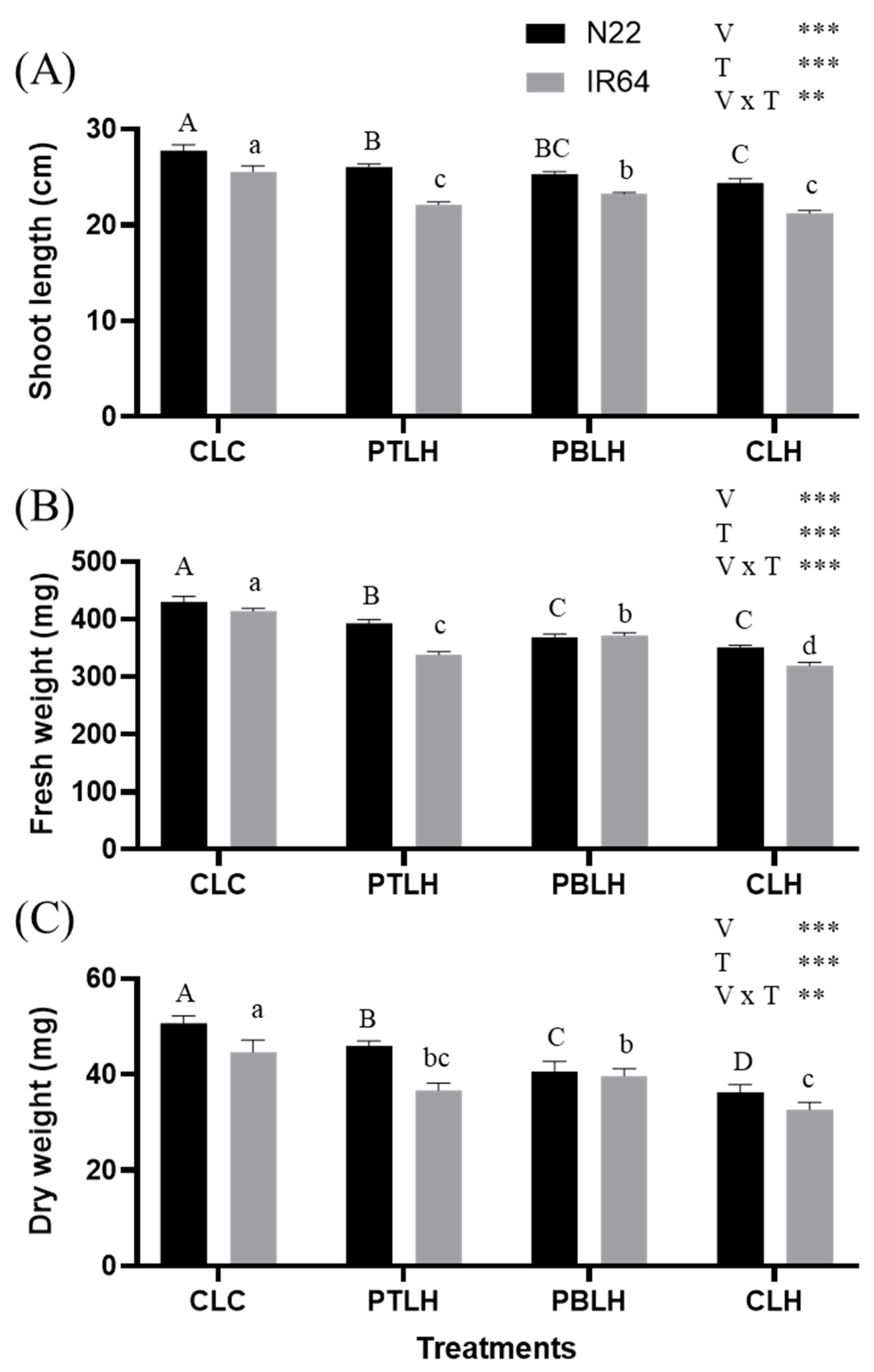
2.3. Biomass Parameters
2.4. H2O2 and MDA Content
3. Discussion
4. Materials and Methods
4.1. Seed Preparation
4.2. Germination Parameters
4.3. Later Heat Stress Application
- (1)
- PTLH: progeny from tillering-stage heat priming for five days, exposed to later heat stress for three days.
- (2)
- PBLH: progeny from booting-stage heat priming for five days, exposed to later heat stress for three days.
- (3)
- CLH: progeny from non-primed parents, exposed to later heat stress for three days.
- (4)
- CLC: progeny from non-primed parents, not exposed to heat stress.
4.4. Shoot Length and Shoot Biomass
4.5. H2O2 and MDA Analyses
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Abbreviations
| PT | Heat priming at the tillering stage |
| PB | Heat priming at the booting stage |
| CC | No heat priming with no heat stress |
| PTH | Heat priming at the tillering stage with heat stress at the flowering stage for seven days |
| PBH | Heat priming at the booting stage with heat stress at the flowering stage for seven days |
| CH | No heat priming with heat stress at the flowering stage for seven days |
| CLC | Progeny from non-primed parents, not exposed to heat stress |
| CLH | Progeny from non-primed parents, exposed to later heat stress for three days |
| PTLH | Progeny from tillering-stage heat priming for five days, exposed to later heat stress for three days |
| PBLH | Progeny from booting-stage heat priming for five days, exposed to later heat stress for three days |
References
- Sen, S.; Chakraborty, R.; Kalita, P. Rice-not just a staple food: A comprehensive review on its phytochemicals and therapeutic potential. Trends Food Sci. Technol. 2020, 97, 265–285. [Google Scholar] [CrossRef]
- Masson-Delmotte, V.; Zhai, P.; Pirani, A.; Connors, S.L.; Péan, C.; Berger, S.; Zhou, B. Climate Change 2021: The Physical Science Basis. In Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; IPCC: Geneva, Switzerland, 2021; Volume 2, p. 2391. [Google Scholar]
- Wu, C.; Cui, K.; Fahad, S. Heat stress decreases rice grain weight: Evidence and physiological mechanisms of heat effects prior to flowering. Int. J. Mol. Sci. 2022, 23, 10922. [Google Scholar] [CrossRef] [PubMed]
- Begcy, K.; Sandhu, J.; Walia, H. Transient heat stress during early seed development primes germination and seedling establishment in rice. Front. Plant Sci. 2018, 9, 1768. [Google Scholar] [CrossRef] [PubMed]
- Kilasi, N.L.; Singh, J.; Vallejos, C.E.; Ye, C.; Jagadish, S.K.; Kusolwa, P.; Rathinasabapathi, B. Heat stress tolerance in rice (Oryza sativa L.): Identification of quantitative trait loci and candidate genes for seedling growth under heat stress. Front. Plant Sci. 2018, 9, 1578. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.; Liu, B.; Piao, S.; Wang, X.; Lobell, D.B.; Huang, Y.; Huang, M.T.; Yao, Y.T.; Bassu, S.; Ciais, P.; et al. Temperature increase reduces global yields of major crops in four independent estimates. Proc. Natl. Acad. Sci. USA 2017, 114, 9326–9331. [Google Scholar] [CrossRef]
- Wolny, E.; Betekhtin, A.; Rojek, M.; Braszewska-Zalewska, A.; Lusinska, J.; Hasterok, R. Germination and the early stages of seedling development in Brachypodium distachyon. Int. J. Mol. Sci. 2018, 19, 2916. [Google Scholar] [CrossRef]
- Tarnawa, Á.; Kende, Z.; Sghaier, A.H.; Kovács, G.P.; Gyuricza, C.; Khaeim, H. Effect of abiotic stresses from drought, temperature, and density on germination and seedling growth of barley (Hordeum vulgare L.). Plants 2023, 12, 1792. [Google Scholar] [CrossRef]
- Tian, B.; Talukder, S.K.; Fu, J.; Fritz, A.K.; Trick, H.N. Expression of a rice soluble starch synthase gene in transgenic wheat improves the grain yield under heat stress conditions. In Vitro Cell. Dev. Biol. Plant 2018, 54, 216–227. [Google Scholar] [CrossRef]
- Xu, Y.; Chu, C.; Yao, S. The impact of high-temperature stress on rice: Challenges and solutions. Crop J. 2021, 9, 963–976. [Google Scholar] [CrossRef]
- Devireddy, A.R.; Tschaplinski, T.J.; Tuskan, G.A.; Muchero, W.; Chen, J.G. Role of reactive oxygen species and hormones in plant responses to temperature changes. Int. J. Mol. Sci. 2021, 22, 8843. [Google Scholar] [CrossRef]
- Wang, X.; Cai, J.; Liu, F.; Dai, T.; Cao, W.; Wollenweber, B.; Jiang, D. Multiple heat priming enhances thermo-tolerance to a later high temperature stress via improving subcellular antioxidant activities in wheat seedlings. Plant Physiol. Biochem. 2014, 74, 185–192. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.; Miao, Y.; Kong, J.; Lindsey, K.; Zhang, X.; Min, L. ROS signaling and its involvement in abiotic stress with emphasis on heat stress-driven anther sterility in plants. Crop Environ. 2024, 3, 65–74. [Google Scholar] [CrossRef]
- Kinoshita, T.; Seki, M. Epigenetic memory for stress response and adaptation in plants. Plant Cell Physiol. 2014, 55, 1859–1863. [Google Scholar] [CrossRef]
- Lämke, J.; Bäurle, I. Epigenetic and chromatin-based mechanisms in environmental stress adaptation and stress memory in plants. Genome Biol. 2017, 18, 124. [Google Scholar] [CrossRef]
- Fan, Y.; Ma, C.; Huang, Z.; Abid, M.; Jiang, S.; Dai, T.; Zhang, W.; Ma, S.; Jiang, D.; Han, X. Heat priming during early reproductive stages enhances thermo-tolerance to post-anthesis heat stress via improving photosynthesis and plant productivity in winter wheat (Triticum aestivum L.). Front. Plant Sci. 2018, 9, 805. [Google Scholar] [CrossRef]
- Hilker, M.; Schmülling, T. Stress priming, memory, and signalling in plants. Plant Cell Environ. 2019, 42, 753–761. [Google Scholar] [CrossRef]
- Zhang, X.; Zhou, Q.; Wang, X.; Cai, J.; Dai, T.; Cao, W.; Jiang, D. Physiological and transcriptional analyses of induced post-anthesis thermo-tolerance by heat-shock pretreatment on germinating seeds of winter wheat. Environ. Exp. Bot. 2016, 131, 181–189. [Google Scholar] [CrossRef]
- Balla, K.; Karsai, I.; Bencze, S.; Veisz, O. Germination ability and seedling vigour in the progeny of heat-stressed wheat plants. Acta Agron. Hung. 2012, 60, 299–308. [Google Scholar] [CrossRef]
- Suriyasak, C.; Oyama, Y.; Ishida, T.; Mashiguchi, K.; Yamaguchi, S.; Hamaoka, N.; Iwaya-Inoue, M.; Ishibashi, Y. Mechanism of delayed seed germination caused by high temperature during grain filling in rice (Oryza sativa L.). Sci. Rep. 2020, 10, 17378. [Google Scholar] [CrossRef]
- Wang, Y.; Sun, X.; Peng, J.; Li, F.; Ali, F.; Wang, Z. Regulation of seed germination: ROS, epigenetic, and hormonal aspects. J. Adv. Res. 2024, 10, 17378. [Google Scholar] [CrossRef]
- Suriyasak, C.; Kawaguchi, R.; Matsumoto, R.; Sawada, Y.; Nong, H.T.; Hamaoka, N.; Ishibashi, Y. Adaptive memory induced by heat stress during grain filling enhances subsequent thermotolerance in rice (Oryza sativa L.). Sci. Rep. 2025, 15, 14135. [Google Scholar] [CrossRef] [PubMed]
- Shah, K.; Chaturvedi, V.; Gupta, S. Climate Change and Abiotic Stress-Induced Oxidative Burst in Rice. In Advances in Rice Research for Abiotic Stress Tolerance; Woodhead Publishing: Sawston, UK, 2019; pp. 505–535. [Google Scholar]
- Liu, J.; Zhang, C.; Wei, C.; Liu, X.; Wang, M.; Yu, F.; Xie, Q.; Tu, J. The RING finger ubiquitin E3 ligase OsHTAS enhances heat tolerance by promoting H2O2-induced stomatal closure in rice. Plant Physiol. 2016, 170, 429–443. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Sun, X.; Xu, F.; Zhang, Y.; Zhang, Q.; Miao, R.; Zhang, Z.J.; Liang, L.J.; Xu, W. Suppression of OsMDHAR4 enhances heat tolerance by mediating H2O2-induced stomatal closure in rice plants. Rice 2018, 11, 38. [Google Scholar] [CrossRef]
- Sailaja, B.; Subrahmanyam, D.; Neelamraju, S.; Vishnukiran, T.; Rao, Y.V.; Vijayalakshmi, P.; Voleti, S.R.; Bhadana, V.P.; Mangrauthia, S.K. Integrated physiological, biochemical, and molecular analysis identifies important traits and mechanisms associated with differential response of rice genotypes to elevated temperature. Front. Plant Sci. 2015, 6, 1044. [Google Scholar] [CrossRef]
- Bahuguna, R.N.; Jha, J.; Pal, M.; Shah, D.; Lawas, L.M.; Khetarpal, S.; Jagadish, K.S. Physiological and biochemical characterization of NERICA-L-44: A novel source of heat tolerance at the vegetative and reproductive stages in rice. Physiol. Plant. 2015, 154, 543–559. [Google Scholar] [CrossRef]
- Niu, Y.; Xiang, Y. An overview of biomembrane functions in plant responses to high-temperature stress. Front. Plant Sci. 2018, 9, 915. [Google Scholar] [CrossRef]
- Bäurle, I. Plant heat adaptation: Priming in response to heat stress. F1000Research 2016, 5, 664. [Google Scholar] [CrossRef]
- Szymańska, R.; Ślesak, I.; Orzechowska, A.; Kruk, J. Physiological and biochemical responses to high light and temperature stress in plants. Environ. Exp. Bot. 2017, 139, 165–177. [Google Scholar] [CrossRef]
- Karwa, S.; Bahuguna, R.N.; Chaturvedi, A.K.; Maurya, S.; Arya, S.S.; Chinnusamy, V.; Pal, M. Phenotyping and characterization of heat stress tolerance at reproductive stage in rice (Oryza sativa L.). Acta Physiol. Plant. 2020, 42, 29. [Google Scholar] [CrossRef]
- Choi, Y.; Gehring, M.; Johnson, L.; Hannon, M.; Harada, J.J.; Goldberg, R.B.; Jacobsen, S.E.; Fischer, R.L. DEMETER, a DNA glycosylase domain protein, is required for endosperm gene imprinting and seed viability in Arabidopsis. Cell 2002, 110, 33–42. [Google Scholar] [CrossRef]
- Wibowo, A.; Becker, C.; Marconi, G.; Durr, J.; Price, J.; Hagmann, J.; Papareddy, R.; Putra, H.; Kageyama, J.; Becker, J.; et al. Hyperosmotic stress memory in Arabidopsis is mediated by distinct epigenetically labile sites in the genome and is restricted in the male germline by DNA glycosylase activity. Elife 2016, 5, e13546. [Google Scholar] [CrossRef] [PubMed]
- Nakao, Y.; Sone, C.; Sakagami, J.I. Genetic diversity of hydro priming effects on rice seed emergence and subsequent growth under different moisture conditions. Genes 2020, 11, 994. [Google Scholar] [CrossRef] [PubMed]
- Kulkarni, M.G.; Street, R.A.; Van Staden, J. Germination and seedling growth requirements for propagation of Dioscorea dregeana (Kunth) Dur. and Schinz—A tuberous medicinal plant. S. Afr. J. Bot. 2007, 73, 131–137. [Google Scholar] [CrossRef]
- Shahidullah, S.M.; Hanafi, M.M.; Ashrafuzzaman, M.; Ismail, M.R.; Salam, M.A.; Khair, A. Biomass accumulation and energy conversion efficiency in aromatic rice genotypes. Comptes Rendus. Biol. 2010, 333, 61–67. [Google Scholar] [CrossRef]
- Libron, J.A.M.A.; Putri, H.H.; Bore, E.K.; Chepkoech, R.; Akagi, I.; Odama, E.; Goto, K.; Tamaru, S.; Yabuta, S.; Sakagami, J.I. Halopriming in the submergence-tolerant rice variety improved the resilience to salinity and combined salinity-submergence at the seedling stage. Plant Physiol. Biochem. 2024, 208, 108494. [Google Scholar] [CrossRef]
- Nakao, Y.; Tamaru, S.; Yabuta, S.; Sakagami, J.I. Vitality of primed rice seeds sown under prolonged dry soil conditions in an upland field. Crop Sci. 2022, 62, 1277–1287. [Google Scholar] [CrossRef]
- Bore, E.K.; Ishikawa, E.; Libron, J.A.M.A.; Goto, K.; Odama, E.; Nakao, Y.; Yabuta, S.; Sakagami, J.I. Primed Seeds of NERICA 4 Stored for Long Periods under High Temperature and Humidity Conditions Maintain Germination Rates. Appl. Sci. 2023, 13, 2869. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
| Variety | Treatment | Germination Percentage (%) | T50 | MGT | |||
|---|---|---|---|---|---|---|---|
| 36 hai | 48 hai | 60 hai | 72 hai | ||||
| N22 | CC | 6.67 a | 73.33 a | 84.44 a | 91.11 a | 43.7 b | 50.3 b |
| PTH | 8.89 a | 46.67 ab | 75.56 ab | 88.89 a | 48.7 ab | 54.1 ab | |
| PBH | 11.11 a | 28.89 ab | 60.00 b | 84.44 a | 53.0 ab | 57.7 ab | |
| CH | 0.00 a | 26.67 b | 42.22 c | 77.78 a | 54.6 a | 61.3 a | |
| IR64 | CC | 11.11 a | 55.56 a | 91.11 a | 91.11 a | 46.4 b | 51.2 b |
| PTH | 4.44 a | 42.22 ab | 66.67 b | 82.22 a | 48.6a b | 55.3 a | |
| PBH | 8.89 a | 44.44 ab | 80.00 ab | 82.22 a | 46.8 b | 52.5 b | |
| CH | 0.00 a | 31.11 b | 62.22 b | 71.11 a | 50.8 a | 56.3 a | |
| Variety | ns | ns | * | ns | ns | ns | |
| Treatment | * | ** | *** | * | ** | ** | |
| Variety × Treatment | ns | ns | * | ns | ns | ns | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ju, Y.; Choi, J.; Yun, S.; Sakagami, J.I. Heat Priming and Heat Stress Enhance Transgenerational Heat Tolerance in the Early Growth Stages of Oryza sativa L. Progeny. Plants 2025, 14, 1593. https://doi.org/10.3390/plants14111593
Ju Y, Choi J, Yun S, Sakagami JI. Heat Priming and Heat Stress Enhance Transgenerational Heat Tolerance in the Early Growth Stages of Oryza sativa L. Progeny. Plants. 2025; 14(11):1593. https://doi.org/10.3390/plants14111593
Chicago/Turabian StyleJu, Younghwan, Juyoung Choi, Sungho Yun, and Jun Ichi Sakagami. 2025. "Heat Priming and Heat Stress Enhance Transgenerational Heat Tolerance in the Early Growth Stages of Oryza sativa L. Progeny" Plants 14, no. 11: 1593. https://doi.org/10.3390/plants14111593
APA StyleJu, Y., Choi, J., Yun, S., & Sakagami, J. I. (2025). Heat Priming and Heat Stress Enhance Transgenerational Heat Tolerance in the Early Growth Stages of Oryza sativa L. Progeny. Plants, 14(11), 1593. https://doi.org/10.3390/plants14111593