
Antifungal Activity of Genistein Against Phytopathogenic Fungi Valsa mali Through ROS-Mediated Lipid Peroxidation


Abstract
1. Introduction
2. Results
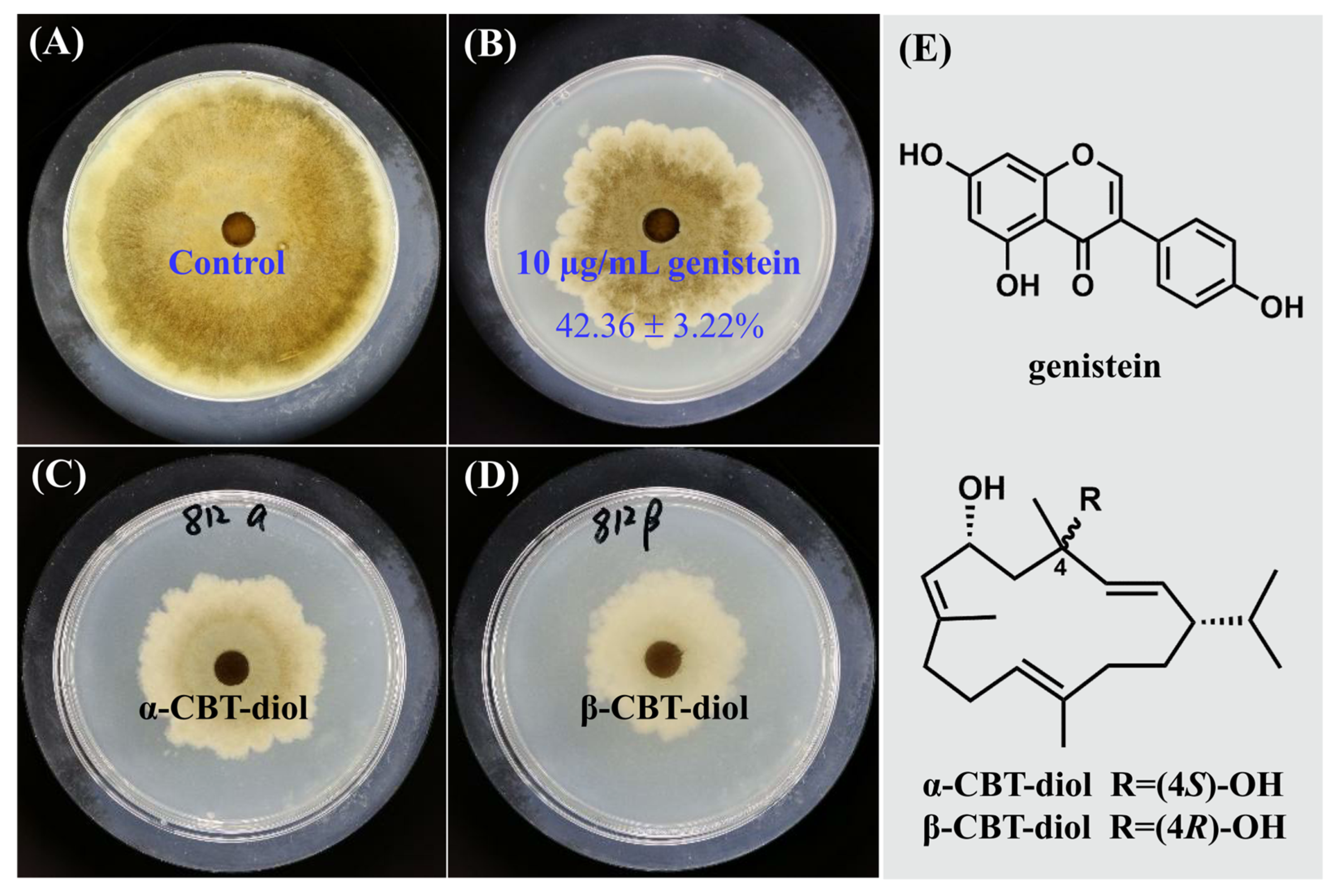
2.1. Inhibitory Effect of Genistein on the Growth of V. mali
2.2. Effects of Genistein on the Morphology and Ultrastructure of V. mali Mycelia
2.3. Effects of Genistein on the Reactive Oxygen Species (ROS) Level of V. mali
2.4. Determination of Antioxidant Enzyme Activity
2.5. Quality Assessment of Sequencing Data
2.6. Differentially Expressed Genes (DEGs) Expression Analysis
2.7. Gene Ontology (GO) Term and Kyoto Encyclopedia of Genes and Genomes (KEGG) Functional Enrichment Analyses of DEGs
2.8. Effects of Genistein on the Cell Membrane System of V. mali
3. Discussion
4. Materials and Methods
4.1. Media, Pathogens and Reagents
4.2. Growth Inhibition Assay of Genistein Against V. mali
4.3. Preparation and Examination of Samples for SEM and TEM
4.4. Determination of the Intracellular ROS Content
4.5. Determination of the Activities of Antioxidant Enzymes
4.6. RNA Extraction and Sequencing
4.7. Analysis of Sequencing Data
4.8. qRT-PCR Analysis
4.9. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Li, P.F.; Tedersoo, L.; Crowther, T.W.; Wang, B.Z.; Shi, Y.; Kuang, L.; Li, T.; Wu, M.; Liu, M.; Luan, L.; et al. Global diversity and biogeography of potential phytopathogenic fungi in a changing world. Nat. Commun. 2023, 14, 6482. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.S.; Meng, Y.G.; Liu, R.H.; Xiao, Y.Z.; Wang, Y.H.; Huang, L.L. Inhibitory effects of Bacillus vallismortis T27 against apple Valsa canker caused by Valsa mali. Pestic. Biochem. Physiol. 2023, 195, 105564. [Google Scholar] [CrossRef]
- Niu, X.D.; Wang, Z.Y.; Wang, C.Y.; Wang, H.S. Dibenzylideneacetone overcomes Botrytis cinerea infection in cherry tomatoes by inhibiting chitinase activity. J. Agric. Food Chem. 2023, 71, 19422–19433. [Google Scholar] [CrossRef]
- Fisher, M.C.; Henk, D.A.; Briggs, C.J.; Brownstein, J.S.; Madoff, L.C.; McCraw, S.L.; Gurr, S.J. Emerging fungal threats to animal, plant and ecosystem health. Nature 2012, 484, 186–194. [Google Scholar] [CrossRef] [PubMed]
- Li, J.Z.; Wang, Y.; Li, Y.X.; Zhuo, X.Y.; Wang, Z.X.; Zhao, X.L.; Wang, M.C.; Hou, Y.L.; Li, X.Y. Research progress and prospect on Valsa mali Miyabe & Yamada. Mol. Plant Breed. 2019, 17, 7222–7227. [Google Scholar]
- Zhao, H.F.; Feng, H.L.; Sun, G.P.; Zheng, Y.Q.; Yan, J.L. Effects of different agents on the prevention and control effect of apple tree rot disease. North. Fruits 2024, 18–20. [Google Scholar] [CrossRef]
- Wang, M.; Duan, H.M.; Li, Z.S.; Meng, X.T.; Lu, W.J. Inhibitory activity of seven fungicides combined with fermentation. J. Xichang Univ. (Nat. Sci. Ed.) 2023, 37, 15–20. [Google Scholar]
- Chen, Y.Z.; Wang, S.R.; Li, T.; Zhang, G.C.; Yang, J. Antifungal activity of 6-methylcoumarin against Valsa mali and its possible mechanism of action. J. Fungi. 2023, 9, 5. [Google Scholar] [CrossRef]
- Duan, C.B.; Du, Y.; Hou, X.; Yan, N.; Dong, W.; Mao, X.; Zhang, Z. Chemical basis of the fungicidal activity of tobacco extracts against Valsa mali. Molecules 2016, 21, 1743. [Google Scholar] [CrossRef] [PubMed]
- Jia, Y.N.; Tang, J.R.; Dang, Y.F.; Yang, Y.Z.; He, S.; Jiang, Y.; Wang, Y.; Feng, J.T.; Ma, Z.Q.; Liu, X.L.; et al. Lansiumamide B induced the degradation of polysaccharides and oligosaccharides showing antifungal activity against Rhizoctonia solani. Ind. Crops Prod. 2024, 214, 118529. [Google Scholar] [CrossRef]
- Zhang, J.X.; Wang, X.L.; Vikash, V.; Ye, Q.; Wu, D.D.; Liu, Y.L.; Dong, W.G. ROS and ROS-mediated cellular signaling. Oxid. Med. Cell. Longev. 2016, 2016, 4350965. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.; Wang, J.; Zhang, P.; Xie, S.N.; Yuan, X.L.; Hou, X.D.; Yan, N.; Fang, Y.D.; Du, Y.M. In vitro and in vivo antifungal activity and preliminary mechanism of cembratrien-diols against Botrytis cinerea. Ind. Crops Prod. 2020, 154, 112745. [Google Scholar] [CrossRef]
- Song, C.G.; Shi, W.L.; Yang, J.; Ding, G.; Chen, W.; Jia, C.G.; Zhang, M.Z.; Qin, J.C. Mechanism study on Monarda didyma essential oil inhibiting Aspergillus flavus infection and aflatoxins accumulation in peanuts. Ind. Crops Prod. 2024, 212, 118311. [Google Scholar] [CrossRef]
- Yang, Q. Antifungal Activity of CBT-diols Against Botrytis cinerea. Master’s Thesis, Chinese Academy of Agricultural Sciences, Beijing, China, June 2020. [Google Scholar]
- OuYang, Q.L.; Tao, N.G.; Jing, G.X. Transcriptional profiling analysis of Penicillium digitatum, the causal agent of citrus green mold, unravels an inhibited ergosterol biosynthesis pathway in response to citral. BMC Genom. 2016, 17, 599. [Google Scholar] [CrossRef]
- Chen, C.; Li, B.H.; Dong, X.L.; Wang, C.X.; Lian, S.; Liang, W.X. Effects of temperature, humidity, and wound age on Valsa mali infection of apple shoot pruning. Plant Dis. 2016, 100, 2394–2401. [Google Scholar] [CrossRef]
- Zhai, H.Z.; Hu, T.L.; Chen, Q.; Cao, K.Q. Control effect of 10 fungicides against apple valsa canker. Plant Prot. 2012, 38, 151–154+171. [Google Scholar]
- Wang, X.H.; Pan, T.T.; Lian, S.; Wang, C.X.; Li, B.H. Effects of environmental factors on the growth and extension of Valsa mali in the xylem of apple branches. Agric. Sci. China 2018, 51, 3291–3301. [Google Scholar]
- Liu, Z.Y.; Wang, S.; Gao, Y.Q.; Huang, L.L.; Feng, H. The cross-resistance of two geographically different Valsa mali strains to SBIs inhibitors and their biotic fitness. J. Northwest For. Univ. 2020, 35, 119–124. [Google Scholar]
- Ma, H. Study on the synergistic technology of Psoralea corylifolia seed extract against apple valsa canker based on Box-Behnken response methodology. Chin. Fruit 2023, 96–99. [Google Scholar] [CrossRef]
- Yang, C.; Xie, S.N.; Ni, L.; Du, Y.M.; Liu, S.; Li, M.Y.; Xu, K. Chemical constituents from Nicotiana tabacum L. and their antifungal activity. Nat. Prod. Commun. 2021, 16, 1–5. [Google Scholar] [CrossRef]
- Chen, X.; Zheng, G.P.; Yu, Z.G. Simultaneous assay of isoflavones in Fructus Sophorae by QAMS. Chin. J. Pharm. Anal. 2013, 33, 1904–1909. [Google Scholar]
- Chen, X.X.; Luan, F.; Liu, H.T. HPLC determination of daidzein, genistein and formononetin in Trifolium pratense. J. Lanzhou Univ. 2014, 40, 35–37. [Google Scholar]
- Jin, X.; Qin, X.H.; Xu, L.; Su, B.; Mu, X.T.; Liu, H.H.; Li, Y.L.; Zhu, Z.; Zhang, D.F. Simultaneous determination of genistein and kaempferol from Kummerowia striata. Pract. Pharm. Clin. Rem. 2015, 18, 1476–1478. [Google Scholar]
- Xing, J.H.; Sun, X.L.; Zhou, J. HPLC determination of genistein, formononetin and biochanin A in Cicer arietinum L. Chin. J. Pharm. Anal. 2009, 29, 73–75. [Google Scholar]
- Xu, J.B. Simultaneous determination of quercetin-7-O-β-D-rhamnoside and genistein in extract of Persimmon leaves by HPLC. J. Chang. Med. Coll. 2016, 30, 264–266. [Google Scholar]
- Yang, Q.M.; Zhang, C.F.; Yang, Z.L. Simultaneous determination of genistin and genistein in Moghania Philippinens (Merr. et Rolfe) Li. from different places by HPLC. Chin. J. Mod. Appl. Pharm. 2012, 29, 744–748. [Google Scholar]
- Yao, L.W.; Chen, X.H.; Shi, Y.; Wei, F.; Ma, S.C. Determination of quercetin, genistein and kaempferol in Cudrania tricuspidata. Chin. J. Pharm. Anal. 2014, 34, 415–417. [Google Scholar]
- Yu, Y.Y.; Zhang, L.H.; Cao, S.W. Determination of daidzein, genistein, formononetin, and biochanin A in Millettla nitida var. hirsutissima extract by RP-HPLC. Chin. Tradit. Herb. Drugs 2006, 37, 1655–1656. [Google Scholar]
- Zheng, L.; Long, F. Determination of genistein in root of Pueraria omeiensis by RP HPLC. China Pract. Med. 2006, 1, 32–33. [Google Scholar]
- Zhang, L. RP-HPLC determination of daidzein, genistein, daidzin and genistin in soybean extract. Chin. Tradit. Herb. Drugs 2001, 32, 118–120. [Google Scholar]
- Konstantinou, E.K.; Gioxari, A.; Dimitriou, M.; Panoutsopoulos, G.I.; Panagiotopoulos, A.A. Molecular pathways of genistein activity in breast cancer cells. Int. J. Mol. Sci. 2024, 25, 5556. [Google Scholar] [CrossRef] [PubMed]
- Wei, T.T.; Chandy, M.; Nishiga, M.; Zhang, A.; Kumar, K.K.; Thomas, D.; Manhas, A.; Rhee, S.; Justesen, J.M.; Chen, L.Y.; et al. Cannabinoid receptor 1 antagonist genistein attenuates marijuana-induced vascular inflammation. Cell 2022, 185, 1676–1693. [Google Scholar] [CrossRef]
- Viña, J.; Escudero, J.; Baquero, M.; Cebrián, M.; Carbonell-Asíns, J.A.; Muñoz, J.E.; Satorres, E.; Meléndez, J.C.; Ferrer-Rebolleda, J.; Cózar-Santiago, M.D.P.; et al. Genistein effect on cognition in prodromal Alzheimer’s disease patients. The GENIAL clinical trial. Alzheimers Res. Ther. 2022, 14, 164. [Google Scholar] [CrossRef] [PubMed]
- Jiang, T.; Dong, Y.H.; Zhu, W.Y.; Wu, T.; Chen, L.Y.; Cao, Y.T.; Yu, X.; Peng, Y.; Wang, L.; Xiao, Y.; et al. Underlying mechanisms and molecular targets of genistein in the management of type 2 diabetes mellitus and related complications. Crit. Rev. Food Sci. Nutr. 2023, 64, 11543–11555. [Google Scholar] [CrossRef]
- Hu, X.J. The Assessment of Potential Breast Health Hazard due to Dietary Genistein Exposure. Ph.D. Thesis, Nanchang University, Nanchang, China, June 2014. [Google Scholar]
- Shi, F.X. Effect of Genistein and Bisphenol-A on Female Reproduction. Ph.D. Thesis, Nanjing Agricultural University, Nanjing, China, June 2019. [Google Scholar]
- Rocha, M.J.; Cruzeiro, C.; Reis, M.; Pardal, M.; Rocha, E. Spatial and seasonal distribution of 17 endocrine disruptor compounds in an urban estuary (Mondego River, Portugal): Evaluation of the estrogenic load of the area. Environ. Monit. Assess. 2014, 186, 3337–3350. [Google Scholar] [CrossRef]
- Teilum, K.; Olsen, J.G.; Kragelund, B.B. Protein stability, flexibility and function. Biochim. Biophys. Acta 2011, 1814, 969–976. [Google Scholar] [CrossRef] [PubMed]
- Naqvi, A.A.T.; Mohammad, T.; Hasan, G.M.; Hassan, M.I. Advancements in docking and molecular dynamics simulations towards ligand-receptor interactions and structure-function relationships. Curr. Top. Med. Chem. 2018, 18, 1755–1768. [Google Scholar] [CrossRef]
- Porta, J.; Vahedi-Faridi, A.; Borgstahl, G.E. Structural analysis of peroxide-soaked MnSOD crystals reveals side-on binding of peroxide to active-site manganese. J. Mol. Biol. 2010, 399, 377–384. [Google Scholar] [CrossRef] [PubMed]
- Murphy, E.J.; Metcalfe, C.L.; Basran, J.; Moody, P.C.; Raven, E.L. Engineering the substrate specificity and reactivity of a heme protein: Creation of an ascorbate binding site in cytochrome c peroxidase. Biochemistry 2008, 47, 13933–13941. [Google Scholar] [CrossRef] [PubMed]
- Dutta, R.K.; Nenavathu, B.P.; Gangishetty, M.K.; Reddy, A.V. Studies on antibacterial activity of ZnO nanoparticles by ROS induced lipid peroxidation. Colloids Surf. B Biointerfaces 2012, 94, 143–150. [Google Scholar] [CrossRef] [PubMed]
- Yan, N.; Du, Y.M.; Liu, X.M.; Zhang, H.B.; Liu, Y.H.; Shi, J.; Xue, S.J.; Zhang, Z.F. Analyses of effects of α-cembratrien-diol on cell morphology and transcriptome of Valsa mali var. mali. Food Chem. 2017, 214, 110–118. [Google Scholar] [CrossRef]
- Ke, Y.; Ding, B.B.; Zhang, M.M.; Dong, T.J.; Fu, Y.; Lv, Q.Y.; Ding, W.P.; Wang, X.D. Study on inhibitory activity and mechanism of chitosan oligosaccharides on Aspergillus Flavus and Aspergillus Fumigatus. Carbohydr. Polym. 2022, 275, 118673. [Google Scholar] [CrossRef] [PubMed]
- Jian, Y.Q.; Chen, X.; Ahmed, T.; Shang, Q.H.; Zhang, S.; Ma, Z.H.; Yin, Y.N. Toxicity and action mechanisms of silver nanoparticles against the mycotoxin-producing fungus Fusarium graminearum. J. Adv. Res. 2021, 38, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Morris, G.M.; Huey, R.; Lindstrom, W.; Sanner, M.F.; Belew, R.K.; Goodsell, D.S.; Olson, A.J. AutoDock4 and AutoDockTools4: Automated docking with selective receptor flexibility. J. Comput. Chem. 2009, 30, 2785–2791. [Google Scholar] [CrossRef] [PubMed]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef]
- Eberhardt, J.; Santos-Martins, D.; Tillack, A.F.; Forli, S. AutoDock Vina 1.2.0: New docking methods, expanded force field, and python bindings. J. Chem. Inf. Model. 2021, 61, 3891–3898. [Google Scholar] [CrossRef]
- Mortazavi, A.; Williams, B.A.; McCue, K.; Schaeffer, L.; Wold, B. Mapping and quantifying mammalian transcriptomes by RNA-Seq. Nat. Methods 2008, 5, 621–628. [Google Scholar] [CrossRef]
- Bray, N.L.; Pimentel, H.; Melsted, P.; Pachter, L. Near-optimal probabilistic RNA-seq quantification. Nat. Biotechnol. 2016, 34, 525–527. [Google Scholar] [CrossRef] [PubMed]
- Garber, M.; Grabherr, M.G.; Guttman, M.; Trapnell, C. Computational methods for transcriptome annotation and quantification using RNA-seq. Nat. Methods 2011, 8, 469–477. [Google Scholar] [CrossRef]
- Liao, Y.; Smyth, G.K.; Shi, W. featureCounts: An efficient general purpose program for assigning sequence reads to genomic features. Bioinformatics 2014, 30, 923–930. [Google Scholar] [CrossRef]
- Anders, S.; Huber, W. Differential expression analysis for sequence count data. Genome Biol. 2010, 11, R106. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. edgeR: A Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef]
- Young, M.D.; Wakefield, M.J.; Smyth, G.K.; Oshlack, A. Gene ontology analysis for RNA-seq: Accounting for selection bias. Genome Biol. 2010, 11, R14. Available online: http://genomebiology.com/2010/11/2/R14 (accessed on 1 March 2023). [CrossRef] [PubMed]
- Yin, Z.Y.; Ke, X.W.; Huang, D.X.; Gao, X.N.; Voegele, R.T.; Kang, Z.S.; Huang, L.L. Validation of reference genes for gene expression analysis in Valsa mali var. mali using real-time quantitative PCR. World J. Microbiol. Biotechnol. 2013, 29, 1563–1571. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Samples | Control-1 | Control-2 | Control-3 | Treated-1 | Treated-2 | Treated-3 |
|---|---|---|---|---|---|---|
| Raw reads | 46,768,938 | 47,448,424 | 48,968,902 | 46,731,424 | 45,432,282 | 45,201,716 |
| Raw bases | 7.02 G | 7.12 G | 7.35 G | 7.01 G | 6.81 G | 6.78 G |
| Clean reads | 45,370,436 | 46,266,524 | 47,631,958 | 45,241,510 | 44,104,334 | 44,120,272 |
| Clean reads/raw reads (%) | 97.01% | 97.51% | 97.27% | 96.81% | 97.08% | 97.61% |
| Clean bases | 6.81 G | 6.94 G | 7.14 G | 6.79 G | 6.62 G | 6.62 G |
| Error rate | 0.02 | 0.02 | 0.02 | 0.02 | 0.02 | 0.02 |
| Q20 | 98.53 | 98.34 | 98.42 | 98.39 | 98.36 | 98.36 |
| Q30 | 95.79 | 95.25 | 95.47 | 95.44 | 95.33 | 95.32 |
| GC pct | 56.73 | 56.87 | 57.02 | 56.59 | 56.62 | 56.71 |
| Samples | Total Reads | Total Map | Unique Map | Multi Map |
|---|---|---|---|---|
| Control-1 | 45,370,436 | 43,541,348 (95.97%) | 43,327,458 (95.5%) | 213,890 (0.47%) |
| Control-2 | 46,266,524 | 44,370,961 (95.9%) | 44,121,004 (95.36%) | 249,957 (0.54%) |
| Control-3 | 47,631,958 | 45,658,834 (95.86%) | 45,424,635 (95.37%) | 234,199 (0.49%) |
| Treated-1 | 45,241,510 | 43,476,439 (96.1%) | 43,277,906 (95.66%) | 198,533 (0.44%) |
| Treated-2 | 44,104,334 | 42,428,393 (96.2%) | 42,239,746 (95.77%) | 188,647 (0.43%) |
| Treated-3 | 44,120,272 | 42,441,929 (96.2%) | 42,227,638 (95.71%) | 214,291 (0.49%) |
| Genes Name | Primers (5′ to 3′) |
|---|---|
| G6PDH [58] | F: TCAGAACAAGTTCGAGGGCGACAA |
| R: TGAGGGCAATAGAGGGCTTGTTCA | |
| VM1G_04618 | F: AGCTTTCCGGTCATTATCACA |
| R: GTCCGAAGTCAATGATCCACA | |
| VM1G_04926 | F: AGCTCCTCTCGGTCGTGCTCA |
| R: CACACCCACGGCCACGTCGTC | |
| VM1G_05255 | F: AAAGGCTCTTCTTGTCGTCCA |
| R: TTTGCTGTCCTGCTCCTCGTC | |
| VM1G_07069 | F: ACACAACTCGCCATATCCCA |
| R: TCCTCTTTGGATTCGACGATG | |
| VM1G_07417 | F: TGATTTCATGGACCTGCCGTT |
| R: TAGATCTGGCTGTACACGCTCT | |
| VM1G_07065 | F: ACCATCCTGACATTGAGCCTA |
| R: AACCTGCCCCTTTTGTACCAC | |
| VM1G_08675 | F: CTCACGATACCCGTATTTGGC |
| R: AATCAAGGGAACATGCGACT | |
| VM1G_09419 | F: CGTGTTGGTCTTCCCATTGACA |
| R: AGAACCTTGCCATTGATCCAC | |
| VM1G_09989 | F: CCGAGACACATCTACATGCAC |
| R: ATATCATGCGGCCAATTCCC | |
| VM1G_11144 | F: GATACGTCTTGATGCACCGAT |
| R: CTTTCCCAGTTGTCCGCTTG | |
| VM1G_05401 | F: AATGTATCACCGCAAAATCCG |
| R: GTACTCTTGACTCAGCGACCA | |
| VM1G_07635 | F: GCGGTCCTTCCCTACCTCC |
| R: CTACTCCCCTGGATAGCGTCT |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, F.; Yang, C.; Li, M.; Liu, S.; Xu, K.; Fu, X. Antifungal Activity of Genistein Against Phytopathogenic Fungi Valsa mali Through ROS-Mediated Lipid Peroxidation. Plants 2025, 14, 120. https://doi.org/10.3390/plants14010120
Li F, Yang C, Li M, Liu S, Xu K, Fu X. Antifungal Activity of Genistein Against Phytopathogenic Fungi Valsa mali Through ROS-Mediated Lipid Peroxidation. Plants. 2025; 14(1):120. https://doi.org/10.3390/plants14010120
Chicago/Turabian StyleLi, Fangjie, Chen Yang, Maoye Li, Su Liu, Kuo Xu, and Xianjun Fu. 2025. "Antifungal Activity of Genistein Against Phytopathogenic Fungi Valsa mali Through ROS-Mediated Lipid Peroxidation" Plants 14, no. 1: 120. https://doi.org/10.3390/plants14010120
APA StyleLi, F., Yang, C., Li, M., Liu, S., Xu, K., & Fu, X. (2025). Antifungal Activity of Genistein Against Phytopathogenic Fungi Valsa mali Through ROS-Mediated Lipid Peroxidation. Plants, 14(1), 120. https://doi.org/10.3390/plants14010120