
Effect of Na, K and Ca Salts on Growth, Physiological Performance, Ion Accumulation and Mineral Nutrition of Mesembryanthemum crystallinum



Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material, Establishment and Cultivation Conditions
2.2. Treatments and Physiological Analysis
2.3. Termination of the Experiments and Measurements
2.4. Data Analysis
3. Results
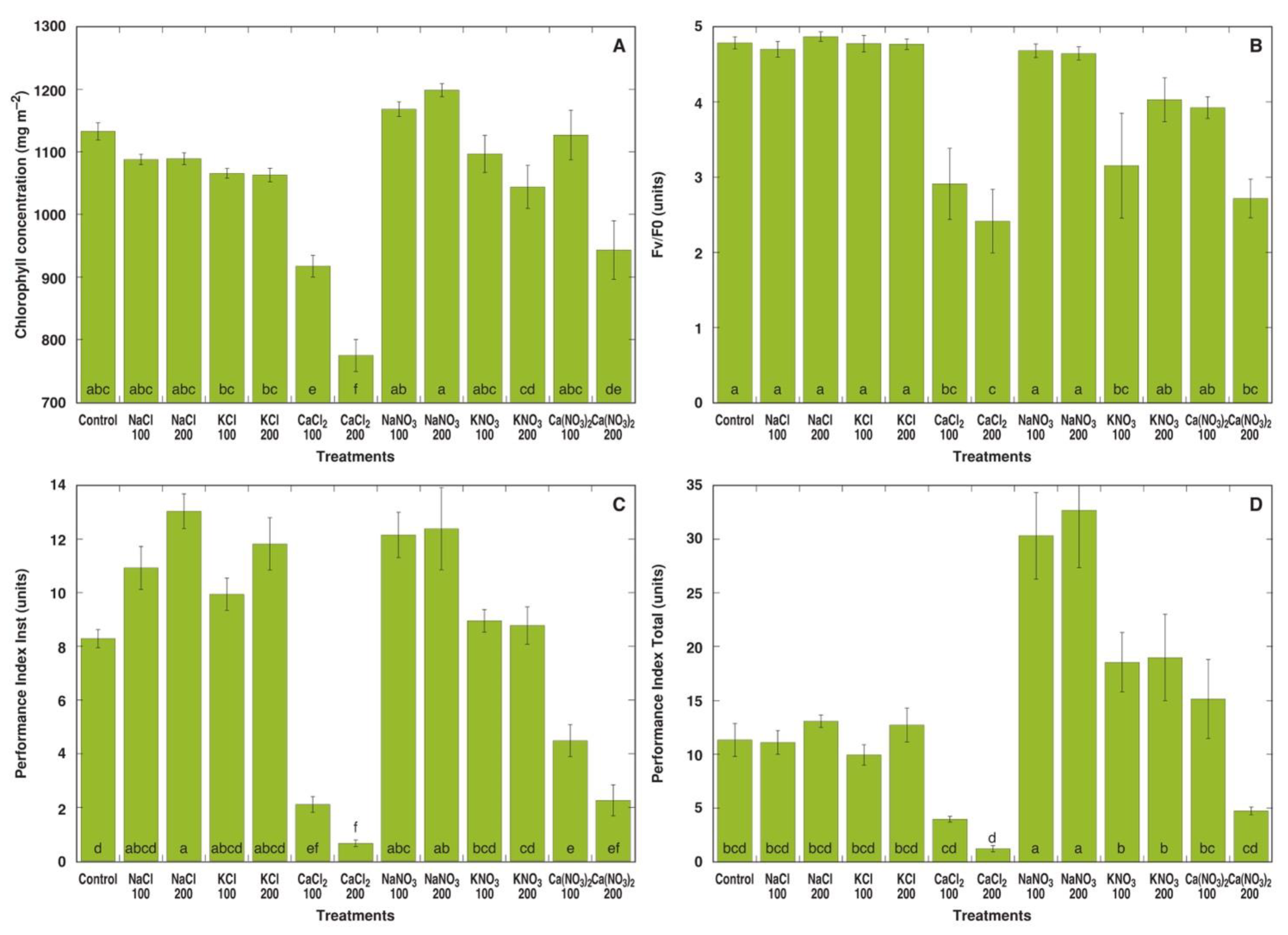
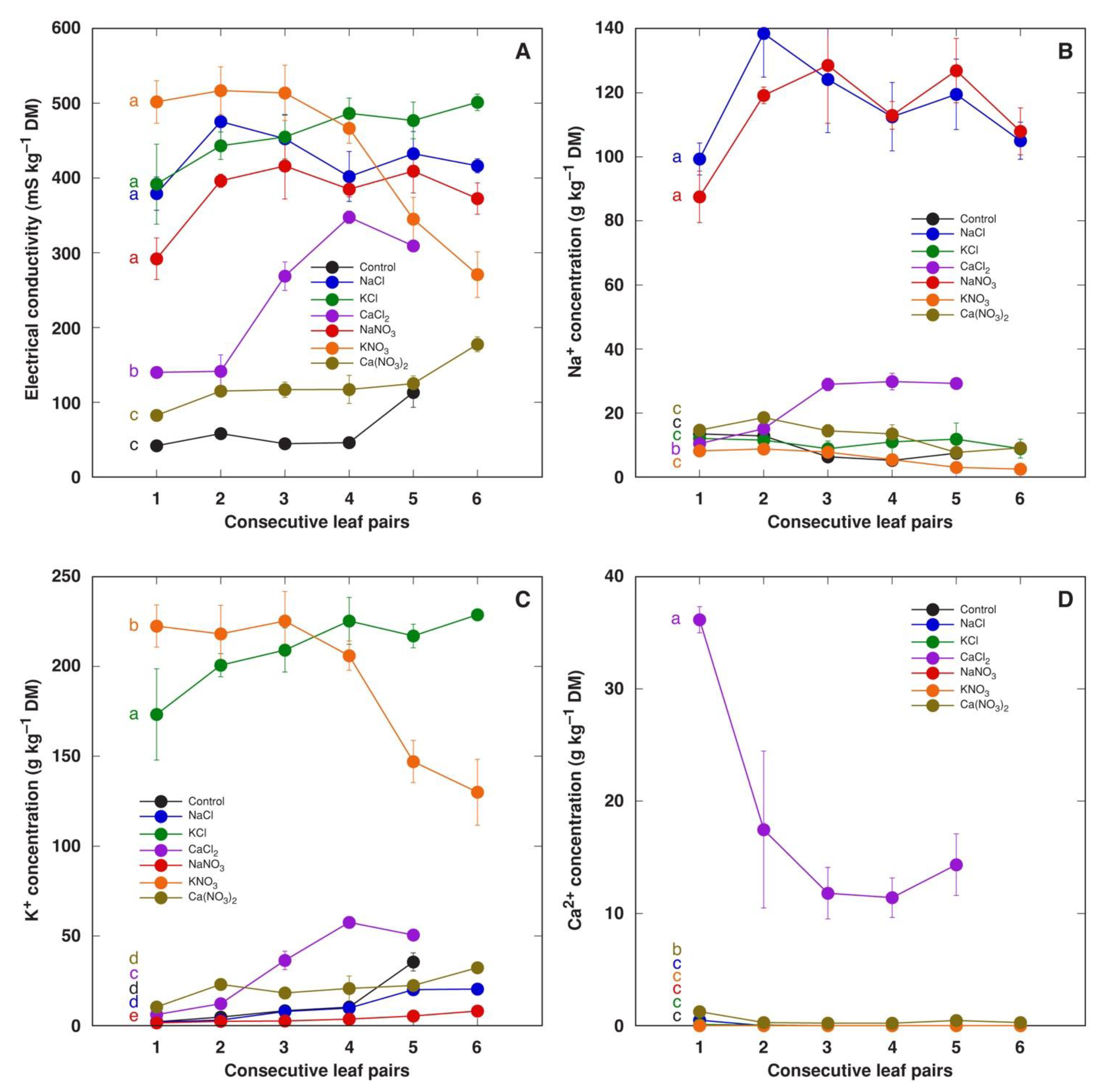
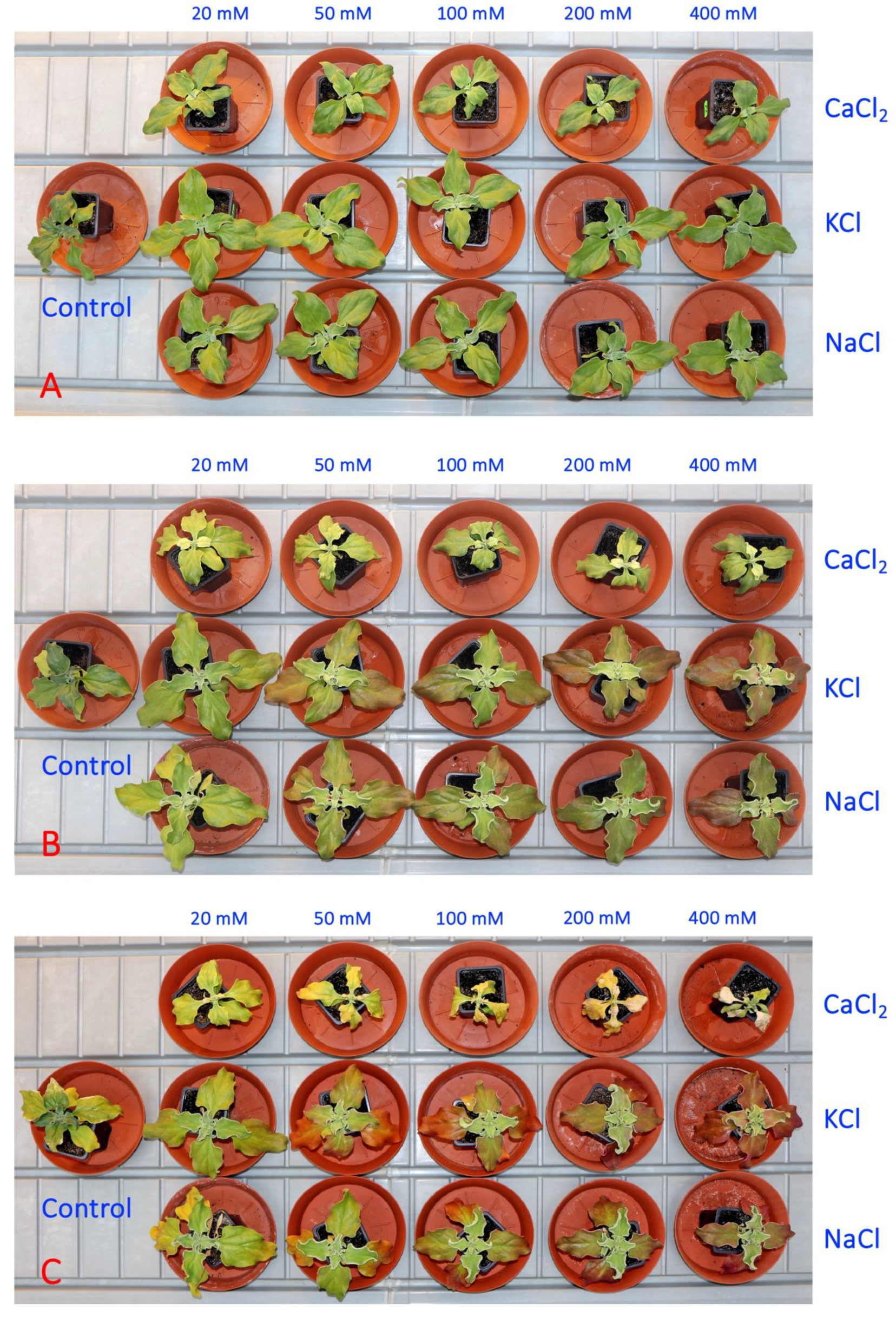
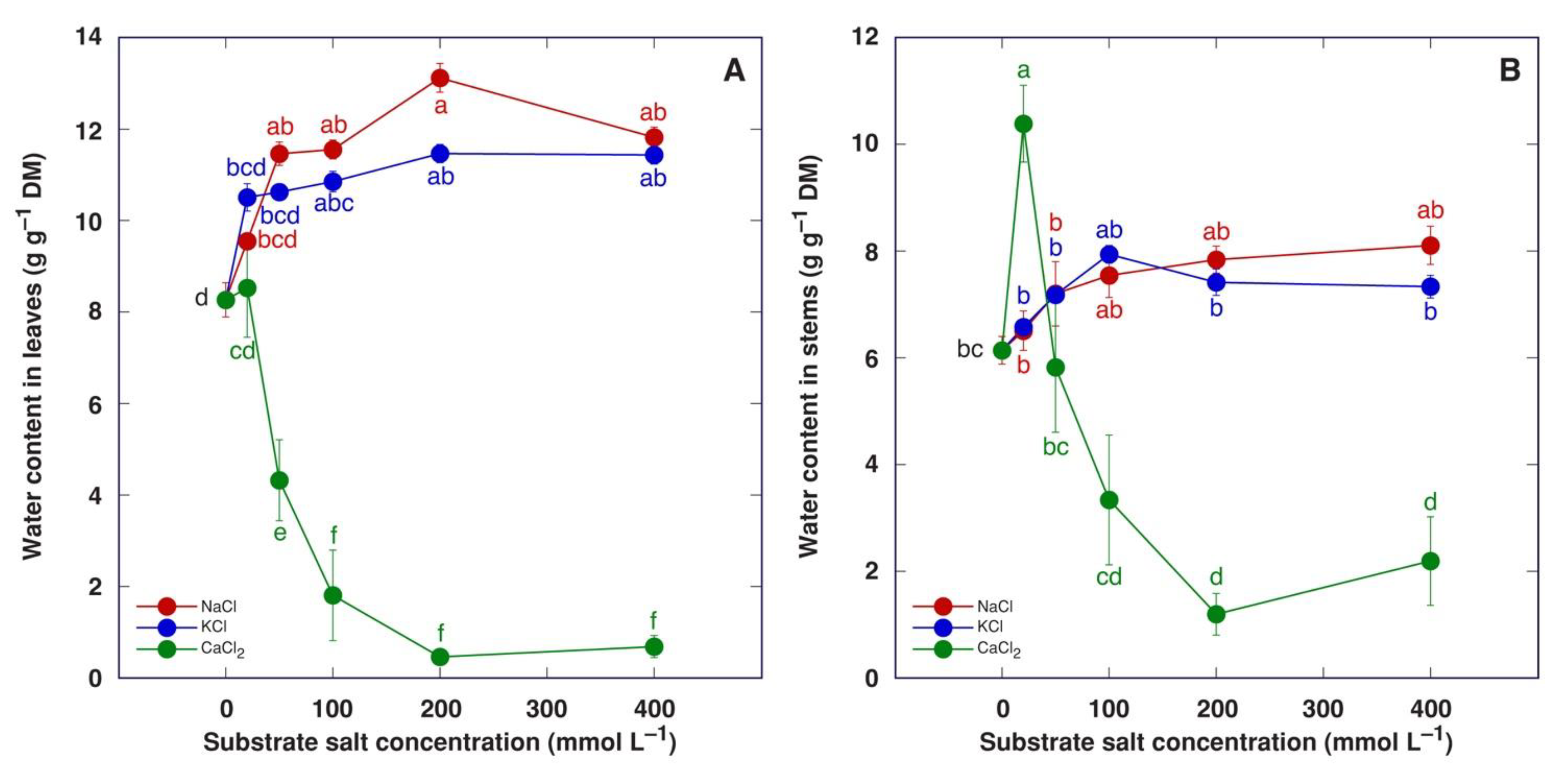
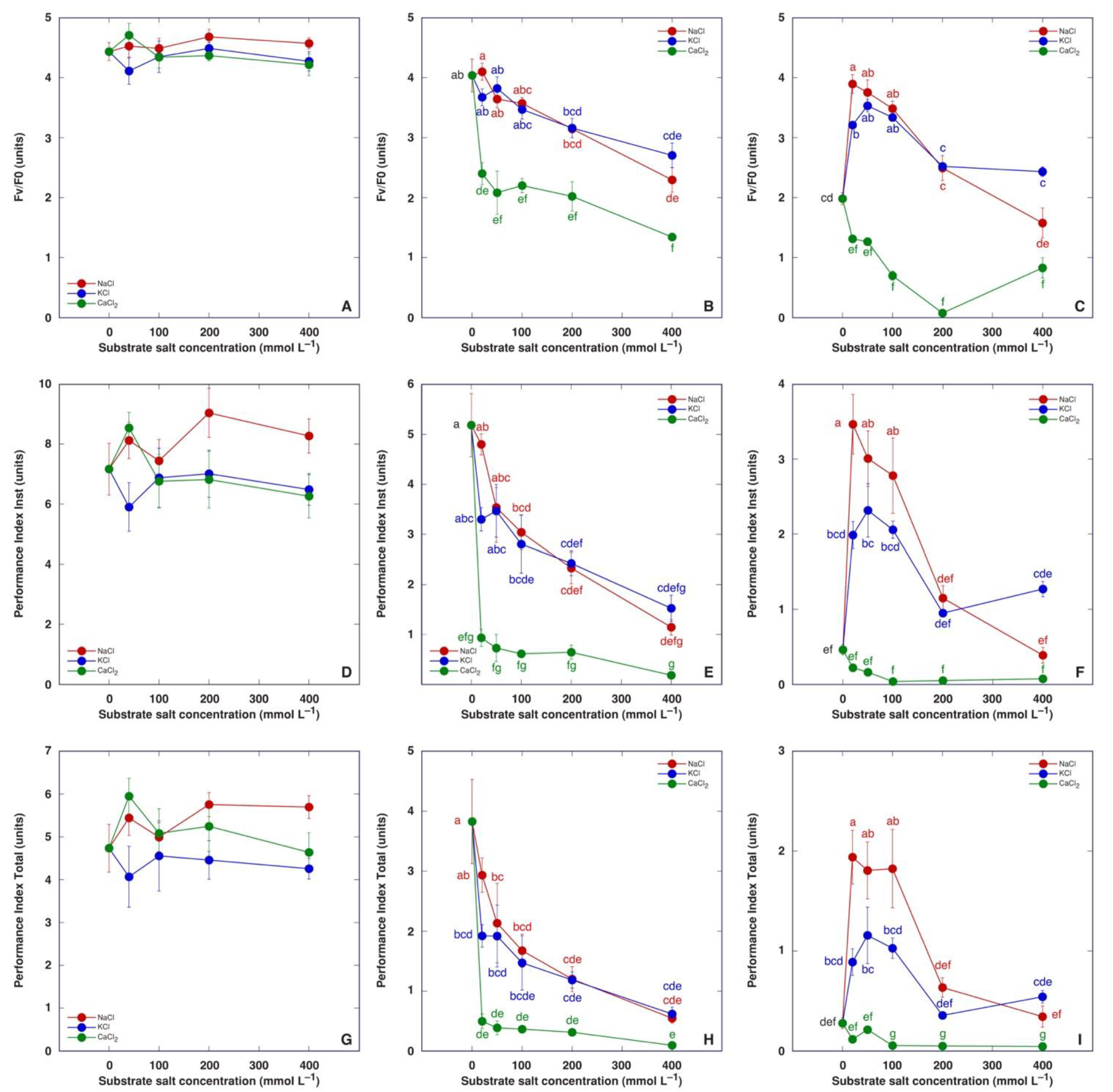
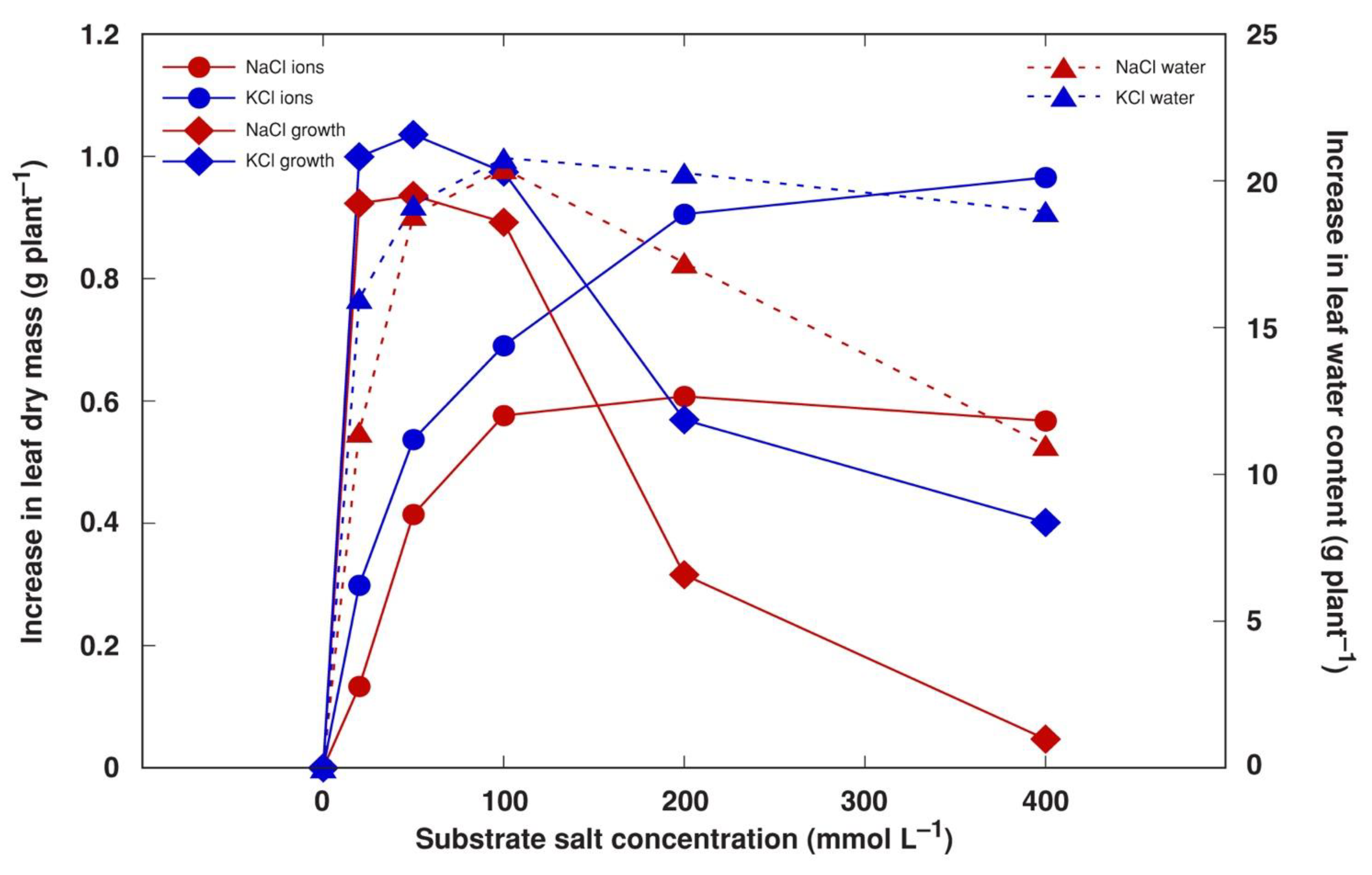
3.1. Experiment 1: Comparison of Six Different Salts
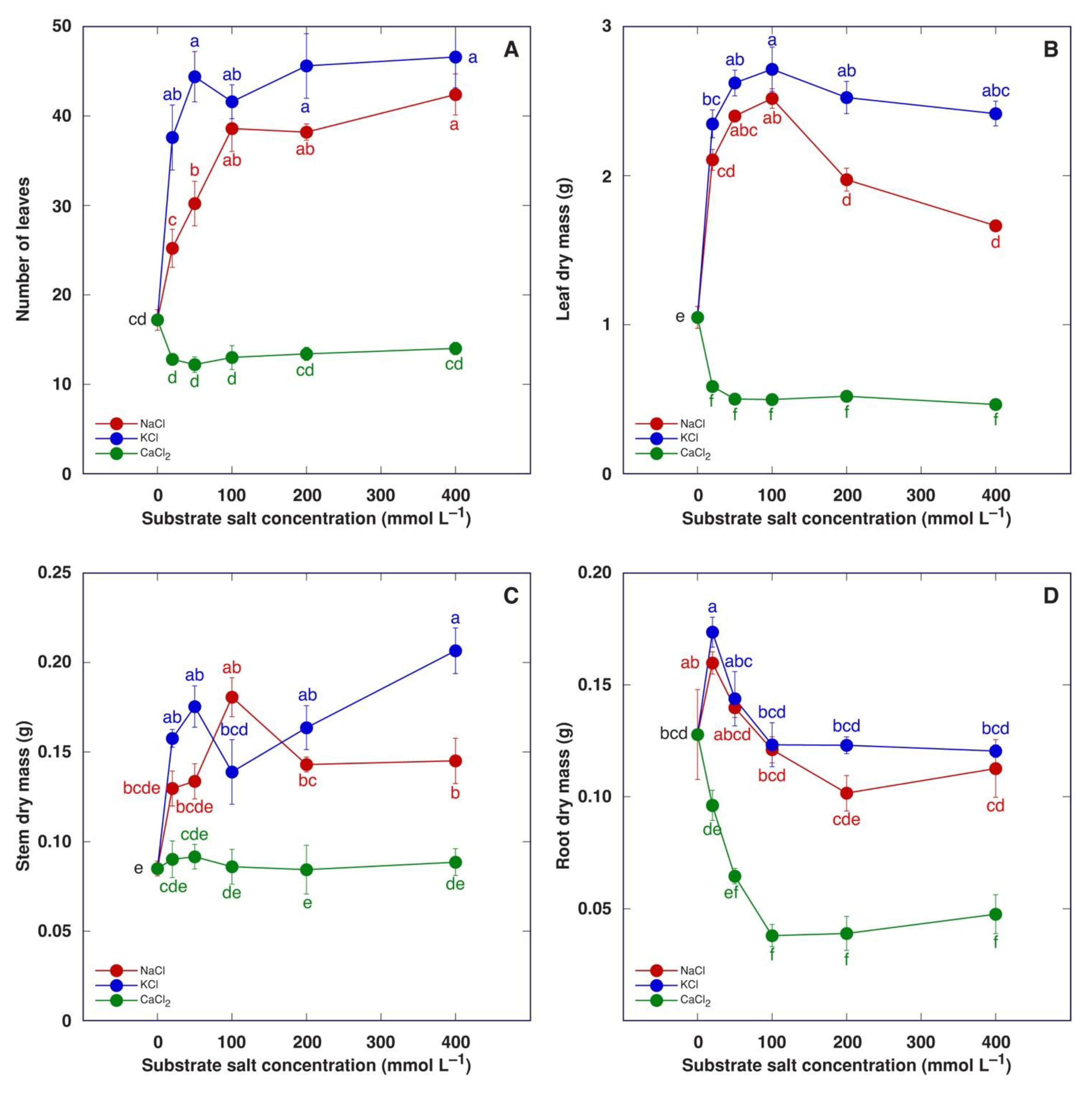
3.2. Experiment 2: Effect of Salinity Gradient of NaCl, KCl and CaCl2
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Duarte, B.; Sleimi, N.; Caçador, I. Biophysical and biochemical constraints imposed by salt stress: Learning from halophytes. Front. Plant Sci. 2014, 5, 746. [Google Scholar] [CrossRef]
- Grigore, M.-N.; Vicente, O. Wild halophytes: Tools for understanding salt tolerance mechanisms of plants and for adapting agriculture to climate change. Plants 2023, 12, 221. [Google Scholar] [CrossRef]
- Lombardi, T.; Bertacchi, A.; Pistelli, L.; Pardossi, A.; Pecchia, S.; Toffanin, A.; Sanmartin, C. Biological and agronomic traits of the main halophytes widespread in the Mediterranean region as potential new vegetable crops. Horticulturae 2022, 8, 195. [Google Scholar] [CrossRef]
- Accogli, R.; Tomaselli, V.; Direnzo, P.; Perrino, E.V.; Albanese, G.; Urbano, M.; Laghetti, G. Edible halophytes and halo-tolerant species in Apulia region (Southeastern Italy); Biogeography, traditional food use and potential sustainable crops. Plants 2023, 12, 549. [Google Scholar] [CrossRef] [PubMed]
- Custódio, L.; Charles, G.; Magné, C.; Barba-Espin, G.; Piqueras, A.; Hernández, J.A.; Ben Hamed, K.; Castañeda-Loaiza, V.; Fernandes, E.; Rodriguez, M.J. Application of in vitro plant tissue culture techniques to halophyte species: A review. Plants 2023, 12, 126. [Google Scholar] [CrossRef]
- Garcia-Caparros, P.; Al-Azzawi, M.J.; Flowers, T.J. Economic uses of salt-tolerant plants. Plants 2023, 12, 2669. [Google Scholar] [CrossRef]
- Hasnain, M.; Abideen, Z.; Ali, F.; Hasanuzzaman, M.; El-Keblawy, A. Potential of halophytes as sustainable fodder production by using saline resources: A review of current knowledge and future directions. Plants 2023, 12, 2150. [Google Scholar] [CrossRef]
- Bohnert, H.J.; Cushman, J.C. The ice plant cometh: Lessons in abiotic stress tolerance. J. Plant Growth Regul. 2000, 19, 334–346. [Google Scholar] [CrossRef]
- Adams, P.; Nelson, D.E.; Yamada, S.; Chmara, W.; Jensen, R.G.; Bohnert, H.J.; Griffiths, H. Growth and development of Mesembryanthemum crystallinum (Aizoaceae). New Phytol. 1998, 138, 171–190. [Google Scholar] [CrossRef] [PubMed]
- Tsukagoshi, H.; Suzuki, T.; Nisjikawa, K.; Agarie, S.; Ishiguro, S.; Higashhiyama, T. RNA-seq analysis of the response of the halophyte, Mesembryanthemum crystallinum (ice plant) to high salinity. PLoS ONE 2015, 10, e0118339. [Google Scholar] [CrossRef]
- Oh, D.-H.; Barkla, B.J.; Vera-Estrella, R.; Pantoja, O.; Lee, S.-Y.; Bohnert, H.J.; Dassanayake, M. Cell type-specific responses to salinity—The epidermal bladder cell transcriptome of Mesembryanthemum crystallinum. New Phytol. 2015, 207, 627–644. [Google Scholar] [CrossRef]
- Chiang, C.-P.; Yim, W.C.; Sun, Y.-H.; Ohnishi, M.; Mimura, T.; Cusjman, J.C.; Yen, H.E. Identification of ice plant (Mesembryanthemum crystallinum L.) microRNAs using NNA-seq and their putative roles in high salinity responses in seedlings. Front. Plant Sci. 2016, 7, 1143. [Google Scholar] [CrossRef]
- Tran, D.Q.; Konishi, A.; Cushman, J.C.; Morokuma, M.; Toyota, M.; Agarie, S. Ion accumulation and expression of ion homeostasis-related genes associated with halophilism, NaCl-promoted growth in a halophyte Mesembryanthemum crystallinum L. Plant Prod. Sci. 2020, 23, 91–102. [Google Scholar] [CrossRef]
- Zhang, X.; Tan, B.; Zhu, D.; Dufresne, D.; Jiang, T.; Chen, S. Proteomics of homeobox7 enhanced salt tolerance in Mesembryanthemum crystallinum. Int. J. Mol. Sci. 2021, 22, 6390. [Google Scholar] [CrossRef]
- Herppich, W.B.; Huyskens-Keil, S.; Schreiner, M. Effects of saline irrigation on growth, physiology and quality of Mesembrynathemum crystallinum L., a rare vegetable crop. J. Appl. Bot. Food Qual. 2008, 82, 47–54. [Google Scholar]
- Atzori, G.; de Vos, A.C.; van Rijsselberghe, M.; Vignolini, P.; Rozema, J.; Mancuso, S.; van Bodegom, P.M. Effects of increased seawater salinity irrigation on growth and quality of the edible Mesembryanthemum crystallinum L. under field conditions. Agric. Water Manag. 2017, 187, 37–46. [Google Scholar] [CrossRef]
- You, X.; Yin, S.; Suo, F.; Xu, Z.; Chu, D.; Kong, Q.; Zhang, S.; Li, Y.; Liu, L. Biochar and fertilizer improved the growth and quality of the ice plant (Mesembryanthemum crystallinum L.) shoots in a coastal soil of Yellow River Delta, China. Sci. Total Environ. 2021, 775, 144893. [Google Scholar] [CrossRef] [PubMed]
- Madhavi, B.G.K.; Choi, G.M.; Bahar, M.E.; Moon, B.E.; Kim, N.E.; Lee, H.-W.; Kim, H.T. Assessment of different salt concentrations on the growth and phytochemical change of the ice plants. J. King Saud Univ. Sci. 2022, 34, 102168. [Google Scholar] [CrossRef]
- He, J.; Koh, S.J.Q.; Qin, L. LED spectral quality and NaCl salinity interact to affect growth, photosynthesis and phytochemical production of Mesembryanthemum crystallinum. Funct. Plant Biol. 2022, 49, 483–495. [Google Scholar] [CrossRef]
- Xia, J.; Mattson, N. Response of common ice plant (Mesembryanthemum crystallinum L.) to sodium chloride concentration in hydroponic nutrient solution. HortScience 2022, 57, 750–756. [Google Scholar] [CrossRef]
- Rodríguez-Hernández, M.C.; Garmendia, I. Optimum growth and quality of the edible ice plant under saline conditions. J. Sci. Food Agric. 2022, 102, 2686–2692. [Google Scholar] [CrossRef]
- Mndi, O.; Sogoni, A.; Jimoh, M.O.; Wilmot, C.M.; Rautenbach, F.; Laubscher, C.P. Interactive effects of salinity stress and irrigation intervals on plant growth, nutritional value, and phytochemical content in Mesembrynathemum crystallinum L. Agriculture 2023, 13, 1026. [Google Scholar] [CrossRef]
- Ghnaya, T.; Nouairi, I.; Slama, I.; Messedi, D.; Grignon, C.; Abdelly, C.; Ghorbel, M.H. Cadmium effects on growth and mineral nutrition of two halophytes: Sesuvium portulacastrum and Mesembryanthemum crystallinum. J. Plant Physiol. 2005, 162, 1133–1140. [Google Scholar] [CrossRef] [PubMed]
- Amari, T.; Ghnaya, T.; Debez, A.; Taamali, M.; Yousseh, N.B.; Lucchini, G.; Sacchi, G.A.; Abdelly, C. Comparative Ni tolerance and accumulation potentials between Messembryanthemum crystallinum (halophyte) and Brassica juncea: Metal accumulation, nutrient status and photosynthetic activity. J. Plant Physiol. 2014, 171, 1634–1644. [Google Scholar] [CrossRef] [PubMed]
- Śliwa-Cebula, M.; Kaszycki, P.; Kaczmarczyk, A.; Nosek, M.; Lis-Kryścin, A.; Miszalski, Z. The common ice plant (Mesembryanthemum crystallinum L.)—Phytoremediation potential for cadmium and chromate-contaminated soils. Plants 2020, 9, 1230. [Google Scholar] [CrossRef]
- Śliwa-Cebula, M.; Koniarz, T.; Szara-Bąk, M.; Baran, A.; Miszalski, Z.; Kaszycki, P. Phytoremediation of metal-contaminated bottom sediments by the common ice plant (Mesembryanthmum crystallinum L.) in Poland. J. Soils Sedim. 2022, 23, 1065–1082. [Google Scholar] [CrossRef]
- Broetto, F.; Duarte, H.M.; Lüttge, U. Responses of chlorophyll fluorescence parameters of the facultative halophyte and C3–CAM intermediate species Mesembryanthemum crystallinum to salinity and high irradiance stress. J. Plant Physiol. 2007, 164, 904–912. [Google Scholar] [CrossRef]
- Ślesak, I.; Libik, M.; Miszalski, Z. The foliar concentration of hydrogen peroxide during salt-induced C3-CAM transition in Mesembryanthemum crystallinum L. Plant Sci. 2008, 174, 221–226. [Google Scholar] [CrossRef]
- Vernon, D.M.; Bohnert, H.J. A novel methyl transferase induced by osmotic stress in the facultative halophyte Mesembryanthemum crystallinum. EMBO J. 1992, 11, 2077–2085. [Google Scholar] [CrossRef] [PubMed]
- Matoh, T.; Watanabe, J.; Takahashi, E. Effects of sodium and potassium salts on the growth of a halophyte Atriplex gmelini. Soil Sci. Plant Nutr. 1986, 32, 451–459. [Google Scholar] [CrossRef]
- Reginato, M.; Sosa, L.; Llanes, A.; Hampp, E.; Vettorazzi, N.; Reinoso, H.; Luna, V. Growth responses and ion accumulation in the halophytic legume Prosopis strombulifera are determined by Na2SO4 and NaCl. Plant Biol. 2014, 16, 97–106. [Google Scholar] [CrossRef] [PubMed]
- Ievinsh, G.; Andersone-Ozola, U.; Jēkabsone, A. Similar responses of relatively salt tolerant plants to Na and K during chloride salinity: Comparison of growth, water content and ion accumulation. Life 2022, 12, 1577. [Google Scholar] [CrossRef]
- Wang, D.; Wang, H.; Han, B.; Wang, B.; Guo, A.; Zheng, D.; Liu, C.; Chang, L.; Peng, M.; Wang, X. Sodium instead of potassium and chloride is an important macronutrient to improve leaf succulence and shoot development for halophyte Sesuvium portulacastrum. Plant Physiol. Biochem. 2012, 51, 53–62. [Google Scholar] [CrossRef]
- Ievinsh, G.; Landorfa-Svalbe, Z.; Andersone-Ozola, U.; Karlsons, A.; Osvalde, A. Salinity and heavy metal tolerance, and phytoextraction potential of Ranunculus sceleratus plants from a sandy coastal beach. Life 2022, 12, 1959. [Google Scholar] [CrossRef]
- Jēkabsone, A.; Kuļika, J.; Romanovs, M.; Andersone-Ozola, U.; Ievinsh, G. Salt tolerance and ion accumulation in several halophytic plant species depending on the type of anion. Int. J. Plant Biol. 2023, 14, 1131–1154. [Google Scholar] [CrossRef]
- Landorfa-Svalbe, Z.; Andersone-Ozola, U.; Ievinsh, G. Type of anion largely determines salinity tolerance in four Rumex species. Plants 2023, 12, 92. [Google Scholar] [CrossRef] [PubMed]
- Flowers, T.J.; Colmer, T.D. Plant salt tolerance: Adaptations in halophytes. Ann. Bot. 2015, 115, 327–331. [Google Scholar] [CrossRef] [PubMed]
- Jēkabsone, A.; Andersone-Ozola, U.; Karlsons, A.; Romanovs, M.; Ievinsh, G. Effect of salinity on growth, ion accumulation and mineral nutrition of different accessions of a crop wild relative legume species, Trifolium fragiferum. Plants 2022, 11, 797. [Google Scholar] [CrossRef]
- Farzana, T.; Guo, Q.; Rahman, M.S.; Rose, T.J.; Barkla, B.J. Salinity and nitrogen source affect productivity and nutritional value of edible halophytes. PLoS ONE 2023, 18, e0288547. [Google Scholar] [CrossRef]
- Khan, M.A.; Ungar, I.A.; Showalter, A.M. Salt stimulation and tolerance in an intertidal stem-succulent halophyte. J. Plant Nutr. 2005, 28, 1365–1374. [Google Scholar] [CrossRef]
- Kalaji, H.M.; Jajoo, A.; Oukarroum, A.; Brestic, M.; Zivcak, M.; Samborska, I.A.; Cetner, M.D.; Łukasik, I.; Goltsev, V.; Ladle, R.J. Chlorophyll a fluorescence as a tool to monitor physiological status of plants under abiotic stress conditions. Acta Physiol. Plant. 2016, 38, 102. [Google Scholar] [CrossRef]
- Tsimilli-Michael, M. Revisiting JIP-test: An educative review on concepts, assumptions, approcimations, definitions and terminology. Photosynthetica 2020, 58, 275–292. [Google Scholar] [CrossRef]
- Swoczyna, T.; Kalaji, H.M.; Bussotti, F.; Mojski, J.; Pollastrini, M. Environmental stress—What can we learn from chlorophyll a fluorescence analysis in woody plants? A review. Front. Plant Sci. 2022, 13, 1048582. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Li, L.; Qin, Y.; Lu, B.; Xu, D.; Zhuang, W.; Shu, X.; Zhang, F.; Wang, N.; Wang, Z. Effects of seasonal changes on chlorophyll fluorescence and physiological characteristics in the two Taxus species. Plants 2023, 12, 2636. [Google Scholar] [CrossRef] [PubMed]
- Banks, J.M. Continuous excitation chlorophyll fluorescence parameters: A review for practicioners. Tree Physiol. 2017, 37, 1128–1136. [Google Scholar] [CrossRef] [PubMed]
- Andersone-Ozola, U.; Jēkabsone, A.; Karlsons, A.; Romanovs, M.; Ievinsh, G. Soil chemical properties and mineral nutrition of Latvian accessions of Trifolium fragiferum, a crop wild relative plant species. Environ. Exp. Biol. 2021, 19, 245–254. [Google Scholar]
- Metsalu, T.; Vilo, J. ClustVis: A web tool for visualizing clustering of multivariate data using Principal Component Analysis and heatmap. Nucleic Acids Res. 2015, 43, W566–W570. [Google Scholar] [CrossRef]
- Yuan, F.; Xu, Y.; Leng, B.; Wang, B. Beneficial effects of salt on halophyte growth: Morphology, cells, and genes. Open Life Sci. 2019, 14, 191–200. [Google Scholar] [CrossRef]
- Rodríguez-Hernández, M.C.; Garmendia, I. Age of plant influences the effect of salinity in yield and mineral content of ice plant. J. Appl. Bot. Food Qual. 2022, 95, 94–99. [Google Scholar]
- Flowers, T.J.; Hajibagheri, M.A.; Clipson, N.J.W. Halophytes. Quart. Rev. Biol. 1986, 61, 313–337. [Google Scholar] [CrossRef]
- Flowers, T.J.; Colmer, T.D. Salinity tolerance in halophytes. New Phytol. 2008, 179, 945–963. [Google Scholar] [CrossRef] [PubMed]
- Lv, S.; Nie, L.; Fan, P.; Wang, X.; Jiang, D.; Chen, X.; Li, Y. Sodium plays a more important role than potassium and chloride in growth of Salicornia europaea. Acta Physiol. Plant. 2012, 34, 503–513. [Google Scholar] [CrossRef]
- Jennings, D.H. Halophytes, succulence and sodium in plants—A unified theory. New Phytol. 1968, 67, 899–911. [Google Scholar] [CrossRef]
- Vera-Estrella, R.; Barkla, B.J.; Bohnert, H.J.; Pantoja, O. Salt stress in Mesembryanthemum crystallinum L. cell suspensions activates adaptive mechanisms similar to those observed in the whole plant. Planta 1999, 207, 426–435. [Google Scholar] [CrossRef] [PubMed]
- Prokopoviča, V.; Ievinsh, G. Ranunculus sceleratus as a model species to decrypt the role of ethylene in plant adaptation to salinity. Plants 2023, 12, 370. [Google Scholar] [CrossRef]
- White, P.J.; Broadley, M.R. Calcium in plants. Ann. Bot. 2003, 92, 487–511. [Google Scholar] [CrossRef]
- Amor, N.B.; Megdiche, W.; Jiménez, A.; Sevilla, F.; Abdelly, C. The effect of calcium on the antioxidant systems in the halophyte Cakile maritima under salt stress. Acta Physiol. Plant. 2010, 32, 453–461. [Google Scholar] [CrossRef]
- Andersone-Ozola, U.; Gaile, L.; Ievinsh, G. Physiological responses of rare coastal marsh plant Triglochin maritima L. to soil chemical heterogeneity. Acta Biol. Univ. Daugavp. 2017, 17, 149–155. [Google Scholar]
- Shimada, N. Excess injury of calcium and magnesium in the crops. Japan Agric. Res. Q. 1972, 7, 173–177. [Google Scholar]
- Nagrão, S.; Schmckel, S.M.; Tester, M. Evaluating physiological responses of plants to salinity stress. Ann. Bot. 2017, 119, 1–11. [Google Scholar] [CrossRef]
- Barkla, B.J.; Vera-Estrella, R.; Camacho-Emiterio, J.; Pantoja, O. Na+/H+ exchange in the halophyte Mesembryanthemum crystallinum L. is associated with cellular sites of Na+ storage. Funct. Plant Biol. 2002, 29, 1017–1024. [Google Scholar] [CrossRef]
- Percey, W.J.; Shabala, L.; Wu, Q.; Su, N.; Breadmore, M.C.; Guijt, R.M.; Bose, J.; Shabala, S. Potassium retention in leaf mesophyll as an element of salinity tissue tolerance in halophytes. Plant Physiol. Biochem. 2016, 109, 346–354. [Google Scholar] [CrossRef] [PubMed]
- Jarrell, W.M.; Beverly, R.B. The dilution effect in plant nutrition studies. Adv. Agron. 1981, 34, 197–224. [Google Scholar]
- Riedell, W.E. Mineral-nutrient synergism and diliution responses to nitrogen fertilizer in field-grown maize. J. Plant Nutr. Soil Sci. 2010, 173, 869–874. [Google Scholar] [CrossRef]
- Li, X.; Dong, J.; Gruda, N.; Chu, W.; Duan, Z. Does the short-term fluctuation of mineral element concentrations in the closed hydroponic experimental facilities affect the mineral concentrations in cucumber plants exposed to elevated CO2? Plant Soil 2021, 465, 125–141. [Google Scholar] [CrossRef]
- Keiller, D.R.; Slocombe, S.P.; Cockburn, W. Analysis of chlorophyll a fluorescence in C3 and CAM forms of Mesembryanthemum crystallinum. J. Exp. Bot. 1994, 45, 325–334. [Google Scholar] [CrossRef]
- Niewiadomska, E.; Pilarska, M. Acclimation to salinity in halophytic ice plant prevents a decline of linear electron transport. Environ. Exp. Bot. 2021, 184, 104401. [Google Scholar] [CrossRef]
- Ose, A.; Andersone-Ozola, U.; Ievinsh, G. Substrate-dependent effect of vermicompost on yield and physiological indices of container-grown Dracocephalum moldavica plants. Agriculture 2021, 11, 1231. [Google Scholar] [CrossRef]
- Wang, H.; Liu, R.L.; Jin, J.Y. Effects of zinc and soil moisture on photosynthetic rate and chlorophyll fluorescence parameters of maize. Biol. Plant. 2009, 53, 191–194. [Google Scholar] [CrossRef]
- Horaczek, T.; Dąbrowski, P.; Kalaji, H.M.; Baczewska-Dąbrowska, A.H.; Pietkiewicz, S.; Stępień, W.; Gozdowski, D. JIP-test as a tool for early detection of the macronutrients deficiency in Miscanthus plants. Photosynthetica 2020, 58, 507–517. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Na (g kg−1) | K (g kg−1) | Cl (g kg−1) | Ca (g kg−1) | P (g kg−1) | Mg (g kg−1) | Fe (mg kg−1) | Mn (mg kg−1) | Zn (mg kg−1) | Cu (mg kg−1) |
|---|---|---|---|---|---|---|---|---|---|---|
| Control | 3.1 ± 0.1 h | 16.7 ± 0.1 g | 1.8 ± 0.1 i | 8.1 ± 0.4 d | 4.3 ± 0.6 c | 7.0 ± 0.5 bc | 111 ± 15 abc | 253 ± 31 ab | 94 ± 9 b | 8.1 ± 0.3 cde |
| NaCl 20 mM | 12.1 ± 0.3 fgh | 8.3 ± 0.5 g | 53.8 ± 1.9 h | 5.0 ± 0.2 d | 2.5 ± 0.0 de | 3.3 ± 0.2 de | 67 ± 4 de | 166 ± 12 cd | 45 ± 1 cd | 4.2 ± 0.3 g |
| NaCl 50 mM | 86.6 ± 3.2 d | 7.4 ± 1.0 g | 88.3 ± 2.2 ef | 5.3 ± 1.1 d | 2.2 ± 0.1 e | 2.9 ± 0.4 de | 87 ± 12 bcd | 123 ± 6 cd | 33 ± 1 d | 3.9 ± 0.2 g |
| NaCl 100 mM | 125.1 ± 4.1 c | 8.5 ± 0.2 g | 105.8 ± 6.8 de | 6.3 ± 0.2 d | 2.5 ± 0.1 de | 3.3 ± 0.5 de | 73 ± 3 cde | 163 ± 13 cd | 36 ± 4 cd | 5.3 ± 1.1 fg |
| NaCl 200 mM | 158.2 ± 3.2 b | 10.7 ± 0.4 g | 152.5 ± 3.8 c | 5.2 ± 0.3 d | 2.8 ± 0.1 de | 2.4 ± 0.1 e | 73 ± 10 cde | 165 ± 26 cd | 40 ± 1 cd | 6.2 ± 0.4 efg |
| NaCl 400 mM | 179.2 ± 5.6 a | 11.3 ± 1.9 g | 165.0 ± 4.3 bc | 4.8 ± 0.3 d | 3.4 ± 0.1 cd | 2.3 ± 0.1 e | 62 ± 7 de | 148 ± 23 cd | 53 ± 3 c | 6.1 ± 0.7 efg |
| KCl 20 mM | 2.2 ± 0.1 h | 80.5 ± 2.3 de | 55.0 ± 2.2 h | 5.7 ± 0.4 d | 1.7 ± 0.1 e | 3.2 ± 0.2 de | 62 ± 4 de | 106 ± 6 cd | 41 ± 5 cd | 5.1 ± 0.3 fg |
| KCl 50 mM | 2.1 ± 0.2 h | 126.1 ± 2.5 c | 86.3 ± 4.0 f | 9.1 ± 2.0 d | 1.9 ± 0.0 e | 4.2 ± 0.7 d | 76 ± 4 cde | 178 ± 16 bc | 53 ± 1 c | 7.4 ± 0.5 def |
| KCl 100 mM | 3.5 ± 0.3 h | 146.5 ± 5.2 b | 115.0 ± 2.5 d | 6.9 ± 0.2 d | 2.0 ± 0.1 e | 3.0 ± 0.1 de | 71 ± 11 de | 137 ± 12 cd | 51 ± 3 c | 6.4 ± 0.4 efg |
| KCl 200 mM | 6.0 ± 0.2 gh | 195.6 ± 5.4 a | 170.8 ± 3.3 b | 9.6 ± 0.6 d | 2.0 ± 0.1 e | 3.4 ± 0.1 de | 51 ± 3 de | 140 ± 13 cd | 42 ± 1 cd | 6.1 ± 0.2 efg |
| KCl 400 mM | 5.4 ± 1.1 h | 205.9 ± 5.0 a | 201.7 ± 7.3 a | 8.8 ± 0.9 d | 2.1 ± 0.0 e | 3.1 ± 0.1 de | 51 ± 6 e | 92 ± 4 d | 43 ± 3 cd | 4.3 ± 0.3 g |
| CaCl2 20 mM | 9.7 ± 0.3 gh | 39.8 ± 0.2 f | 63.8 ± 1.4 gh | 34.4 ± 0.6 c | 6.3 ± 0.2 b | 11.4 ± 0.0 a | 147 ± 0 b | 305 ± 15 a | 128 ± 4 a | 10.3 ± 1.1 abc |
| CaCl2 50 mM | 12.7 ± 0.1 fgh | 67.5 ± 1.9 e | 81.9 ± 0.4 fg | 54.1 ± 0.1 b | 7.2 ± 0.3 ab | 10.8 ± 0.1 a | 139 ± 2 ab | 308 ± 10 a | 138 ± 2 a | 8.9 ± 0.1 abcd |
| CaCl2 100 mM | 16.4 ± 0.1 efg | 79.9 ± 2.8 de | 91.9 ± 3.3 ef | 71.4 ± 3.1 a | 7.4 ± 0.5 ab | 8.5 ± 0.1 b | 119 ± 4 ab | 289 ± 6 a | 134 ± 3 a | 10.2 ± 0.5 bcde |
| CaCl2 200 mM | 21.2 ± 0.1 ef | 90.0 ± 1.7 d | 100.0 ± 0.7 def | 77.8 ± 6.2 a | 7.9 ± 0.0 a | 7.2 ± 0.3 bc | 132 ± 4 a | 276 ± 1 a | 133 ± 3 a | 11.1 ± 0.6 ab |
| CaCl2 400 mM | 25.0 ± 0.2 e | 88.0 ± 6.4 d | 111.3 ± 2.2 d | 76.1 ± 1.5 a | 7.1 ± 0.0 ab | 6.6 ± 0.2 c | 137 ± 7 ab | 293 ± 13 a | 144 ± 1 a | 12.9 ± 0.1 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jēkabsone, A.; Karlsons, A.; Osvalde, A.; Ievinsh, G. Effect of Na, K and Ca Salts on Growth, Physiological Performance, Ion Accumulation and Mineral Nutrition of Mesembryanthemum crystallinum. Plants 2024, 13, 190. https://doi.org/10.3390/plants13020190
Jēkabsone A, Karlsons A, Osvalde A, Ievinsh G. Effect of Na, K and Ca Salts on Growth, Physiological Performance, Ion Accumulation and Mineral Nutrition of Mesembryanthemum crystallinum. Plants. 2024; 13(2):190. https://doi.org/10.3390/plants13020190
Chicago/Turabian StyleJēkabsone, Astra, Andis Karlsons, Anita Osvalde, and Gederts Ievinsh. 2024. "Effect of Na, K and Ca Salts on Growth, Physiological Performance, Ion Accumulation and Mineral Nutrition of Mesembryanthemum crystallinum" Plants 13, no. 2: 190. https://doi.org/10.3390/plants13020190
APA StyleJēkabsone, A., Karlsons, A., Osvalde, A., & Ievinsh, G. (2024). Effect of Na, K and Ca Salts on Growth, Physiological Performance, Ion Accumulation and Mineral Nutrition of Mesembryanthemum crystallinum. Plants, 13(2), 190. https://doi.org/10.3390/plants13020190