Abstract
Volatile compounds play indispensable roles in the interactions among host plants, herbivores and natural enemies. Previous studies showed that the addition of buckwheat strips in cotton fields could attract Peristenus spretus, the dominant parasitoid of Apolygus lucorum, and enhance its parasitic activity. Through the combined analysis of Y-tube olfactometer, solid-phase microextraction (SPME), gas chromatography-mass spectrometer (GC-MS) and electroantennography (EAG), we found that male and female P. spretus responded to compounds present in buckwheat flowers. The five major components of buckwheat flowers, cis-3-hexenyl acetate (Z3HA), 4-methylanisole, 4-oxoisophorone, p-methylphenol and 2-ethylhexyl salicylate, all had a significant attraction to P. spretus adults and led to positive electroantennogram responses, especially for 10 mg/mL 4-oxoisophorone, indicating the components played a key role in the selection behavior of P. spretus to buckwheat flowers. Additionally, field trials showed that the five volatiles could significantly increase the parasitism by P. spretus. Our study screened the key active components of buckwheat flower volatiles that have an attractive effect on P. spretus, revealing its behavioral selection mechanism and emphasizing the important role of plant volatiles on host selection and parasitism of parasitic wasps, providing a theoretical basis for the development of attractants for P. spretus and the reduction of pesticides in the field to promote conservation biological control (CBC) of A. lucorum.
1. Introduction
Conservation biological control (CBC) refers to protecting natural enemies or improving their control ability by changing the field environment or pesticide application methods [1]. Compared with classical or augmentative biological control, CBC pays more attention to the conservation and protection of natural enemy insect populations [2], which is an effective way to achieve sustainable pest control and maintain ecological balance. Utilizing the volatile organic compounds (VOCs) to attract natural enemies, increasing the number of attracted natural enemies or establishing a “Push-Pull” system [3,4] are major research focuses of CBC.
VOCs are volatile secondary substances continuously released by plants during their growth and development processes [5], playing an irreplaceable role in maintaining the ecological balance in tritrophic interactions among host plants, herbivorous insects and natural enemies. VOCs provide herbivores with cues for long-distance host identification and orientation, acquisition of nutritional resources and mating sites, as well as guidance on oviposition and mating behaviors [6]. In addition, VOCs can act as information substances for natural enemy searching behavior [6,7,8,9]. In corn fields, nerolidol, cis-jasmone and trans-β-farnesene were effective in attracting natural enemies [10]. Three volatiles from the pagoda tree Sophora japonica L., α-pinene, linalool and hexanal at the concentrations of 10−4~10−6 g/mL, 10−4 g/mL and 10−5 g/mL, respectively, were significantly attractive to the ladybug, Harmonia axyridis (Pallas), an important natural enemy of insect pests such as scale insects and aphids [11]. The volatiles bornylene, 6-methyl-5-hepten-2-one and (+)-6-methyl-5-hepten-2-ol induced by wheat aphid were the most important chemical signaling substances for the parasitoid Aphidius avenae Haliday, as well as predatory Coccinella septempunctata L., Propylaea japonica (Thunberg), Chrysoperla sinica (Tjeder) and Chrysopa pallens (Rambur) to search for their hosts [12,13].
The mirid bug, Apolygus lucorum (Hemiptera: Miridae), is one of the most important pests of cotton, cereals, vegetables and fruit crops in China, especially in the Yangtze River Basin and the Yellow River Basin [14]. A. lucorum adults and nymphs can damage host plants through piercing and sucking feeding activities, and their wide environmental adaptability, high population growth rate and strong dispersal ability can lead to a loss in cotton yield of 20–30% in some years [15]. A. lucorum, especially males, had a strong electrophysiological response to butyrate and green leaf volatiles (GLVs) [16]. Previous studies showed that fragrant volatiles emitted by flowers could mediate the preference of A. lucorum for flowering host plants [17]. Moreover, temporal shifts in plant volatile emission may modulate host plant foraging of A. lucorum and appear to guide its colonization of different host plants [18]. When applied to fields, flower-emitted VOCs acted as attractants for several mirids, including Adelphocoris suturalis, Ad. lineolatus and Ad. fasciaticollis [19].
Peristenus spretus Chen et van Achterberg (Hymenoptera: Braconidae) is a key solitary koinobiont endoparasitoid native to China, with a distribution range of 23–39° N [20,21]. When P. spretus was released in field cages at the rate of one female parasitoid per 50 A. lucorum nymphs, the parasitism rate reached 77.8% [22]. The foraging behavior of natural enemies can be regulated by olfactory cues, such as the odor produced by flowers and vegetative plant parts [2,23]. P. spretus females use volatiles released by plants infested by A. lucorum nymphs to locate host plants with potential hosts, and the behavioral attraction of P. spretus to flowering host plants is similar to that of adult A. lucorum [24].
Buckwheat, Fagopyrum esculentum Moench, is a valuable nectary source for parasitoids and also a common grain crop in China with a long flowering period and well-developed nectaries [25]. Buckwheat is widely used in CBC due to its value to improve the performance of parasitic wasps [23,26]. The introduction of flowering plants in or around cultivated fields has been used to enhance the control of pests by natural enemies [27]. For example, adding 2 m-wide buckwheat strips in cotton fields attracted P. spretus and enhanced their parasitic activity [28]. We hypothesize that buckwheat flower volatiles are a key factor in the attraction of buckwheat to parasitic wasps and may have a positive effect on their function in the field. Further, knowledge of the bioactive volatiles produced by buckwheat might permit use of synthetic analogs to attract P. spretus to crop fields without sacrificing part of the field for buckwheat production.
In this study, we used a Y-tube olfactometer, solid-phase microextraction (SPME), gas chromatography-mass spectrometer (GC-MS) and electroantennography (EAG) to assess the behavioral and physiological responses of P. spretus adults to volatiles produced by buckwheat flowers under laboratory and field conditions.
2. Results
2.1. Behavioral Responses to Buckwheat Plants
The behavioral responses of adult wasps to volatiles from buckwheat flowers in a Y-tube olfactometer were assessed and analyzed, indicating that volatile odors from buckwheat flowers have a significant trapping effect on both male (64.15%) and female (65.22%) P. spretus (female: χ2 = 4.26, df = 1, p = 0.04; male: χ2 = 4.25, df = 1, p = 0.04) compared with clean air (Figure 1).
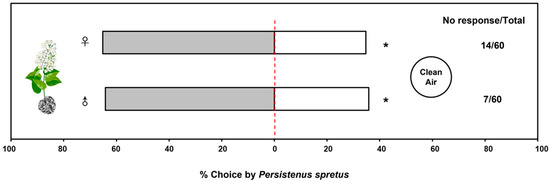
Figure 1.
Behavioral response of Peristenus spretus adults to buckwheat plants in a Y-tube olfactometer. Data in the horizontal bar chart are the percentage of individuals that responded in each treatment out of the total number of individuals that responded, and the numbers next to the chart indicate the number of individuals who did not respond to either treatment out of the total individuals in the trial. 0.01 < p < 0.05 (*) means significantly difference.
2.2. Analysis of Buckwheat Volatiles
After collecting buckwheat flower volatiles through SPME technology, the major substances in buckwheat flower volatiles were screened and identified by GC-MS, and five compounds were obtained, while compounds often associated with air (e.g., toluene and benzene) or laboratory equipment (e.g., siloxanes or phthalates) were not included in our list of putative plant volatiles [29]. According to the sequence of GC retention time (RT), they were (1) cis-3-hexenyl acetate (Z3HA, RT = 10.07 min), (2) 4-methylanisole (RT = 11.12 min), (3) 4-oxoisophorone (RT = 16.30 min), (4) p-methylphenol (RT = 24.93 min) and (5) 2-ethylhexyl salicylate (RT = 29.16 min). Additionally, the compound Z3HA had the greatest concentration (128.31 ± 33.60 mg/mL), followed by p-methylphenol (5.48 ± 2.41 mg/mL), 2-ethylhexyl salicylate (1.87 ± 0.11 mg/mL), 4-methylanisole (1.56 ± 0.52 mg/mL) and 4-oxoisophorone (1.49 ± 0.4 mg/mL) (Figure 2).
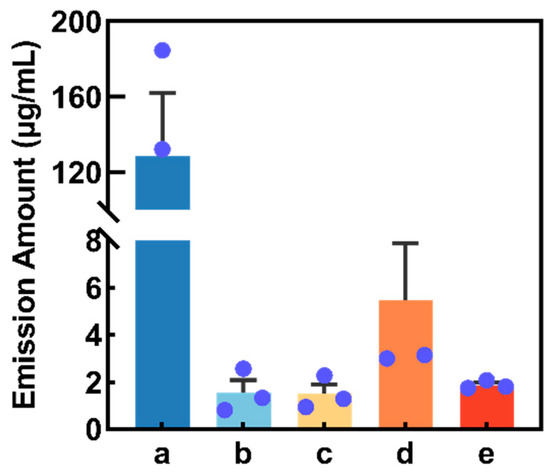
Figure 2.
Concentration of volatile compounds collected from buckwheat flowers (4 h collection period). The blue dot in the figure represents the original data. (a) cis-3-Hexenyl acetate. (b) 4-Methylanisole. (c) 4-Oxoisophorone. (d) p-Methylphenol. (e) 2-Ethylhexyl salicylate.
2.3. EAG Responses to Single Compounds
The EAG response relative value of male P. spretus increased with increasing concentrations for all compounds identified by GC-MS and reached the maximum value at 10 mg/mL for 4-oxoisophorone. Equally, the EAG response relative value for female P. spretus increased with the increase of the concentrations of Z3HA, 4-methylanisole and p-methylphenol and was the largest at 10 mg/mL 4-oxoisophorone. However, the EAG amplitude response of females decreased at 0.1 mg/mL 4-oxoisophorone, as well as 0.1 and 1 mg/mL 2-ethylhexyl salicylate (Figure 3).
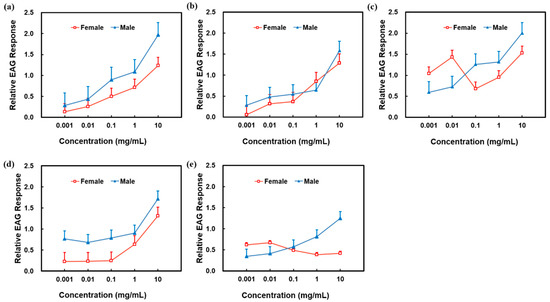
Figure 3.
EAG response of Peristenus spretus antennae to buckwheat flowers. EAG relative value = (EAG amplitude of the test stimulus − mean EAG amplitude of the control)/(mean EAG amplitude of the reference compound − mean EAG amplitude of the control). (a) cis-3-Hexenyl acetate. (b) 4-Methylanisole. (c) 4-Oxoisophorone. (d) p-Methylphenol. (e) 2-Ethylhexyl salicylate.
2.4. Behavioral Responses to Principal Compounds
In a Y-tube olfactometer, the above-mentioned five electrophysiological volatiles all had significant attraction to male (Z3HA: χ2 = 18.62, df = 1, p < 0.01; 4-methylanisole: χ2 = 35.00, df = 1, p < 0.01; p-methylphenol: χ2 = 36.00, df = 1, p < 0.01; 4-oxoisophorone: χ2 = 20.57, df = 1, p < 0.01; 2-ethylhexyl salicylate: χ2 = 18.00, df = 1, p < 0.01) and female (Z3HA: χ2 = 25.00, df = 1, p < 0.01; 4-methylanisole: χ2 = 28.00, df = 1, p < 0.01; p-methylphenol: χ2 = 20.57, df = 1, p < 0.01; 4-oxoisophorone: χ2 = 14.29, df = 1, p < 0.01; 2-ethylhexyl salicylate: χ2 = 8.00, df = 1, p < 0.01) parasitoids. Thus, all five compounds played key roles in the preference for buckwheat flowers by P. spretus (Figure 4).
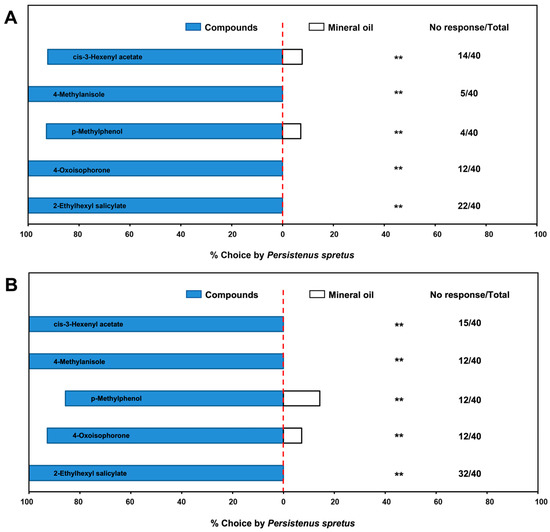
Figure 4.
Behavioral response of Peristenus spretus males (A) and females (B) to main components of buckwheat flower volatiles in a Y-tube olfactometer. Data in the horizontal bar chart are the percentage of individuals that responded in each treatment out of the total number of individuals that responded, and the numbers next to the chart indicate the number of individuals who did not respond to either treatment out of the total individuals in the trial. p < 0.01 (**) means highly significantly different.
2.5. Parasitism in the Field
Field trials showed that the five main components of buckwheat flower volatiles significantly enhanced parasitism of A. lucorum nymphs by P. spretus in both cotton fields and vineyards (cotton field: F = 5.92, df = 5, 71, p < 0.001; vineyard: F = 12.46, df = 5, 71, p < 0.001). In cotton fields and vineyards, the five volatiles of buckwheat flowers increased parasitism by 28.25~40.78% and 18.05~27.86%, rezspectively, with the most significant potentiation of parasitism by Z3HA (Figure 5).
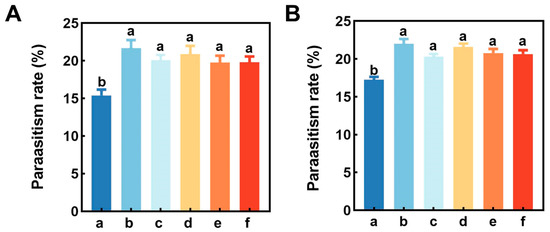
Figure 5.
Parasitism rate of Apolygus lucorum nymphs by Peristenus spretus wasps in barrel traps baited with 10 mg/mL concentration of buckwheat major volatile compounds in cotton field (A) and vineyard (B) test areas. We have added the meaning of the lower case letters above the bar chart. (a) Mineral oil, (b) cis-3-Hexenyl acetate, (c) 4-Methylanisole, (d) 4-Oxoisophorone, (e) p-Methylphenol, (f) 2-Ethylhexyl salicylate. Letters above the bars indicate significant differences.
3. Discussion
Plant volatiles play an important role in the trophic relationship, not only providing beneficial and herbivorous insects with clues related to food orientation, habitat location and oviposition sites but also helping predatory and parasitic natural enemies locate their hosts, thus enabling plants to engage in indirect defense or enhance their direct resistance to herbivores [30,31]. In addition, plants and herbivores often determine the physiological and behavioral responses of parasitic wasps. In this study, we found the flowers of the nectar source plant, buckwheat, were significantly attractive to P. spretus adults. Further, we identified five floral volatiles (Z3HA, 4-methylanisole, 4-oxoisophorone, p-methylphenol and 2-ethylhexyl salicylate) from buckwheat flowers, all of which had electrophysiological and behavioral activities on both male and female P. spretus adults, indicating that these compounds played a key role in the attraction of P. spretus to buckwheat flowers. Our results suggest that integration of buckwheat strips into crop fields may enhance CBC of A. lucorum by P. spretus.
The main components of buckwheat flower volatiles were Z3HA and p-methylphenol which were confirmed to be attractive to P. spretus males and females through the behavioral determination of electrophysiological volatiles. P-Methylphenol is often used as bait for traps due to its enticing effect on pests and parasitic wasps [32]. Z3HA is one of the dominant GLVs emitted by wounded plants [33], as well as the main component of some floral volatiles, especially in the HS-SPME of flowers, such as Jasminum sambac [34], Rubus idaeus [35], Lonicera japonica (Thunberg) [36] and living rose [37]. In addition, the attraction effect of Z3HA on parasitic wasps has also been demonstrated in other parasitoids. Campoletis chlorideae Uchida is an endophytic wasp of Helicoverpa armigera (Hübner). Z3HA not only had an attractive effect on the parasitoid, but a specific dose of Z3HA significantly increased parasitism of H. armigera by C. chlorideae [38]. Thus, use of synthetic analogs of plant volatiles which are attractive to natural enemies has potential to improve pest control without sacrificing cropland, which is of great significance in CBC. Further field studies are necessary to evaluate the efficacy of synthetic attractants based on buckwheat volatiles for A. lucorum control by P. spretus.
Insect olfaction varies with compound type and concentration [39] and the species and sex of organism [40]. In general, EAG amplitude increased with ascending concentrations for the five compounds within a certain range. The EAG response relative value of male P. spretus was the maximum when the concentrations of Z3HA, 4-methylanisole, 4-oxoisophorone, p-methylphenol and 2-ethylhexyl salicylate were 10 mg/mL, and that of females reached the maximum when the concentrations of Z3HA, 4-methylanisole, 4-oxoisophorone and p-methylphenol were 10 mg/mL while 2-ethylhexyl salicylate was at 0.01 mg/mL. In summary, the EAG response of P. spretus could be affected by the type and concentration of volatile compounds, which is an important guideline for selecting suitable compounds and concentrations to make attractants for biological control of A. lucorum in the field.
Insects mainly recognize external chemical signals through olfaction; their olfactory receptors are mainly distributed on antennae where chemical communication among the tritrophic levels is mainly accomplished [41]. This process also requires the involvement of multiple proteins, such as odorant-binding proteins (OBPs), chemosensory proteins (CSPs), odorant receptors (ORs) and ionotropic receptors (IRs) [42]. Competitive binding experiments found that Z3HA had high binding affinity to AgosOBP8, which can participate in olfactory and taste recognition at the same time, playing a physiological role in regulating behavior of Aphis gossypii [43]. In addition, SlituOR12, which could affect the detection of host location and oviposition site of moth females, with expressed oocytes having extremely high sensitivity to Z3HA [41]. It could be inferred that Z3HA plays an important role in the host recognition process of insects and affects their searching behavior.
Previous laboratory studies revealed that feeding on buckwheat flowers could significantly increase the daily parasitism rate of P. spretus by 39.8% compared with water [25]. In addition, Li et al. [28] evaluated the parasitism rate of A. lucorum in field plots of 13 plant species and found that the parasitism rate among buckwheat flowers was the highest, i.e., 2.9× that of cotton. Additionally, we here demonstrated that volatile compounds of buckwheat flowers could also significantly increase the parasitism rate of P. spretus under field conditions. Our results provide insight on the role of buckwheat floral VOCs in P. spretus behavior and enable future studies toward the development of a CBC program for A. lucorum.
4. Materials and Methods
4.1. Biological and Chemical Materials
4.1.1. Insects
Mirid bugs. Adults and nymphs of A. lucorum were obtained from Langfang Experimental Station, Chinese Academy of Agricultural Sciences (CAAS), Hebei Province, China (116°36′7″ E, 39°30′31″ N). These insects were continuously reared in 20 × 10 × 6 cm transparent plastic boxes at 25 ± 1 °C, 60 ± 5% RH and 14:10 h (L:D) photoperiod. Adults of A. lucorum were provided with fresh green bean pods (Phaseolus vulgaris L.), rinsed with sodium hypochlorite (NaClO) solution (0.5%), as both food and an oviposition substrate, whereas nymphs were reared on kernels of organic corn (Zea mays L.) [15,44].
Parasitoids. P. spretus adults were obtained from a laboratory colony at Langfang Experimental Station, CAAS, where this parasitoid had been continuously reared for >1 year in plexiglass rearing cages (30 × 30 × 25 cm) at 25 ± 1 °C, 60 ± 5% RH and 14:10 h (L:D) photoperiod, using 3rd to 5th instar nymphs of A. lucorum as hosts while corn kernels continued to be offered for consumption [20].
4.1.2. Plants
Buckwheat seeds were sown in the growth medium mixed with peat soil: vermiculite: medium loam in the ratio of 6: 1:1 (by volume) in a greenhouse at Langfang Experimental Station, CAAS, under the following conditions: 26 ± 1 °C, 60 ± 10% RH and 14:10 h (L:D) photoperiod. Cotton (CCRI49) seeds were obtained from the Institute of Cotton Research of CAAS and sown in a field at Langfang Experimental Station of CAAS in May, and no fertilizers, pesticides or herbicides were used on the plants.
4.2. Behavioral Bioassays with Buckwheat Flowers
Referring to Yu et al. [14] and Xiu et al. [45], a Y-type olfactory device (inner diameter: 2 cm; length of main and selector arms: 15 cm; angle between selector arms: 60°) was used to evaluate the behavioral responses of P. spretus adults to buckwheat flowers. A QC-3 atmospheric sampler (Beijing Municipal Institute of Labor Protection, Beijing, China) was used as the airflow power source to connect activated Z3HArcoal, a distilled water humidification device, a gas flow control meter, a glass odor source vessel and the Y-tube test arena with Teflon tubes and to seal the connections with parafilm (Figure 6).
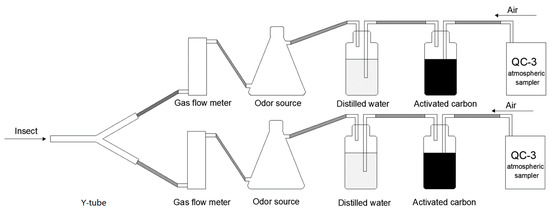
Figure 6.
Schematic of Y-tube olfactometer.
Two pots of full-bloom buckwheat flowers were selected to conduct our olfactometer bioassays at 25 ± 1 °C from 900 h to 1700 h. Before the test, the buckwheat plants were rinsed with ultra-pure water; the soil part was wrapped tightly with tin foil; and care was taken to avoid plant damage. An intact buckwheat plant was then placed into one odor source vessel, and the other odor source was an empty air control.
One active, unmated and healthy 2-day-old P. spretus adult (n = 60 males and 60 females) was introduced to the initial test chamber. When parasitoid wasps crossed 1/3rd of either test arm within 5 min and remained there for more than 10 s, it was considered to be a selection; if wasps did not respond as such they were considered to be unresponsive and discarded. After testing five parasitic wasps, the two arms of the Y-tube were reversed, and after testing 10 wasps a clean Y-tube was used. At the end of the test, the Y-tube, odor source vessel and Teflon tubes were washed with 95% ethanol, soaked and rinsed with distilled water and air-dried at room temperature.
4.3. Collection and Analysis of Buckwheat Volatiles
Buckwheat flower volatiles were collected by SPME. Prior to the start of the experiment, the SPME device was activated in the GC-MS injection port for 30 min (230 °C). Latex gloves were worn to pick buckwheat flowers from plants, which accounted for about 1/3rd of the transparent glass sample bottle (10 mL). The sample bottle was sealed and placed horizontally on the lab bench. SPME collections were then made for 4 h with a fiber needle coated with quartz (INNOTEG, Beijing, China), and a total of three biological replicates were set up.
GC-MS (GC: Agilent 7890A, equipped with a DB-WAX chromatographic column [30 m × 0.25 mm × 0.25 µm]; MS: Agilent 5975C) was used to identify and analyze the major substances of buckwheat flowers. GC and MS working conditions were similar parameters as those in previous studies [45]. The injector temperature for GC analysis was 230 °C; the oven temperature was kept at 50 °C for 1 min, then raised by 5 °C/min to 180 °C for 2 min, then increased by 10 °C/min to 230 °C and held for 2 min. Helium was the carrier gas, at an average flow rate of 1 mL/min. The ion source temperature was 230 °C. The volatile samples were first identified automatically by NIST 14, and then their retention times and external standard method were, respectively, used for compound identification and qualitative analyses. Some 1 mL five standard compounds at different concentrations (0.001, 0.002, 0.05, 0.1 and 0.2 mg/mL) which were dissolved in hexane, respectively, were used to establish an external standard curve, whose y-axis was the peak areas of mass chromatograms, and x-axis was the substance content (mg/L). The emission amount of buckwheat flower volatiles can be obtained by substituting the peak area of the corresponding compound.
4.4. EAG Recordings
Electroantennographic tests were performed with an EAG detector system (Syntech Ltd., Hilversum, The Netherlands) with similar parameters as those in Liu [46] and Zhang et al. [47], including the control recording of electrodes, amplifier and signal electronic processing, signal display and recording system, and stimulus application system.
Active 2-day-old P. spretus adults (n = 3 males and 3 females for each treatment) with intact antennae were selected to assess their antennal responses to the volatiles of buckwheat flowers. One antenna was cut off along at the base with a scalpel, and the tip of the antenna was also removed before the antenna was placed between the reference electrode and recording electrode. charcoal filtered, humidified air (0.4 L/min) was blown over the antenna for a few minutes to allow the baseline to stabilize. Then, the electrodes were inserted into the EAG probe for signal acquisition, amplification and conversion analysis. The order of the test solutions was: control (paraffin oil), five standard compounds at different concentrations (0.001, 0.01, 0.1, 1 and 10 µg/µL) which were dissolved in paraffin oil and the standard reference compound (10 µg/µL cis-3-hexen-1-ol) [47]. Ten µL of each test solution was placed on a rectangular filter paper strip (0.5 cm × 4 cm) and placed in a glass Pasteur pipette for presentation to the antenna. The duration of each stimulation was 0.5 s, and the interval between two stimuli was 30 s during which time clean humidified air was blown over the antenna. Each antenna was stimulated seven times.
4.5. Behavioral Bioassays with Synthetic Compounds
The Y-tube olfactometer was used to evaluate behavioral responses of 2-day-old active P. spretus adults (n = 40 males and 40 females for each treatment) to major components of buckwheat flower volatiles. One side of the test tube was mineral oil as control, while the other side was respectively placed with (1) 10 mg/mL Z3HA, (2) 10 mg/mL 4-methylanisole, (3) 10 mg/mL 4-oxoisophorone, (4) 10 mg/mL p-methylphenol and (5) 10 mg/mL 2-ethylhexyl salicylate. Ten μL of each test solution was pipetted onto the filter paper strip and put into the odor source bottle, following protocols as above.
4.6. Field Trials
Field parasitism rate trials were conducted in August 2022 in cotton fields (0.13 ha) and vineyards (0.11 ha), respectively, located at Langfang Experimental Station, CAAS. During the experimental period, no pesticides were applied. Prior to each trial (replicate), bamboo poles with barrel traps (Insect collector: upper diam: 16.5 cm; lower diam: 13.0 cm; height: 12.5 cm; Pherobio Technology Co., Ltd., Beijing, China) were randomly placed in each test plot at 10 m distances, with each trap 20 cm from the top of the plants. A dilution of 200 µL of five volatiles (Z3HA, 4-methylanisole, 4-oxoisophorone, p-methylphenol, 2-ethylhexyl salicylate) at a concentration of 10 mg/mL was separately placed in a rubber septum (length: 1.84 cm; upper diameter: 0.89 cm; lower diameter: 0.46 cm) in each trap, and mineral oil was used as the control. Each barrel trap was filled with 100 2nd–3rd instar A. lucorum nymphs and four beans as food source. Then, a total of 50 pairs of 2-day-old P. spretus adults were released at the center and four corners of each plot. Two days later, A. lucorum nymphs were transferred indoors and fed with corn for 6 d until they were dissected to assess P. spretus parasitism. Five volatiles and one control were placed in each of the six traps in the field to test effects of different volatile compounds on parasitism by the wasps, and 12 trials (replicates) were conducted in cotton fields and vineyards respectively.
4.7. Statistical Analysis
Chi-square tests were used to analyze the Y-tube olfactometer data to detect differences between the pairs of treatments. χ2 and P values were calculated, and non-responsive adults were excluded from the analysis. The emission amounts, relative EAG response of each volatile and field parasitism rates were compared using one-way ANOVA, followed by Duncan’s new multiple range tests. Chi-square tests and one-way ANOVAs were conducted using SPSS 25.0.
5. Conclusions
From the perspective of chemical ecology, our study found that buckwheat flowers and their five active volatile components (Z3HA, 4-methylanisole, 4-oxoisophorone, p-methylphenol and 2-ethylhexyl salicylate) had a significant attraction on P. spretus adults through the combined analysis of Y-type insect olfactometer, SPME, GC-MS and EAG. In addition, this study clarified the EAG response of buckwheat flowers on P. spretus and the key role of buckwheat flowers in the selection behavior of P. spretus; the antennal potential response of males and females was especially the strongest under 10 mg/mL of 4-Oxoisophorone. Moreover, field experiments revealed the positive effect of five volatiles on the parasitism of P. spretus, which could increase the parasitism rate by ca. 40%. This study illustrated the selective behavior of P. spretus on buckwheat flowers and its chemical recognition mechanism for buckwheat flower volatiles, which provided a theoretical basis for the development of P. spretus attractants and CBC of A. lucorum. Further work is necessary to evaluate the potential benefits of buckwheat volatiles to feral P. spretus and to determine the relative attractiveness of volatiles from buckwheat flowers versus synthetic volatiles on P. spretus attraction and parasitism.
Author Contributions
S.X.: data curation and analysis, investigation, writing—original draft; T.Z.: methodology, supervision, validation; L.W.III: conceptualization, writing—review and editing; Y.Y.: conceptualization, writing—review and editing; Y.L.: conceptualization, data curation and analysis, funding acquisition, methodology, project administration, writing—review and editing. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the National Key R&D Program of China (2022YFD1400300), China Agriculture Research System of MOF and MARA (CARS-15-19), and the funding from MARA-CABI Joint Laboratory for Bio-safety.
Data Availability Statement
Not applicable.
Acknowledgments
We thank Yu Gao from the Institute of Plant Protection, Chinese Academy of Agricultural Sciences, for her help in arranging field trials and Bing Liu from the Institute of Plant Protection, Chinese Academy of Agricultural Sciences, for assistance with field trials design.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript or in the decision to publish the results.
References
- Orre Gordon, G.U.S.; Wratten, S.D.; Jonsson, M.; Simpson, M.; Hale, R. ‘Attract and reward’: Combining a herbivore-induced plant volatile with floral resource supplementation-Multitrophic level effects. Biol. Control 2013, 64, 106–115. [Google Scholar] [CrossRef]
- Cheng, Y. Study on the effect of urban space insectary plants on attracting natural enemy insects. Master’s Thesis, Beijing Forestry University, Beijing, China, 2020. [Google Scholar]
- Jonsson, M.; Wratten, S.D.; Landis, D.A.; Gurr, G.M. Recent advances in conservation biological control of arthropods by arthropods. Biol. Control 2008, 45, 172–175. [Google Scholar] [CrossRef]
- Begg, G.S.; Cook, S.M.; Dye, R.; Ferrante, M.; Franck, P.; Lavigne, C.; Lövei, G.L.; Mansion-Vaquie, A.; Pell, J.K.; Petit, S.; et al. A functional overview of conservation biological control. Crop Prot. 2017, 97, 145–158. [Google Scholar] [CrossRef]
- Sheng, Z.Y.; Li, W.Z.; Yuan, G.H. Advances in studies on the relationship between plant odor diversity and insects. Chin. J. Appl. Entomol. 2019, 56, 652–661. [Google Scholar] [CrossRef]
- Xu, H.; Turlings, T.C.J. Plant volatiles as mate-finding cues for insects. Trends Plant Sci. 2018, 23, 100–111. [Google Scholar] [CrossRef]
- Kafle, B.D.; Morawo, T.; Fadamiro, H. Host-induced plant volatiles mediate ability of the parasitoid Microplitis croceipes to discriminate between unparasitized and parasitized Heliothis virescens larvae and avoid superparasitism. J. Chem. Ecol. 2020, 46, 967–977. [Google Scholar] [CrossRef]
- Gontijo, L.; Cascone, P.; Giorgini, M.; Michelozzi, M.; Rodrigues, H.S.; Spiezia, G.; Iodice, L.; Guerrieri, E. Relative importance of host and plant semiochemicals in the foraging behavior of Trichogramma achaeae, an egg parasitoid of Tuta absoluta. J. Pest Sci. 2019, 92, 1479–1488. [Google Scholar] [CrossRef]
- Liu, F.; Lou, Y.G.; Cheng, J.A. Herbivory insect induced plant volatiles: Evolutionary products of plant-herbivore-natural enemy interactions. Entomol. Knowl. 2003, 40, 481–486. [Google Scholar]
- Su, J.W.; Cai, Z.P.; Qiao, F.; Miao, L.; Yin, S.Q.; Zheng, P.Q. Numbers of natural enemies of corn pests attracted by plant volatile lures. Chin. J. Appl. Entomol. 2020, 57, 196–205. [Google Scholar] [CrossRef]
- Xue, J.L.; He, J.; Xie, Y.P. Attractive effect of plant volatiles on Harmonia axyridis (Pallas). Chin. J. Appl. Environ. Biol. 2008, 14, 494–498. [Google Scholar]
- Liu, Y.; Guo, G.X.; Chen, J.L.; Ni, H.X. Behavioral and electrophysiological responses of four predatory insect species to semiochemicals of wheat. Acta Entomol. Sin. 2005, 48, 161–165. [Google Scholar]
- Quiroz, A.; Pettersson, J.; Pickett, J.A.; Wadhams, L.J.; Niemeyer, H.M. Semiochemicals mediating spacing behavior of bird cherry-oat aphid, Rhopalosiphum padi feeding on cereals. J. Chem. Ecol. 1997, 23, 2599–2607. [Google Scholar] [CrossRef]
- Yu, H.L.; Zhang, Y.J.; Wyckhuys, K.A.; Wu, K.M.; Gao, X.W.; Guo, Y.Y. Electrophysiological and behavioral responses of Microplitis mediator (Hymenoptera: Braconidae) to caterpillar-induced volatiles from cotton. Environ. Entomol. 2010, 39, 600–609. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.H.; Wu, K.M.; Cai, X.M.; Liu, Y.Q. A rearing method for mirids using the green bean, Phaseolus vulgaris in the laboratory. Acta Phytophylacica Sin. 2008, 35, 215–219. [Google Scholar]
- Chen, Z.C.; Su, L.; Ge, F.; Su, J.W. Electroantennogram responses of the green leaf bug, Lygus lucorum Meyer-Dür (Hemiptera: Miridae), to sex pheromone analogs and plant volatiles. Acta Entomol. Sin. 2010, 53, 47–54. [Google Scholar]
- Pan, H.S.; Lu, Y.H.; Xiu, C.L.; Geng, H.H.; Cai, X.M.; Sun, X.L.; Zhang, Y.J.; Williams, L.I.; Wyckhuys, K.A.G.; Wu, K.M. Volatile fragrances associated with flowers mediate host plant alternation of a polyphagous mirid bug. Sci. Rep. 2015, 5, 14805. [Google Scholar] [CrossRef]
- Pan, H.S.; Xiu, C.L.; Livy, W.I.; Lu, Y.H. Plant volatiles modulate seasonal dynamics between hosts of the polyphagous mirid bug Apolygus lucorum. J. Chem. Ecol. 2021, 47, 87–98. [Google Scholar] [CrossRef]
- Xiu, C.L.; Pan, H.S.; Liu, B.; Luo, Z.X.; Williams, L., III; Yang, Y.Z.; Lu, Y.H. Perception of and behavioral responses to host plant volatiles for three Adelphocoris species. J. Chem. Ecol. 2019, 45, 779–788. [Google Scholar] [CrossRef]
- Luo, S.P.; Zhang, F.; Wu, K.M. Effect of temperature on the reproductive biology of Peristenus spretus (Hymenoptera: Braconidae), a biological control agent of the plant bug Apolygus lucorum (Hemiptera: Miridae). Biocontrol Sci. Technol. 2015, 25, 1410–1425. [Google Scholar] [CrossRef]
- Luo, S.P.; Zhang, F.; Wu, K.M. Interspecific competition between Peristenus spretus and Peristenus relictus (Hymenoptera: Braconidae), larval parasitoids of Apolygus lucorum (Hemiptera: Miridae). Biol. Control 2018, 117, 115–122. [Google Scholar] [CrossRef]
- Luo, S.P.; Lu, Y.H.; Men, X.Y.; Zhang, F.; Wu, K.M. Parasitism of Apolygus lucorum by Peristenus spretus at different release levels in jujube orchards. Chin. J. Biol. Control 2016, 32, 698–702. [Google Scholar] [CrossRef]
- Colazza, S.; Peri, E.; Cusumano, A. Chemical ecology of floral resources in conservation biological control. Ann. Rev. Entomol. 2022, 68, 13–29. [Google Scholar] [CrossRef] [PubMed]
- Dai, W.J.; Pan, H.S.; Xiu, C.L.; Luo, S.P.; Yang, Y.Z.; Lu, Y.H. Behavioral selection of endoparasitoid Peristenus spretus to green mirid bug Apolygus lucorum and its damaged host plants. J. Plant Prot. 2018, 45, 194–200. [Google Scholar] [CrossRef]
- Xia, S.K.; Luo, S.P.; Li, J.Q.; Yang, Q.; Dai, C.; Yang, Y.; Lu, Y. Fructose and glucose in buckwheat nectar enhance Peristenus spretus (Hymenoptera: Braconidae) survival and parasitism of the mirid Apolygus lucorum. Biol. Control 2021, 161, 104710. [Google Scholar] [CrossRef]
- Russell, M. A meta-analysis of physiological and behavioral responses of parasitoid wasps to flowers of individual plant species. Biol. Control 2015, 82, 96–103. [Google Scholar] [CrossRef]
- Shields, M.W.; Johnson, A.C.; Pandey, S.; Cullen, R.; González- Chang, M.; Wratten, S.D.; Gurr, G.M. History, current situation and challenges for conservation biological control. Biol. Control 2019, 131, 25–35. [Google Scholar] [CrossRef]
- Li, J.H.; Liu, B.; Pan, H.S.; Luo, S.P.; Wyckhuys, K.A.G.; Yuan, H.; Lu, Y. Buckwheat strip crops increase parasitism of Apolygus lucorum in cotton. BioControl 2019, 64, 645–654. [Google Scholar] [CrossRef]
- Mohammed, K.; Agarwal, M.; Du, X.B.; Newman, J.; Ren, Y.L. Behavioural responses of the parasitoid Aphytis melinus to volatiles organic compounds (VOCs) from Aonidiella aurantii on its host fruit Tahitian lime fruit Citrus latifolia. Biol. Control 2019, 133, 103–109. [Google Scholar] [CrossRef]
- Li, M.Y.; Xia, S.K.; Zhang, T.; Williams, L.I.; Xiao, H.J.; Lu, Y.H. Volatiles from cotton plants infested by Agrotis segetum (Lep.: Noctuidae) attract the larval parasitoid Microplitis mediator (Hym.: Braconidae). Plants 2022, 11, 863. [Google Scholar] [CrossRef]
- Effah, E.; Holopainen, J.K.; McCormick, A.C. Potential roles of volatile organic compounds in plant competition. Perspect Plant Ecol. Evol. Syst. 2019, 38, 58–63. [Google Scholar] [CrossRef]
- Cai, H.H.; Zhang, T.; Su, Y.H.; Wang, Z.Y.; Zhang, X.F.; Wang, S.S.; Liu, Y.Q. Influence of trap color, type, and placement on capture efficacy for Protaetia brevitarsis (Coleoptera: Scarabaeidae). J. Econ. Entom. 2021, 114, 225–230. [Google Scholar] [CrossRef] [PubMed]
- Harvey, R.M.; Bateman, A.P.; Jain, S.; Li, Y.J.; Martin, S.; Petrucci, G.A. Optical properties of secondary organic aerosol from cis-3-hexenol and cis-3-hexenyl acetate: Effect of chemical composition, humidity, and phase. Environ. Sci. Technol. 2016, 50, 4997–5006. [Google Scholar] [CrossRef] [PubMed]
- Pragadheesh, V.S.; Yadav, A.; Chanotiya, C.S.; Rout, P.K.; Uniyal, G.C. Monitoring the emission of volatile organic compounds from flowers of Jasminum sambac using solid-phase micro-extraction fibers and gas chromatography with mass spectrometry detection. Nat. Prod. Commun. 2011, 6, 1333–1338. [Google Scholar] [CrossRef] [PubMed]
- Robertson, G.W.; Griffliths, D.W.; Woodford, J.A.T.; Birch, A.N.E. Changes in the chemical composition of volatiles released by the flowers and fruits of the red raspberry (Rubus idaeus) cultivar glen prosen. Phytochemistry 1995, 38, 1175–1179. [Google Scholar] [CrossRef]
- El-Sayed, A.M.; Mitchell, V.J.; McLaren, G.F.; Manning, L.M.; Bunn, B.; Suckling, D.M. Attraction of New Zealand flower thrips, Thrips obscuratus, to cis-jasmone, a volatile identified from Japanese honeysuckle flowers. J. Chem. Ecol. 2009, 35, 656–663. [Google Scholar] [CrossRef] [PubMed]
- Mookherjee, B.D.; Trenkle, R.W.; Wilson, R.A. Live vs. dead. Part II. A comparative analysis of the headspace volatiles of some important fragrance and flavor raw materials. J. Essent. Oil Res. 1989, 1, 85–90. [Google Scholar] [CrossRef]
- Sun, Y.L.; Dong, J.F.; Huang, L.Q.; Wang, C.Z. The cotton bollworm endoparasitoid Campoletis chlorideae is attracted by cis-jasmone or cis-3-hexenyl acetate but not by their mixtures. Arthropod-Plant Interact. 2020, 14, 169–179. [Google Scholar] [CrossRef]
- Wang, P.; Zhang, L. Olfaction in the process of herbivorous insect food selection. J. Environ. Entomol. 2021, 43, 633–641. [Google Scholar] [CrossRef]
- He, Y.Y.; Wang, K.; Zhang, Y.J.; Wu, Q.J.; Wang, S.L. Research progress of olfactory protein of parasitoid wasps. J. Environ. Entomol. 2019, 41, 1232–1243. [Google Scholar]
- Zhou, Z.; Wang, M.Q.; Hu, Y.; Chen, H.Y. Morphological structure of the antennal sensilla of Nesidiocoris tenuis observed with a scanning electron microsope. Chin. J. Appl. Entomol. 2012, 49, 631–635+635–636. [Google Scholar] [CrossRef]
- Leal, W.S.; Choo, Y.M.; Xu, P.; da Silva, C.S.; Ueira-Vieira, C. Differential expression of olfactory genes in the southern house mosquito and insights into unique odorant receptor gene isoforms. Proc. Natl. Acad. Sci. USA 2013, 110, 18704–18709. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.K.; Zhang, S.; Luo, J.Y.; Wang, C.Y.; Lü, L.-M.; Zhang, L.J.; Zhu, X.Z.; Wang, L.; Cui, J.J. Molecular characterization and ligand-binding properties of six odorant-binding proteins (OBPs) from Aphis gossypii. J. Asia-Pac. Entomol. 2018, 21, 914–925. [Google Scholar] [CrossRef]
- Pan, H.S.; Liu, B.; Lu, Y.H. Host-plant switching promotes the population growth of Apolygus lucorum: Implications for laboratory rearing. Bull. Entomol. Res. 2018, 109, 309–315. [Google Scholar] [CrossRef]
- Xiu, C.L.; Dai, W.J.; Pan, H.S.; Zhang, W.; Luo, S.P.; Wyckhuys, K.A.G.; Yang, Y.Z.; Lu, Y.H. Herbivore-induced plant volatiles enhance field-level parasitism of the mirid bug Apolygus lucorum. Biol. Control 2019, 135, 41–47. [Google Scholar] [CrossRef]
- Liu, H.N.; Xiu, C.L.; Zhang, T.; Lu, Y.H. Odor perception in the cotton bollworm, Helicoverpa armigera, exposed to Juglans regia, a marginal host plant. J. Chem. Ecol. 2022, 48, 618–627. [Google Scholar] [CrossRef]
- Zhang, X.F.; Liu, H.M.; Zhang, A.H.; Fang, C.; Zhang, T. Pear leaf volatiles and their electroantennogram responses to Illiberis pruni. J. Anhui Agric. Sci. 2017, 45, 145–148. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).