
Probiotics, Proline and Calcium Induced Protective Responses of Triticum aestivum under Drought Stress
 ,
,  ,
,  , , and
, , and
Abstract
1. Introduction
2. Results
2.1. Effect of Stress-Protecting Compounds on RWC of Wheat Seedlings Exposed to Prolonged Drought
2.2. Impact of Stress-Protecting Compounds on Morphometric Parameters of Wheat Seedlings Exposed to Prolonged Drought Stress
2.3. Impact of Stress-Protecting Compounds on Accumulation of Photosynthetic Pigments of Wheat Seedlings Exposed to Prolonged Drought Stress
2.4. Impact of Stress-Protecting Compounds on Ethylene Emission of Wheat Seedlings Exposed to Prolonged Drought
2.5. Effect of Stress-Protecting Compounds on Biochemical Responses of Wheat Plant Exposed to Prolonged Drought
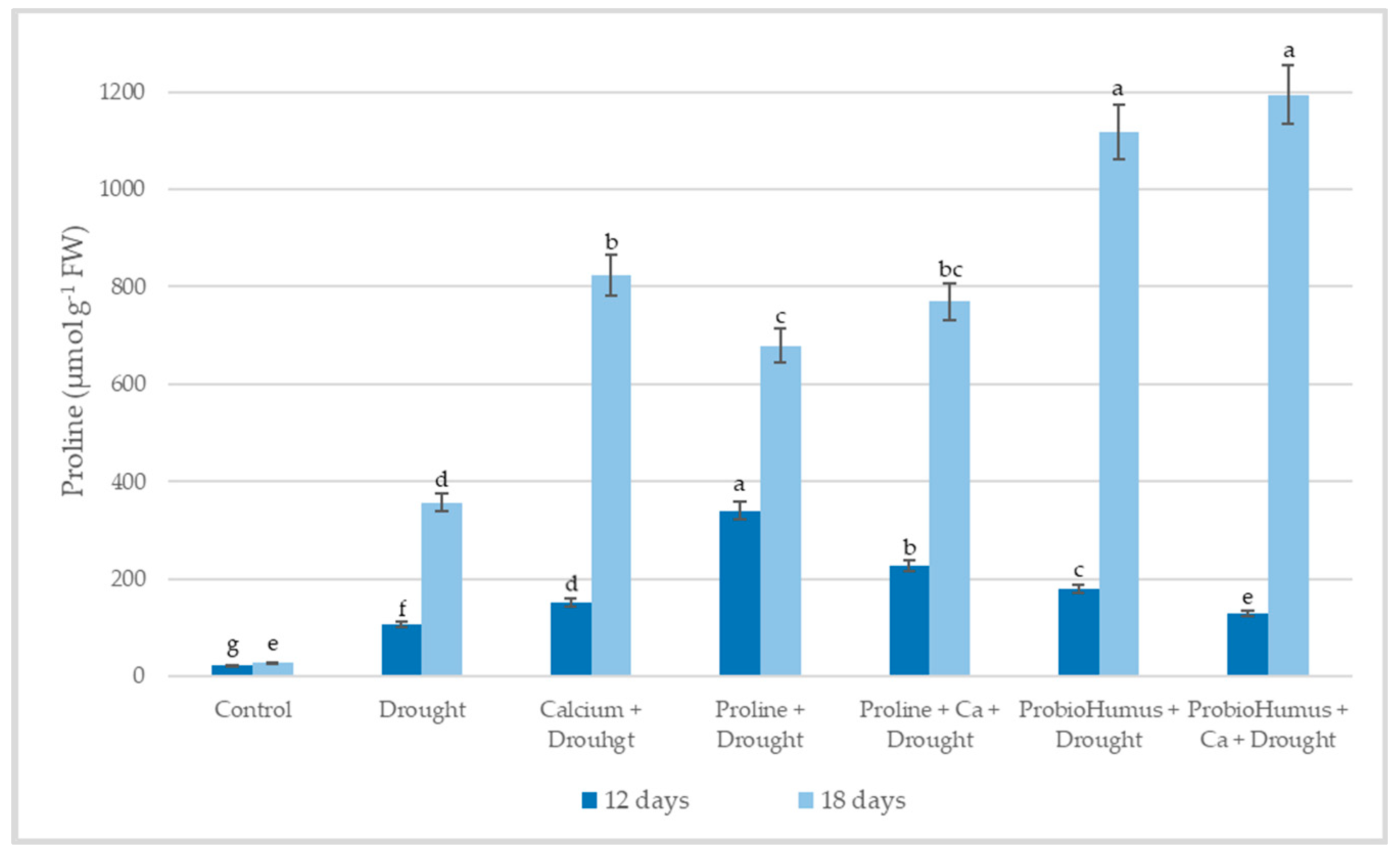
2.5.1. Free Proline
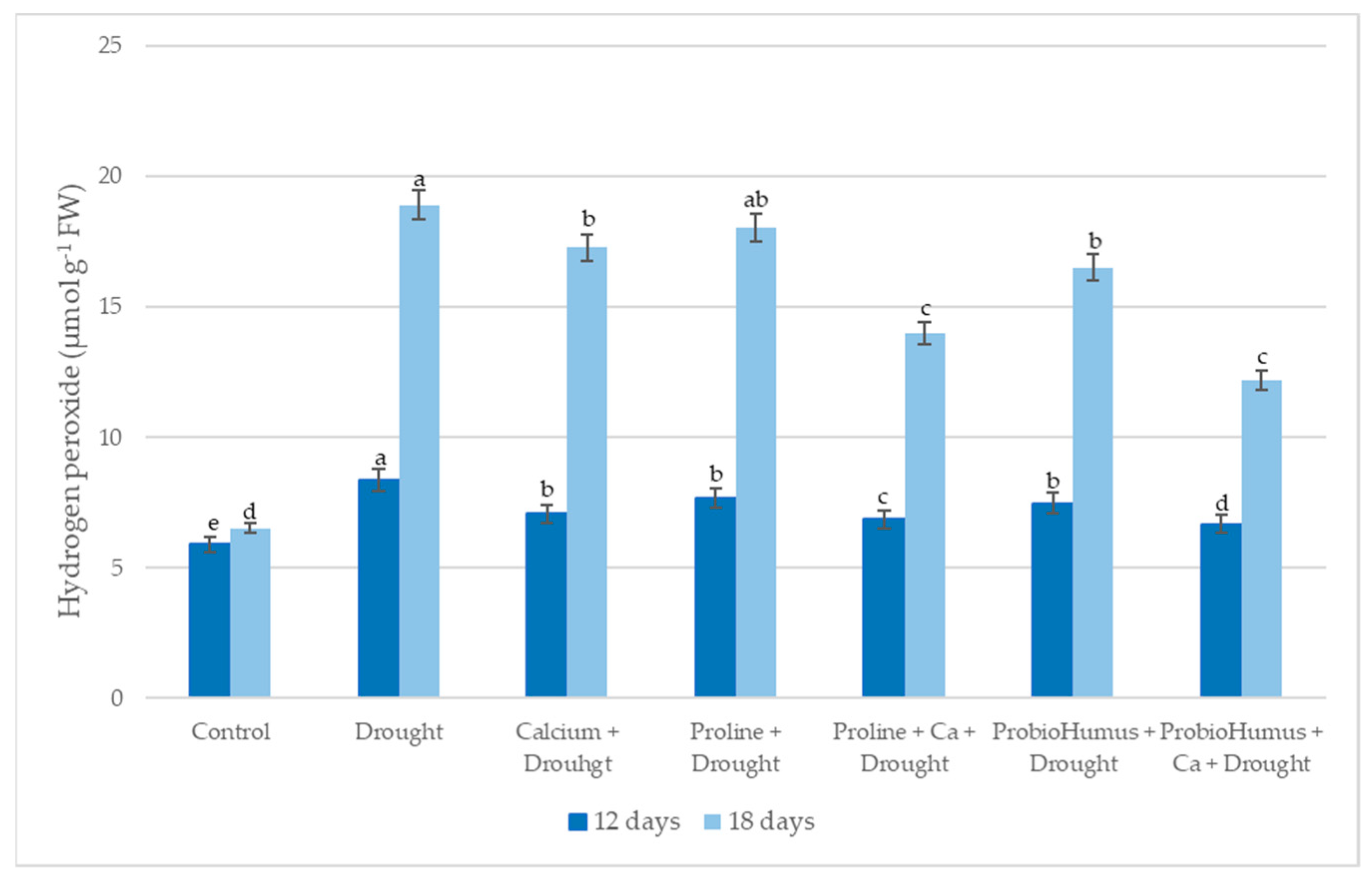
2.5.2. Hydrogen Peroxide (H2O2)
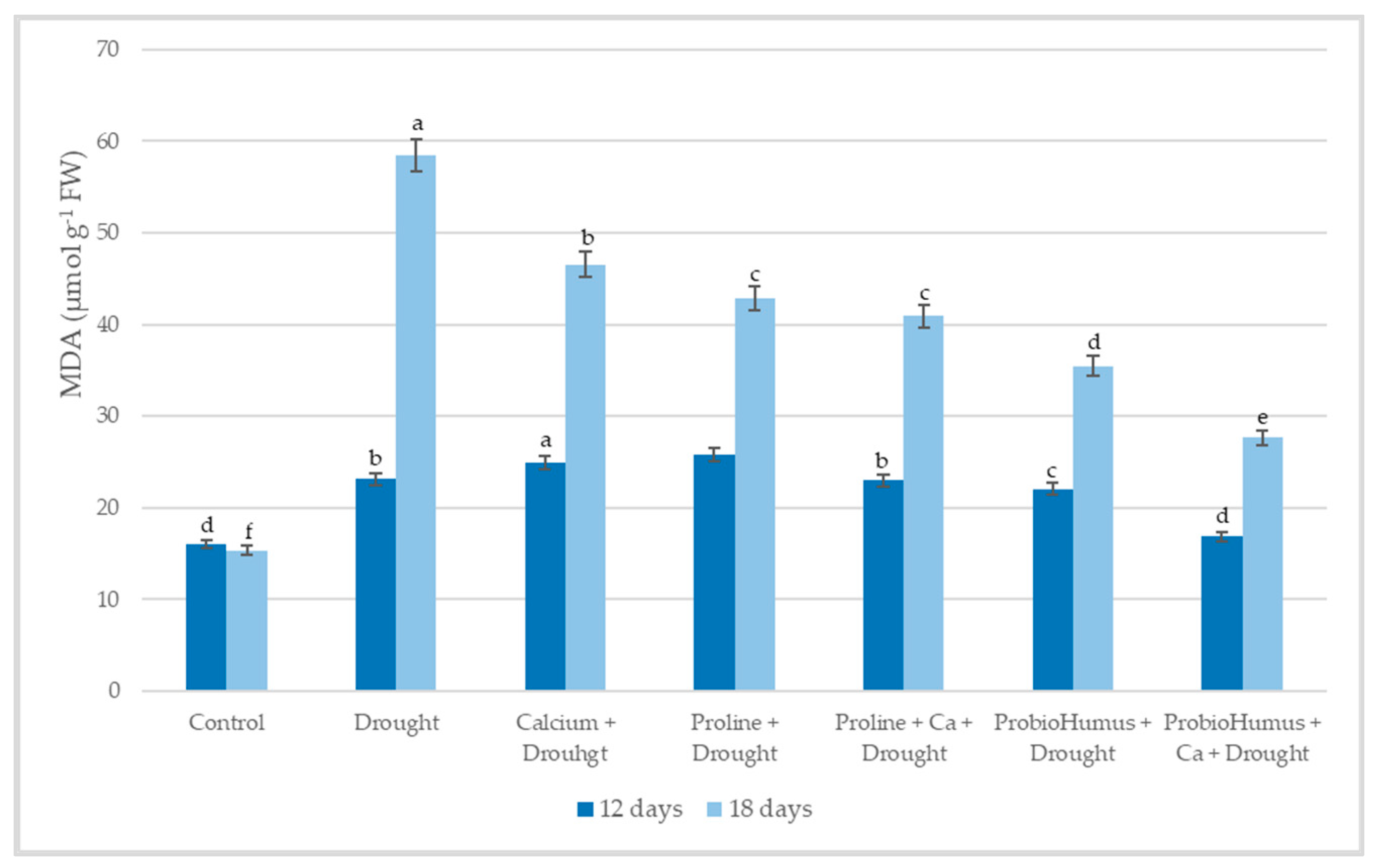
2.5.3. Malondialdehyde (MDA)
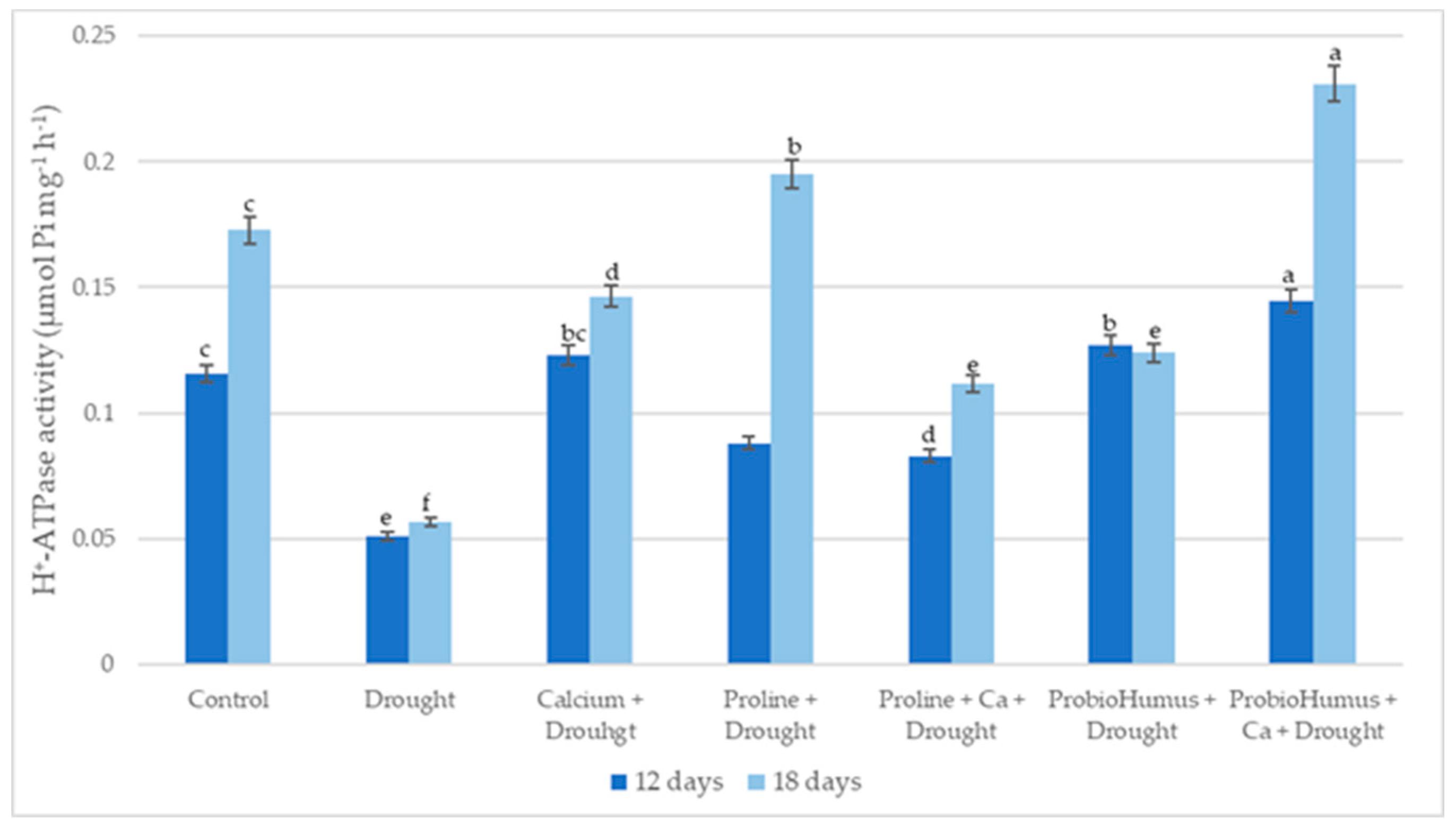
2.5.4. PM ATPase Activity
2.6. Effect of Stress-Protecting Compounds on Late Embryogenesis Abundant (lea) Genes Expression Levels of Wheat Plant Exposed to Prolonged Drought
3. Discussion
4. Materials and Methods
4.1. Plant Material and Growth Conditions
Treatments
- (a)
- Calcium was added to the soil in the form of CaCO3 (MKDS) (hereafter referred to as Ca) at a rate of 3.71 g per pot, based on 70 g m−2.
- (b)
- ProbioHumus—probiotic concentrate (Baltic Probiotics, Rucavas pagasts, Latvia) was used for seed priming 2 µL/g and diluted with water 1:100 was used for seedling spraying 10 mL per pot at the 3–4 leaf stage (BBCH-scale 3–4) [71]. Composition of microorganisms: Bacillus subtilis (103 CFU/mL), Saccharomyces cerevisiae, Bifidobacterium animalis, B. bifidum, B. longum (104 CFU/mL), Lactobacillus diacetylactis, L. casei, L. delbrueckii, L. plantarum, (105 CFU/mL), Lactococcus lactis (102 CFU/mL), Streptococcus thermophilus, Rhodopseudomonas palustris and R. sphaeroides (104 CFU/mL).
- (c)
- L-proline 1 mM aqueous solution (Roth) was used for seedling spraying 10 mL per pot at the 3–4 leaf stage (BBCH-scale 3–4) [71].
4.2. Experimental Design
4.3. Sampling
4.4. Morphometrical Measurements
4.5. Relative Water Content (RWC)
4.6. Assessment of Biochemical Parameters
4.6.1. Lipid Peroxidation According MDA
4.6.2. H2O2
4.6.3. Proline
4.6.4. Photosynthetic Pigments
4.6.5. Ethylene
4.6.6. PM ATPase Activity Assay
4.7. Molecular Techniques
4.7.1. RNA Extraction and Reverse Transcription
4.7.2. Real-Time Quantitative PCR
4.7.3. Primers
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fahad, S.; Bajwa, A.A.; Nazir, U.; Anjum, S.A.; Farooq, A.; Zohaib, A.; Sadia, S.; Nasim, W.; Adkins, S.; Saud, S.; et al. Crop production under drought and heat stress: Plant responses and management options. Front. Plant Sci. 2017, 8, 1147. [Google Scholar] [CrossRef]
- Camaille, M.; Fabre, N.; Clément, C.; Ait Barka, E. Advances in wheat physiology in response to drought and the role of plant growth promoting rhizobacteria to trigger drought tolerance. Microorganisms 2021, 9, 687. [Google Scholar] [CrossRef]
- Fita, A.; Rodríguez-Burruezo, A.; Boscaiu, M.; Prohens, J.; Vicente, O. Breeding and domesticating crops Adapted to drought and salinity: A new paradigm for increasing food production. Front. Plant Sci. 2015, 6, 978. [Google Scholar] [CrossRef]
- Nuccio, M.L.; Paul, M.J.; Bate, N.J.; Cohn, J.; Cutler, S.R. Where are the drought tolerant crops? An assessment of more than two decades of plant biotechnology effort in crop improvement. Plant Sci. 2018, 273, 110–119. [Google Scholar] [CrossRef] [PubMed]
- Kanwal, S.; Ilyas, N.; Batool, N.; Arshad, M. Amelioration of drought stress in wheat by combined application of PGPR, compost, and mineral fertilizer. J. Plant Nutr. 2017, 40, 1250–1260. [Google Scholar] [CrossRef]
- Liu, J.; Niu, Y.; Zhang, J.; Zhou, Y.; Ma, Z.; Huang, X. Ca2+ channels and Ca2+ signals involved in abiotic stress responses in plant cells: Recent advances. Plant Cell Tissue Organ Cult. 2018, 132, 413–424. [Google Scholar] [CrossRef]
- Rezayian, M.; Niknam, V.; Ebrahimzadeh, H. Different effects of calcium and penconazole on primary and secondary metabolites of Brassica napus under drought. Physiol. Mol. Biol. Plants 2019, 25, 497–509. [Google Scholar] [CrossRef]
- Chen, C.; Xin, K.; Liu, H.; Cheng, J.; Shen, X.; Wang, Y.; Zhang, L. Pantoea alhagi, a novel endophytic bacterium with ability to improve growth and drought tolerance in wheat. Sci. Rep. 2017, 7, 41564. [Google Scholar] [CrossRef] [PubMed]
- Lalarukh, I.; Al-Dhumri, S.A.; Al-Ani, L.K.T.; Hussain, R.; Al Mutairi, K.A.; Mansoora, N.; Amjad, S.F.; Abbas, M.H.H.; Abdelhafez, A.A.; Poczai, P.; et al. A Combined use of rhizobacteria and moringa leaf extract mitigates the adverse effects of drought stress in wheat (Triticum aestivum L.). Front. Microbiol. 2022, 13, 813415. [Google Scholar] [CrossRef] [PubMed]
- Yadav, V.K.; Yadav, R.C.; Choudhary, P.; Sharma, S.K.; Bhagat, N. Mitigation of drought stress in wheat (Triticum aestivum L.) by inoculation of drought tolerant Bacillus paramycoides DT-85 and Bacillus paranthracis DT-97. J. Appl. Biol. Biotechnol. 2022, 10, 59–69. [Google Scholar] [CrossRef]
- Santoyo, G.; Guzmán-Guzmán, P.; Parra-Cota, F.I.; Santos-Villalobos, S.D.L.; Orozco-Mosqueda, M.; Glick, B.R. Plant growth stimulation by microbial consortia. Agronomy 2021, 11, 219. [Google Scholar] [CrossRef]
- Abbas, M.; Abdel-Lattif, H.; Shahba, M. Ameliorative effects of calcium sprays on yield and grain nutritional composition of Maize (Zea mays L.) cultivars under drought stress. Agriculture 2021, 11, 285. [Google Scholar] [CrossRef]
- Mohammed, I.S.; Al-Maliki, S. Interaction between calcium silicate and Pseudomonas Fluorescens and irrigation intervals affecting soil microbial biomass and yield of wheat. J. NVEO 2022, 9, 628–645. [Google Scholar]
- Tuteja, N.; Mahajan, S. Calcium signaling network in plants: An overview. Plant Signal Behav. 2007, 2, 79–85. [Google Scholar] [CrossRef] [PubMed]
- Navarro-León, E.; López-Moreno, F.J.; Fernández, M.A.; Maldonado, J.J.; Yánez, J.; Blasco, B.; Ruiz, J.M. A New calcium vectoring technology: Concentration and distribution of Ca and agronomic efficiency in pepper plants. Agronomy 2022, 12, 410. [Google Scholar] [CrossRef]
- Kurtyka, R.; Małkowski, E.; Kita, A.; Karcz, W. Effect of calcium and cadmium on growth and accumulation of cadmium, calcium, potassium and sodium in maize seedlings. Pol. J. Environ. Stud. 2008, 17, 51–56. [Google Scholar]
- Hosseini, S.A.; Réthoré, E.; Pluchon, S.; Ali, N.; Billiot, B.; Yvin, J.C. Calcium application enhances drought stress tolerance in sugar beet and promotes plant biomass and beetroot sucrose concentration. Int. J. Mol. Sci. 2019, 20, 3777. [Google Scholar] [CrossRef]
- Ozturk, M.; Turkyilmaz, U.B.; García-Caparrós, P.; Khursheed, A.; Gul, A.; Hasanuzzaman, M. Osmoregulation and its actions during the drought stress in plants. Physiol. Plant. 2021, 17, 1321–1335. [Google Scholar] [CrossRef] [PubMed]
- Ali, Q.; Ashraf, M.; Shahbaz, M.; Humera, H.A.F.I.Z.A. Ameliorating effect of foliar applied proline on nutrient uptake in water stressed maize (Zea mays L.) plants. Pak. J. Bot. 2008, 40, 211–219. [Google Scholar]
- Abdelaal, K.A.; Attia, K.A.; Alamery, S.F.; El-Afry, M.M.; Ghazy, A.I.; Tantawy, D.S.; Hafez, Y.M. Exogenous application of proline and salicylic acid can mitigate the injurious impacts of drought stress on barley plants associated with physiological and histological characters. Sustainability 2020, 12, 1736. [Google Scholar] [CrossRef]
- Farhad; Kibria, M.G.; Mian, M.H.; Murata, Y.; Hoque, M.A. Mitigating water stress in wheat by foliar application of proline. Int. J. Expt. Agric. 2015, 5, 8–14. [Google Scholar]
- Farooq, M.; Nawaz, A.; Chaudhry, M.A.M.; Indrasti, R.; Rehman, A. Improving resistance against terminal drought in bread wheat by exogenous application of proline and gamma-aminobutyric acid. J. Agron. Crop Sci. 2017, 9, 464–472. [Google Scholar] [CrossRef]
- Zheng, J.; Su, H.; Lin, R.; Zhang, H.; Xia, K.; Jian, S.; Zhang, M. Isolation and characterization of an atypical LEA gene (IpLEA) from Ipomoea pes-caprae conferring salt/drought and oxidative stress tolerance. Sci. Rep. 2019, 9, 14838. [Google Scholar] [CrossRef] [PubMed]
- Hsiao, T.C. Plant responses to water stress. Ann. Rev. Plant Physiol. 1973, 24, 519–570. [Google Scholar] [CrossRef]
- Sultan, M.A.R.F.; Hui, L.; Yang, L.J.; Xian, Z.H. Assessment of drought tolerance of some Triticum L. species through physiological indices. Czech J. Genet. Plant Breed. 2012, 48, 178–184. [Google Scholar] [CrossRef]
- Yang, H.; Zhao, L.; Zhao, S.; Wang, J.; Shi, H. Biochemical and transcriptomic analyses of drought stress responses of LY1306 tobacco strain. Sci Rep. 2017, 7, 17442. [Google Scholar] [CrossRef] [PubMed]
- Qayyum, A.; Al Ayoubi, S.; Sher, A.; Bibi, Y.; Ahmad, S.; Shen, Z.; Jenks, M.A. Improvement in drought tolerance in bread wheat is related to an improvement in osmolyte production, antioxidant enzyme activities, and gaseous exchange. Saudi J. Biol. Sci. 2021, 28, 5238–5249. [Google Scholar] [CrossRef]
- Jeyasri, R.; Muthuramalingam, P.; Satish, L.; Pandian, S.K.; Chen, J.-T.; Ahmar, S.; Wang, X.; Mora-Poblete, F.; Ramesh, M. An overview of abiotic stress in cereal crops: Negative impacts, regulation, biotechnology and integrated omics. Plants 2021, 10, 1472. [Google Scholar] [CrossRef]
- Farooq, M.; Wahid, A.; Kobayashi, N.; Fujita, D.; Basra, S.M.A. Plant drought stress: Effects, mechanisms and management. Agron. Sustain. Dev. 2009, 29, 185–212. [Google Scholar] [CrossRef]
- Khushboo, B.K.; Singh, P.; Raina, M.; Sharma, V.; Kumar, D. Exogenous application of calcium chloride in wheat genotypes alleviates negative effect of drought stress by modulating antioxidant machinery and enhanced osmolyte accumulation. In Vitro Cell. Dev. Biol. Plant. 2018, 54, 495–507. [Google Scholar] [CrossRef]
- Hayat, S.; Hayat, Q.; Alyemeni, M.N.; Wani, A.S.; Pichtel, J.; Ahmad, A. Role of proline under changing environments: A review. Plant Signal Behav. 2012, 7, 1456–1466. [Google Scholar] [CrossRef] [PubMed]
- Weng, M.; Cui, L.; Liu, F.; Zhang, M.; Shan, L.; Yang, S.; Deng, X.-P. Effects of Drought stress on antioxidant enzymes in seedlings of different wheat genotypes. Pak. J. Bot. 2015, 47, 49–56. [Google Scholar]
- Iqbal, S.; Bano, A.; Ilyas, N. Drought and abscisic acid (ABA) induced changes in protein and pigment contents of four wheat (Triticum aestivum L.) Accessions. J. Agric. Res. 2010, 48, 1–13. [Google Scholar]
- Dbira, S.; Hassan, M.A.; Gramazio, P.; Ferchichi, A.; Vicente, O.; Prohens, J.; Boscaiu, M. Variable levels of tolerance to water stress (drought) and associated biochemical markers in Tunisian barley landraces. Molecules 2018, 23, 613. [Google Scholar] [CrossRef] [PubMed]
- Naveed, M.; Hussain, M.B.; Zahir, Z.A.; Mitter, B.; Sessitsch, A. Drought stress amelioration in wheat through inoculation with Burkholderia phytofirmans strain PsJN. Plant Growth Regul. 2014, 73, 121–131. [Google Scholar] [CrossRef]
- Rashid, U.; Yasmin, H.; Hassan, M.N.; Naz, R.; Nosheen, A.; Sajjad, M.; Ilyas, N.; Keyani, R.; Jabeen, Z.; Mumtaz, S.; et al. Drought-tolerant Bacillus megaterium isolated from semi-arid conditions induces systemic tolerance of wheat under drought conditions. Plant Cell Rep. 2022, 41, 549–569. [Google Scholar] [CrossRef]
- Azeem, M.; Haider, M.Z.; Javed, S.; Saleem, M.H.; Alatawi, A. Drought stress amelioration in maize (Zea mays L.) by inoculation of Bacillus spp. strains under sterile soil conditions. Agriculture 2022, 12, 50. [Google Scholar] [CrossRef]
- Khan, Z.; Rho, H.; Firrincieli, A.; Hung, S.H.; Luna, V.; Masciarelli, O.; Kim, S.-H.; Doty, S.L. Growth enhancement and drought tolerance of hybrid poplar upon inoculation with endophyte consortia. Curr. Plant Biol. 2016, 6, 38–47. [Google Scholar] [CrossRef]
- Jiang, M.Y.; Zhang, J.H. Abscisic acid and antioxidant defense in plant cells. Acta Bot. Sin. 2004, 46, 1–9. [Google Scholar]
- Todorova, D.; Sergiev, I.; Katerova, Z.; Shopova, E.; Dimitrova, L.; Brankova, L. Assessment of the biochemical responses of wheat seedlings to soil drought after application of selective herbicide. Plants 2021, 10, 733. [Google Scholar] [CrossRef]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef] [PubMed]
- Luna, C.M.; Pastori, G.M.; Driscoll, S.; Groten, K.; Bernard, S.; Foyer, C.H. Drought controls on H2O2 accumulation, catalase (CAT) activity and CAT gene expression in wheat. J. Exp. Bot. 2004, 56, 417–423. [Google Scholar] [CrossRef] [PubMed]
- Hung, S.-H.; Yu, C.-W.; Lin, C. Hydrogen peroxide functions as a stress signal in plants. Bot. Bull. Acad. Sin. 2005, 46, 1–10. [Google Scholar]
- Gontia-Mishra, I.; Sapre, S.; Sharma, A.; Tiwari, S. Amelioration of drought tolerance in wheat by the interaction of plant growth-promoting rhizobacteria. Plant Biol. 2016, 18, 992–1000. [Google Scholar] [CrossRef] [PubMed]
- Gong, H.-J.; Chen, K.-M.; Chen, G.-C.; Wang, S.-M.; Zhang, C.-L. Drought stress stimulates p-nitrophenyl phosphate hydrolysis rate of the plasma membrane H+-ATPase from wheat leaves. Plant Growth Regul. 2003, 40, 139–145. [Google Scholar] [CrossRef]
- Feng, X.; Liu, W.; Zeng, F.; Chen, Z.; Zhang, G.; Wu, F. K+ uptake, H+-ATPase pumping activity and Ca2+ efflux mechanism are involved in drought tolerance of barley. Environ. Exp. Bot. 2016, 129, 57–66. [Google Scholar] [CrossRef]
- Chang, N.; Ziwen, Z.; Yeyun, L.; Xianchen, Z. Exogenously applied Spd and Spm enhance drought tolerance in tea plants by increasing fatty acid desaturation and plasma membrane H+-ATPase activity. Plant Physiol. Biochem. 2022, 170, 225–233. [Google Scholar]
- Zhao, H.C.; Wang, B.C.; Cai, S.X.; Sakanishi, A. Effect of water stress on redox system in plasmic membrane of mature leaf of young P. betuloefolia Bqe. Colloids Surf. B. 2000, 18, 99–103. [Google Scholar] [CrossRef]
- Du, H.; Zhou, X.; Yang, Q.; Liu, H.; Kurtenbach, R. Changes in H+-ATPase activity and conjugated polyamine contents in plasma membrane purified from developing wheat embryos under short-time drought stress. Plant Growth Regul. 2015, 75, 1–10. [Google Scholar] [CrossRef]
- Qiu, Q.S.; Zhang, N. Water stress inhibits P-nitrophenyl phosphate hydrolysis activity of the plasma membrane H+-ATPase from soybean hypocotyls. Aust. J. Plant Physiol. 2000, 27, 717–721. [Google Scholar] [CrossRef]
- Butt, M.; Ayyub, C.M.; Amjad, M.; Ahmad, R. Proline application enhances growth of chilli by improving physiological and biochemical attributes under salt stress. Pak. J. Agric. Sci. 2016, 53, 43–49. [Google Scholar]
- Liang, X.; Zhang, L.; Natarajan, S.K.; Becker, D.F. Proline mechanisms of stress survival. Antioxid. Redox. Signal. 2013, 19, 998–1011. [Google Scholar] [CrossRef] [PubMed]
- Planchet, E.; Verdu, I.; Delahaie, J.; Cukier, C.; Girard, C.; Morère-Le Paven, M.C.; Limami, A.M. Abscisic acid-induced nitric oxide and proline accumulation in independent pathways under water-deficit stress during seedling establishment in Medicago truncatula. J. Exp. Bot. 2014, 65, 2161–2170. [Google Scholar] [CrossRef]
- Alexieva, V.; Sergiev, I.; Mapelli, S.; Karanov, E. The effect of drought and ultraviolet radiation on growth and stress markers in pea and wheat. Plant Cell Environ. 2001, 24, 1337–1344. [Google Scholar] [CrossRef]
- Nayyar, H. Variation in Osmoregulation in differentially drought-sensitive wheat genotypes involves calcium. Biol. Plant. 2003, 47, 541–547. [Google Scholar] [CrossRef]
- Maghsoudi, K.; Emam, Y.; Niazi, A.; Pessarakli, M.; Arvin, M.J. P5CS expression level and proline accumulation in the sensitive and tolerant wheat cultivars under control and drought stress conditions in the presence/absence of silicon and salicylic acid. J. Plant Interact. 2018, 13, 461–471. [Google Scholar] [CrossRef]
- Kour, D.; Rana, K.L.; Yadav, A.N.; Sheikh, I.; Kumar, V.; Dhaliwal, H.S.; Saxena, A. Amelioration of drought stress in foxtail millet BV growth promoting attributes. Environ. Sustain. 2020, 3, 23–34. [Google Scholar] [CrossRef]
- Ain-Lhout, F.; Zunzunegui, M.; Diaz Barradas, M.C.; Tirado, R.; Garcia Novo, F. Comparison of proline accumulation in two mediterranean shrubs subjected to natural and experimental water deficit. Plant Soil 2001, 230, 175–183. [Google Scholar] [CrossRef]
- Paul, M.J.; Primavesi, L.F.; Jhurreea, D.; Zhang, Y. Trehalose metabolism and signaling. Annu. Rev. Plant Biol. 2008, 59, 417–441. [Google Scholar] [CrossRef]
- Gusain, Y.S.; Singh, U.S.; Sharma, A.K. Bacterial mediated amelioration of drought stress in drought tolerant and susceptible cultivars of rice (Oryza sativa L.). Afr. J. Biotechnol. 2015, 14, 764–773. [Google Scholar]
- Kaushal, M. Portraying rhizobacterial mechanisms in drought tolerance: A way forward toward sustainable agriculture. In PGPR Amelioration in Sustainable Agriculture, 1st ed.; Singh, A.K., Kumar, A., Singh, P.K., Eds.; Woodhead Publishing: Sawston, UK, 2019; pp. 195–216. [Google Scholar]
- Vurukonda, S.S.K.P.; Vardharajula, S.; Shrivastava, M.; SkZ, A. Enhancement of drought stress tolerance in crops by plant growth promoting rhizobacteria. Microbiol. Res. 2016, 184, 13–24. [Google Scholar] [CrossRef] [PubMed]
- Valluru, R.; Davies, W.J.; Reynolds, M.P.; Dodd, I.C. Foliar Abscisic acid-to-ethylene accumulation and response regulate shoot growth sensitivity to mild drought in wheat. Front. Plant Sci. 2016, 7, 461. [Google Scholar] [CrossRef] [PubMed]
- Jankovska-Bortkevič, E.; Gavelienė, V.; Koryznienė, D.; Jankauskienė, J.; Mockevičiūtė, R.; Jurkonienė, S. Response of winter oilseed rape to imitated temperature fluctuations in autumn-winter period. Environ. Exp. Bot. 2019, 166, 103801. [Google Scholar] [CrossRef]
- Glick, B.R. Modulation of plant ethylene levels by the bacterial enzyme ACC deaminase. FEMS Microbiol. Lett. 2005, 251, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.P.; Shelke, G.M.; Kumar, A.; Jha, P.N. Biochemistry and genetics of ACC deaminase: A weapon to “stress ethylene” produced in plants. Front. Microbiol. 2015, 6, 937. [Google Scholar] [PubMed]
- Priyanka; Agrawal, T.; Kotasthane, A.S.; Kosharia, A.; Kushwah, R.; Zaidi, N.W.; Singh, U.S. Crop specific plant growth promoting effects of ACCd enzyme and siderophore producing and cynogenic fluorescent Pseudomonas. 3 Biotech 2017, 7, 27. [Google Scholar]
- Gupta, A.; Mishra, R.; Rai, S.; Bano, A.; Pathak, N.; Fujita, M.; Kumar, M.; Hasanuzzaman, M. Mechanistic insights of plant growth promoting bacteria mediated drought and salt stress tolerance in plants for sustainable agriculture. Int. J. Mol. Sci. 2022, 23, 3741. [Google Scholar] [CrossRef]
- Nezhadahmadi, A.; Prodhan, Z.H.; Faruq, G. Drought tolerance in wheat. Sci. World J. 2013, 610721. [Google Scholar] [CrossRef] [PubMed]
- Ali-Benali, M.A.; Alary, R.; Joudrier, P.; Gautier, M.F. Comparative expression of five Lea Genes during wheat seed development and in response to abiotic stresses by real-time quantitative RT-PCR. Biochim. Biophys. Acta Gene Struct. Expr. 2005, 1730, 56–65. [Google Scholar] [CrossRef]
- Weber, E.; Bleiholder, H.; Lancashire, P.D.; Langelüddecke, R.; Stauss, R.; Van der Boom, T.; Witzen-Berger, A. Growth stages of mono- and dicotyledonous plants. In BBCH Monograph., 1st ed.; Meier, U., Ed.; Julius Kühn-Institut: Quedlinburg, Germany, 2018; pp. 18–21. [Google Scholar]
- Hodges, D.; DeLong, J.; Forney, C.; Prange, R.K. Improving the thiobarbituric acid-reactive-substances assay for estimating lipid peroxidation in plant tissues containing anthocyanin and other interfering compounds. Planta 1999, 207, 604–611. [Google Scholar] [CrossRef]
- Velikova, V.; Yordanov, I.; Edreva, A. Oxidative stress and some antioxidant systems in acid rain-treated bean plants: Protective role of exogenous polyamines. Plant Sci. 2000, 151, 59–66. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Wellburn, A.R. The spectral determination of chlorophylls a and b, as well as total carotenoids, using various solvents with spectrophotometers of different resolution. J. Plant Physiol. 1994, 144, 307–313. [Google Scholar] [CrossRef]
- Child, R.D.; Chauvaux, N.; John, K.; Van Onckelen, H.A.; Ulvskov, P. Ethylene biosynthesis in oilseed rape pods in relation to pod shatter. J. Exp. Bot. 1998, 49, 829–838. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid method for the quantification of microgram quantities of proteins utilising the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Darginavičienė, J.; Pašakinskienė, I.; Maksimov, G.; Rognli, O.A.; Jurkonienė, S.; Šveikauskas, V.; Bareikienė, N. Changes in plasmalemma K+ Mg2+-ATPase dephosphorylating activity and H+ transport in relation to freezing tolerance and seasonal growth of Festuca pratensis Huds. J. Plant Physiol. 2008, 165, 825–832. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using Real-Time Quantitative PCR and the 2-ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Average Length (cm) | Average Weight (g) | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Fresh | Dry | ||||||||
| 6 Days | 12 Days | 18 Days | 6 Days | 12 Days | 18 Days | 6 Days | 12 Days | 18 Days | |
| Control | 33.17 d | 35.80 c | 39.45 a | 0.26 c | 0.33 d | 0.60 a | 0.032 d | 0.040 e | 0.065 b |
| Drought | 32.06 e | 33.67 e | 34.65 e | 0.23 e | 0.31 e | 0.15 f | 0.025 e | 0.037 f | 0.052 e |
| Calcium + Drouhgt | 33.20 d | 35.07 d | 38.32 b | 0.28 b | 0.37 c | 0.26 c | 0.035 c | 0.044 d | 0.062 c |
| Proline + Drought | 33.96 c | 36.22 c | 35.23 d | 0.25 d | 0.32 de | 0.16 ef | 0.032 d | 0.044 d | 0.060 d |
| Proline + Ca + Drought | 33.98 c | 36.76 bc | 37.45 c | 0.27 bc | 0.39 b | 0.16 e | 0.039 b | 0.051 b | 0.063 c |
| ProbioHumus + Drought | 34.28 b | 37.12 b | 38.43 b | 0.27 bc | 0.38 bc | 0.20 d | 0.040 b | 0.049 c | 0.065 b |
| ProbioHumus + Ca + Drought | 35.85 a | 39.38 a | 39.74 a | 0.31 a | 0.42 a | 0.28 b | 0.049 a | 0.055 a | 0.070 a |
| Treatment | Chlorophyll a | Chlorophyll b | Chlorophyll Ratio a/b | |||
|---|---|---|---|---|---|---|
| mg g−1 FW | ||||||
| 6 Days | 12 Days | 6 Days | 12 Days | 6 Days | 12 Days | |
| Control | 1.27 b | 1.19 a | 0.53 a | 0.54 a | 2.38 bc | 2.21 e |
| Drought | 1.20 e | 0.45 g | 0.49 d | 0.09 e | 2.45 b | 4.92 a |
| Calcium + Drouhgt | 1.21 de | 0.54 d | 0.51 bc | 0.14 cd | 2.35 c | 3.73 d |
| Proline + Drought | 1.24 c | 0.46 f | 0.50 c | 0.12 d | 2.46 b | 3.69 d |
| Proline + Ca + Drought | 1.32 a | 0.49 e | 0.51 bc | 0.13 cd | 2.57 a | 3.65 d |
| ProbioHumus + Drought | 1.22 d | 0.59 c | 0.52 b | 0.15 c | 2.34 c | 3.92 bc |
| ProbioHumus + Ca +Drought | 1.27 b | 0.75 b | 0.53 a | 0.17 b | 2.39 bc | 4.15 b |
| Gene | LEA Proteins Group | Primer Pairs | Primer Sequences (5′-3′) |
|---|---|---|---|
| 26S | F/R | CCGGTTGTTATGCCAATAGCA/GCGGCGCAGCAGTTCT | |
| Td11 | 2 | F/R | AGGTGATCGATGACAACGGTG/ACCCTCGGTGTCCTTGTGG |
| Td29b | 4 | F/R | CGCACCCAGCTAGTAAGTTCG/CCCAGCCCAGTAATAACCCAT |
| Td27e | 2 | F/R | CAGCACTGAGCCGACGG/ACGTGGAACTAGAAGGCATTGAC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mockevičiūtė, R.; Jurkonienė, S.; Šveikauskas, V.; Zareyan, M.; Jankovska-Bortkevič, E.; Jankauskienė, J.; Kozeko, L.; Gavelienė, V. Probiotics, Proline and Calcium Induced Protective Responses of Triticum aestivum under Drought Stress. Plants 2023, 12, 1301. https://doi.org/10.3390/plants12061301
Mockevičiūtė R, Jurkonienė S, Šveikauskas V, Zareyan M, Jankovska-Bortkevič E, Jankauskienė J, Kozeko L, Gavelienė V. Probiotics, Proline and Calcium Induced Protective Responses of Triticum aestivum under Drought Stress. Plants. 2023; 12(6):1301. https://doi.org/10.3390/plants12061301
Chicago/Turabian StyleMockevičiūtė, Rima, Sigita Jurkonienė, Vaidevutis Šveikauskas, Mariam Zareyan, Elžbieta Jankovska-Bortkevič, Jurga Jankauskienė, Liudmyla Kozeko, and Virgilija Gavelienė. 2023. "Probiotics, Proline and Calcium Induced Protective Responses of Triticum aestivum under Drought Stress" Plants 12, no. 6: 1301. https://doi.org/10.3390/plants12061301
APA StyleMockevičiūtė, R., Jurkonienė, S., Šveikauskas, V., Zareyan, M., Jankovska-Bortkevič, E., Jankauskienė, J., Kozeko, L., & Gavelienė, V. (2023). Probiotics, Proline and Calcium Induced Protective Responses of Triticum aestivum under Drought Stress. Plants, 12(6), 1301. https://doi.org/10.3390/plants12061301