
Designing Novel Strategies for Improving Old Legumes: An Overview from Common Vetch


Abstract
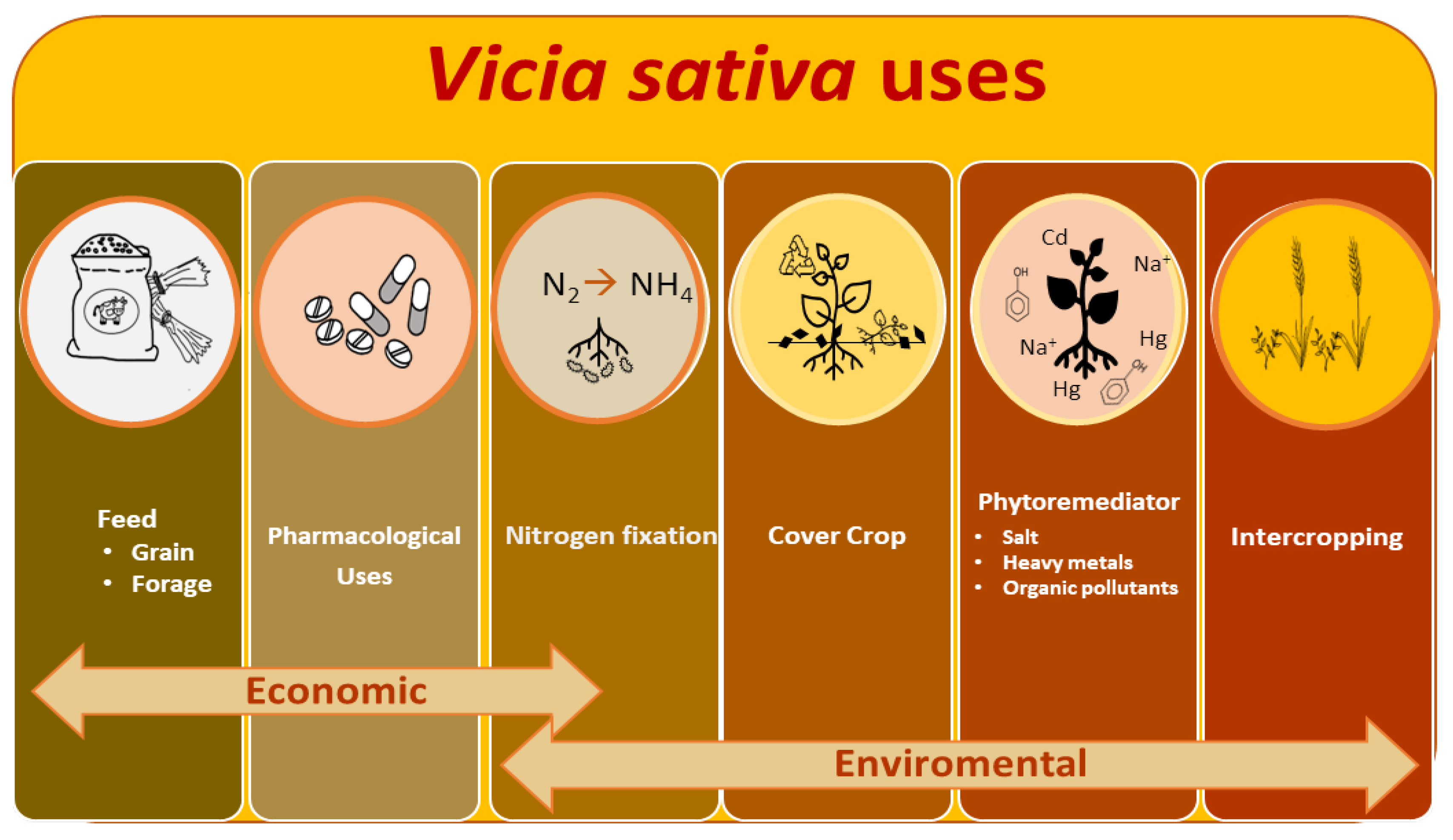
1. Introduction
2. Taxonomy
3. A Historical Crop
4. Worldwide Vetch Cultivation
5. Nutritional and Pharmacological Properties
6. Environmental Benefits

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pollutant | Developed Assay | References |
|---|---|---|
| Cd | Cd tolerance. Oxidative damage accumulation. | [96] |
| Cd and Zn | Zn and Cd accumulation in different tissues | [95] |
| Zn | Zn tolerance | [98] |
| Cu | Cu tolerance | [91,92] |
| Salt | Tolerance to salt. Na and K accumulation | [84] |
| Hg | Hg accumulation in different tissues | [94,100] |
| Ni | Ni accumulation. Oxidative damage accumulation. | [97] |
| Sulfosulfuron herbicide | Tolerance to sulfosulfuron | [85] |
| Diesel fuel | Tolerance to diesel | [90] |
| Phenol derivatives | Polychlorinated biphenyl (PCB) dissipation | [87] |
| Phenolics | Tolerance to phenolics. Effects on biomass, nodulation and nitrogen fixation activity | [89] |
| Mepiquat | Tolerance to mepiquat | [88] |
7. Pests and Diseases on Common Vetches
8. Germplasm Gene Banks and Common Vetch Genetic Diversity
9. Generating Genomic and Transcriptomic Tools
| Type of Molecular Marker | Target | References |
|---|---|---|
| Retrotranspon-derived Sequence-Specific Amplified Polymorphism (SSAP) | Genomic sequences | [128] |
| Amplified Fragment Length Polymorphism (AFLP) | Genomic sequences | [129] |
| Seed reserve protein patterns | Protein | [130,132] |
| Start Codon Targeted (SCoT) marker | cDNA sequences | [131] |
| Inter Simple Sequence Repeats (ISSR) | Genomic sequences | [132] |
| cDNA-SSR | cDNA sequences | [132,135,136,138,140,141,142] |
| SNP | cDNA sequences | [132,139,142] |
| double-digest restriction-site associated DNA sequencing (ddRAD-Seq) | Genomic sequences | [139] |
| genomic-SSR | Genomic sequences | [140] |
9.1. Genomic Data and Transcriptomic Characterization of Some Traits and Developmental Stages
9.2. Genomic Data and Transcriptomic Characterization of Stress Responses
| Process | Analyzed Plant Tissue | References |
|---|---|---|
| Flower Development | Floral organs (dorsal, lateral and ventral petals, sepals, stamens, carpels) leaf, and roots | [146] |
| Flowering time | Aerial part at different stages | [145] |
| Pod Shattering | Pod ventral sutures | [151] |
| Drought Tolerance | Whole plant under different drought treatments | [137] |
| Drought Stress | Root, stem, and leaf tissue under PEG treatments | [156] |
| Drought Stress | Comparative leaf versus root | [155] |
| Drought response and tolerance | Aerial part of tolerant and sensitive varieties | [142] |
| Cold–drought combined stress | Comparative leaf versus root | [158] |
| Drought Stress | Aerial part under PEG treatments | [157] |
| Salinity Stress | Leaf versus root | [153] |
| Hydrogen Cyanide Synthesis | Seed development | [53] |
10. Perspectives for Future Breeding Strategies
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Parissi, Z.; Irakli, M.; Tigka, E.; Papastylianou, P.; Dordas, C.; Tani, E.; Abraham, E.M.; Theodoropoulos, A.; Kargiotidou, A.; Kougiteas, L.; et al. Analysis of Genotypic and Environmental Effects on Biomass Yield, Nutritional and Antinutritional Factors in Common Vetch. Agronomy 2022, 12, 1678. [Google Scholar] [CrossRef]
- Huang, Y.F.; Gao, X.L.; Nan, Z.B.; Zhang, Z.X. Potential value of the common vetch (Vicia sativa L.) as an animal feedstuff: A review. J. Anim. Physiol. Anim. Nutr. 2017, 101, 807–823. [Google Scholar] [CrossRef]
- Lithourgidis, A.; Dordas, C.; Damalas, C.A.; Vlachostergios, D.N. Annual intercrops: An alternative pathway for sustainable agriculture. Aust. J. Crop Sci. 2011, 5, 396–410. [Google Scholar]
- Dalias, P.; Neocleous, D. Comparative Analysis of the Nitrogen Effect of Common Agricultural Practices and Rotation Systems in a Rainfed Mediterranean Environment. Plants 2017, 6, 61. [Google Scholar] [CrossRef]
- Maxted, N. An ecogeographical study of Vicia subgenus Vicia. In Systematic and Ecogeographic Studies on Crop Genepools; IPGRI: Rome, Italy, 1995; Volume 8, p. 184. [Google Scholar]
- Maxted, N. A phenetic investigation of Vicia L. subgenus Vicia (Leguminosae, Vicieae). Bot. J. Linn. Soc. 1993, 111, 155–182. [Google Scholar] [CrossRef]
- Hanelt, P.; Mettin, D. Biosystematics of the Genus Vicia L. (Leguminosae). Annu. Rev. Ecol. Syst. 1989, 20, 199–223. [Google Scholar] [CrossRef]
- Kupicha, F.K. The infrageneric structure of Vicia. Notes R. Bot. Gard. Edinb. 1976, 34, 287–326. [Google Scholar]
- Leht, M. Phylogenetics of Vicia (Fabaceae) based on morphological data. Feddes Repert. 2009, 120, 379–393. [Google Scholar] [CrossRef]
- Tate, M.; Ennerking, D. Vetches: From feed to food? Grain Legumes 2006, 47, 12–13. [Google Scholar]
- Potokina, E. Vicia sativa L. aggregate (Fabaceae) in the flora of former USSR. Genet. Resour. Crop Evol. 1997, 44, 199–209. [Google Scholar] [CrossRef]
- Jaaska, V. Isoenzyme diversity and phylogenetic affinities in Vicia subgenus Vicia (Fabaceae). Genet. Resour. Crop Evol. 1997, 44, 557–574. [Google Scholar] [CrossRef]
- Jaaska, V. Isozyme Variation and Phylogenetic Relationships in Vicia subgenus Cracca (Fabaceae). Ann. Bot. 2005, 96, 1085–1096. [Google Scholar] [CrossRef][Green Version]
- van de Wouw, M.; Maxted, N.; Chabane, K.; Ford-Lloyd, B.V. Molecular taxonomy of Vicia ser. Vicia based on Amplified Fragment Length Polymorphisms. Plant Syst. Evol. 2001, 229, 91–105. [Google Scholar] [CrossRef]
- Yeater, K.M.; Bollero, G.; Bullock, D.; Rayburn, A.L. Flow cytometric analysis for ploidy level differentiation of 45 hairy vetch accessions. Ann. Appl. Biol. 2004, 145, 123–127. [Google Scholar] [CrossRef]
- van de Wouw, M.; Maxted, N.I.; Ford-Lloyd, B.V. A multivariate and cladistic study of Vicia L. ser. Vicia (Fabaceae) based on analysis of morphological characters. Plant Syst. Evol. 2003, 237, 19–39. [Google Scholar] [CrossRef]
- Ennerking, D.; Tate, M. Global vetch production. Grain Legumes 2006, 47, 14–15. [Google Scholar]
- Zohary, D.; Hopf, M. Domestication of Plants in the Old World: The Origin and Spread of Cultivated Plants in West Asia, Europe and the Nile Valley (No. Ed. 3); Oxford University Press: Oxford, UK, 2000. [Google Scholar]
- Maxted, N.; Bennett, S. Plant Genetic Resources of Legumes in the Mediterranean; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2001; Volume 39. [Google Scholar]
- Tardío, J.; Pardo-de-Santayana, M.; Lazaro, A.; Aceituno, L.; Molina, M. Inventario Español de Conocimientos Tradicionales Relativos a la Biodiversidad Agrícola Volumen 2; Ministerio de Agricultura, Pesca y Alimentación: Madrid, Spain, 2022. [Google Scholar]
- Zohary, D.; Hopf, M.; Weiss, E. Domestication of Plants in the Old World: The Origin and Spread of Domesticated Plants in Southwest Asia, Europe, and the Mediterranean Basin; Oxford University Press: Oxford, UK, 2012. [Google Scholar]
- Varron, M. Rerum Rusticarum: Libri III; Cubero-Salmeron, J.I., Translator; Consejería de Agricultura y Pesca, Junta de Andalucía: Seville, Spain, 2010; 1st century BC. [Google Scholar]
- Holgado-Redondo, A.; Columela, L.J.M. (1st Century)-Translation; “De re Rustica”: De los Trabajos de Campo; Holgado-Redondo, A., Translator; Ministerio de Agricultura: Madrid, Spain, 1988; 339p, ISBN 84-323-0622-3. [Google Scholar]
- Hernandez, L.; Huerta, J. Secundus Plinius The Elder (1st Century) Historia Natural; UNAM-Universidad Nacional Autónoma de Mexico: Mexico City, Mexico, 1976; Hernandez, L. (books 1–25), Huerta, J. (books 26–37). [Google Scholar]
- Oroz-Reta, J.; Marcos-Casquero., M.A. Isidore of Seville (6–7th Century)-Translation “Etymologiae” Etimologias; Oroz-Reta, J.; Marcos-Casquero, M.A., Translators; BAC: Madrid, Spain, 1982; Volume 15. [Google Scholar]
- Cubero, J.I. Historia General de la Agricultura; Guadalmazán: Córdoba, Spain, 2018; ISBN 9788494155239. p. 840. [Google Scholar]
- Carabaza-Bravo, J.M.; L-Jayr, A.; Al-Filāḥa, K. Tratado de Agricultura; (11st–12nd Century) Translation; Carabaza-Bravo, J.M., Ed.; Instituto de Cooperación con el Mundo Arabe: Madrid, Spain, 1991. [Google Scholar]
- Jarava, J. “Historia de las Yerbas y Plantas” (from Dioscoride Anazarbeo); Gorda, L.G., Ed.; Heirs of A. Byrcman: Antwerp, Belgium, 1557. [Google Scholar]
- Weber, L.H.; Schifino-Wittmann, M.T. The Vicia sativa L. aggregate (Fabaceae) in southern Brazil: Karyotypes, phenology and qualitative morphology. Genet. Resour. Crop Evol. 1999, 46, 207–211. [Google Scholar] [CrossRef]
- Gomez-Ortega, C. Continuacion de la Flora Española, ó Historia de las Plantas de España, Que Escribía Don Joseph Quer. Ibarra, J.: Madrid, Spain, 1784; Volume V–VI. [Google Scholar]
- Faostat. 2023. Available online: https://www.fao.org (accessed on 15 February 2023).
- Nguyen, V.; Riley, S.; Nagel, S.; Fisk, I.; Searle, I.R. Common Vetch: A Drought Tolerant, High Protein Neglected Leguminous Crop With Potential as a Sustainable Food Source. Front. Plant Sci. 2020, 11, 818. [Google Scholar] [CrossRef]
- Grela, E.R.; Samolinska, W.; Rybinski, W.; Kiczorowska, B.; Kowalczuk-Vasilev, E.; Matras, J.; Wesolowska, S. Nutritional and Anti-Nutritional Factors in Vicia sativa L. Seeds and the Variability of Phenotypic and Morphological Characteristics of Some Vetch Accessions Cultivated in European Countries. Animals 2021, 11, 44. [Google Scholar] [CrossRef]
- Larbi, A.; El-Moneim, A.M.A.; Nakkoul, H.; Jammal, B.; Hassan, S. Intra-species variations in yield and quality determinants in Vicia species: 3. Common vetch (Vicia sativa ssp. sativa L.). Anim. Feed Sci. Technol. 2011, 164, 241–251. [Google Scholar] [CrossRef]
- Huang, Y.F.; Matthew, C.; Li, F.; Nan, Z.B. Common vetch varietal differences in hay nutritive value, ruminal fermentation, nutrient digestibility and performance of fattening lambs. Animal 2021, 15, 100244. [Google Scholar] [CrossRef]
- Salehi, B.; Abu-Reidah, I.M.; Sharopov, F.; Karazhan, N.; Sharifi-Rad, J.; Akram, M.; Daniyal, M.; Khan, F.S.; Abbaass, W.; Zainab, R.; et al. Vicia plants—A comprehensive review on chemical composition and phytopharmacology. Phytother. Res. 2021, 35, 790–809. [Google Scholar] [CrossRef]
- Abbasi, A.M.; Shah, M.H.; Li, T.; Fu, X.; Guo, X.; Liu, R.H. Ethnomedicinal values, phenolic contents and antioxidant properties of wild culinary vegetables. J. Ethnopharmacol. 2015, 162, 333–345. [Google Scholar] [CrossRef]
- Shinwari, M.I.; Khan, M.A. Folk use of medicinal herbs of Margalla Hills National Park, Islamabad. J. Ethnopharmacol. 2000, 69, 45–56. [Google Scholar] [CrossRef]
- Prabhu, S.; Vijayakumar, S.; Yabesh, J.E.; Ravichandran, K.; Sakthivel, B. Documentation and quantitative analysis of the local knowledge on medicinal plants in Kalrayan hills of Villupuram district, Tamil Nadu, India. J. Ethnopharmacol. 2014, 157, 7–20. [Google Scholar] [CrossRef]
- Marc, E.; Nellya, A.; Annick, D.D.; Frederic, D. Plants used as remedies antirheumatic and antineuralgic in the traditional medicine of Lebanon. J. Ethnopharmacol. 2008, 120, 315–334. [Google Scholar] [CrossRef]
- Nelson, K.; Lyles, J.T.; Li, T.; Saitta, A.; Addie-Noye, E.; Tyler, P.; Quave, C.L. Anti-Acne Activity of Italian Medicinal Plants Used for Skin Infection. Front. Pharmacol. 2016, 7, 425. [Google Scholar] [CrossRef]
- Saleem, M.; Karim, M.; Qadir, M.; Ahmed, B.; Rafiq, M.; Ahmad, B. In vitro antibacterial activity and phytochemical analysis of hexane extract of Vicia sativa. Bangladesh J. Pharmacol. 2014, 9, 189–193. [Google Scholar] [CrossRef]
- Megías, C.; Pastor-Cavada, E.; Torres-Fuentes, C.; Girón-Calle, J.; Barragán, M.A.; Juan, R.; Pastor, J.E.; Vioque, J. Chelating, antioxidant and antiproliferative activity of Vicia sativa polyphenol extracts. Eur. Food Res. Technol. 2009, 230, 353–359. [Google Scholar] [CrossRef]
- Ford, R. Vetch pod rupture associated with unrelated streak-inducing viruses of peas. Phytopathology 1965, 55, 935. [Google Scholar]
- Mao, Z.; Fu, H.; Nan, Z.; Wan, C. Fatty acid, amino acid, and mineral composition of four common vetch seeds on Qinghai-Tibetan plateau. Food Chem. 2015, 171, 13–18. [Google Scholar] [CrossRef]
- Fırıncıoğlu, H.K.; Ünal, S.; Erbektaş, E.; Doğruyol, L. Relationships between seed yield and yield components in common vetch (Vicia sativa ssp. sativa) populations sown in spring and autumn in central Turkey. Field Crops Res. 2010, 116, 30–37. [Google Scholar] [CrossRef]
- Firincioğlu, H.K.; Tate, M.; Ünal, S.; Doğruyol, L.; Özcan, İ. A Selection Strategy for Low Toxin Vetches (Vicia sativa spp.). Turk. J. Agric. For. 2007, 31, 303–311. [Google Scholar]
- Matić, R.; Nagel, S.; Robertson, S.; Young, I.; Mihailović, V.; Mikić, A.; Kirby, G. Vetch (Vicia spp) expansion and use in Australia. Biotechnol. Anim. Husb. 2005, 21, 203–207. [Google Scholar] [CrossRef]
- Daryanto, S.; Wang, L.; Jacinthe, P.A. Global Synthesis of Drought Effects on Food Legume Production. PLoS ONE 2015, 10, e0127401. [Google Scholar] [CrossRef]
- Koumas, A.; Economides, S. Replacement of Soybean Meal by Broad Bean or Common Vetch Seed in Lamb and Kid Fattening Diets. Tech. Bull. 1987, 88, 1–5. [Google Scholar]
- Delaere, I. The Chemistry of Vivia sativa L. Selection; University of Adelaide, Department of Plant Science: Adelaide, Australia, 1996. [Google Scholar]
- Rathjen, J.M. The Potential for Vicia sativa L. as a Grain Legume for South Australia/Thesis Jane Mary Rathjen. Ph.D. Thesis, The University of Adelaide, Adelaide, Australia, 1997. [Google Scholar]
- Li, M.; Zhao, L.; Zhou, Q.; Fang, L.; Luo, D.; Liu, W.; Searle, I.R.; Liu, Z. Transcriptome and Coexpression Network Analyses Provide In-Sights into the Molecular Mechanisms of Hydrogen Cyanide Synthesis during Seed Development in Common Vetch (Vicia sativa L.). Int. J. Mol. Sci. 2022, 23, 2275. [Google Scholar] [CrossRef]
- Akande, K.E.; Fabiyi, E.F. Effect of Processing Methods on Some Antinutritional Factors in Legume Seeds for Poultry Feeding. Int. J. Poult. Sci. 2010, 9, 996–1001. [Google Scholar] [CrossRef]
- Lambein, F.; Kuo, Y.H.; Ikegami, F.; Kusama-Eguchi, K.; Enneking, D. Grain legumes and human health. In Food Legumes for Nutritional Security and Sustainable Agriculture, Proceedings of the 4th International Food Legumes Research Conference, New Delhi, India, 18–22 October 2005; Indian Society of Genetics and Plant Breeding: New Delhi, India, 2009; pp. 422–432. [Google Scholar]
- Ampomah, O.; Huss-Danell, K. Genetic diversity of rhizobia nodulating native Vicia spp. in Sweden. Syst. Appl. Microbiol. 2016, 39, 203–210. [Google Scholar] [CrossRef]
- Daramola, D.A.; Hatzell, M.C. Energy Demand of Nitrogen and Phosphorus Based Fertilizers and Approaches to Circularity. ACS Energy Lett. 2003, 8, 1493–1501. [Google Scholar] [CrossRef]
- Lei, X.; Wang, E.T.; Chen, W.F.; Sui, X.H.; Chen, W.X. Diverse bacteria isolated from root nodules of wild Vicia species grown in temperate region of China. Arch. Microbiol. 2008, 190, 657–671. [Google Scholar] [CrossRef]
- Jorrin, B.; Imperial, J. Population Genomics Analysis of Legume Host Preference for Specific Rhizobial Genotypes in the Rhizobium leguminosarum bv. Viciae Symbioses. Mol. Plant Microbe Interact. 2015, 28, 310–318. [Google Scholar] [CrossRef] [PubMed]
- Alvarez-Martinez, E.R.; Valverde, A.; Ramirez-Bahena, M.H.; Garcia-Fraile, P.; Tejedor, C.; Mateos, P.F.; Santillana, N.; Zuniga, D.; Peix, A.; Velazquez, E. The analysis of core and symbiotic genes of rhizobia nodulating Vicia from different continents reveals their common phylogenetic origin and suggests the distribution of Rhizobium leguminosarum strains together with Vicia seeds. Arch. Microbiol. 2009, 191, 659–668. [Google Scholar] [CrossRef] [PubMed]
- Laus, M.C.; van Brussel, A.A.; Kijne, J.W. Exopolysaccharide structure is not a determinant of host-plant specificity in nodulation of Vicia sativa roots. Mol. Plant Microbe Interact. 2005, 18, 1123–1129. [Google Scholar] [CrossRef]
- Laus, M.C.; van Brussel, A.A.; Kijne, J.W. Role of cellulose fibrils and exopolysaccharides of Rhizobium leguminosarum in attachment to and infection of Vicia sativa root hairs. Mol. Plant Microbe Interact. 2005, 18, 533–538. [Google Scholar] [CrossRef]
- Muszynski, A.; Laus, M.; Kijne, J.W.; Carlson, R.W. Structures of the lipopolysaccharides from Rhizobium leguminosarum RBL5523 and its UDP-glucose dehydrogenase mutant (exo5). Glycobiology 2011, 21, 55–68. [Google Scholar] [CrossRef]
- Tak, T.; van Spronsen, P.C.; Kijne, J.W.; van Brussel, A.A.; Boot, K.J. Accumulation of lipochitin oligosaccharides and NodD-activating compounds in an efficient plant--Rhizobium nodulation assay. Mol. Plant Microbe Interact. 2004, 17, 816–823. [Google Scholar] [CrossRef]
- Recourt, K.; Schripsema, J.; Kijne, J.W.; van Brussel, A.A.; Lugtenberg, B.J. Inoculation of Vicia sativa subsp. nigra roots with Rhizobium leguminosarum biovar Viciae results in release of nod gene activating flavanones and chalcones. Plant Mol. Biol. 1991, 16, 841–852. [Google Scholar] [CrossRef]
- Göttfert, M. Regulation and function of rhizobial nodulation genes. FEMS Microbiol. Rev. 1993, 10, 39–63. [Google Scholar] [CrossRef] [PubMed]
- Álvarez-Aragón, R.; Manuel Palacios, J.M.; Ramirez-Parra, E. Rhizobial symbiosis promotes drought tolerance in Vicia sativa and Pisum sativum. Environ. Exp. Bot. 2023, 208, 105268. [Google Scholar] [CrossRef]
- Zhang, E.; Li, L.; Huang, G.; Huang, P.; Chai, Q. Regulation of fertilizer application on yield and root growth of spring wheat-faba bean intercropping system. Ying Yong Sheng Tai Xue Bao J. Appl. Ecol. 2002, 13, 939–942. [Google Scholar]
- Allende-Montalbán, R.; Martín-Lammerding, D.; del Mar Delgado, M.; Porcel, M.A.; Gabriel, J.L. Nitrate Leaching in Maize (Zea mays L.) and Wheat (Triticum aestivum L.) Irrigated Cropping Systems under Nitrification Inhibitor and/or Intercropping Effects. Agriculture 2022, 12, 478. [Google Scholar] [CrossRef]
- Garduno-Castro, Y.; Espinoza-Ortega, A.; Gonzalez-Esquivel, C.E.; Mateo-Salazar, B.; Arriaga-Jordan, C.M. Intercropped oats (Avena sativa)—common vetch (Vicia sativa) silage in the dry season for small-scale dairy systems in the highlands of central Mexico. Trop. Anim. Health Prod. 2009, 41, 827–834. [Google Scholar] [CrossRef] [PubMed]
- Keba, W.; Tolemariam, T.; Mohammed, A. Straw dry matter yield and quality of finger millet intercropped with selected vetch species at different seeding ratios in western Oromia, Ethiopia. Heliyon 2022, 8, e10433. [Google Scholar] [CrossRef]
- Hontoria, C.; Garcia-Gonzalez, I.; Quemada, M.; Roldan, A.; Alguacil, M.M. The cover crop determines the AMF community composition in soil and in roots of maize after a ten-year continuous crop rotation. Sci. Total Environ. 2019, 660, 913–922. [Google Scholar] [CrossRef] [PubMed]
- Genard, T.; Etienne, P.; Laine, P.; Yvin, J.C.; Diquelou, S. Nitrogen transfer from Lupinus albus L., Trifolium incarnatum L. and Vicia sativa L. contribute differently to rapeseed (Brassica napus L.) nitrogen nutrition. Heliyon 2016, 2, e00150. [Google Scholar] [CrossRef]
- Wang, Q.; Zhang, C.; Li, J.; Wu, X.; Long, Y.; Su, Y. Intercropping Vicia sativa L. Improves the Moisture, Microbial Community, Enzyme Activity and Nutrient in Rhizosphere Soils of Young Kiwifruit Plants and Enhances Plant Growth. Horticulturae 2021, 7, 335. [Google Scholar] [CrossRef]
- Ogilvie, C.M.; Ashiq, W.; Vasava, H.B.; Biswas, A. Quantifying Root-Soil Interactions in Cover Crop Systems: A Review. Agriculture 2021, 11, 218. [Google Scholar] [CrossRef]
- Baldwin, K.; Creamer, N. Cover Crops for Organic Farms; Center for Enviromental Farming Systems, NCSU-NCA&TSU-NCDA&CS; North Carolina Cooperative Extension Service: Raleigh, NC, USA, 2006; pp. 1–22. [Google Scholar]
- Trenton, R.; Carrie, O.; Kelsey, H.; Hannah, W.; Tyler, D. Understanding cover crops. Agric. Nat. Resour. 2018, FS2156, 1–8. [Google Scholar]
- Wiesmeier, M.; Lungu, M.; Hübner, R.; Cerbari, V. Remediation of degraded arable steppe soils in Moldova using vetch as green manure. Solid Earth 2015, 6, 609–620. [Google Scholar] [CrossRef]
- Rodrigo-Comino, J.; Terol, E.; Mora, G.; Giménez-Morera, A.; Cerdà, A. Vicia sativa Roth. Can Reduce Soil and Water Losses in Recently Planted Vineyards (Vitis vinifera L.). Earth Syst. Environ. 2020, 4, 827–842. [Google Scholar] [CrossRef]
- Renzi, J. Efecto de la estructura de cultivo y grado de madurez a cosecha sobre el rendimiento y la calidad de semillas de Vicia sativa L. y Vicia. villosa Roth., bajo riego. MSc. Thesis, Universidad nacional del Sur, Bahia Blanca, Argentina, 2009. [Google Scholar]
- Alonso-Ayuso, M.; Gabriel, J.L.; Pancorbo, J.L.; Quemada, M. Interseeding cover crops into maize: Characterization of species performance under Mediterranean conditions. Field Crops Res. 2020, 249, 107762. [Google Scholar] [CrossRef]
- Ali, H.; Khan, E.; Sajad, M.A. Phytoremediation of heavy metals-Concepts and applications. Chemosphere 2013, 91, 869–881. [Google Scholar] [CrossRef]
- Ibañez, S.; Medina, M.I.; Agostini, E. Vicia: A green bridge to clean up polluted environments. Appl. Microbiol. Biotechnol. 2020, 104, 13–21. [Google Scholar] [CrossRef] [PubMed]
- Lastiri-Hernández, M.A.; Alvarez-Bernal, D.; Bermúdez-Torres, K.; Cárdenas, G.C.; Ceja-Torres, L.F. Phytodesalination of a moderately saline soil combined with two inorganic amendments. Bragantia 2019, 78, 579–586. [Google Scholar] [CrossRef]
- Alonso-Prados, J.L.; Hernández-Sevillano, E.; Llanos, S.; Villarroya, M.; García-Baudín, J.M. Effects of sulfosulfuron soil residues on barley (Hordeum vulgare), sunflower (Helianthus annuus) and common vetch (Vicia sativa). Crop Prot. 2002, 21, 1061–1066. [Google Scholar] [CrossRef]
- Ibanez, S.G.; Sosa Alderete, L.G.; Medina, M.I.; Agostini, E. Phytoremediation of phenol using Vicia sativa L. plants and its antioxidative response. Environ. Sci. Pollut. Res. 2012, 19, 1555–1562. [Google Scholar] [CrossRef] [PubMed]
- Halfadji, A.; Portet-Koltalo, F.; Touabet, A.; Le Derf, F.; Morin, C.; Merlet-Machour, N. Phytoremediation of PCB: Contaminated Algerian soils using native agronomics plants. Environ. Geochem. Health 2022, 44, 117–132. [Google Scholar] [CrossRef]
- Tan, M.; Temel, S. Effect of mepiquat chloride, a growth retardant, on seed yield and yield components in common vetch (Vicia sativa). Indian J. Agric. Sci. 2005, 75, 160–161. [Google Scholar]
- Machrafi, Y.; Prévost, D.; Beauchamp, C.J. Toxicity of phenolic compounds extracted from bark residues of different ages. J. Chem. Ecol. 2006, 32, 2595–2615. [Google Scholar] [CrossRef]
- Adam, G.; Duncan, H. The effect of diesel fuel on common vetch (Vicia sativa L.) plants. Environ. Geochem. Health 2003, 25, 123–130. [Google Scholar] [CrossRef]
- Muccifora, S.; Bellani, L.M. Effects of copper on germination and reserve mobilization in Vicia sativa L. seeds. Environ. Pollut. 2013, 179, 68–74. [Google Scholar] [CrossRef] [PubMed]
- Bellani, L.M.; Muccifora, S.; Giorgetti, L. Response to copper bromide exposure in Vicia sativa L. seeds: Analysis of genotoxicity, nucleolar activity and mineral profile. Ecotoxicol. Environ. Saf. 2014, 107, 245–250. [Google Scholar] [CrossRef]
- Ibañez, S.G.; Villasuso, A.L.; Racagni, G.E.; Agostini, E.; Medina, M.I. Phenol modulates lipid kinase activities in Vicia sativa plants. Environ. Exp. Bot. 2016, 122, 109–114. [Google Scholar] [CrossRef]
- Sierra, M.J.; Millán, R.; Esteban, E.; Cardona, A.I.; Schmid, T. Evaluation of mercury uptake and distribution in Vicia sativa L. applying two different study scales: Greenhouse conditions and lysimeter experiments. J. Geochem. Explor. 2008, 96, 203–209. [Google Scholar] [CrossRef]
- Bogatu, C.; Masu, S.; Lazarovici, M. Metals extraction from polluted soils by using of pillared zeolite and Vicia sativa. In Proceedings of the 14th Symposium on Analytical and Environmental Problems, Szeged, Hungary, 24 September 2007. [Google Scholar]
- Rui, H.; Zhang, X.; Shinwari, K.I.; Zheng, L.; Shen, Z. Comparative transcriptomic analysis of two Vicia sativa L. varieties with contrasting responses to cadmium stress reveals the important role of metal transporters in cadmium tolerance. Plant Soil 2018, 423, 241–255. [Google Scholar] [CrossRef]
- Ivanishchev, V.V.; Abramova, E.A. Accumulation of nickel ions in seedlings of Vicia sativa L. and manifestations of oxidative stress. Environ. Sci. Pollut. Res. 2015, 22, 7897–7905. [Google Scholar] [CrossRef]
- Masu, S.; Lixandru, B.; Bogatu, C. Zinc extraction from polluted soils by using zeolite and Vicia sativa plant. In Proceedings of the 3rd International Conference on Life Cycle Management, Zurich, Switzerland, 27–29 August 2007. [Google Scholar]
- Zhang, F.; Zhang, H.; Wang, G.; Xu, L.; Shen, Z. Cadmium-induced accumulation of hydrogen peroxide in the leaf apoplast of Phaseolus aureus and Vicia sativa and the roles of different antioxidant enzymes. J. Hazard. Mater. 2009, 168, 76–84. [Google Scholar] [CrossRef]
- Zhang, X.; Zhang, L.; Chen, L.; Lu, Y.; An, Y. Ectopic expression gamma-glutamylcysteine synthetase of Vicia sativa increased cadmium tolerance in Arabidopsis. Gene 2022, 823, 146358. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Rui, H.; Zhang, F.; Hu, Z.; Xia, Y.; Shen, Z. Overexpression of a Functional Vicia sativa PCS1 Homolog Increases Cadmium Tolerance and Phytochelatins Synthesis in Arabidopsis. Front. Plant Sci. 2018, 9, 107. [Google Scholar] [CrossRef] [PubMed]
- Ibañez, S.G.; Merini, L.J.; Barros, G.G.; Medina, M.I.; Agostini, E. Vicia sativa-Rhizospheric bacteria interactions to improve phenol remediation. Int. J. Environ. Sci. Technol. 2014, 11, 1679–1690. [Google Scholar] [CrossRef]
- Anderson, T.A.; Guthrie, E.A.; Walton, B.T. Bioremediation in the rhizosphere. Environ. Sci. Technol. 1993, 27, 2630–2636. [Google Scholar] [CrossRef]
- Ibañéz, S.G.; Oller, A.L.W.; Paisio, C.E.; Alderete, L.G.S.; González, P.S.; Medina, M.I.; Agostini, E. The challenges of remediating metals using phytotechnologies. In Heavy Metals in the Environment: Microorganisms and Bioremediation; CRC Press: Boca Raton, FL, USA, 2018; pp. 173–191. [Google Scholar]
- Renzi, J.P.; Cantamutto, M.A. Vicias: Bases Agronómicas para el Manejo en la Región Pampeana; Ediciones INTA: Buenos Aires, Argentina, 2013; ISBN 978-987-521-470-5. [Google Scholar]
- InfoAgro. El Cultivo de la Veza (Vicia spp.). Available online: https://www.infoagro.com/documentos/el_cultivo_veza____em_Vicia__em__spp__.asp (accessed on 15 February 2023).
- Cordo, H.A.; Logarzo, G.; Braun, K.; Di Iorio, O.R. Plagas y Enemigos naturales. In Catálogo de Insectos Fitófagos de la Argentina y sus Plantas Asociadas; Sociedad Entomológica Argentina Ediciones: Buenos Aires, Argentina, 2004; Volume 6, p. 734. [Google Scholar]
- Xu, S.; Li, Y.-Z. Research advances on fungal diseases of Vicia sativa. Acta Prataculturae Sin. 2016, 25, 203. [Google Scholar]
- Allen, R.; Hayward, A.C.; Halliday, W.G.; Fulcher, J.G. Bacterial Blight of Vicia sativa: Aetiology of the Disease and Identification of the Pathogen. Aust. J. Biol. Sci. 1970, 23, 597–606. [Google Scholar] [CrossRef]
- Martín-Sanz, A.; Palomo, J.L.; Caminero, C. First Report of Bacterial Blight Caused by Pseudomonas syringae pv. syringae on Common Vetch in Spain. Plant Dis. 2009, 93, 1348. [Google Scholar] [CrossRef]
- Sastry, K.S.; Mandal, B.; Hammond, J.; Scott, S.W.; Briddon, R.W. Vicia sativa (Vetch). In Encyclopedia of Plant Viruses and Viroids; Springer: New Delhi, India, 2019; pp. 2697–2699. [Google Scholar]
- Terzakis, M.; Avgelis, A.D.; Jones, A.T.; Katis, N.I. Artichoke yellow ringspot virus infecting Vetch (Vicia sativa) in Greece. Phytoparasitica 2002, 30, 195–197. [Google Scholar] [CrossRef]
- McCord, R.W.; Gudauskas, R.T. Properties of a strain of bean yellow mosaic virus isolated from vetch, Vicia sativa. Phytopathology 1968, 58, 1294–1297. [Google Scholar]
- Musil, M.; Leskova, O.; Kleja, S. The occurrence of broad bean stain virus on vetch plants in Slovakia. Ochr. Rostl. 1978, 14, 161–166. [Google Scholar]
- Asaad, N.Y.; Kumari, S.G.; Haj-Kassem, A.A.; Shalaby, A.B.A.; Al-Shaabi, S.; Malhotra, R.S. Detection and characterization of Chickpea Chlorotic Stunt Virus in Syria. J. Phytopathol. 2009, 157, 756–761. [Google Scholar] [CrossRef]
- Kumari, S.G.; Attar, N.; Mustafayev, E.; Akparov, Z. First Report of Faba bean necrotic yellows virus Affecting Legume Crops in Azerbaijan. Plant Dis. 2009, 93, 1220. [Google Scholar] [CrossRef] [PubMed]
- Rubiales, D.; Sadiki, M.; Román, B. First Report of Orobanche foetida on Common Vetch (Vicia sativa) in Morocco. Plant Dis. 2005, 89, 528. [Google Scholar] [CrossRef]
- Rubiales, D.; Fernandez-Aparicio, M.; Rodriguez, M.J. First Report of Crenate Broomrape (Orobanche crenata) on Lentil (Lens culinaris) and Common Vetch (Vicia sativa) in Salamanca Province, Spain. Plant Dis. 2008, 92, 1368. [Google Scholar] [CrossRef]
- Evidente, A.; Cimmino, A.; Fernandez-Aparicio, M.; Rubiales, D.; Andolfi, A.; Melck, D. Soyasapogenol B and trans-22-dehydrocam- pesterol from common vetch (Vicia sativa L.) root exudates stimulate broomrape seed germination. Pest Manag. Sci. 2011, 67, 1015–1022. [Google Scholar] [CrossRef]
- Perez-de-Luque, A.; Lozano, M.D.; Cubero, J.I.; Gonzalez-Melendi, P.; Risueno, M.C.; Rubiales, D. Mucilage production during the incompatible interaction between Orobanche crenata and Vicia sativa. J. Exp. Bot. 2006, 57, 931–942. [Google Scholar] [CrossRef]
- Cordoba, E.M.; Fernandez-Aparicio, M.; Gonzalez-Verdejo, C.I.; Lopez-Grau, C.; Muñoz-Muñoz, M.V.; Nadal, S. Search for Resistant Genotypes to Cuscuta campestris Infection in Two Legume Species, Vicia sativa and Vicia ervilia. Plants 2021, 10, 738. [Google Scholar] [CrossRef]
- Maalouf, F.; Somanagouda, P.; Rajendran, K.; Hamwieh, A.; Goyal, A.; Agrawal, S.K. Breeding for post-emergence herbicide tolerance in cool-season food legumes. In Proceedings of the International Year Pulses ICARDA Conference, Marrakesh, Morocco, 18–20 April 2016. [Google Scholar]
- Kumar, N.; Nath, C.; Hazra, K. Efficient weed management in pulses for higher productivity and profitability. Indian J. Agron. 2016, 61, 5199–5213. [Google Scholar]
- Fraser, J.; McCartney, D.; Najda, H.; Mir, Z. Yield potential and forage quality of annual forage legumes in southern Alberta and northeast Saskatchewan. Can. J. Plant Sci. 2004, 84, 143–155. [Google Scholar] [CrossRef][Green Version]
- Ramírez-Villegas, J.; Jarvis, A.; Fujisaka, S.; Hanson, J.; Leibing, C. Crop and forage genetic resources: International interdependence in the face of climate change. In Crop Genetic Resources as a Global Commons: Challenges in International Law and Governance; Halewood, M., López Noriega, I., Louafi, S., Eds.; Routledge: Abingdon, UK, 2013; pp. 78–98. [Google Scholar]
- Pellegrini, P.A.; Balatti, G.E. Noah’s arks in the XXI century. A typology of seed banks. Biodivers. Conserv. 2016, 25, 2753–2769. [Google Scholar] [CrossRef]
- Dempewolf, H.; Baute, G.; Anderson, J.; Kilian, B.; Smith, C.; Guarino, L. Past and Future Use of Wild Relatives in Crop Breeding. Crop Sci. 2017, 57, 1070–1082. [Google Scholar] [CrossRef]
- Sanz, A.M.; Gonzalez, S.G.; Syed, N.H.; Suso, M.J.; Saldaña, C.C.; Flavell, A.J. Genetic diversity analysis in Vicia species using retrotransposon-based SSAP markers. Mol. Genet. Genom. 2007, 278, 433–441. [Google Scholar] [CrossRef]
- Potokina, E.K.; Aleksandrova, T.G. Genetic singularity coefficients of common vetch Vicia sativa L. accessions determined with molecular markers. Genetika 2008, 44, 1508–1516. [Google Scholar] [CrossRef]
- De la Rosa, L.; González, J.M. The genetic diversity associated with seed proteins in a collection of Spanish underground vetches (Vicia sativa L. subsp. amphicarpa (Dorthes) Asch. et Graebn.). Genet. Resour. Crop Evol. 2010, 57, 565–573. [Google Scholar] [CrossRef]
- Chai, X.; Dong, R.; Liu, W.; Wang, Y.; Liu, Z. Optimizing Sample Size to Assess the Genetic Diversity in Common Vetch (Vicia sativa L.) Populations Using Start Codon Targeted (SCoT) Markers. Molecules 2017, 22, 567. [Google Scholar] [CrossRef]
- De la Rosa, L.; López-Román, M.I.; González, J.M.; Zambrana, E.; Marcos-Prado, T.; Ramírez-Parra, E. Common Vetch, Valuable Germplasm for Resilient Agriculture: Genetic Characterization and Spanish Core Collection Development. Front. Plant Sci. 2021, 12, 617873. [Google Scholar] [CrossRef]
- Edwards, D.; Batley, J. Plant genome sequencing: Applications for crop improvement. Plant Biotechnol. J. 2010, 8, 2–9. [Google Scholar] [CrossRef]
- Kumar, J.; Gupta, D.S.; Gupta, S.; Dubey, S.; Gupta, P.; Kumar, S. Quantitative trait loci from identification to exploitation for crop improvement. Plant Cell Rep. 2017, 36, 1187–1213. [Google Scholar] [CrossRef]
- Chung, J.W.; Kim, T.S.; Suresh, S.; Lee, S.Y.; Cho, G.T. Development of 65 novel polymorphic cDNA-SSR markers in common vetch (Vicia sativa subsp. sativa) using next generation sequencing. Molecules 2013, 18, 8376–8392. [Google Scholar] [CrossRef]
- Liu, Z.; Liu, P.; Luo, D.; Liu, W.; Wang, Y. Exploiting Illumina sequencing for the development of 95 novel polymorphic EST-SSR markers in common vetch (Vicia sativa subsp. sativa). Molecules 2014, 19, 5777–5789. [Google Scholar] [CrossRef]
- Zhu, Y.; Liu, Q.; Xu, W.; Zhang, J.; Wang, X.; Nie, G.; Yao, L.; Wang, H.; Lin, C. De Novo Assembly and Discovery of Genes That Involved in Drought Tolerance in the Common Vetch. Int. J. Mol. Sci. 2019, 20, 328. [Google Scholar] [CrossRef]
- Raveendar, S.; Lee, G.A.; Jeon, Y.A.; Lee, Y.J.; Lee, J.R.; Cho, G.T.; Cho, J.H.; Park, J.H.; Ma, K.H.; Chung, J.W. Cross-amplification of Vicia sativa subsp. sativa microsatellites across 22 other Vicia species. Molecules 2015, 20, 1543–1550. [Google Scholar] [CrossRef]
- Shirasawa, K.; Kosugi, S.; Sasaki, K.; Ghelfi, A.; Okazaki, K.; Toyoda, A. Genome features of common vetch (Vicia sativa) in natural habitats. Plant Direct 2021, 5, e352. [Google Scholar] [CrossRef]
- Ma, L.; Wang, X.; Yan, M.; Liu, F.; Zhang, S.; Wang, X. Genome survey sequencing of common vetch (Vicia sativa L.) and genetic diversity analysis of Chinese germplasm with genomic SSR markers. Mol. Biol. Rep. 2022, 49, 313–320. [Google Scholar] [CrossRef] [PubMed]
- Kim, T.S.; Raveendar, S.; Suresh, S.; Lee, G.A.; Lee, J.R.; Cho, J.H.; Lee, S.Y.; Ma, K.H.; Cho, G.T.; Chung, J.W. Transcriptome Analysis of Two Vicia sativa Subspecies: Mining Molecular Markers to Enhance Genomic Resources for Vetch Improvement. Genes 2015, 6, 1164–1182. [Google Scholar] [CrossRef]
- De la Rosa, L.; Zambrana, E.; Ramirez-Parra, E. Molecular bases for drought tolerance in common vetch: Designing new molecular breeding tools. BMC Plant Biol. 2020, 20, 71. [Google Scholar] [CrossRef]
- Ladizinsky, G.; Shefer, Y. Polyploidy in the Vicia sativa aggregate. New Phytol. 1982, 91, 541–547. [Google Scholar] [CrossRef]
- Ladizinsky, G.; Temkin, R. The cytogenetic structure of Vicia sativa aggregate. Theor. Appl. Genet. 1978, 53, 33–42. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Q.; Cui, Y.; Dong, R.; Luo, D.; Fang, L.; Nan, Z.; Liu, Z. Integrative Analyses of Transcriptomes and Metabolomes Reveal Associated Genes and Metabolites with Flowering Regulation in Common Vetch (Vicia sativa L.). Int. J. Mol. Sci. 2022, 23, 6818. [Google Scholar] [CrossRef]
- Liu, Z.; Ma, L.; Nan, Z.; Wang, Y. Comparative transcriptional profiling provides insights into the evolution and development of the zygomorphic flower of Vicia sativa (Papilionoideae). PLoS ONE 2013, 8, e57338. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Wang, J.; Kuang, H.; Hou, Z.; Gong, P.; Bai, M.; Zhou, S.; Yao, X.; Song, S.; Yan, L.; et al. Elimination of an unfavorable allele conferring pod shattering in an elite soybean cultivar by CRISPR/Cas9. Abiotech 2022, 3, 110–114. [Google Scholar] [CrossRef] [PubMed]
- Aguilar-Benitez, D.; Rubio, J.; Millán, T.; Gil, J.; Die, J.V.; Castro, P. Genetic analysis reveals PDH1 as a candidate gene for control of pod dehiscence in chickpea. Mol. Breed. 2020, 40, 40. [Google Scholar] [CrossRef]
- Parker, T.A.; Berny Mier y Teran, J.C.; Palkovic, A.; Jernstedt, J.; Gepts, P. Pod indehiscence is a domestication and aridity resilience trait in common bean. New Phytol. 2020, 225, 558–570. [Google Scholar] [CrossRef]
- Abd El-Moneim, A.M. Selection for Non-Shattering Common Vetch, Vicia sativa L. Plant Breed. 1993, 110, 168–171. [Google Scholar] [CrossRef]
- Dong, R.; Dong, D.; Luo, D.; Zhou, Q.; Chai, X.; Zhang, J.; Xie, W.; Liu, W.; Dong, Y.; Wang, Y.; et al. Transcriptome Analyses Reveal Candidate Pod Shattering-Associated Genes Involved in the Pod Ventral Sutures of Common Vetch (Vicia sativa L.). Front. Plant Sci. 2017, 8, 649. [Google Scholar] [CrossRef]
- Sun, Y.; Li, J.; Xing, J.; Yu, X.; Lu, Y.; Xu, W.; Zhao, N.; Liu, Z.; Guo, Z. Evaluation of salt tolerance in common vetch (Vicia sativa L.) germplasms and the physiological responses to salt stress. J. Plant Physiol. 2022, 278, 153811. [Google Scholar] [CrossRef]
- Lin, X.; Wang, Q.; Min, X.; Liu, W.; Liu, Z. Comparative Transcriptomic Analysis of Root and Leaf Transcript Profiles Reveals the Coordinated Mechanisms in Response to Salinity Stress in Common Vetch. Int. J. Mol. Sci. 2022, 23, 8477. [Google Scholar] [CrossRef] [PubMed]
- Min, X.; Lin, X.; Ndayambaza, B.; Wang, Y.; Liu, W. Coordinated mechanisms of leaves and roots in response to drought stress underlying full-length transcriptome profiling in Vicia sativa L. BMC Plant Biol. 2020, 20, 165. [Google Scholar] [CrossRef] [PubMed]
- Tenopala, J.; Gonzalez, F.; De la Barrera, E. Physiological responses of the green manure, Vicia sativa, to drought. Bot. Sci. 2012, 90, 305–311. [Google Scholar] [CrossRef][Green Version]
- Wei, X.; Jin, X.; Ndayambaza, B.; Min, X.; Zhang, Z.; Wang, Y.; Liu, W. Transcriptome-Wide Characterization and Functional Identification of the Aquaporin Gene Family During Drought Stress in Common Vetch. DNA Cell Biol. 2019, 38, 374–384. [Google Scholar] [CrossRef]
- Zhu, Y.; Liu, Q.; Xu, W.; Yao, L.; Wang, X.; Wang, H.; Xu, Y.; Li, L.; Duan, C.; Yi, Z.; et al. Identification of novel drought-responsive miRNA regulatory network of drought stress response in common vetch (Vicia sativa). Open Life Sci. 2021, 16, 1111–1121. [Google Scholar] [CrossRef]
- Min, X.; Wang, Q.; Wei, Z.; Liu, Z.; Liu, W. Full-length transcriptional analysis reveals the complex relationship of leaves and roots in responses to cold-drought combined stress in common vetch. Front. Plant Sci. 2022, 13, 976094. [Google Scholar] [CrossRef]
- Khatun, M.; Sarkar, S.; Era, F.M.; Islam, A.K.M.M.; Anwar, M.P.; Fahad, S.; Datta, R.; Islam, A.K.M.A. Drought Stress in Grain Legumes: Effects, Tolerance Mechanisms and Management. Agronomy 2021, 11, 2374. [Google Scholar] [CrossRef]
- Nadeem, M.; Li, J.; Yahya, M.; Sher, A.; Ma, C.; Wang, X.; Qiu, L. Research Progress and Perspective on Drought Stress in Legumes: A Review. Int. J. Mol. Sci. 2019, 20, 2541. [Google Scholar] [CrossRef]
- Nguyen, V.; Searle, I.R. An Efficient Root Transformation System for Recalcitrant Vicia sativa. Front. Plant Sci. 2022, 12, 781014. [Google Scholar] [CrossRef]
- Rolli, E.; Marasco, R.; Vigani, G.; Ettoumi, B.; Mapelli, F.; Deangelis, M.L.; Gandolfi, C.; Casati, E.; Previtali, F.; Gerbino, R.; et al. Improved plant resistance to drought is promoted by the root-associated microbiome as a water stress-dependent trait. Environ. Microbiol. 2015, 17, 316–331. [Google Scholar] [CrossRef]
- Pastor-Cavada, E.; Drago, S.R.; González, R.J.; Juan, R.; Pastor, J.E.; Alaiz, M.; Vioque, J. Physical and nutritional properties of extruded products based on whole grain with the addition of wild legumes (Vicia lutea subsp. lutea var. hirta and Vicia sativa subsp. sativa). Int. J. Food Sci. Technol. 2013, 48, 1949–1955. [Google Scholar] [CrossRef]
- Hernandez-Aguirre, A.I.; Téllez-Pérez, C.; San Martín-Azócar, A.; Cardador-Martínez, A. Effect of Instant Controlled Pressure-Drop (DIC), Cooking and Germination on Non-Nutritional Factors of Common Vetch (Vicia sativa spp.). Molecules 2020, 25, 151. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ramírez-Parra, E.; De la Rosa, L. Designing Novel Strategies for Improving Old Legumes: An Overview from Common Vetch. Plants 2023, 12, 1275. https://doi.org/10.3390/plants12061275
Ramírez-Parra E, De la Rosa L. Designing Novel Strategies for Improving Old Legumes: An Overview from Common Vetch. Plants. 2023; 12(6):1275. https://doi.org/10.3390/plants12061275
Chicago/Turabian StyleRamírez-Parra, Elena, and Lucía De la Rosa. 2023. "Designing Novel Strategies for Improving Old Legumes: An Overview from Common Vetch" Plants 12, no. 6: 1275. https://doi.org/10.3390/plants12061275
APA StyleRamírez-Parra, E., & De la Rosa, L. (2023). Designing Novel Strategies for Improving Old Legumes: An Overview from Common Vetch. Plants, 12(6), 1275. https://doi.org/10.3390/plants12061275