
Facilitation by Haloxylon persicum Shrubs Enhances Density and Richness of Soil Seed Bank of Annual Plants in a Hyper-Arid Ecosystem

Abstract
1. Introduction
2. Results
2.1. Seed Bank Composition
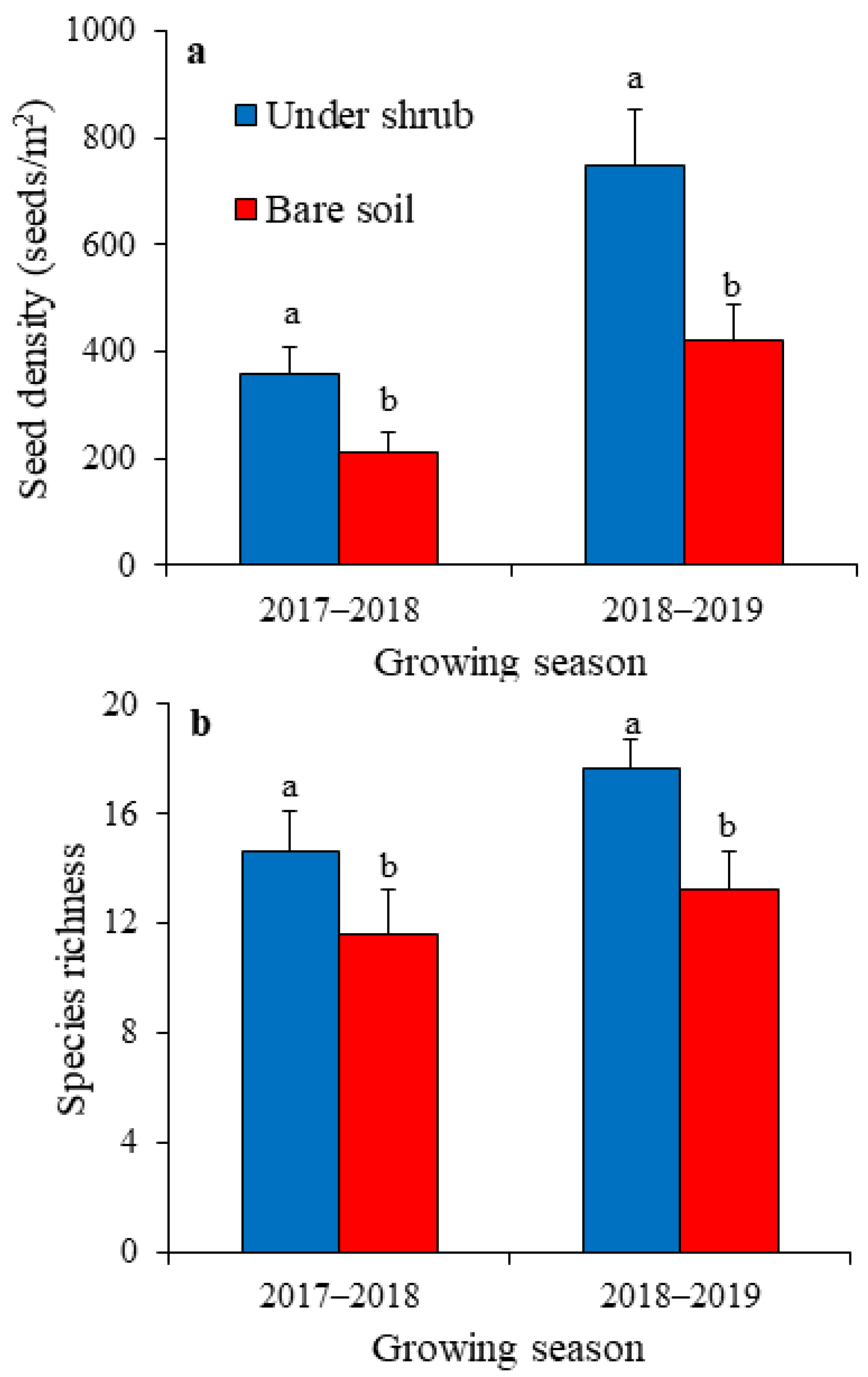
2.2. Shrub Effects on the Soil Seed Bank of Annual Plants
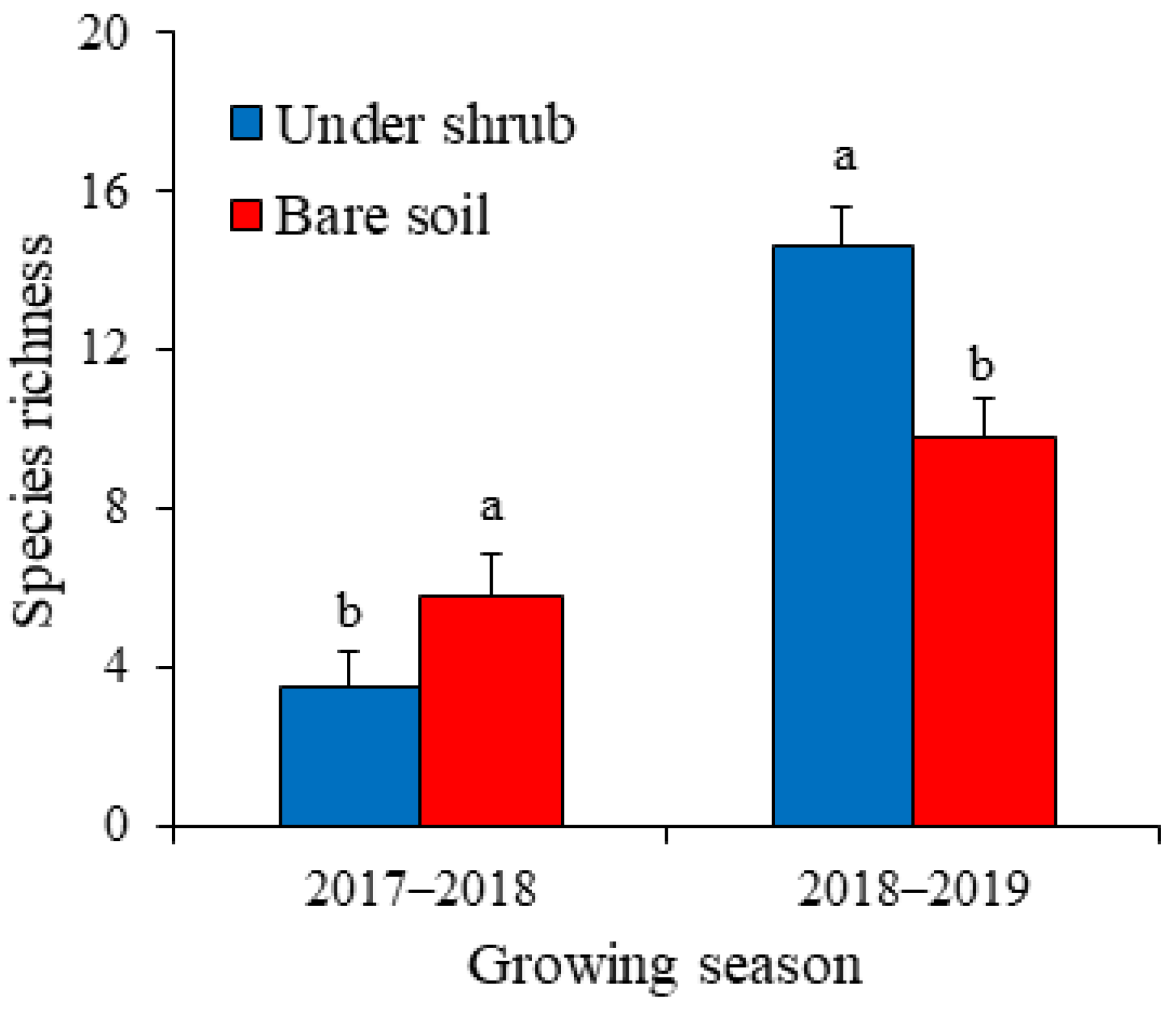
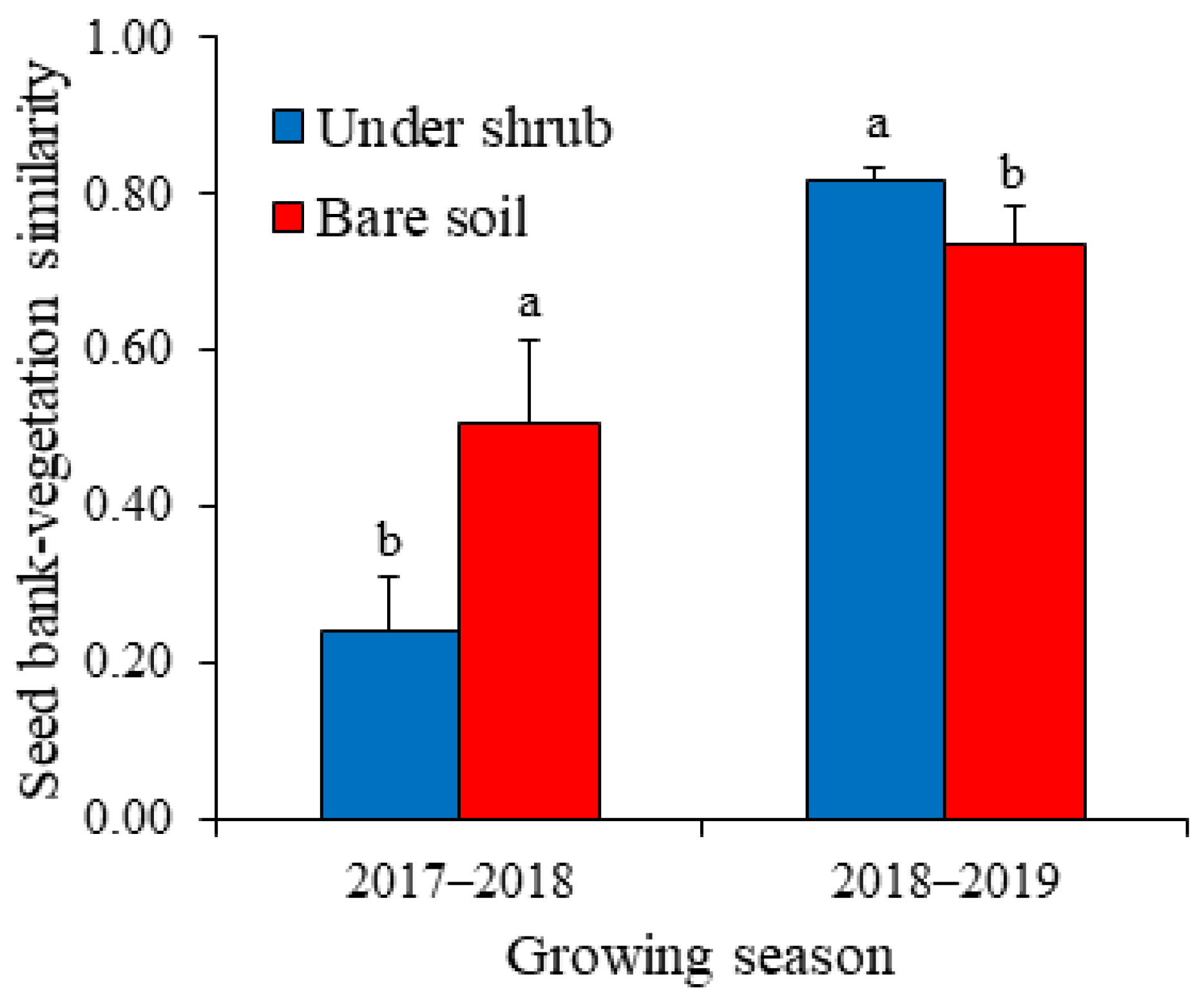
2.3. Shrub Effects on Above-Ground Annual Vegetation and Seed Bank–Vegetation Similarity
2.4. Shrub Effects on Soil Properties
3. Discussion
4. Materials and Methods
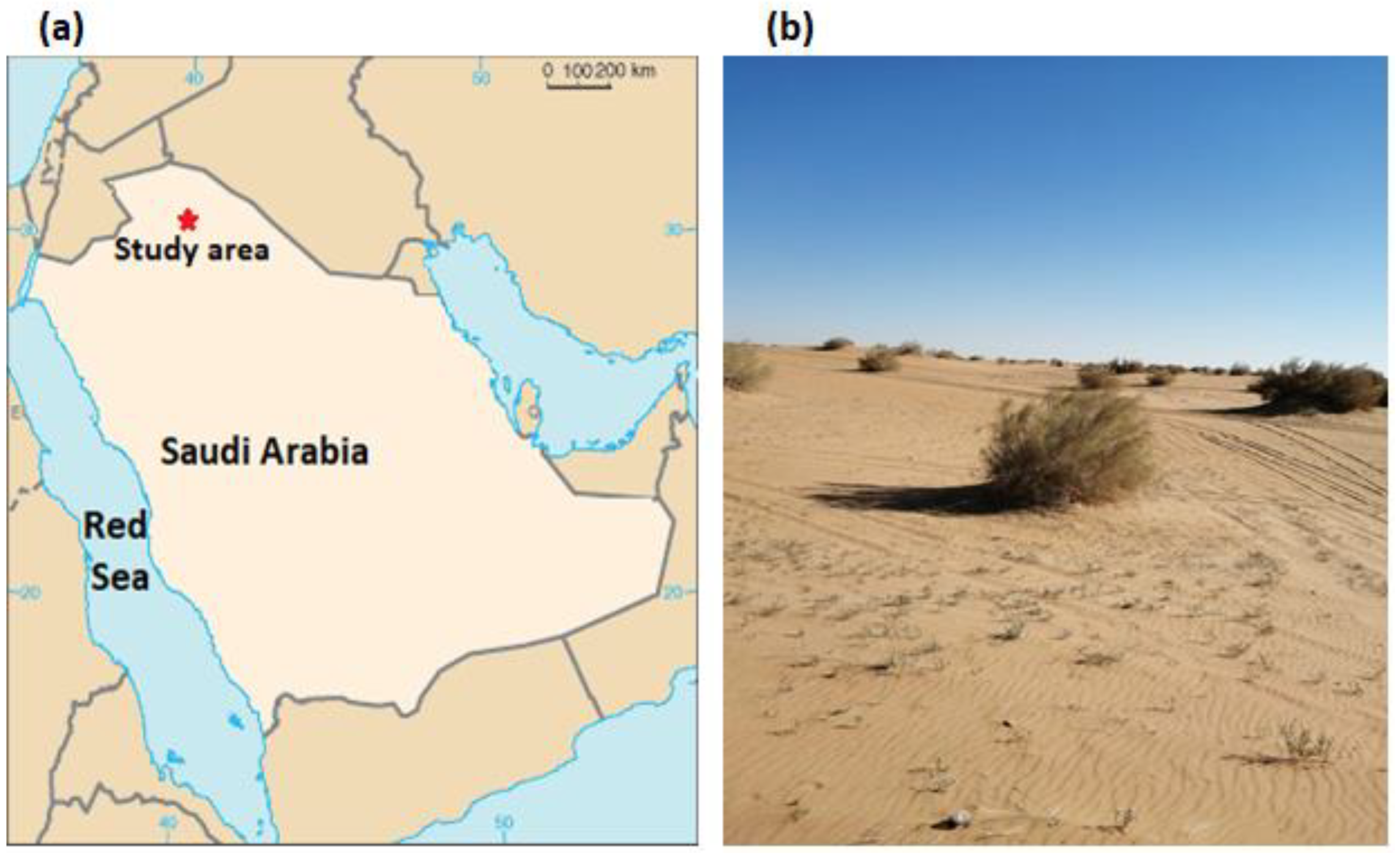
4.1. Study Area
4.2. Shrub Effects on Above-Ground Annual Plants
4.3. Shrub Effects on the Soil Seed Bank of Annual Plants
4.4. Soil Analysis
4.5. Data Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | 2017–2018 | 2018–2019 | ||||
|---|---|---|---|---|---|---|
| Density (Seeds m−2) | RII | Density (Seeds m−2) | RII | |||
| Under Shrub | Bare Soil | Under Shrub | Bare Soil | |||
| Agriophyllum minus Fisch. & C.A.Mey. | 6.7 | 3.3 | 0.40 ns | 14.2 | 6.7 | 0.58 * |
| Aizoon hispanicum L. | 0.8 | 0.0 | – | 0.8 | 0.0 | – |
| Anastatica hierochuntica L. | 0.8 | 0.0 | – | 0.0 | 0.0 | |
| Anisosciadium lanatum Boiss. | 0.0 | 0.0 | 0.8 | 0.0 | – | |
| Anthemis haussknechtii Boiss. & Reut. | 16.7 | 10.8 | 0.21 * | 35.8 | 21.7 | 0.34 * |
| Arnebia decumbens (Vent.) Coss. & Kralik | 1.7 | 0.8 | – | 1.7 | 0.8 | – |
| Asteriscus hierochunticus (Michon) Wiklund | 1.7 | 0.8 | – | 4.2 | 2.5 | 0.57 ns |
| Astragalus arpilobus Kar. & Kir. | 15.0 | 9.2 | 0.36 * | 32.5 | 19.2 | 0.29 * |
| Astragalus asterias Hohen | 7.5 | 3.3 | 0.37 * | 15.8 | 10.0 | 0.43 ns |
| Bassia eriophora (Schrad.) Asch. | 9.2 | 4.2 | 0.53 * | 21.7 | 13.3 | 0.35 * |
| Bassia muricata (L.) Asch. | 31.7 | 20.8 | 0.28 * | 64.2 | 36.7 | 0.32 * |
| Brassica tournefortii Gouan | 10.0 | 6.7 | 0.22 * | 22.5 | 12.5 | 0.49 * |
| Cakile arabica Velen. & Bornm. | 21.7 | 14.2 | 0.23 * | 45.8 | 28.3 | 0.31 * |
| Calendula tripterocarpa Rupr. | 4.1 | 1.7 | 0.42 ns | 7.5 | 5.0 | 0.43 ns |
| Cleome amblyocarpa Barratte & Murb. | 0.0 | 0.0 | 1.7 | 0.0 | – | |
| Cutandia memphitica (Spreng.) K.Richt. | 13.3 | 8.3 | 0.24 * | 26.7 | 16.7 | 0.32 * |
| Diplotaxis acris (Forssk.) Boiss. | 1.7 | 0.0 | – | 6.7 | 1.7 | 0.80 * |
| Eremobium aegyptiacum (Spreng.) Asch. & Schweinf. ex Boiss. | 30.8 | 22.5 | 0.16 * | 65.0 | 38.3 | 0.26 * |
| Erodium laciniatum (Cav.) Willd. | 5.0 | 2.5 | 0.47 ns | 8.3 | 3.3 | 0.53 ns |
| Horwoodia dicksoniae Turrill | 2.5 | 0.8 | 0.67 ns | 3.3 | 0.0 | – |
| Ifloga spicata (Forssk.) Sch. Bip. | 18.3 | 10.8 | 0.39 * | 39.2 | 22.5 | 0.31 * |
| Limonium lobatum (L.f.) Chaz. | 0.8 | 0.8 | – | 0.8 | 0.0 | – |
| Malcolmia grandiflora (Bunge) Kuntze | 4.1 | 1.7 | 0.42 ns | 6.7 | 4.2 | 0.24 ns |
| Malva parviflora L. | 8.3 | 5.0 | 0.33 ns | 15.0 | 9.2 | 0.26 * |
| Matthiola longipetala (Vent.) DC. | 9.2 | 5.0 | 0.41 * | 15.8 | 6.7 | 0.59 * |
| Medicago laciniata (L.) Mill. | 0.8 | 0.0 | – | 0.8 | 0.0 | – |
| Mesembryanthemum nodiflorum L. | 0.0 | 0.0 | 0.8 | 0.0 | – | |
| Neurada procumbens L. | 1.7 | 0.8 | 0.50 ns | 1.7 | 0.8 | – |
| Notoceras bicorne (Aiton) Amo | 1.7 | 0.8 | – | 3.3 | 2.5 | – |
| Oligomeris linifolia (Vahl ex Hornem.) J.F.Macbr. | 1.7 | 0.8 | – | 3.3 | 2.5 | – |
| Opophytum forsskalii (Hochst. ex Boiss.) N.E.Br. | 0.0 | 0.0 | 0.8 | 0.0 | – | |
| Paronychia arabica (L.) DC. | 0.8 | 0.0 | – | 1.7 | 0.8 | 0.50 ns |
| Plantago boissieri Hausskn. & Bornm. | 25.8 | 16.7 | 0.24 * | 50.0 | 33.3 | 0.27 * |
| Plantago amplexicaulis Cav. | 4.2 | 2.5 | 0.33 ns | 8.3 | 3.3 | 0.57 * |
| Plantago ciliata Desf. | 9.2 | 2.5 | 0.75 * | 19.2 | 10.0 | 0.33 * |
| Plantago ovata Forssk. | 10.0 | 4.2 | 0.60 * | 19.2 | 9.2 | 0.55 * |
| Pteranthus dichotomus Forssk. | 0.8 | 0.0 | – | 1.7 | 0.0 | – |
| Rumex vesicarius L. | 5.8 | 0.8 | 0.87 * | 11.7 | 2.5 | 0.74 * |
| Savignya parviflora (Delile) Webb | 10.8 | 7.5 | 0.17 * | 25.0 | 14.2 | 0.43 * |
| Schimpera arabica Hochst. & Steud. ex Steud. | 1.7 | 0.8 | – | 5.0 | 2.5 | 0.71 ns |
| Schismus barbatus (L.) Thell. | 43.3 | 31.7 | 0.18 * | 89.2 | 54.2 | 0.24 * |
| Silene arabica Boiss. | 15.8 | 9.2 | 0.28 * | 35.0 | 20.8 | 0.42 * |
| Spergularia bocconei (Scheele) Graebn. | 0.8 | 0.0 | – | 3.3 | 0.8 | 0.67 ns |
| Trigonella stellata Forssk. | 4.2 | 1.7 | 0.58 ns | 11.7 | 5.0 | 0.51 * |
References
- Baskin, C.; Baskin, J.M. Seeds: Ecology, Biogeography, and Evolution of Dormancy and Germination, 2nd ed.; Academic Press: San Diego, CA, USA, 2014. [Google Scholar]
- King, T.J. The roles of seed mass and persistent seed banks in gap colonisation in grassland. Plant Ecol. 2007, 193, 233–239. [Google Scholar] [CrossRef]
- Ludewig, K.; Hansen, W.; Klinger, Y.P.; Eckstein, R.L.; Otte, A. Seed bank offers potential for active restoration of mountain meadows. Restor. Ecol. 2021, 29, e13311. [Google Scholar] [CrossRef]
- Sotomayor, D.A.; Gutiérrez, J.R. Seed bank of desert annual plants along an aridity gradient in the southern Atacama coastal desert. J. Veg. Sci. 2015, 26, 1148–1158. [Google Scholar] [CrossRef]
- Fenner, M.; Thompson, K. The Ecology of Seeds; Cambridge University Press: Cambridge, UK, 2005. [Google Scholar]
- Yahdjian, L.; Sala, O. Vegetation structure constrains primary production response to water availability in the Patagonian steppe. Ecology 2006, 87, 952–962. [Google Scholar] [CrossRef] [PubMed]
- Ward, D. The Biology of Deserts; Oxford University Press: New York, NY, USA, 2009. [Google Scholar]
- Pake, C.E.; Venable, D.L. Seed banks in desert annuals: Implications for persistence and coexistence in variable environments. Ecology 1996, 77, 1427–1435. [Google Scholar] [CrossRef]
- Honnay, O.; Bossuyt, B.; Jacquemyn, H.; Shimono, A.; Uchiyama, K. Can a seed bank maintain the genetic variation in the above ground plant population? Oikos 2008, 117, 1–5. [Google Scholar] [CrossRef]
- De Villiers, A.J.; Van Rooyen, M.W.; Theron, G.K. Similarity between the soil seed bank and the standing vegetation in the Strandveld Succulent Karoo, South Africa. Land Degrad. Dev. 2003, 14, 527–540. [Google Scholar] [CrossRef]
- Gomaa, N.H. Soil seed bank in different habitats of the Eastern Desert of Egypt. Saudi J. Biol. Sci. 2012, 19, 211–220. [Google Scholar] [CrossRef]
- Gomaa, N.H. Microhabitat variations and seed bank-vegetation relationships in a desert wadi ecosystem. Flora 2014, 209, 725–732. [Google Scholar] [CrossRef]
- Royo, A.A.; Ristau, T.E. Stochastic and deterministic processes regulate spatio-temporal variation in seed bank diversity. J. Veg. Sci. 2013, 24, 724–734. [Google Scholar] [CrossRef]
- Noy-Meir, I. Desert ecosystems: Environment and producers. Annu. Rev. Ecol. Syst. 1973, 4, 25–41. [Google Scholar] [CrossRef]
- Quevedo-Robledo, L.; Pucheta, E.; Ribas-Fern’andez, Y. Influences of interyear rainfall variability and microhabitat on the germinable seed bank of annual plants in a sandy Monte Desert. J. Arid Environ. 2010, 74, 167–172. [Google Scholar] [CrossRef]
- Gomaa, N.H. Reproductive traits, flowering phenology and seed bank dynamics in Retama raetam (Fabaceae) in a hyper-arid environment. Arid Land Res. Manag. 2019, 33, 70–90. [Google Scholar] [CrossRef]
- Aguiar, M.R.; Sala, O.E. Seed distribution constrains the dynamics of the Patagonian steppe. Ecology 1997, 78, 93–100. [Google Scholar] [CrossRef]
- Flores, J.; Jurado, E. Are nurse-protege interactions more common among plants from arid environments? J. Veg. Sci. 2003, 14, 911–916. [Google Scholar] [CrossRef]
- Filazzola, A.; Lortie, C.J. A systematic review and conceptual framework for the mechanistic pathways of nurse plants. Glob. Ecol. Biogeogr. 2014, 23, 1335–1345. [Google Scholar] [CrossRef]
- Holmgren, M.; Scheffer, M.; Huston, M.A. The interplay of facilitation and competition in plant communities. Ecology 1997, 78, 1966–1975. [Google Scholar] [CrossRef]
- Casper, B.B. Demographic consequences of drought in the herbaceous perennial Cryptantha flava: Effects of density, associations with shrubs, and plant size. Oecologia 1996, 106, 144–152. [Google Scholar] [CrossRef]
- Zhang, G.; Zhao, W.; Zhou, H.; Yang, Q.; Wang, X. Extreme drought stress shifts net facilitation to neutral interactions between shrubs and sub-canopy plants in an arid desert. Oikos 2018, 127, 381–391. [Google Scholar] [CrossRef]
- Gomaa, N.H.; Hegazy, A.K.; Latef, A.A.H.A. Facilitation effects of Haloxylon salicornicum shrubs on associated understory annuals, and a modified “Stress-Gradient” hypothesis for droughty times. Plants 2020, 9, 1726. [Google Scholar] [CrossRef]
- O’Brien, M.J.; Pugnaire, F.I.; Armas, C.; Rodríguez-Echeverría, S.; Schöb, C. The shift from plant–plant facilitation to competition under severe water deficit is spatially explicit. Ecol. Evol. 2017, 7, 2441–2448. [Google Scholar] [CrossRef] [PubMed]
- Kemp, P.R. Seed banks and vegetation processes in deserts. In Ecology of Soil Seed Banks; Leck, M.A., Parker, V.T., Simpson, R.L., Eds.; Academic Press: San Diego, CA, USA, 1989; pp. 257–281. [Google Scholar]
- Pugnaire, F.; Lázaro, R. Seed Bank and Understorey Species Composition in a Semi-arid Environment: The Effect of Shrub Age and Rainfall. Ann. Bot. 2000, 86, 807–813. [Google Scholar] [CrossRef]
- Giladi, I.; Segoli, M.; Ungar, E.D. Shrubs and herbaceous seed flow in a semi-arid landscape: Dual functioning of shrubs as trap and barrier. J. Ecol. 2013, 101, 97–106. [Google Scholar] [CrossRef]
- Debussche, M.; Isenmann, P. Bird-dispersed seed rain and seedling establishment in patchy Mediterranean vegetation. Oikos 1994, 69, 414–426. [Google Scholar] [CrossRef]
- Beck, M.J.; Vander Wall, S.B. Seed dispersal by scatter-hoarding rodents in arid environments. J. Ecol. 2010, 98, 1300–1309. [Google Scholar] [CrossRef]
- Varela, O.; Ordano, M.; Toledo, G.; Lizardo, G.; Rotger, S.; Montero, A.; Cisneros, M.C. Diversity and density of the desert seed bank: Interplays between cacti and nurse shrub species. J. Arid Environ. 2021, 191, 104536. [Google Scholar] [CrossRef]
- Tielbörger, K.; Kadmon, R. Temporal environmental variation tips the balance between facilitation and interference in desert plants. Ecology 2000, 81, 1544–1553. [Google Scholar] [CrossRef]
- Gutiérrez, J.R.; Meserve, P.L. El Niño effects on soil seed bank dynamics in north-central Chile. Oecologia 2003, 134, 511–517. [Google Scholar] [CrossRef]
- Tessema, Z.K.; Ejigu, B.; Nigatu, L. Tree species determine soil seed bank composition and its similarity with understory vegetation in a semi-arid African savanna. Ecol. Processes 2017, 6, 1–9. [Google Scholar] [CrossRef]
- Spiegel, O.; Nathan, R. Incorporating dispersal distance into the disperser effectiveness framework: Frugivorous birds provide complementary dispersal to plants in a patchy environment. Ecol. Lett. 2007, 10, 718–728. [Google Scholar] [CrossRef]
- Wezel, A.; Rajot, J.L.; Herbrig, C. Influence of shrubs on soil characteristics and their function in Sahelian agro-ecosystems in semi-arid Niger. J. Arid Environ. 2000, 44, 383–398. [Google Scholar] [CrossRef]
- Batanouny, K.H.; Zayed, K.M.; Emad, H.M. Ecology of the soil seed bank in sandstone ecosystem within Cairo-Suez desert, Egypt. Saudi J. Biol. Sci. 2000, 7, 46–63. [Google Scholar]
- Li, Y.J.; Bao, W.K.; Wu, N. Spatial patterns of the soil seed bank and extant vegetation across the dry Minjiang river valley in Southwest China. J. Arid Environ. 2011, 75, 1083–1089. [Google Scholar] [CrossRef]
- El-Keblawy, A.; Abdelfatah, M. Impacts of native and invasive exotic Prosopis congeners on soil and associated flora in the arid United Arab Emirates. J. Arid Environ. 2014, 100–101, 1–8. [Google Scholar] [CrossRef]
- Zhang, Z.; Li, X.; Dong, X.; Jia, X.; He, M.; Tan, H. Rainfall interception by sand-stabilizing shrubs related to crown structure. Sci. Cold Arid Reg. 2009, 1, 107–119. [Google Scholar]
- Padilla, F.M.; Pugnaire, F.I. The role of nurse plants in the restoration of degraded environments. Front. Ecol. Environ. 2006, 4, 196–202. [Google Scholar] [CrossRef]
- García, E. Efecto de la Estructura de la Copa en la Partición de Lluvia de Tres Especies Arbustivas en Clima Smiárido. Ph.D. Thesis, University of Almería, Almería, Spain, 2006. [Google Scholar]
- Peco, B.; Ortego, M.; Levassor, C. Similarity between seed bank and vegetationin Mediterranean grassland: A predictive model. J. Veg. Sci. 1998, 9, 815–828. [Google Scholar] [CrossRef]
- Li, N.; Feng, G.; Tian, C. Characteristics and dynamics of the soil seed bank at the north edge of Taklimakan Desert. Sci. China Ser. D Earth Sci. 2007, 50, 122–127. [Google Scholar] [CrossRef]
- Wu, T.; Wang, X.Q.; Gai, S.G. Effect of Grazing in Spring-Summer on Soil Seed Bank and Vegetation in Southern Part of Gurbantunggut Desert. J. Desert Res. 2009, 29, 499–507. [Google Scholar]
- Li, F.R.; Zhao, L.Y.; Wang, S.F. Effects of enclosure management on the structure of soil seed bank and standing vegetation in degraded sandy grasslands of eastern Inner Mongolia. Acta Pratacult. Sin. 2003, 12, 90–99. [Google Scholar]
- Rathore, V.S.; Singh, J.P.; Bhardwaj, S.; Nathawat, N.S.; Kumar, M.; Roy, M.M. Potential of native shrubs Haloxylon salicornicum and Calligonum polygonoides for restoration of degraded lands in Arid Western Rajasthan, India. Environ. Manag. 2015, 55, 205–216. [Google Scholar] [CrossRef]
- Hegazy, A.K.; Lovett-Doust, J. Plant Ecology in the Middle East; Oxford University Press: Oxford, UK, 2016. [Google Scholar]
- Song, J.I.E.; Feng, G.U.; Tian, C.; Zhang, F. Strategies for adaptation of Suaeda physophora, Haloxylon ammodendron and Haloxylon persicum to a saline environment during seed-germination stage. Ann. Bot. 2005, 96, 399–405. [Google Scholar] [CrossRef] [PubMed]
- Al-khalifah, N.S.; Shanavaskhan, A.E. On the distribution, status and phenology of Ghada (Haloxylon persicum Bunge) in the Arabian Peninsula. Trop. Ecol. 2007, 48, 51–60. [Google Scholar]
- Casati, P.; Andrew, C.S.; Edward, G.E. Characterization of NADP-malic enzyme from two species of Chenopodiaceae: Haloxylon persicum (C4) and Chenopodium album (C3). Phytochemistry 1999, 52, 985–992. [Google Scholar] [CrossRef]
- Orlovsky, N.; Birnbaum, E. The role of Haloxylon species for combating desertification in Central Asia. Plant Biosyst. 2002, 136, 233–240. [Google Scholar] [CrossRef]
- El-Sheikh, M.A.; Abbadi, G.A.; Bianco, P.M. Vegetation ecology of phytogenic hillocks (nabkhas) in coastal habitats of Jal Az-Zor National Park, Kuwait: Role of patches and edaphic factors. Flora 2010, 205, 832–840. [Google Scholar] [CrossRef]
- Alghanem, S.M. Ecological and botanical diversity in Haloxylon persicum community at Al-Qassim Region in Kingdom of Saudi Arabia. Am. J. Environ. Prot. 2018, 6, 43–49. [Google Scholar] [CrossRef]
- Chaudhary, S.A. Flora of the Kingdom of the Saudi Arabia; Ministry of Agriculture and Water: Riyadh, Saudi Arabia, 1999; Volume I.
- Chaudhary, S.A. Flora of the Kingdom of the Saudi Arabia; Ministry of Agriculture and Water: Riyadh, Saudi Arabia, 2000; Volume II.
- Chaudhary, S.A. Flora of the Kingdom of the Saudi Arabia; Ministry of Agriculture and Water: Riyadh, Saudi Arabia, 2001; Volume III.
- Black, C.A. Methods of Soil Analysis; American Society of Agronomy: Madison, WI, USA, 1965; Part 2. [Google Scholar]
- Gupta, P.K. Soil, Plant, Water and Fertilizer Analysis; Agro-Botanica Publisher: Bikaner, India, 1999. [Google Scholar]
- Armas, C.; Ordinales, R.; Pugnaire, F.I. Measuring plant interactions: A new comparative index. Ecology 2004, 85, 2682–2686. [Google Scholar] [CrossRef]





| Factor | Seed Density | Species Richness |
|---|---|---|
| Shrub | 134.563 *** | 87.870 *** |
| Growing season | 219.466 *** | 34.020 *** |
| Shrub × growing season | 19.624 *** | 2.777 ns |
| Growing Season | Seed Density | Species Richness |
|---|---|---|
| 2017–2018 | 0.26 ± 0.02 *b | 0.12 ± 0.03 *b |
| 2018–2019 | 0.28 ± 0.02 *a | 0.14 ± 0.03 *a |
| Factor | Species Richness | Seed Bank–Vegetation Similarity |
|---|---|---|
| Shrub | 18.821 *** | 20.102 *** |
| Growing season | 692.707 *** | 387.073 *** |
| Shrub × growing season | 147.559 *** | 71.490 *** |
| Parameter | Shrub | Growing Season | Shrub × Growing Season |
|---|---|---|---|
| Organic carbon (%) | 18.160 *** | 0.077 ns | 0.009 ns |
| Moisture content (%) | 7.430 ** | 1867.863 *** | 26.047 *** |
| Electrical conductivity (dS/m) | 155.033 *** | 1.948 ns | 0.007 ns |
| Sand | 634.707 *** | 0.036 ns | 0.024 ns |
| Silt + clay | 634.707 *** | 0.036 ns | 0.024 ns |
| N (mg/kg) | 33.869 *** | 0.006 ns | 0.024 ns |
| P (mg/kg) | 815.276 *** | 0.002 ns | 0.143 ns |
| K (mg/kg) | 242.818 *** | 0.005 ns | 0.053 ns |
| Soil Variable | 2017–2018 | 2018–2019 | ||
|---|---|---|---|---|
| Under Shrub | Bare Soil | Under Shrub | Bare Soil | |
| Organic carbon (%) | 0.31 ± 0.08 a | 0.22 ± 0.08 b | 0.30 ± 0.08 a | 0.21 ± 0.08 b |
| Moisture content (%) | 1.15 ± 0.16 b | 1.31 ± 0.18 a | 4.04 ± 0.32 a | 3.87 ± 0.32 b |
| Electrical conductivity (dS/m) | 0.73 ± 0.04 a | 0.55 ± 0.07 b | 0.71 ± 0.04 a | 0.53 ± 0.06 b |
| Sand (%) | 84.4 ± 1.0 b | 92.1 ± 0.6 a | 84.1 ± 1.6 b | 91.8 ± 1.2 a |
| Silt + clay (%) | 15.6 ± 1.0 a | 7.9 ± 0.6 b | 15.9 ± 1.6 a | 8.2 ± 1.2 b |
| N (mg/kg) | 102.7 ± 8.1 a | 90.9 ± 7.7 b | 103.2 ± 7.0 a | 91.4 ± 6.7 b |
| P (mg/kg) | 2.49 ± 0.11 a | 1.35 ± 0.16 b | 2.47 ± 0.12 a | 1.37 ± 0.15 b |
| K (mg/kg) | 117.3 ± 7.6 a | 80.4 ± 9.9 b | 118.5 ± 6.0 a | 79.7 ± 9.8 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gomaa, N.H.; Hegazy, A.K.; Alhaithloul, H.A.S. Facilitation by Haloxylon persicum Shrubs Enhances Density and Richness of Soil Seed Bank of Annual Plants in a Hyper-Arid Ecosystem. Plants 2023, 12, 1276. https://doi.org/10.3390/plants12061276
Gomaa NH, Hegazy AK, Alhaithloul HAS. Facilitation by Haloxylon persicum Shrubs Enhances Density and Richness of Soil Seed Bank of Annual Plants in a Hyper-Arid Ecosystem. Plants. 2023; 12(6):1276. https://doi.org/10.3390/plants12061276
Chicago/Turabian StyleGomaa, Nasr H., Ahmad K. Hegazy, and Haifa Abdulaziz S. Alhaithloul. 2023. "Facilitation by Haloxylon persicum Shrubs Enhances Density and Richness of Soil Seed Bank of Annual Plants in a Hyper-Arid Ecosystem" Plants 12, no. 6: 1276. https://doi.org/10.3390/plants12061276
APA StyleGomaa, N. H., Hegazy, A. K., & Alhaithloul, H. A. S. (2023). Facilitation by Haloxylon persicum Shrubs Enhances Density and Richness of Soil Seed Bank of Annual Plants in a Hyper-Arid Ecosystem. Plants, 12(6), 1276. https://doi.org/10.3390/plants12061276