Abstract
Satureja pilosa Velen., S. coerulea Janka and S. kitaibelii Wierzb. ex Heuff. are Balkan endemic species, and they are distributed in restricted territories, primarily found in dry grasslands, rocky slopes, and stony habitats. This study presents the results of the first embryological and micromorphological analyses of three Satureja species (S. pilosa, S. kitaibelii, and S. coerulea) from the Bulgarian flora. The aim of this study was to establish the features of the male and female reproductive sphere, as well as surface characteristics of leaves, stem, and calyx in order to understand the mode of reproduction, character, size and state of species populations and delimitation. For the embryological study, flowers and flower buds in different developmental stages were collected from plants of natural populations and treated with the classic paraffin method. Reproductive capacity was assessed using the following approaches: (1) acetocarmine test for pollen viability, (2) tetrazolium test (TTZ) for seed viability, and (3) germination test. The surfaces characteristics of leaves, stem, and calyx were analyzed by scanning electron microscopy (SEM). As a result, the study revealed the structures of the male (M) and female (F) generative spheres as well as the processes of gamete development, pollination, and endosperm and embryo formation. It was found that the three Satureja species exhibited a high pollen viability but low seed viability and germination. The SEM analysis showed both common and distinct micromorphology features regarding epidermis, calyx and stem surfaces among the three Satureja species. Notably, the S. coerulea surfaces (leaves, calyx, stem) were clearly distinguishable from the other two Satureja species. Regarding the nutlet surfaces, it was observed that the exocarp surfaces of S. coerulea and S. kitaibelii have a reticulate convex type surface and comprise two types of cells: (1) highly bulging, bubble-like cells; and (2) flat cells with numerous striations. On the other hand, the exocarp of S. pilosa displayed rectangular or polygonal shapes without bubble-like cells, and it had a tabular to slightly convex type surface. Additionally, nutlets (seeds) of both S. coerulea and S. pilosa exhibited distinct papilla formations resembling non-glandular trichomes seen on the ends of the nutlets for S. coerulea and over the entire surface for S. pilosa. The characteristics of the embryological structures and processes, along with the absence of apomixis, characterized the three studied Satureja species as sexually reproducing. The established balanced processes and stable structures contribute to their high reproductive potential and population stability. However, these traits may also decrease their adaptability to environmental changes.
Keywords:
pollen; embryology; nutlets; leaves; surfaces; germinations; male and female gametophyte; embryo; endosperm 1. Introduction
Satureja is one of the largest genera in the Lamiaceae family [1]. The different Satureja plants are commonly often used as culinary herbs, and they are widely cultivated [2,3]. Most Satureja species have been studied for their chemical composition and biological activities [4,5,6,7]. The species of this genus are distributed in various regions of the world, and most of the species are found in the Mediterranean region and surrounding areas [8]. According to Flora Europaea, there are 12 naturally distributed Satureja species across Europe, and five of them are found in Bulgaria [9].
Satureja pilosa, S. coerulea and S. kitaibelii are Balkan endemic species. They are naturally distributed in the Bulgarian flora in dry grasslands, rocky slopes, and stony habitats under extreme conditions [6,7,9]. The Satureja species have multiple mechanisms to adapt and survive in these unfavorable environmental conditions. These mechanisms include dry tolerance (small leaves with a thick, waxy cuticle), cold tolerance (high-altitude habitats), allelopathy (through the production of allelopathic compounds that inhibit the growth of competing plant species) [10], chemical defense (through the production of essential oil (EO) containing antimicrobial, antifungal, and insecticidal compounds) [6,7], and reproductive strategies. Due to the increased interest in these species and their limited distribution, it is important to implement measures to protect their gene pool and ensure their long-term survival. Generally, endemic species are only found in a restricted geographic location and are often of high conservation significance [11]. Ex situ, in situ collections and biotechnological tools are important approaches for conservation of endemic plants for several reasons: (1) they can help to preserve plant diversity by maintaining populations; (2) they are a source of plant material for research; (3) help with the development of new crops; (4) preserve the genetic diversity of endemic species, etc. [11]. Reproduction plays a key role in plant species and population biology, as well as in the perpetuation of the species [12,13]. The successful cultivation and preservation of rare and endemic plant species requires comprehensive information on their reproductive strategy [14,15,16]. General knowledge on the reproductive strategy of threatened and endemic plants plays a key role not only in systematic studies and evolution [17] but also in the development of effective conservation strategies [18].
The review of the literature sources showed limited data on the embryological structures and processes, type of reproduction and reproductive capacity of species in the genus Satureja. Only the flower structure (pollination system and self-incompatibility) in S. sahendica Bornm. and S. bachtiarica Bunge have been reported [19]. On the other hand, a number of other species in the Lamiaceae family have been subjects of embryological studies [20,21,22,23]. The embryological features of Nepeta cataria L., Salvia nemorosa L., Lamium maculatum L., L. garganicum L., and L. bifidum Cirillo are well-documented [20,21,22,24]. These studies reported that anthers were tetrasporangiate and four-layered; tapetum was of the secretory type; and meiosis in the anthers was of simultaneous type. Furthermore, the cited authors established three-celled hexacolpate pollen, which is typical for the Lamiaceae species [20,21,22,24]. Overall, the embryological features of plants provide valuable information about the plants’ reproductive biology, including aspects such as pollen development, fertilization, and seed formation.
Scanning electron microscopy (SEM) analyses, on the other hand, provide a detailed view of the surface morphology of various plant parts (leaves, stems, flowers, pollen, seeds). SEM is a powerful tool for describing the surface morphology, microstructure and subcellular ultrastructure of plants [25]. This information can be useful in identifying different species of Satureja plants, as well as understanding their adaptations to different environmental conditions. Several papers reported SEM analyses of Satureja species, mainly for S. hortensis L., and S. montana L. [26,27,28,29]. Indeed, these Satureja species are widely cultivated for their EO, which is synthesized and accumulated in glandular trichomes [6,30]. Furthermore, SEM analyses could provide useful information in studying the surfaces and morphology of leaves, nutlets (seeds) and pollen, especially for Satureja species where there are several unresolved taxonomic questions [31]. These issues were a result of difficulties in species delimitation for the following reasons: (1) Satureja species can easily hybridize within the same population as well between different Satureja species [32]; (2) subsequently, there is a high degree of morphological variability and overlapping characteristics within and among different species because of hybridization [31,32,33]; (3) there are geographical variations in morphology [32]. Comparative SEM analyses of the surface features of nutlets (seeds), leaves, and pollen in the three Balkan endemic species S. pilosa, S. coerulea and S. kitaibelii distributed in the Bulgarian flora have not been reported. The objectives of this study were (1) to establish the features of male and female reproductive spheres of S. pilosa, S. coerulea and S. kitaibelii in connection with revealing the mode of reproduction, character, size and state of their populations; and (2) to analyze and compare surfaces of nutlets (seeds), leaves, calyx and stem in order to provide new data for species knowledge and delimitation.
2. Results
2.1. Embryological Analyses
The embryological characteristics observed in the study were consistent with those described for the Lamiaceae, and were found to be largely similar across the studied species. Therefore, the images and description of results from this study are common for S. pilosa, S. coerulea and S. kitaibelii.
2.1.1. Anther and Development of the Male Gametophyte
The anther was four-locular (Figure 1A). The anther wall was four-layered, arranged according to the dicotyledonous type of the Davis classification [34]. The four-layered walls consisted of epidermis, endothecium, middle layer and secretory tapetum. The epidermis was represented by single row of one-nucleate cells. Their forms were rectangular, and during anther ontogenesis, they enlarged tangentially and rounded up outside. The middle layer was one-rowed. The shapes and sizes of cells of the middle layer were heterogeneous. This layer degenerated about the homotypic division of meiosis in microspore mother cells (MMCs). At the stage of one-nucleate pollen (Figure 1D), the endothecium developed fibrous thickenings. Initially, the tapetum was glandular with one-nucleate cells. During anther ontogenesis, highly elongated tapetum cells underwent rapid multiplication of nuclei due to successive mitotic divisions without subsequent cytokinesis. After the formation of unicellular pollen, the tapetum cells enlarged and intruded into the anther locules and formed “placentoids” (Figure 1D). This layer remained cellular until the maturity of the anther. The sporogenous tissue was one-rowed (Figure 1B). The primary sporogenous cells functioned directly as MMCs. After meiosis and simultaneous microsporogenesis consecutively running in them predominantly, tetrahedral tetrads formed (Figure 1C). The mature pollen grains were two-celled (Figure 1E).
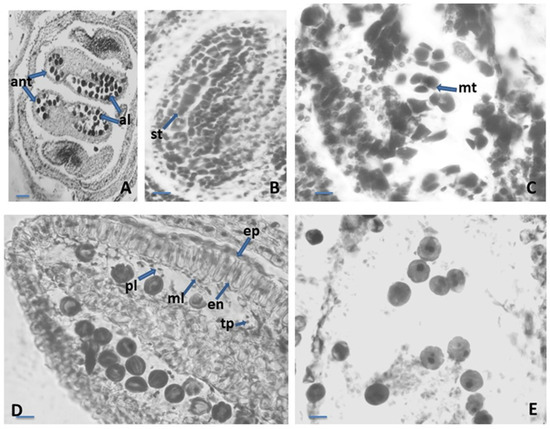
Figure 1.
Anther and development of the male gametophyte. (A)—tetrasporangiate anther; (B)—sporogenous tissue in the anther locule; (C)—microspore tetrads in the anther locule; (D)—one-nucleate pollen grains and anther wall with epidermis, endothecium, middle layer and tapetum forming placentoids; (E)—two-celled mature pollen grains, ant—anther, al—anther locule, st—sporogenous tissue, mt—microspore tetrad, ep—epidermis, en—endothecium, ml—middle layer, tp- tapetum, pl—placentoid. Scale bar: for (A) = 50 μm; for (B–E) = 20 μm.
2.1.2. Ovule and Development of the Female Gametophyte
The ovary in the studied species was syncarpous, superior, and four locular (Figure 2A). Each locule consisted of a single anatropous ovule, with tenuinucellate unitegument on axile placentation (Figure 2B,C). In the early stages of ovule development, a unicellular archesporium formed below the surface, directly functioning as the microspore mother cell. (MMC). The archesporogenesis proceeded without cover cell cutting. As a result of macrosporogenesis in MMC, a linear macrospore tetrad was formed (Figure 2D). We observed that the embryo sac started its development of the micropylar megaspore, besides the chalazal megaspore, and one embryo sac in the ovule locule was observed.

Figure 2.
Ovule and development of the female gametophyte. (A)—Structure of the Satureja flowers; (B)–Ovary locules with anatropous unitegmic ovule in each locule; (C)—Anatropous unitegmic ovule; (D)—Linear macrospore tetrad in the ES cavity with macrospore mother cell; ov—ovary, ant—anther locule, ol—ovary locule, mt—macrospore tetrad, mmc—macrospore mother cell. Scale bar: for (A) = 50 μm; for (B–D) = 20 μm.
The female gametophyte in the studied Satureja species followed the basal Polygonum type of development (Figure 3A). The mature embryo sac (ES) is typical for this type of structure and arrangement of its elements. The arrangement of elements was as follows: (1) egg apparatus composed from an egg cell and two hook synergids with filiform apparatus at the micropylar end of ES; (2) two polar nuclei situated near the egg apparatus, below the egg cell; and (3) three antipodals at the chalazal end of the ES (Figure 3A). The embryo and endosperm started their development after double porogamous fertilization. The endosperm was cellular but initially passed through a free nuclear stage. Embryogenesis began after endosperm genesis and can be defined as the Onagrad type according to Johansen’s classification of embryogenesis [35,36]. Apomixis was not observed. Proterandry was found to take place in flower development: when mature pollen is formed in the anthers, a tetrad megaspore is formed in the ovule (Figure 3C).
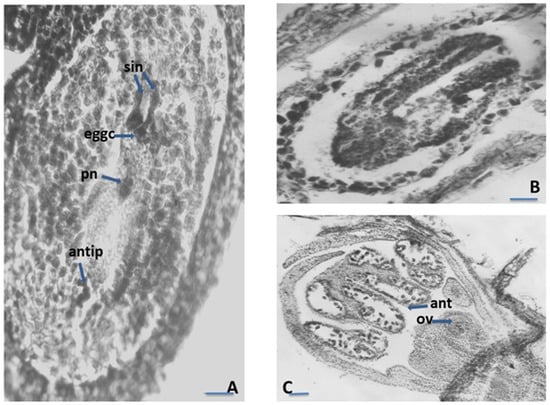
Figure 3.
Ovule and development of the female gametophyte. (A)—Mature Polygonum-type embryo sac (ES); (B)—Embryo with cotyledons in ES cavity; (C)—Proterandry in the Satureja flower; syn—synergids, eggc—egg cell, pn—polar nucleus cell, antip—antipodal, ov—ovule, ant—anther locule; Scale bar: for (A,B) = 20 μm; for (C) = 50 μm.
2.1.3. Nutlets (Seed) and Pollen Viability Testing
According to the requirements of tetrazolium methods (TTZ test) [37,38], the seeds of S. coerulea, S. kitaibelii and S. pilosa were divided into six classes (Figure 4). The first class (Class I) included the seeds with embryos stained in dark red (Figure 4A). The second class (Class II) included the seeds with pink colored embryos (Figure 4B). The third class (Class III) included the seeds with embryos in which only the root was stained in red (Figure 4C). The fourth class (Class IV) included the seeds with colorless embryos (Figure 4D). The fifth class (Class V) comprised the empty seeds (Figure 4E). The sixth class (Class VI) included the seeds with undeveloped (dried) embryos (Figure 4F). According to Moore’s classification [39], the seeds in Classes I, II and III were considered as viable, and those in Classes IV, V and VI were considered unviable. Therefore, the amount of viable seeds (embryos) estimated in % was as follows: S. pilosa: 31.18%, S. coerulea: 39.58%, S. kitaibelii: 10.58% (Figure 5).
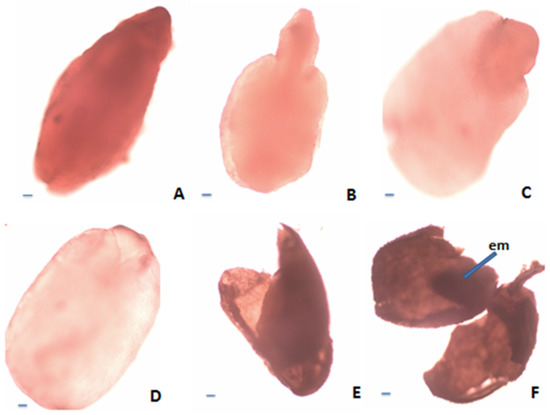
Figure 4.
Estimation of nutlet (seed) viability according to tetrazolium test: (A)—Viable embryo (stained in dark red); (B)—Viable embryo (stained in pink); (C)—Viable embryo (only the root stained in red); (D)—Nonviable embryo (unstained); (E)—Empty seed; (F)—Seed with undeveloped embryo. Scale bar = 100 μm.
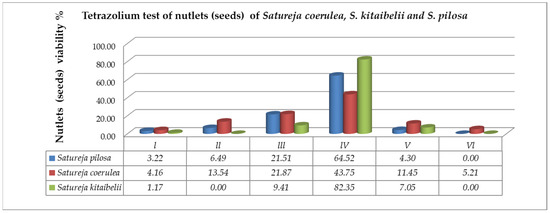
Figure 5.
Evaluation of nutlet (seed) viability in classes after TZ testing on the studied Satureja species: Class I—seeds with embryos stained in dark red; Class II—seeds with pink colored embryos; Class III—seeds with embryos in which only the root is stained in red; Class IV—seeds with colorless embryos; Class V—empty seeds; Class VI—seeds with undeveloped (dried) embryos.
After acetocarmine staining, the cytoplasm and nuclei of viable pollen grains were stained in red, while unviable, empty and shrunken pollen grains remained unstained (Figure 6). The results of the study showed high viability of the mature pollen: 90.01 ± 3.35 for S. pilosa, 94.14 ± 3.82 for S. coerulea, and 93.44 ± 3.88% for S. kitaibelii.
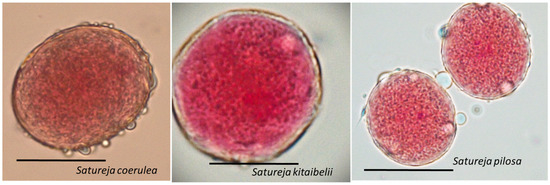
Figure 6.
Viable pollen of Satureja coerulea, S. kitaibelii and S. pilosa. The photos were taken with LM Motic DMA (×40); Scale bar = 100 µm.
2.1.4. Nutlet (Seed) Germination
Satureja kitaibelii seeds were not tested for germination due to insufficient seeds for three replicates because most of the seeds of this species were empty and non-viable. Overall, the seeds of S. coerulea and S. pilosa were tested for germination energy (GE) and germination (G). The results are presented in Table 1. Low germination energy and germination rates (below 50%) were observed in both species—S. coerulea and S. pilosa. Both indicators (GE and G) were the lowest in variant 2 (fluorescent white light with addition of LED red and blue in ratio of 7:1) for S. pilosa (9 and 18%). The highest GE values were observed for variant 3 (fluorescent white light with addition of LED red and blue in ratio of 4:1) in S. coerulea (31%). Natural light (V 1) was found to be the most suitable for seed germination (G%) compared to applied artificial light combinations in both tested species.

Table 1.
Germination energy (GE%) and germination (G%) of Satureja coerulea and Satureja pilosa.
2.2. Scanning Electron Microscopy (SEM) Analyses
2.2.1. Leaves, Stem, and Calyx Surfaces
Overall, all studied Satureja species parts (leaves, stem, calyx, corolla) were covered with trichomes. The indumentum of this species presented glandular and non-glandular trichomes.
Satureja coerulea
The micromorphological characteristics of S. coerulea (leaves, calyx, and stem) are presented in Figure 7A–F, Figure 8A–D and Figure 9A–C. The leaves are small and smooth. The SEM analyses showed that the main epidermal cells are irregular to rectangular in shape, with slightly raised to smooth periclinal walls. The stomata are elliptical and submerged in the epidermis, with distinct cuticles and scattered epicuticular wax plates (Figure 7A,E). Both non-glandular and glandular trichomes are present on the leaf surface, with the latter being peltate and located on the abaxial surface (Figure 7A–D). Stereo microscope analyses showed that the calyx is short, with five shallowly incised teeth, ten veins, and anthocyanin coloring (Figure 8). The calyx surfaces have highly undulating epidermis surfaces with convex periclinal walls, and both non-glandular and glandular trichomes are distributed over the entire surface (Figure 8A–D). The stem surface is smooth to raised with striations and covered with unicellular to multicellular, unbranched non-glandular trichomes and epicuticular wax plates (Figure 9A–C).
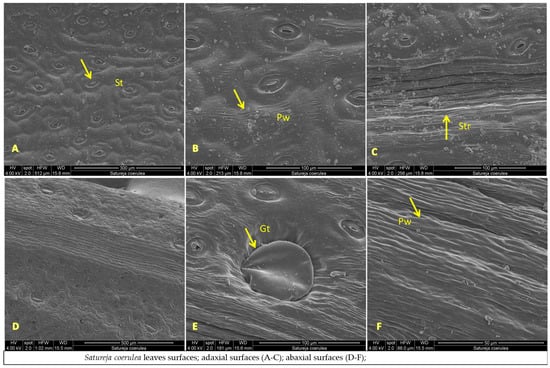
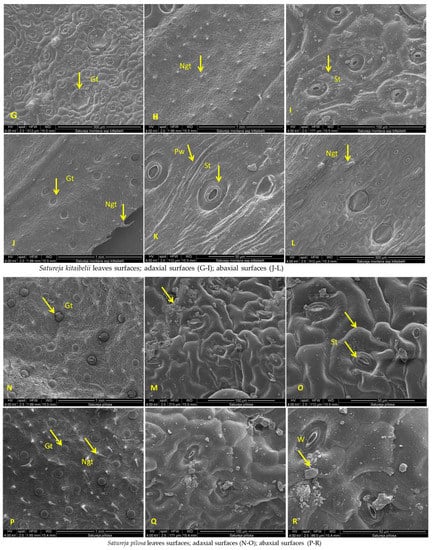
Figure 7.
SEM analysis of leaf surfaces of Satureja coerulea (A–F), S. kitaibelii (G–L), S. pilosa (M–R); St—stomata; Gt—glandular trichomes; Ngt—non-glandular trichomes; W—waxes; Pw—periclinal wall; Str—striation.

Figure 8.
SEM analysis of calyx surfaces of Satureja coerulea (A–C), S. kitaibelii (D–F), S. pilosa (G–I). Scale bar: for the calyx LM = 100 µm. The photos were taken with Stereo Microscope Motic DM 143.
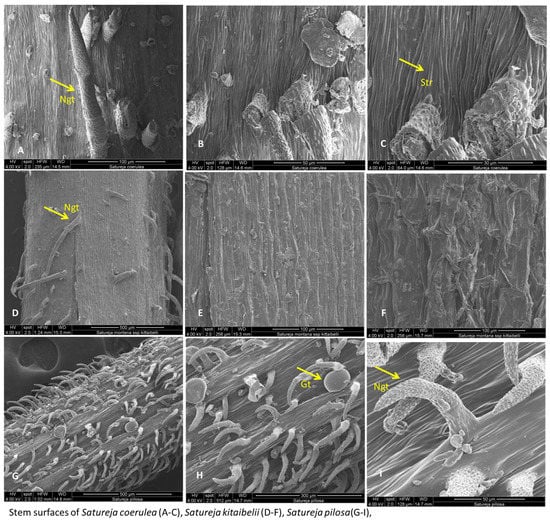
Figure 9.
SEM analysis of stem surfaces of Satureja coerulea (A–C), S. kitaibelii (D–F), S. pilosa (G–I). Gt—glandular trichomes; Ngt—non-glandular trichomes; Str- striation.
Satureja kitaibelii
The SEM analyses of S. kitaibelii revealed that the epidermal surfaces of its leaves are smooth to slightly convex with striation (Figure 7G–I). The main epidermal cells are isodiametric, with a rounded edge and straight anticlinal walls. The waxes are continuous layers with single crusts of varying thicknesses (Figure 7G–I). The stomata are located on both surfaces, and they are mostly diacytic. The cells of the stomatal complex of S. kitaibelii have specific striations (Figure 7I,K). The guard cells are round shaped and small. The observed indumentum includes both non-glandular and glandular peltate-type trichomes on both surfaces of leaves (Figure 7G–L). The non-glandular trichomes are of two types: multicellular and unbranched along the edge of leaves (Figure 7J), and unicellular and uniseriate as papilla (Figure 7G,H,L). The stereo microscope analyses showed that the calyx of S. kitaibelii is cylindrical, slightly tubular, 6–7 mm long, with 10 veins and anthocyanin coloring (Figure 8). The calyx surfaces are highly wrinkled, with multiple non-glandular and glandular capitate- and peltate-type trichomes (Figure 8E–H). The stem surfaces are covered with waxes and two types of non-glandular trichomes, namely unicellular conical curved and multicellular unbranched (Figure 9D–F).
Satureja pilosa
The leaves of S. pilosa are oblong-lanceolate, acuminate. SEM analyses of the leaf surfaces showed that the main epidermal cells are isodiametric with curve anticlinal walls (Figure 7). The edges of the anticlinal walls are oval to point, and the periclinal walls are striated (Figure 7N–R). The stomata are oval shapes with a double rim of waxes at the level of the epidermis (Figure 7O,Q,R). The stomata are in both surfaces (ad, ab), and leaves are amphistomatic (Figure 7). Overall, the whole plant is covered with non-glandular simple conical trichomes and glandular peltate- or capitate-type trichomes (Figure 7 and Figure 8). The wax crusts are observed over the entire surface. The sepals are cylindrical and elongated (5–6 mm), and the teeth are triangularly incised and covered with multicellular simple, conical hairs and peltate-type glandular trichomes (Figure 8I–K). The stem is the most hairy, with longitudinal striations (Figure 9G–I). The surface of the epidermal cells of the stem is ribbed, with indented and slightly convex ribs covered with a waxy film. Wax plates are observed over the entire surface of the stem. The non-glandular hairs are unicellular or multicellular (mostly four-cellular), conical, with papillose surfaces. The glandular trichomes are of the capitate type with a short stalk and a large head.
2.2.2. Nutlet (Seed) Surfaces
The seed surfaces and morphology of three Satureja species were analyzed with stereo microscope and SEM analyses. The pictures are presented in Figure 10 and Figure 11.

Figure 10.
Nutlets (seeds) of Satureja species viewed with Stereo Microscope Motic DM 143. Satureja coerulea (1A–1D): (A)—view of nutlets from all sides, top, lateral, and tip; (B–D)—papillae; (C)—unevenly triangular tip; (D)—adaxial surfaces, papillae; Satureja kitaibelii (2A–2D): A—view of nutlets from all sides top, lateral, and tip; (B–D)—lateral view, the triangular tip; Satureja pilosa 10; (3A–3D): A—view of nutlets from all sides top, lateral, and tip; (B–D)—papillae, unevenly triangular tip. Scale bar = 100 µm.
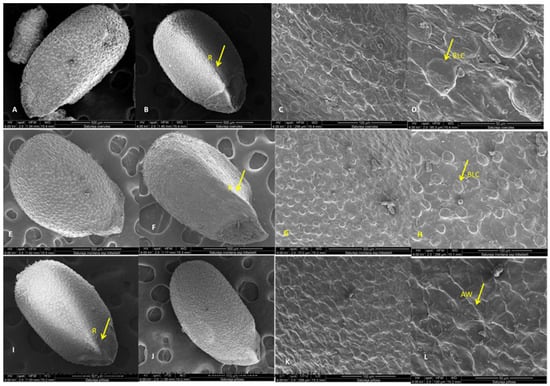
Figure 11.
SEM analysis of nutlet surfaces of Satureja coerulea, S. kitaibelii, and S. pilosa; (A,E,I)—the general view; (B,F,J)—triangular edge; (C,D,G,H,K,L)—surfaces. BLC—bubble like cells; R—rib; AW—anticlinal walls.
Satureja coerulea
The nutlets of this species are dark brown, with lighter speckled areas and indistinct dark stripes that start from the top and reach toward the middle (Figure 10(1A–1D)). The nutlets of S. coerulea varied in size (from 113.32 μm to 182.38 μm) and form. The form varied from very small, round, bubble-like to elongated ovals (Figure 10(1A)). On the abaxial side of the nutlets, there is a ridge that starts from the middle of the surface. The tips of the nutlets are uneven and triangularly beveled on both sides (Figure 10(1C)). The exocarp surfaces have specific papilloma formations seen on the end of the nutlets (light microscope, Figure 10(1A–1C)). The SEM analysis showed that the surface of the exocarp is a reticulate convex type and comprises two types of cells. The first type consists of highly bulging, bubble-like cells and flat cells with numerous striations (Figure 11A–D). A thickened rim is observed at the base of the bulging cells. Comb-like connections are established between the two types of cells. The anticlinal walls are straight with slight striations, and the periclinal walls vary from smooth to wavy grooves. The waxes are observed all over the surface.
Satureja kitaibelii
The nutlets of S. kitaibelii are dark brown, flattened oval, with a winged triangular pointed tip, and length varied from 67.95 μm to 76.38 μm (Figure 10(2A–2D)).
Dark brown stripes are observed on the upper side from the tip, which starts from the top and reaches the middle of the nutlets (Figure 10(2A)). The nutlet of this Satureja species has a triangular edge in the middle from its lower part. The micromorphological SEM analyses showed that the epidermal cells of the exocarp are composed of two types. The first type of cell is with rhomboid forms, strongly convex, while the second type of cell is with polygonal shapes, tabular type (Figure 11E–H). The anticlinal walls are undulating with a rounded edge, while the periclinal walls are smooth with striations. According to the terminology and classification described by Barthlott and Ehler (1977), the surface of S. kitaibelii is convex type. The entire surface is covered with waxes.
Satureja pilosa
The seeds of S. pilosa are dark brown, from round to oval elongated forms, and varied in size from 129.22 μm to 140.07 μm. The tips of seeds are triangularly beveled, winged, and in lighter brown colors. On the adaxial surfaces of the seeds, there are three to four different lengths of longitudinal brown stripes that start from the tip, and they are located along the entire length of the seed (Figure 10(3A–3D)). The tip is arrow-shaped, beveled, and laterally, it is triangularly concave, with a lighter color (Figure 10(3A–3D)). Formations like warts (papillae) can be observed on the entire surface of the seeds (Figure 10(3A–3D)). The micromorphological SEM analyses showed that the epidermal cells of exocarp are of rectangular or polygonal shapes (Figure 11I–L). The anticlinal walls are undulating, with a pointed edge and indistinct striations, while periclinal walls are smooth to wavy convex. According to the terminology and classification described by Barthlott and Ehler (1977), the surface of S. pilosa is tabular to slightly convex type. The entire surface is covered with waxes.
3. Discussion
3.1. Embryological Analyses
This is the first study on the sporogenesis and gametogenesis of the three Balkan endemic Satureja species. The established features of the structures and development of gametophytes (M,F) are similar to those described in other species from Lamiaceae [20,21,22,23,34,40,41,42,43]. The main characteristics of gametophytes (M,F) of Satureja species are: four-sporangiate anthers with four-layered walls, tetrahedral microspore tetrads, anatropous tenuinucellate ovule, development of Polygonum-type female gametophyte and Onagrad-type embryo. The formation of placentoids, observed at the phase of one-cellular pollen that coincides with the beginning of degeneration of the tapetum, was described earlier in other Lamiaceae species such as Sideritis scardica Griseb [44] and Marrubium friwaldskyanum Boiss [45]. Furthermore, this structure was reported as a typical feature of the genera Lavandula, Salvia, and Stachys from Lamiaceae [41]. The observation of this structure in other genera such as Sideritis and Marrubium, as well as in this study, suggests that placentoids are a typical feature of the species belonging to the Lamiaceae family. Most of the reported data for other species from Lamiaceae showed that the chalazal megaspore of the tetrad was the functional megaspore, which becomes the embryo sac mother cell (MMC) [34,40,41,42,43]. In this study, it was established that in the Satureja species, the female gametophyte starts its development not only from the chalazal megaspore, but also from the micropylar one. The presence of more than one functional megaspore has been observed in other representatives of Lamiaceae. For example, in Lavandula spica L., Salvia officinalis L., Salvia sclarea L. and Nepeta hindostana (B. Heyne ex Roth) Haines, the development of megagametophytes starts with the four megaspores of the tetrad or only with the sub-micropylar one; or the micropylar and chalazal megaspores; or the sub-micropylar and chalazal megaspores; or the epi-chalazal and chalazal megaspores [41].
We observed that the synergids in the three studied Saturea species were hook-shaped, with well-shaped filiform apparatus. This characteristic was also identified in other Lamiaceae species [41] and can also be considered as a typical feature of all species from the Lamiaceae family.
The described characteristics of the reproductive sphere of the studied Satureja species define them as sexually reproducing species. This reproductive type ensures stability in the size of the populations of these species. The strict sexuality found in the three target species (no apomixis was observed) limits their adaptive abilities. Combined with the estimated low seed viability, this will affect the reproductive potential of their populations.
The high estimated percentage of fertile pollen in the three studied Satureja species is prerequisite to the successful pollination, fertilization and subsequent embryo (seed) formation. It is a result of the observed normal running of the processes of microsporogenesis and development of the male gametophyte. During the microsporogenesis ending predominantly with the formation of tetrahedral microspore tetrads, deviations were not observed. The development of male gametophytes in the pollen was successful, without significant degeneration. Despite this high percentage of viable mature pollen established in the three studied Satureja species and sufficient for normal fertilization and embryo (seed) formation, the estimated seed viability was low. A reduction in the percentage of viable seeds has also been found in other Lamiaceae species. For example, for Salvia nemorosa, Daskalova [22] estimated that a rather high percentage of seeds were empty and sterile (about 25%). According to the cited author, the large percentage of empty seeds obtained is due to degenerative processes occurring in the female generative sphere, affecting single macrospore tetrads and elements of the embryo sac (most often the egg cell and mature embryo sacs). Our results with the tested Satureja seeds showed that the percentage of empty seeds varied from 4.3 to 11.45% (Figure 5), while the percentage of unstained seeds varied between 64.52% and 82.35%. These results indicated that the high percentage of nonviable seeds in Satureja is probably due to the impact of environmental conditions rather than to deviations in megagametogenesis and the development of female gametophytes. The high percentage of mature uncolored seeds indicated that the processes leading to the formation of embryos proceeded normally, and the non-viability was a result of secondary disturbances in the quality of the obtained seeds under the influence of environmental factors. Between these factors, the fragmentation of the population of the studied species has a strong impact on their reproductive success. Habitat fragmentation was recognized as a major threat to plant–pollinator interactions [46,47,48,49]. Due to avoidance of small plant populations or isolated fragments by wide-ranging pollinators, reproduction may be reduced and eventually lead to extinction [48,49]. All of these factors affect the reproductive potential of the researched Satureja species. The results obtained for seed vitality corresponded with the results of the germination energy and germination tests (Table 1). Empty seeds and low vigor explain the low germination test results. Light, as an environmental factor, also affected seed germination. Members of the Satureja genus prefer sunny sites with high light intensity, and that is in accordance with the highest seed germination percentage of both species in variant 1 (natural daylight): 39% for S. pilosa and 40% for S. coerulea.
3.2. Scanning Electron Microscopy (SEM) Analysis
3.2.1. Leaves, Stem, and Calyx Surfaces
The micromorphology of leaf, calyx and stem surfaces of S. coerulea, S. kitaibelii and S. pilosa were analyzed by SEM. The microstructure of leaves, nutlets, calyx, and indumentum of Satureja species is an important systematic indicator for their taxonomy [50,51]. This research showed that the three Satureja species have many common and different micromorphological features. Overall, the indumenta of S. coerulea, S. kitaibelii and S. pilosa were presented by glandular and non-glandular trichomes, which is not unusual because, for Lamiaceae, this is specific [26,30]. Furthermore, morphology and distribution of trichomes can vary between Satureja species [26,30]. Peltate-type trichomes were the most frequently observed in the three species, but they were not located in all plant parts of the Satureja species. For example, on the stem of S. coerulea, only non-glandular trichomes were observed. This is not accidental because, as indicated in previous research on other Satureja species, the hairs show an organ preference [28,30]. The capitate-type trichomes were detected only on the surfaces of S. pilosa. The three types of non-glandular trichomes were present on S. pilosa, two types on S. kitaibelii, and one type on S. coerulea. Multicellular and unbranched trichomes were observed along the edge of leaves, while unicellular and uniseriate papillae trichomes were observed on the leaf surfaces (adaxial and abaxial) of S. kitaibelii (Figure 7G–L). The non-glandular trichomes on the surfaces of S. pilosa were several types: simple, short conical trichomes, unicellular, and multicellular (mostly four-cellular) conical trichomes (Figure 7N–R, Figure 8I–K and Figure 9G–I). All described non-glandular trichomes of S. pilosa had specific papillose surfaces that were reported for the first time.
The varying shapes, types and specific characteristics of epidermal cells and stomatal complexes of plants are important taxonomic values, and they are successfully used in taxonomy [52]. In this regard, the three studied Satureja species showed different forms of main epidermal cells and stomatal complexes that are related with their taxonomic position. The main epidermal cells of S. coerulea are irregular to rectangular in shape, with slightly raised or smooth periclinal walls, while in S. kitaibelii and S. pilosa, the forms of main cells are isodiametric, and the periclinal walls are striated. The cuticle striation is an important diagnostic indication for the ecological adaptation of species and their taxonomy [53]. Previous investigations of foliar epidermal surfaces of species from Lamiaceae showed that epidermal cells varied from irregular, isodiametric, and rectangular forms, and based on these characters, the authors developed a taxonomic key [50]. The stomatal apparatus is another important characteristic of a species [52]. In the present study’s SEM observations of the three Satureja species, amphistomatic leaves were detected, which is in agreement with reports for other Satureja species [54]. Previous research on leaves of S. pilosa reported hypostomatic leaves [26], which was not confirmed in this study. The stomatal complexes of the studied Satureja taxa showed distinctive patterns. The stomata of S. coerulea are elliptical and submerged in the epidermis, with distinct cuticles, while the stomata of S. kitaibelii are round, small, and diacytic. Furthermore, the cells of the stomatal complex of S. kitaibelii have specific striations, and the stomata of S. pilosa are oval shapes with a double rim of waxes at the level of the epidermis. Micromorphological analyses of epidermis and stomata of other Satureja species have been performed previously [26,28,30,55,56]. The conclusion of all cited authors was that the main epidermal cell and stomatal complex show variation, but diacytic-type stomata are a common feature of most species in the Lamiaceae family. Generally, the epidermis, calyx and stem surfaces of Bulgarian samples of S. pilosa, S. kitaibelii, and S. coerulea were analyzed for the first time. The micromorphological analysis revealed that the surfaces of S. coerulea are clearly distinct from those of the other two Satureja species.
3.2.2. Nutlet (Seed) Surfaces
The taxonomic importance of the micromorphology of nutlet surfaces of Satureja species is well-documented [1]. As known in Lamiaceae, the fruits are schizocarpic. After reaching maturity, the fruit is separated into four nutlets (cocci), and the fruit coat and seed fuse together [57]; the fruit is identified as the seed in Lamiaceae. Scanning electron microscopy (SEM) has been widely used to study the surface features of nutlets in Satureja species because the micromorphology of the fruits and ornamentation are much more important in taxonomic studies [58]. SEM analysis and stereomicroscope observation in this study on nutlets of S. coerulea, S. kitaibelii, and S. pilosa showed common but distinguishing features. The common features of the nutlets were their colors, forms, and variations in size. In addition, brown longitudinal lines were observed on the adaxial surfaces of nutlets for the three studied species. These marks were here described for the first time for S. coerulea, S. kitaibelii, and S. pilosa. As was described in the Results section, the tips of the nutlets are triangular and beveled on both sides unevenly. For this reason, determining the shapes of the nutlets depends on how they are arranged in space, so they appear differently (Figure 10 and Figure 11). Most of the previous research on Satureja micromorphology grouped the nutlets into two main types and several subgroups, but there is no uniform classification [26,59]. For example, according to Husain et al. [26], in the first group were the nutlets with protuberances and without trichomes and oil glands (S. montana group) [26]. In the second group, the authors included nutlets with surfaces that were without protuberances, with papillae, trichomes and sessile oil glands, where they placed S. pilosa [26]. In another study, Kaya et al. [59] divided Satureja nutlets into two main types: those with more or less smooth surfaces and those with sculptured surfaces, respectively. Furthermore, they subdivided the first type into four subtypes: undulate-reticulate, reticulate, reticulate-protuberculate and papillate-tuberculate, where they put S. coerulea [59]. The cited authors [59] found tiny, stalked glandular and eglandular hairs at the nutlets’ apex and on the median edge in S. cilicica, S. coerulea, S. icarica, S. parnassiaca subsp. sipylea, S. pilosa, S. spinosa, S. thymbra, and S. wiedemanniana. In this study, glandular trichomes were not found on nutlets of any of the three studied Satureja species. The exocarp surfaces of S. coerulea and S. pilosa have specific papilloma formations, such as the non-glandular trichomes seen on the end of the nutlets of S. coerulea, while they are on all surfaces of S. pilosa.
Generally, our study showed different characteristics for the surfaces of the nutlets from S. coerulea, S. kitaibelii, and S. pilosa. Therefore, it was difficult to determine into which group the nutlets we studied fell.
4. Materials and Methods
4.1. Collection of the Plant Materials
Plant materials of the three Satureja species were collected from natural populations as follows: Satureja pilosa—locality Selce, Stara planina (42°37′04.3″ N 25°33′02.2″ E; 749 masl); Satureja coerulea—locality Markovo, Rhodope mountain (42°02′30.7″ N 24°42′54.1″ E; 638 masl); Satureja kitaibelii—locality Kostenkovci, Stara planina (42°57′35.7″ N 25°25′00.1″ E; 604 masl).
4.2. Embryological Analyses
The reproductive potential of the three studied Satureja species was investigated. The main parameters were estimated as follows: (1) structures and processes in the male and female generative sphere; (2) pollen and nutlet (seed) viability. The flower buds, flowers at different development stages, and nutlets were collected from natural populations of the species. For revealing the peculiarities of structures and development of male and female gametophytes, previously described procedures were followed [44]. According to the methods described by Sundara [60], the fixed material was dehydrated by passing it through increasing ethanol solutions and embedding in paraffin wax. The paraffin-embedded material was cut using a rotary microtome Leica RM2125RT into thicknesses of 8 to 12 μm. Permanent slides were prepared. The description of characteristics of structures in the generative sphere of the studied species was made based on observations using an Olympus Light CX21 microscope (Olympus Corporation, Shinjuku, Tokyo, Japan). The microphotographs were taken with an “Infinity lite” digital camera 1.4 Mpx (Lumenera Corporation, Ottawa, ON, Canada).
4.2.1. Pollen Viability
The quality of the produced mature pollen grains was estimated through pollen viability according to an acetocarmine test [61]. The mature pollen grains in 30 anthers from different individual plants per species were treated with a solution of 1% acetocarmine. On the basis of the intensity of staining from the acetocarmine solution, the pollen grains were classified as follows: (1) viable pollen grains (pollen grains stained in red, with clear reticulated sculpture and strongly distinguished vegetative and generative cells or sperm); (2) nonviable pollen grains (colorless or transparent pollen grains). The fertile and infertile pollen grains were counted on a visible field using a light microscope (described above) at magnification 100× or 400×. The pollen viability was calculated and presented in percentages.
4.2.2. Nutlet (Seed) Viability Testing
Nutlet (embryo) viability was assessed using a tetrazolium test [38]. This topographical tetrazolium method differentiates live from dead seeds based on the activity of the respiration enzymes in seeds. When nutlets are in contact with a solution of 2,3,5-triphenyl tetrazolium chloride, the activity of dehydrogenase enzymes increases. The hydrogen ions reduce the colorless tetrazolium solution into a compound named formazan. As known, formazan stains the cells (respiring) in red if they are alive, and the dead cells remain colorless. Thus, the staining pattern after application of the tetrazolium test reveals the live and dead areas of the embryo and enables one to determine if seeds have the capacity to produce normal seedlings [37]. Approximately 100 mature seeds per species were used, and the procedure for preparation of nutlets was as previously described [45]. According to criteria described by Moore [39], the viable embryos display entire embryo staining (in red or in pink) or staining of their basal part—the root; the nonviable embryos display abnormal or no staining.
4.2.3. Testing of Nutlet (Seed) Germination under Different Light Regimes
The test for germination with different wavelengths (colors) of light was conducted under light intensity of 250 micromoles. A photoperiod of 16 h light and 8 h dark was used. The tested nutlets were arranged in Petri dishes, and they were positioned at a distance of 60 cm from the light sources. The temperature was set at 23 °C during the day and 20 °C at night. Fifty seeds per species were used in each Petri dish in three replications for each variant. Every day, distilled water was added to ensure the seeds remained moist. The experimental design was as previously reported [45]. (1) The first variant (V1) was natural daylight; (2) the second variant (V2) was fluorescent white light with the addition of LED red and blue in a ratio of 7:1; (3) the third variant (V3) was fluorescent white light with the addition of LED red and blue in a ratio of 4:1; and (4) the fourth variant (V4) was fluorescent white light. According to the procedure described in BDS, GE was measured on the fifth day, and total G was measured on the tenth day. All data were analyzed with descriptive statistics, performed using the averages and standard deviation (SD) of the germination energy (%) and total germination (%) obtained from the samples in triplicate. The wavelengths of the light color specifications were reported previously [45].
4.3. Scanning Electron Microscopy (SEM) Analysis
For this investigation, an FEI Quanta 600 scanning electron microscope (SEM) at the Microscopy Facility of Oregon State University in the United States was utilized. Sample preparation involved placing small samples into a fixative consisting of 1% paraformaldehyde and 2.5% glutaraldehyde in 0.1 M sodium cacodylate buffer of pH 7.4. The samples were soaked in fixative for 2 h, followed by two rinses in 0.1 M Cacodylate buffer for 15 min each, and then underwent a dehydration series in acetone (10%, 30%, 50%, 70%, 90%, 95%, and 100%), with each step lasting 10–15 min. The samples were then subjected to critical point drying (two "bomb flushes" at chamber pressure to 5 °C, followed by filling the chamber with CO2). After venting for 5 min, the procedure was repeated. The dry samples were mounted onto an aluminum SEM stub using double-stick carbon tape and sputter-coated with a Cressington 108A sputter coater from Ted Pella with Au/Pd, 60/40 mix. The shape and surface structure of the nutlets (seeds) of the species were described morphologically. In this study, the terminology and classification described by Barthlott and Ehler [62] were adopted.
5. Conclusions
This was the first study of the reproductive capacity and micromorphological (SEM) characteristics of leaf, calyx, stem and nutlet surfaces of three Balkan-endemic Satureja species (S. coerulea, S. kitaibelii, and S. pilosa). This study revealed that all three species reproduce sexually. The established characteristics of the processes in the reproductive sphere suggest low plasticity of the species and is probably the reason for their endemism. Furthermore, the results showed that the reproductive potential of the populations of the three Balkan-endemic Satureja species depended on the environmental conditions and the established fragmentation of their habitats. Indeed, the balanced processes and stable structures in the male and female generative spheres, combined with high pollen viability, as observed in this study, provide high reproductive potential, but the produced nutlets had low viability and germination. Furthermore, high numbers of nutlets were empty with undeveloped embryos, which is a major problem for species distribution. The habitats of these endemic species are fragmented, found on dry grasslands, rocky slopes, and stony places where environmental conditions are rather extreme. Therefore, the unfavorable conditions affect pollination and seed production of S. coerulea, S. kitaibelii and S. pilosa. Consequently, the most efficient approach for preserving these species would comprise measures aimed at conserving their habitats and gene pool, such as (1) monitoring the state of the populations; (2) limiting anthropogenic pressure on the populations; (3) establishing in situ and ex situ collections; and (4) developing the three species into field crops for EO production.
Overall, the SEM analyses of S. coerulea, S. kitaibelii, and S. pilosa showed variability between the three species. Satureja coerulea microstructural surfaces (leaves, calyx, stem) were clearly dissimilar to those of the two other Satureja species. Regarding the nutlet surfaces, this study established that the exocarp surfaces of S. coerulea and S. kitaibelii had a reticulate convex-type surface, while S. pilosa exocarp was a tabular to slightly convex type, without bubble-like cells. The seeds of both S. coerulea and S. pilosa had specific papilla formations that were not found in S. kitaibelii. Furthermore, the non-glandular papillae in S. coerulea were observed only on the back side of its nutlets, while those in S. pilosa were observed over the entire surface of the nutlets.
Author Contributions
Conceptualization, I.S., V.D.Z. and E.Y.-T.; methodology, E.Y.-T., L.H.K.-V., R.N. and I.S.; formal analysis, L.H.K.-V.; investigation, I.S., V.D.Z. and E.Y.-T.; resources, I.S., V.D.Z., E.Y.-T., L.H.K.-V. and R.N.; data curation, I.S. and E.Y.-T.; writing—original draft preparation, I.S. and E.Y.-T.; writing—review and editing, I.S., V.D.Z., E.Y.-T., L.H.K.-V. and R.N.; supervision, V.D.Z.; project administration, I.S. All authors have read and agreed to the published version of the manuscript.
Funding
The project was funded by the Agricultural University in Plovdiv, Bulgaria (project 11–18 of the AU Research Center), and by Oregon State University, USA.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data is contained within the article.
Acknowledgments
The authors are grateful for the financial support provided by the Agricultural University in Plovdiv, Bulgaria (project 11–18 of the AU Research Center). We thank Teresa Sawyer of Oregon State University for the help with the scanning electron microscopy sample preparation and analyses.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Doroszenko, A. Taxonomic Studies of Satureja Complex (Labiate). Ph.D. Thesis, Edinburgh University and Royal Botanic Garden, Edinburgh, UK, 1986. Available online: https://www.researchgate.net/publication/319540144_Taxonomic_studies_on_the_satureja_complex_Labiatae (accessed on 14 April 2023).
- Başer, K.H.C.; Özek, T.; Kirimer, N.; Tümen, G. A Comparative study of the essential oils of wild and cultivated Satureja hortensis L. J. Essent. Oil Res. 2004, 16, 422–424. [Google Scholar] [CrossRef]
- Nooshkam, H.M.; Hadian, J.; Alemardan, A.; Mohammad, R.M. Drug yield and essential oil and carvacrol contents of two species of Satureja (S. khuzistanica Jamzad and S. rechingeri Jamzad) cultivated in two different locations. J. Appl. Res. Med. Arom. Plants 2017, 6, 126–130. [Google Scholar] [CrossRef]
- Ćavar, S.; Maksimović, M.; Šolić, M.E.; Jerković-Mujkić, A.; Bešta, R. Chemical composition and antioxidant and antimicrobial activity of two Satureja essential oils. Food Chem. 2008, 111, 648–653. [Google Scholar] [CrossRef]
- Skočibušić, M.; Bezić, N.; Dunkić, V. Phytochemical composition and antimicrobial activities of the essential oils from Satureja subspicata Vis. growing in Croatia. Food Chem. 2006, 96, 20–28. [Google Scholar] [CrossRef]
- Semerdjieva, I.B.; Zheljazkov, V.; Cantrell, C.L.; Astatkie, T.; Ali, A. Essential oil yield and composition of the Balkan endemic Satureja pilosa Velen. (Lamiaceae). Molecules 2020, 25, 827. [Google Scholar] [CrossRef]
- Zheljazkov, V.D.; Semerdjieva, I.B.; Cantrell, C.L.; Astatkie, T.; Aćimović, M. Phytochemical variability of essential oils of two Balkan endemic species: Satureja pilosa Velen. and S. kitaibelii Wierzb. ex Heuff. (Lamiaceae). Molecules 2022, 27, 3153. [Google Scholar] [CrossRef]
- Tutin, T.G.; Heywood, V.H.; Burges, N.A.; Mooze, D.M.; Valeutine, D.H.; Walters, S.M.; Webb, D.A. Genus Satureja. In Flora Europaea; Tutin, T.G., Ed.; Cambridge University Press: Cambridge, MA, USA, 1972; Volume 3, pp. 108–124. [Google Scholar]
- Anchev, M. Genus Satureja. In Flora of Bulgaria; Yordanov, D., Ed.; BAS: Sofia, Bulgaria, 1989; Volume 9, pp. 335–342. (In Bulgarian) [Google Scholar]
- Šućur, J.; Popović, A.; Petrović, M.; Anačkov, G.T.; Bursić, V.; Kiprovski, B.; Prvulović, D. Allelopathic effects and insecticidal activity of aqueous extracts of Satureja montana L. J. Serbian Chem. Soc. 2015, 80, 475–484. [Google Scholar] [CrossRef]
- Coelho, N.; Gonçalves, S.; Romano, A. Endemic plant species conservation: Biotechnological approaches. Plants 2020, 9, 345. [Google Scholar] [CrossRef]
- Yıldız, F.; Aslay, M.; Kandemir, A.; Kaya, O. Reproductive biology of Fritillaria aurea Schott (Liliaceae), a rare species endemic to Turkey. Diversity 2022, 14, 1052. [Google Scholar] [CrossRef]
- Gopalakrishnan, K.K.; Thomas, T.D. Reproductive biology of Pittosporum dasycaulon Miq., (Family Pittosporaceae) a rare medicinal tree endemic to Western Ghats. Bot. Stud. 2014, 55, 15. [Google Scholar] [CrossRef]
- Tilman, D.; Lehman, C. Human-caused environmental change: Impacts on plant diversity and evolution. Proc. Natl. Acad. Sci. USA. 2001, 98, 5433–5440. [Google Scholar] [CrossRef]
- Murray, B.R.; Thrall, P.H.; Gill, A.M.; Nicotra, A.B. How plant life-history and ecological traits relate to species rarity and commonness at varying spatial scales. Austral Ecol. 2002, 27, 291–310. [Google Scholar] [CrossRef]
- Castro, S.; Silveira, P.; Navarro, L. How flower biology and breeding system affect the reproductive success of the narrow endemic Polygala vayredae Costa (Polygalaceae). Bot. J. Linnean Soc. 2008, 157, 67–81. [Google Scholar] [CrossRef]
- Anderson, G.J. Systematics and reproductive biology. In Experimental and Molecular Approaches to Plant Systematics; Hoch, P.C., Stephenson, A.G., Eds.; Monogr Syst. Bot 53; Missouri Botanical Garden: St. Louis, MI, USA, 1995; pp. 263–272. [Google Scholar]
- Bernardello, G.; Anderson, G.J.; Lopez, S.P.; Cleland, M.A.; Stuessy, T.F.; Crawford, D.J. Reproductive biology of Lactoris fernandeziana (Lactoridaceae). Am. J. Bot. 1999, 86, 829–840. [Google Scholar] [CrossRef]
- Sharia, A.; Sefidkon, F. Investigation of pollination system and self-incompatibility intwo species of Satureja sahendica and S. bachtiarica. Iran. J. Rangelands Forest Plants 2020, 27, 204–215. [Google Scholar] [CrossRef]
- Daskalova, T.; Genova, E. Histological structure of the anthers and microsporogenesis in Hyssopus officinalis L. ssp. aristatus (Godr.) Briq. (Lamiaceae). Bot. Jahrb. Syst. 1996, 118, 297–302. [Google Scholar]
- Daskalova, T. Histological structure of the microsporangia, microsporogenesis and development of the male gametophyte in Nepeta cataria (Lamiaceae). Phytol. Balcan. 2004, 10, 241–246. Available online: http://www.bio.bas.bg/~phytolbalcan/PDF/10_2-3/10_2-3_18_Daskalova.pdf (accessed on 17 January 2023).
- Daskalova, T. On some specificities of seed formation in Salvia nemorosa (Lamiaceae). Phytol. Balcan 2004, 10, 7984. Available online: https://www.researchgate.net/publication/237790347 (accessed on 17 January 2023).
- Daskalova, T. Polygamy in some species of Nepeta (Lamiaceae). Phytol. Balcan. 2005, 11, 79–98. Available online: http://www.bio.bas.bg/~phytolbalcan/PDF/11_1/11_1_10_Daskalova.pdf (accessed on 17 January 2023).
- Daskalova, T. Polygamy in Lamium maculatum L. C. R. Acad. Bulg. Sci. 2008, 61, 327–334. Available online: https://www.researchgate.net/scientific-contributions/Tzvetana-Daskalova-2015103073 (accessed on 17 January 2023).
- Wightman, R. An overview of cryo-scanning electron microscopy techniques for plant imaging. Plants 2022, 11, 1113. [Google Scholar] [CrossRef] [PubMed]
- Husain, S.; Marin, P.D.; Šilić, Č.; Qaiser, M.; Petrović, B. A micromorphological study of some representative genera in the tribe Saturejeae (Lamiaceae). Bot. J. Lin. Soc. 1990, 103, 59–80. [Google Scholar] [CrossRef]
- Satil, F.; Tümen, G.; Akçelik, A.; Baser, K.H.C. Comparative morphological, anatomical, ecological and chemical studies on endemic Satureja parnassica subsp. sipylea from Turkey. Acta Bot. Croatica 2002, 61, 207–220. Available online: https://hrcak.srce.hr/3459 (accessed on 18 January 2023).
- Marin, M.; Ascensao, L.; Lakušić, B. Trichomes of Satureja horvatii Šilić (Lamiaceae): Micromorphology and histochemistry. Arch. Biol. Sci. 2012, 64, 995–1000. [Google Scholar] [CrossRef]
- Malmir, M.; Serrano, R.; Reza Gohari, A.; Silva, O. Characterization of Satureja khuzestanica Leaf as a herbal medicine. Microsc. Microanal. 2014, 20, 1425–1435. [Google Scholar] [CrossRef]
- Dodoš, T.; Janković, S.; Marin, P.D.; Rajčević, N. Essential oil composition and micromorphological traits of Satureja montana L., S. subspicata Bartel Ex Vis., and S. kitaibelii Wierzb. Ex Heuff. Plant Organs. Plants 2021, 10, 511. [Google Scholar] [CrossRef]
- Bräuchler, C.; Meimberg, H.; Heubl, G. Molecular phylogeny of Menthinae (Lamiaceae, Nepetoideae, Mentheae)—Taxonomy, biogeography and conflicts. Mol. Phylogen. Evol. 2010, 55, 501–523. [Google Scholar] [CrossRef]
- Khadivi-Khub, A.; Salehi-Arjmand, H.; Movahedi, K.; Hadian, J. Molecular and morphological variability of Satureja bachtiarica in Iran. Plant Syst. Evol. 2015, 301, 77–93. [Google Scholar] [CrossRef]
- Hedge, I.C.; Feinbrun, N. Satureia thymbrifolia Hedge & Feinbrun sp. nov. Israel. J. Bot. 1969, 17, 213. Available online: http://www.worldfloraonline.org/taxon/wfo-0000304558 (accessed on 14 May 2023).
- Davis, G.L. Systematic Embryology of the Angiosperms; John and Wiley and Sons: Hoboken, NJ, USA, 1966. [Google Scholar]
- Johansen, D.A. A critical survey of the present status of plant embryology. Bot. Rev. 1945, 11, 87–107. [Google Scholar] [CrossRef]
- Johansen, D.A. Plant Embryology; Chronica Botanica: Waltham, MA, USA, 1950. [Google Scholar]
- Copeland, L.O.; Mc Donald, M.B. Principles of Seed Science and Thechnology, 4th ed.; Kluwer Academic Publishers: Hingham, MA, USA, 2001. [Google Scholar] [CrossRef]
- Peters, J. Tetrazolium testing handbook. In Contribution No 29 to the Handbook on Seed Testing Revised; The Association of Official Seed Analysts (AOSA); PMB: Las Cruces, NM, USA, 2000. [Google Scholar]
- Moore, R.P. Handbook on Tetrazolium Testing; International Seed Testing Association: Zürich, Switzerland, 1985; p. 99. [Google Scholar]
- Poddubnaya-Arnoldi, V.A. Characteristics of Flowering Plants Families According to the Cytoembryological Features; Nauka: Moscow, Russia, 1982. (In Russian) [Google Scholar]
- Kamellina, O.; Dzevaltovsky, A. Lamiaceae. In Comparative Embryology of Flowering Plants; Batygina, M., Yakovlev, T.B., Eds.; Nauka: Leningrad, Russia, 1987; pp. 225–236. (In Russian) [Google Scholar]
- Kai-yu, P.; Wen Jie, W.; Shi-liang, Z. Embryological study on Mosla chinensis (Lamiaceae). Acta Bot. Sinica 1997, 39, 111–116. Available online: https://www.jipb.net/EN/abstract/abstract26600.shtml (accessed on 18 January 2023).
- Jafari Marandi, S.; Arbabian, S.; Musavi, M. Structure and development of male and female gametophyte in Ocimum basilicum L. Caryologia 2017, 70, 102–111. [Google Scholar] [CrossRef]
- Yankova-Tsvetkova, E.; Yurukova-Grancharova, P.; Vitkova, A. Reproductive biology of the Balkan endemic Sideritis scardica (Lamiaceae). Bot. Serbica 2013, 37, 83–87. Available online: https://botanicaserbica.bio.bg.ac.rs/arhiva/pdf/2013_37_1_580_full.pdf (accessed on 17 January 2023).
- Zheljazkov, V.D.; Semerdjieva, I.B.; Stevens, J.F.; Wu, W.; Cantrell, C.L.; Yankova-Tsvetkova, E.; Koleva-Valkova, L.H.; Stoyanova, A.; Astatkie, T. Phytochemical investigation and reproductive capacity of the Bulgarian endemic plant species. Marrubium friwaldskyanum Boiss. (Lamiaceae). Plants 2022, 11, 114. [Google Scholar] [CrossRef]
- Aizen, M.A.; Ashworth, L.; Galetto, L. Reproductive success in fragmented habitats: Do compatibility systems and pollination specialization matter? J. Veg. Sci. 2002, 13, 885–892. Available online: https://www.jstor.org/stable/3236934 (accessed on 18 January 2023). [CrossRef]
- Aizen, M.A.; Feinsinger, P. Habitat fragmentation, pollination, and plant reproduction in a Chaco dry forest. Ecology 1994, 75, 330–351. Available online: https://www.jstor.org/stable/pdf/1939538.pdf (accessed on 25 May 2023). [CrossRef]
- Buchmann, S.L.; Nabhan, G.P. The Forgotten Pollinators; Island Press: Washington, DC, USA, 1996. [Google Scholar]
- Kearns, C.A.; Inouye, D.W.; Waser, N.M. Endangered mutualisms: The conservation of plant-pollinator interactions. Ann. Rev. Ecol. Syst. 1998, 29, 83. [Google Scholar] [CrossRef]
- Gul, S.; Ahmad, M.; Zafar, M.; Bahadur, S.; Celep, F.; Sultana, S.; Begum, N.; Hanif, U.; Zaman, W.; Shuaib, M.; et al. Taxonomic significance of foliar epidermal morphology in Lamiaceae from Pakistan. Mycrosc. Res. Techniq. 2019, 82, 1507–1528. [Google Scholar] [CrossRef]
- Zaman, W.; Fazal Ullah, F.; Parmar, G.; Saqib, S.; Ayaz, A.; Park, S.J. Foliar micromorphology of selected medicinal Lamiaceae taxa and their taxonomic implication using scanning electron microscopy. Mycrosc. Res. Techniq. 2022, 85, 3217–3236. [Google Scholar] [CrossRef]
- Metcalfe, C.R.; Chalk, L. Anatomy of the Dicotyledons, Systematic Anatomy of the Leaf and Stem, 2nd ed.; Claredon Press: Oxford, UK, 1979; Volume 1. [Google Scholar]
- Barthlott, W. Epidermal and seed surface characters of plants: Systematic applicability and some evolutionary aspects. Nord. J. Bot. 1981, 1, 345–355. [Google Scholar] [CrossRef]
- Satil, F.; Kaya, A. Leaf anatomy and hairs of Turkish Satureja L. (Lamiaceae). Acta Biol. Cracov. Ser. Bot. 2007, 49, 67–76. Available online: https://abcbot.pl/pdf/49_1/09satil.pdf (accessed on 20 April 2023).
- Redžić, S.; Tuka, M.; Pajević, A. Research into microscopic structure and essential oils of endemic medicinal plant species Satureja subspicata Bartl. Ex Vis. (Lamiaceae). Bosn. J. Basic Med. Sci. 2006, 6, 25–31. [Google Scholar] [CrossRef] [PubMed]
- Dunkić, V.; Bezić, N.; Ljubešić, N.; Bočina, I. Glandular hair ultrastructure and essential oils in Satureja subspicata Vis. ssp. subspicata and ssp. liburnica Šilić. Acta Biol. Cracov. 2007, 49, 45–51. Available online: https://abcbot.pl/pdf/49_2/06_dunkic.pdf (accessed on 20 April 2023).
- McDonald, I.; Kwong, M.B.I.; Francis, Y. Flowers-Seeds. In Flower Seeds: Biology and Technology; McDonald, I., Kwong, F., Eds.; CAB International: Wallingford, UK, 2005; p. 372. Available online: https://www.scribd.com/document/239950465/Flower-Seeds-Biology-and-Technology (accessed on 18 January 2023).
- Duletiæ-Lauševiæ, S.; Marin, P.D. Pericarp structure and myxocarpy in selected genera of Nepetoideae (Lamiaceae). Nord. J. Bot. 1999, 19, 435–446. [Google Scholar] [CrossRef]
- Kaya, A.; Satil, F.; Gogel, F. Nutlet surface micromorphology of Turkish Satureja (Lamiaceae). Biologia 2009, 64, 902–907. [Google Scholar] [CrossRef]
- Sundara, R.S. Practical Manual of Plant Anatomy and Embryology; Anmol Publications Pvt Ltd.: New Delhi, India, 2000; Available online: https://www.bagchee.com/books/BB28913/practical-manual-of-plant-anatomy-and-embryology (accessed on 18 January 2023).
- Heslop-Harrison, J.S. Pollen Capture Adhesion and Hydration. In Sexual Plant Reproduction; Cresti, M., Tizzi, A., Eds.; Springer: Berlin/Heidelberg, Germany, 1992; pp. 81–88. Available online: https://link.springer.com/chapter/10.1007/978-3-642-77677-9_8 (accessed on 18 January 2023).
- Barthlott, W.; Ehler, N. Raster-Elektronenmikroskopie der Epidermis-Oberfl€achen von Spermatophyten. Trop. Subtrop. Pflanzenwelt 19; Akad. Wiss. Lit. Mainz; Franz Steiner Verlag: Stuttgart, Germany, 1977; p. 105. [Google Scholar]












Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).