
Effect of Drought Stress on Capsaicin and Antioxidant Contents in Pepper Genotypes at Reproductive Stage


 ,
, 

 ,
,  ,
,  , and
, and 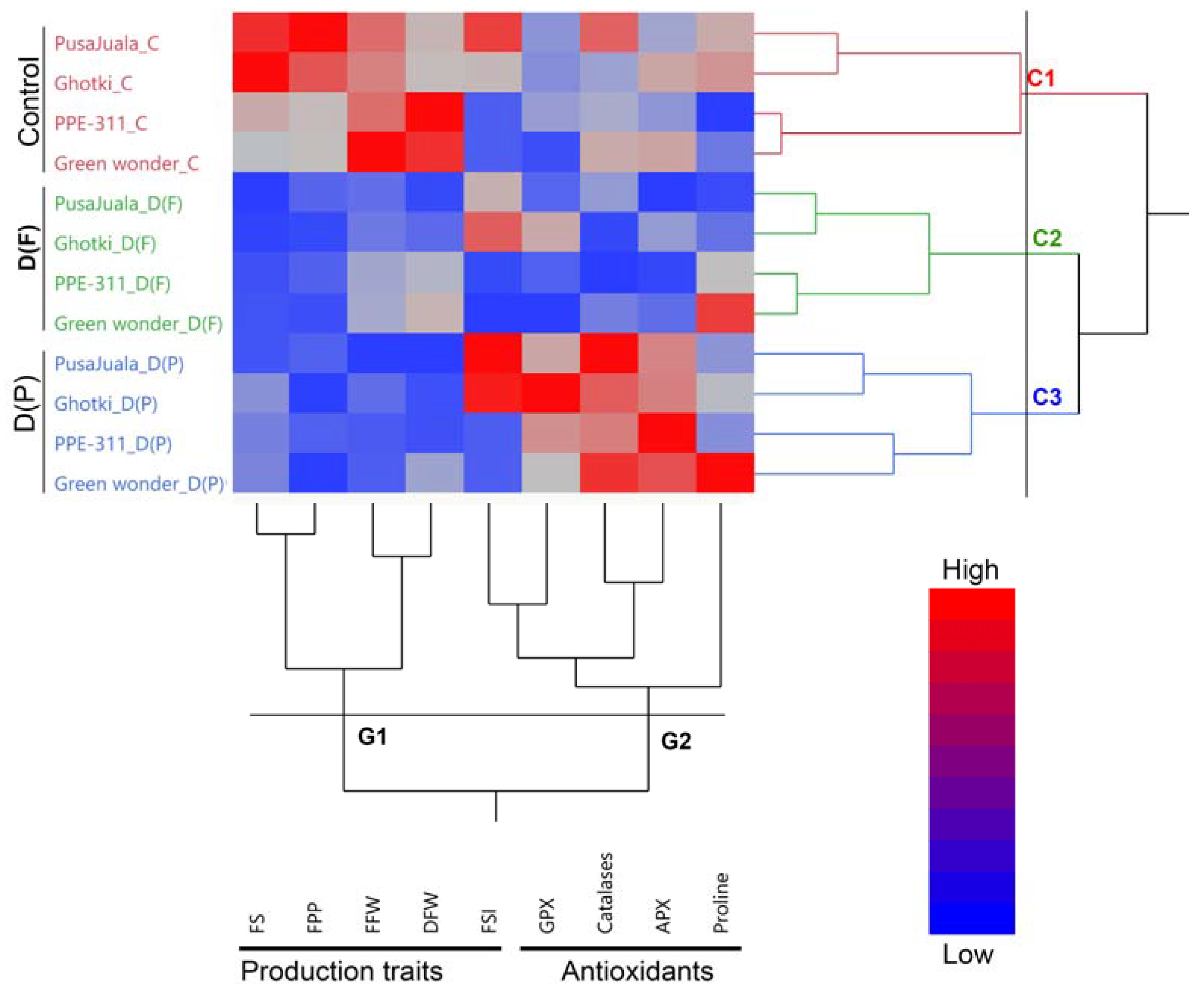{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Flower and Fruit Data
2.3. Detection of Capsaicin Contents by Scoville Scale
2.4. Proline Determination
2.5. Antioxidant Enzyme Assay
2.6. Statistical Analysis
3. Results
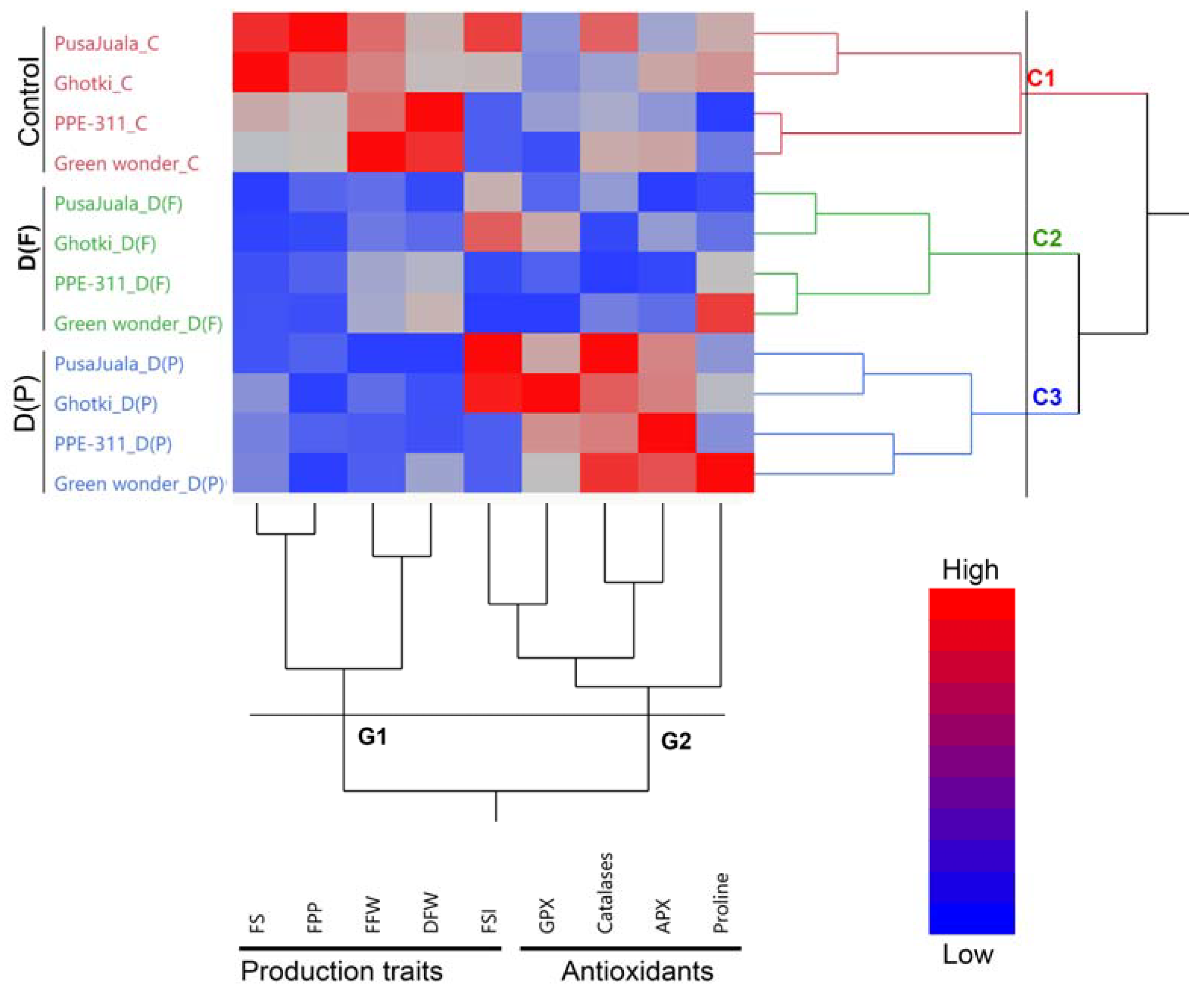
3.1. Clustering and Comparative Analysis of Pepper Genotypes and Traits
3.2. Effect of Drought Stress on Antioxidants and Production Traits
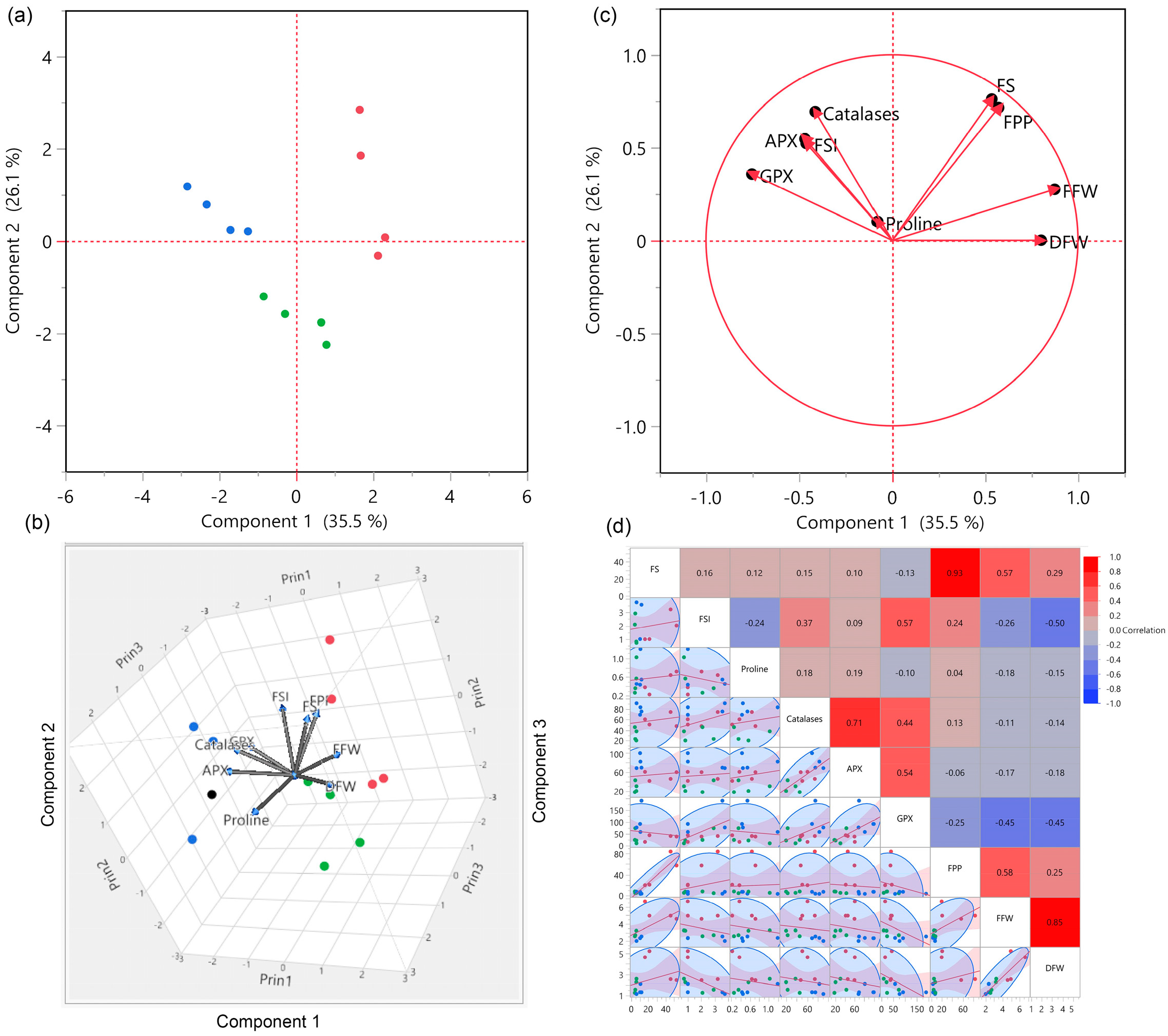
3.3. Association of Production Traits with Antioxidants
3.4. Principal Component Analysis
3.5. Mean Performance and Graphical Presentation of Traits
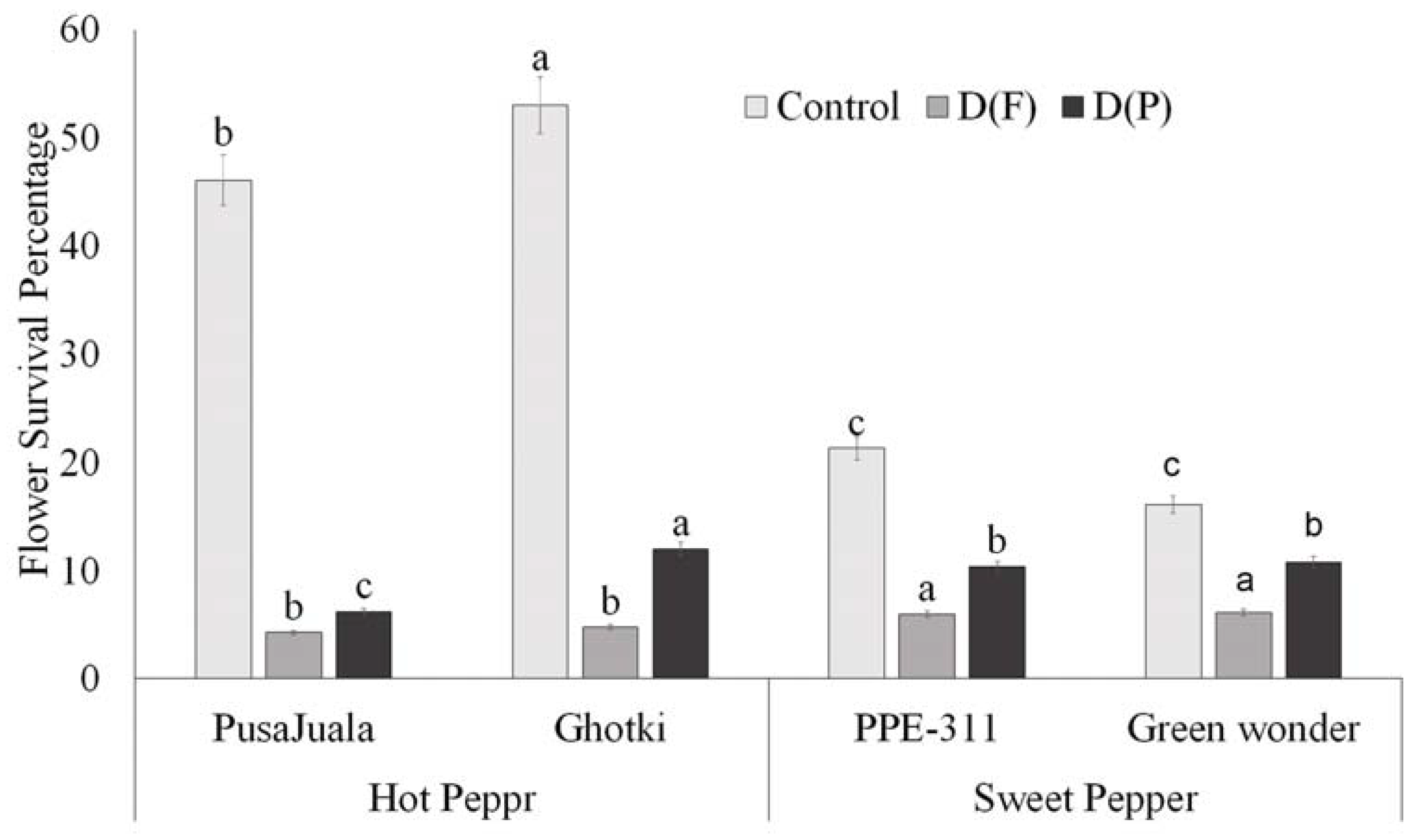
3.5.1. Flower Survival Percentage
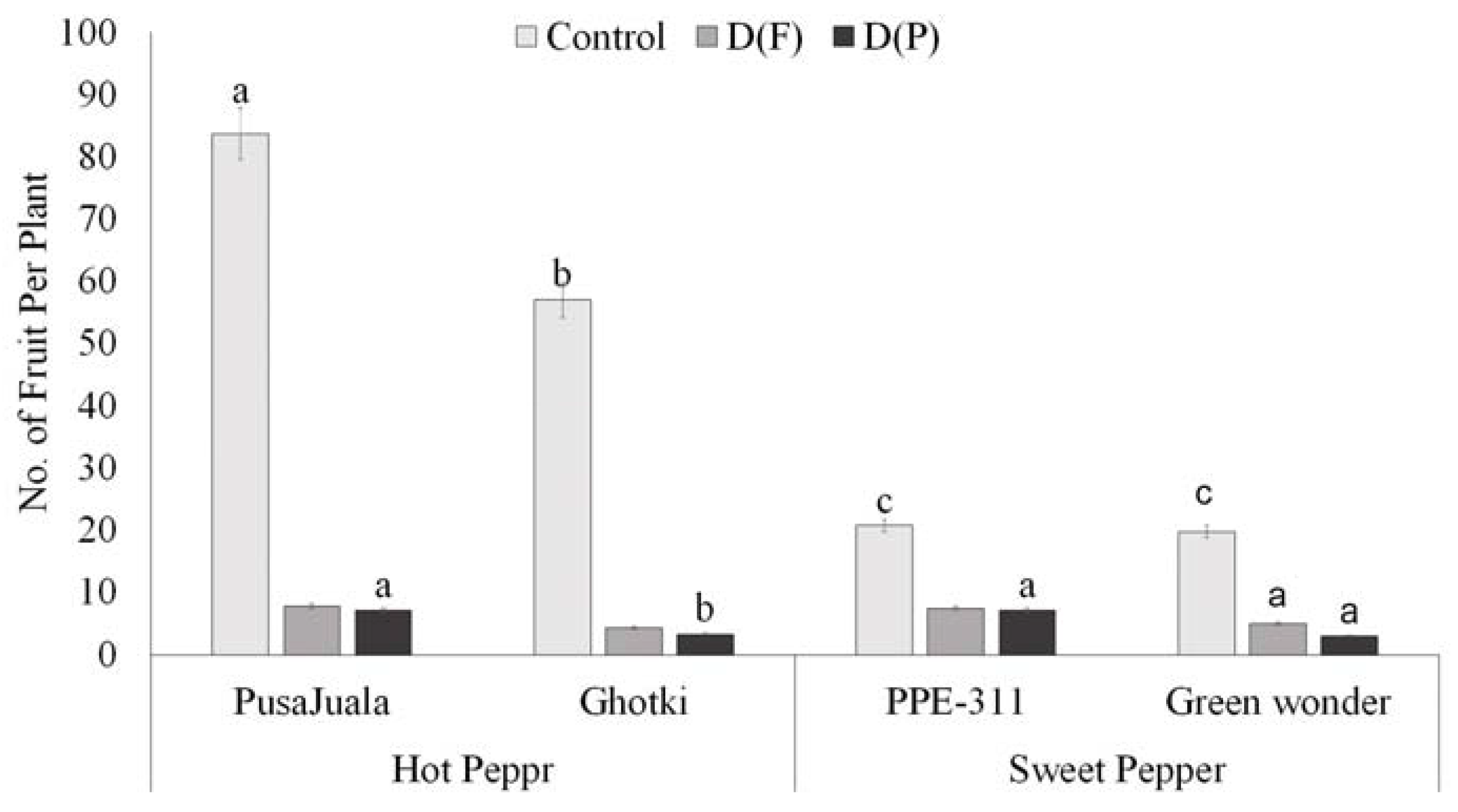
3.5.2. Number of the Fruits Per Plant
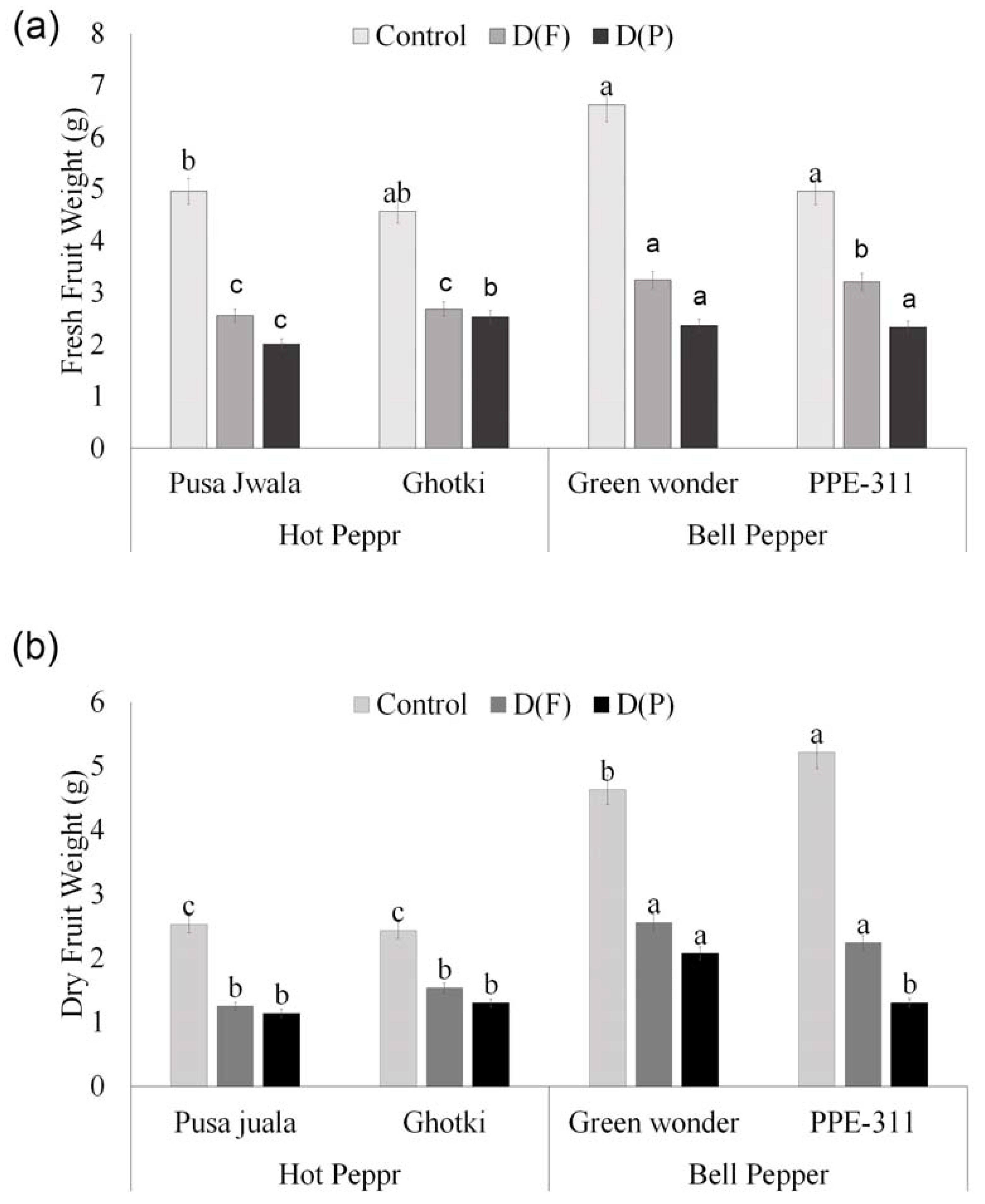
3.5.3. Fruit Weight
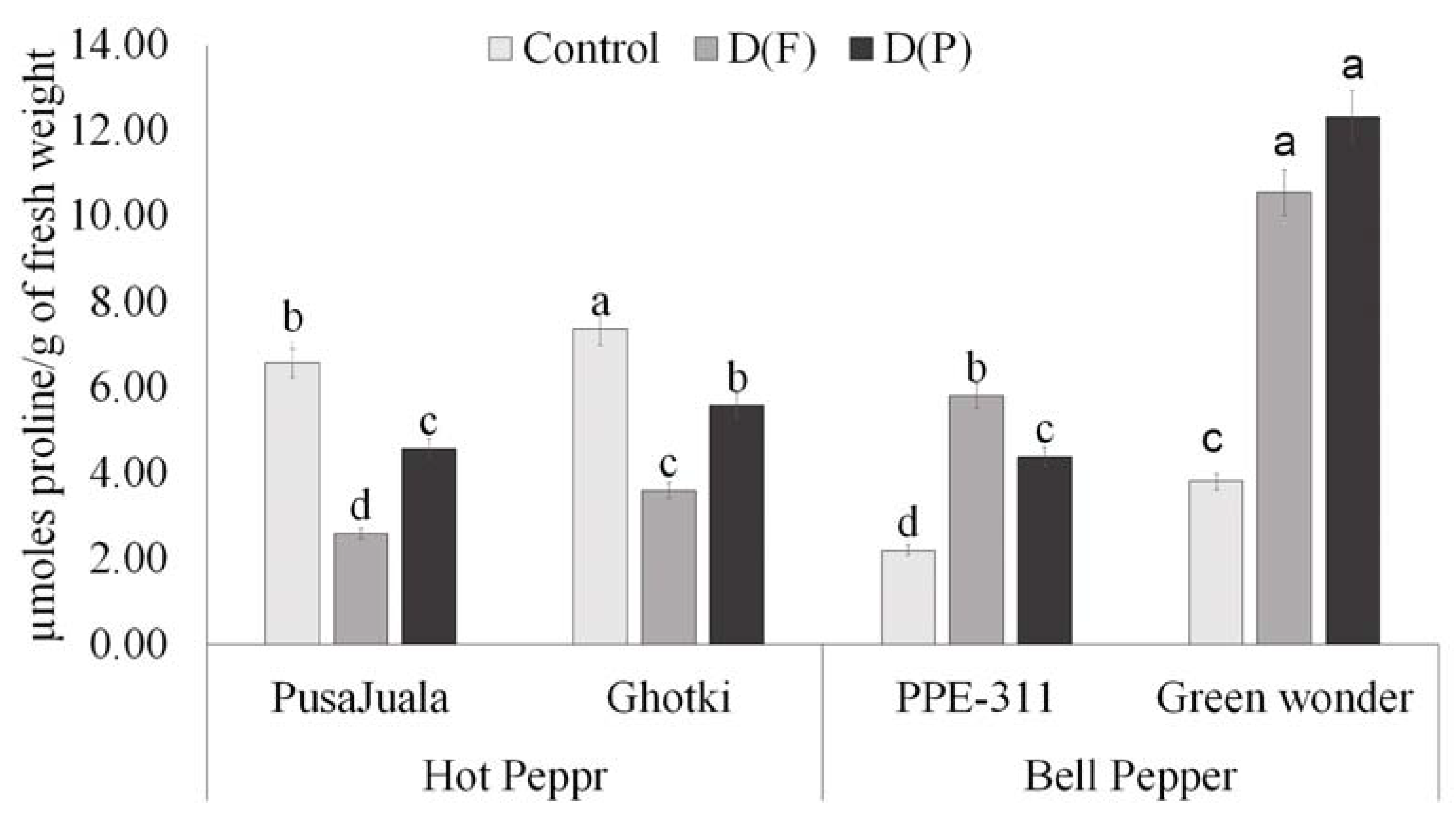
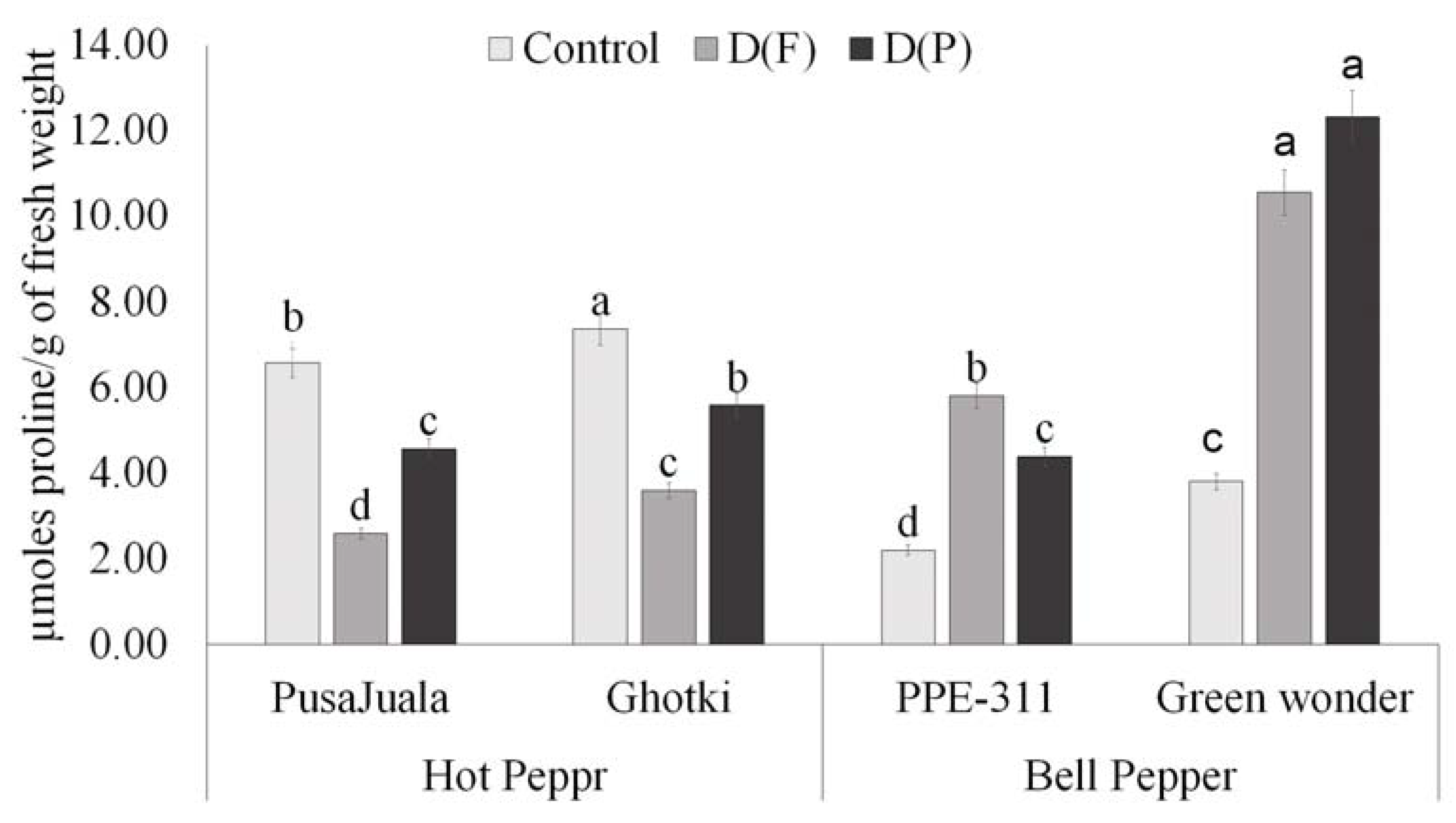
3.5.4. Proline Contents
3.5.5. Capsaicin Contents
3.5.6. Antioxidant Enzyme Activity
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lee, Y.; Howard, L.R.; Villalón, B. Flavonoids and Antioxidant Activity of Fresh Pepper (Capsicum Annuum) Cultivars. J. Food Sci. 1995, 60, 473–476. [Google Scholar] [CrossRef]
- Howard, L.R.; Talcott, S.T.; Brenes, C.H.; Villalon, B. Changes in Phytochemical and Antioxidant Activity of Selected Pepper Cultivars (Capsicum Species) as Influenced by Maturity. J. Agric. Food Chem. 2000, 48, 1713–1720. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.J.; Crosby, K.M.; Pike, L.M.; Yoo, K.S.; Leskovar, D.I. Impact of Genetic and Environmental Variation on Development of Flavonoids and Carotenoids in Pepper (Capsicum Spp.). Sci. Hortic. 2005, 106, 341–352. [Google Scholar] [CrossRef]
- Kidmose, U.; Yang, R.-Y.; Thilsted, S.; Christensen, L.P.; Brandt, K. Content of carotenoids in commonly consumed Asian vegetables and stability and extractability during frying. J. Food Compos. Anal. 2006, 19, 562–571. [Google Scholar] [CrossRef]
- Nishino, H.; Murakoshi, M.; Ii, T.; Takemura, M.; Kuchide, M.; Kanazawa, M.; Yang Mou, X.; Wada, S.; Masuda, M.; Ohsaka, Y.; et al. Carotenoids in Cancer Chemoprevention. Cancer Metastasis Rev. 2002, 21, 257–264. [Google Scholar] [CrossRef]
- Nishino, H.; Murakoshi, M.; Tokuda, H.; Satomi, Y. Cancer Prevention by Carotenoids. Arch. Biochem. Biophys. 2009, 483, 165–168. [Google Scholar] [CrossRef] [PubMed]
- Asplund, K. Antioxidant Vitamins in the Prevention of Cardiovascular Disease: A Systematic Review. J. Intern. Med. 2002, 251, 372–392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Byers, T.; Perry, G. Dietary Carotenes, Vitamin C, and Vitamin E as Protective Antioxidants in Human Cancers. Annu. Rev. Nutr. 1992, 12, 139–159. [Google Scholar] [CrossRef] [PubMed]
- Cichewicz, R.H.; Thorpe, P.A. The Antimicrobial Properties of Chile Peppers (Capsicum Species) and Their Uses in Mayan Medicine. J. Ethnopharmacol. 1996, 52, 61–70. [Google Scholar] [CrossRef]
- Wahba, N.M.; Ahmed, A.S.; Ebraheim, Z.Z. Antimicrobial Effects of Pepper, Parsley, and Dill and Their Roles in the Microbiological Quality Enhancement of Traditional Egyptian Kareish Cheese. Foodborne Pathog. Dis. 2010, 7, 411–418. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, T.; Iwai, K. Chapter 4 Constituents of Red Pepper Species: Chemistry, Biochemistry, Pharmacology, and food Science of the Pungent Principle of Capsicum Species. Alkaloids Chem. Pharmacol. 1984, 3, 227–299. [Google Scholar]
- Ting, Y.S.; Chang, Y.-Y. Ni-Lun Capsaicin Biosynthesis in Water-Stressed Hot Pepper Fruits. Bot. Bull. Acad. Sin. 2005, 46, 25–42. [Google Scholar]
- Aza-González, C.; Núñez-Palenius, H.G.; Ochoa-Alejo, N. Molecular Biology of Capsaicinoid Biosynthesis in Chili Pepper (Capsicum Spp.). Plant Cell Rep. 2011, 30, 695–706. [Google Scholar] [CrossRef] [PubMed]
- Sein, Z.M.M.; Zhi, X. Interannual Variability of Summer Monsoon Rainfall over Myanmar. Arab. J. Geosci. 2016, 9, 1–19. [Google Scholar] [CrossRef]
- Nawaz, U.; Shahid, S.; Ahmad, R.; Ibrahim, M.U.; Wajid, A.; Manan, A.; Ibrar, M. Qureshi Indus Basin Irrigation System and Climatic Trends Regarding Glacial Regimes and Water Runoff. J. Environ. Agric. Sci. 2016, 9, 28–36. [Google Scholar]
- Mahmood, T.; Khalid, S.; Abdullah, M.; Ahmed, Z.; Shah, M.K.N.; Ghafoor, A.; Du, X.; Kausar, M.; Shah, N.; Ghafoor, A.; et al. Insights into Drought Stress Signaling in Plants and the Molecular Genetic Basis of Cotton Drought Tolerance. Cells 2020, 9, 105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahmood, T.; Abdullah, M.; Ahmar, S.; Yasir, M.; Iqbal, M.S.; Yasir, M.; Rehman, S.U.; Ahmed, S.; Rana, R.M.; Ghafoor, A.; et al. Incredible Role of Osmotic Adjustment in Grain Yield Sustainability under Water Scarcity Conditions in Wheat (Triticum Aestivum L.). Plants 2020, 9, 1208. [Google Scholar] [CrossRef]
- Khaliq, T.; Ullah, A.; Ahmad, M.; Hussain, N.; Ali, A.; Ahmad, A. Quantification of Root-Shoot Development and Water Use Efficiency in Autumn Maize (Zea Mays L.) Under Different Irrigation Strategies. J. Environ. Agric. Sci. 2016, 6, 16–22. [Google Scholar]
- Hussain, S.; Iqbal, M.M.; Iqbal, M.; Aziz, O.; Murtaza, D.G.; Iqbal, S.; Mehmood, S. Effect of Different Irrigation Practices and Plastic Mulch on Water Use Efficiency, Growth and Yield of Spring Maize. Basic Res. J. 2015, 4. [Google Scholar]
- Jangid, K.K.; Dwivedi, P. Physiological Responses of Drought stress in Tomato: A Review. Int. J. Agric. Environ. Biotechnol. 2016, 9, 53. [Google Scholar] [CrossRef]
- Gaikwad, S.E.; Shinde, R.A.; Thoke, R.B.; Aparadh, V.T. Potential of Drought Stress in Two Varieties of Capsicum Annum Grown in Maharashtra. Int. J. Res. Pharm. Chem. 2013, 3, 303–307. [Google Scholar]
- Hall, R.D.; Holden, M.A.; Yeoman, M.M. The Accumulation of Phenylpropanoid and Capsaicinoid Compounds in Cell Cultures and Whole Fruit of the Chilli Pepper, Capsicum Frutescens Mill. Plant Cell Tissue Organ Cult. 1987, 8, 163–176. [Google Scholar] [CrossRef]
- Kopta, T.; Sekara, A.; Pokluda, R.; Ferby, V.; Caruso, G. Screening of Chilli Pepper Genotypes as a Source of Capsaicinoids and Antioxidants under Conditions of Simulated Drought Stress. Plants 2020, 9, 364. [Google Scholar] [CrossRef] [Green Version]
- Jeeatid, N.; Techawongstien, S.; Suriharn, B.; Chanthai, S.; Bosland, P.W.; Techawongstien, S. Influence of Water Stresses on Capsaicinoid Production in Hot Pepper (Capsicum Chinense Jacq.) Cultivars with Different Pungency Levels. Food Chem. 2018, 245, 792–797. [Google Scholar] [CrossRef] [PubMed]
- Phimchan, P.; Chanthai, S.; Bosland, P.W.; Techawongstien, S. Enzymatic Changes in Phenylalanine Ammonia-Lyase, Cinnamic-4-Hydroxylase, Capsaicin Synthase, and Peroxidase Activities in Capsicum under Drought Stress. J. Agric. Food Chem. 2014, 62, 7057–7062. [Google Scholar] [CrossRef]
- Zamljen, T.; Zupanc, V.; Slatnar, A. Influence of Irrigation on Yield and Primary and Secondary Metabolites in Two Chilies Species, Capsicum Annuum L. and Capsicum Chinense Jacq. Agric. Water Manag. 2020, 234, 106104. [Google Scholar] [CrossRef]
- Reddy, V. Capsaicin and colour extraction from different verities of green and red chilli peppers of andhrapradesh. Int. J. Adv. Sci. Tech. Res. 2013, 2. [Google Scholar]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid Determination of Free Proline for Water-Stress Studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Ruley, A.T.; Sharma, N.C.; Sahi, S.v. Antioxidant Defense in a Lead Accumulating Plant, Sesbania Drummondii. Plant Physiol. Biochem. 2004, 42, 899–906. [Google Scholar] [CrossRef]
- Aebi, H. Catalase in Vitro. Methods Enzymol. 1984, 105, 121–126. [Google Scholar] [CrossRef]
- Steel, R.G.D. Principles and Procedures of Statistics: A Biometrical Approach; WCB McGraw-Hill series in probability and statistics; McGraw-Hill: New York, NY, USA, 1980. [Google Scholar]
- Lau, N.R.; Lara, F.M.; García, Y.M.; Moreno, Z.; Antonio, A.G.; Machado, I.E.; Estévez, M.M. Water Deficit Affects the Accumulation of Capsaicinoids in Fruits of Capsicum Chinense Jacq. Am. Soc. Hortic. Sci. 2011, 46, 487–492. [Google Scholar]
- Hussain, H.A.; Men, S.; Hussain, S.; Chen, Y.; Ali, S.; Zhang, S.; Zhang, K.; Li, Y.; Xu, Q.; Liao, C.; et al. Interactive Effects of Drought and Heat Stresses on Morpho-Physiological Attributes, Yield, Nutrient Uptake and Oxidative Status in Maize Hybrids. Sci. Rep. 2019, 9, 3890. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zafar, S.A.; Hameed, A.; Nawaz, M.A.; MA, W.; Noor, M.A.; Hussain, M. Mehboob-ur-Rahman Mechanisms and Molecular Approaches for Heat Tolerance in Rice (Oryza Sativa, L.) under Climate Change Scenario. J. Integr. Agric. 2018, 17, 726–738. [Google Scholar] [CrossRef]
- Mahendran, S.; Bandara, D.C. Effects of soil moisture stress at different growth stages on vitamin C, capsaicin and β-carotene contents of chilli (Capsicum annuum L.) fruits and their impact on yield. Tropical Agric. Res. 2000, 12, 95–106. [Google Scholar]
- Panchal, S.C.; Bhatnagar, R.; Momin, R.A.; Chauhan, N.P. Capsaicin and Ascorbic Acid Content of Chilli as Influenced by Cultural Practices. CAPSICUM Amp. Eggplant Newsl. 2001, 20, 19–22. [Google Scholar]
- Ashraf, M. Inducing Drought Tolerance in Plants: Recent Advances. Biotechnol. Adv. 2010, 28, 169–183. [Google Scholar] [CrossRef]
- Chen, T.H.H.; Murata, N. Glycinebetaine: An Effective Protectant against Abiotic Stress in Plants. Trends Plant Sci. 2008, 13, 499–505. [Google Scholar] [CrossRef]
- Verbruggen, N.; Hermans, C. Proline Accumulation in Plants: A Review. Amino Acids 2008, 35, 753–759. [Google Scholar] [CrossRef]
- Munns, R.; Tester, M. Mechanisms of Salinity Tolerance. Annu. Rev. Plant Biol. 2008, 59, 651–681. [Google Scholar] [CrossRef] [Green Version]
- Szabados, L.; Savouré, A. Proline: A Multifunctional Amino Acid. Trends Plant Sci. 2010, 15, 89–97. [Google Scholar] [CrossRef]
- Kaur, K.; Kaur, N.; Gupta, A.K.; Singh, I. Exploration of the Antioxidative Defense System to Characterize Chickpea Genotypes Showing Differential Response towards Water Deficit Conditions. Plant Growth Regul. 2013, 70, 49–60. [Google Scholar] [CrossRef]
- Sikha, S.; Sunil, P.; Jamwal, A.J.; Bhattacharya, S.; Bhattacharya, S. Impact of Water-Deficit and Salinity Stress on Seed Germination and Seedling Growth of Capsicum Annuum “Solan Bharpur”. Int. Res. J. Biol. Sci. 2013, 2, 9–15. [Google Scholar]
- Bennett, D.J.; Kirby, G.W. Constitution and Biosynthesis of Capsaicin. J. Chem. Soc. C Org. 1968, 442–446. [Google Scholar] [CrossRef]
- Iwai, K.; Suzuki, T.; Fujiwake, H. Formation and Accumulation of Pungent Principle of Hot Pepper Fruits, Capsaicin and Its Analogues, in Capsicum Annuun Var. Annuun Cv. Karayatsubusa at Different Growth Stages after Flowering. Agric. Biol. Chem. 1979, 43, 2493–2498. [Google Scholar] [CrossRef] [Green Version]
- Govindarajan, V.S. Capsicum—Production, Technology, Chemistry, and Quality—Part II. Processed Products, Standards, World Production and Trade. C R C Crit. Rev. Food Sci. Nutr. 1986, 23, 207–288. [Google Scholar] [CrossRef] [PubMed]
- Kosuge, S.; Furuta, M. Studies on the Pungent Principle of Capsicum,: Part Xiv Chemical Constitution of the Pungent Principle. Agric. Biol. Chem. 1970, 34, 248–256. [Google Scholar] [CrossRef]
- Estrada, B.; Pomar, F.; Díaz, J.; Merino, F.; Bernal, M.A. Pungency Level in Fruits of the Padron Pepper with Different Water Supply. Sci. Hortic. 1999, 81, 385–396. [Google Scholar] [CrossRef]
- Hodges, D.M.; Lester, G.E.; Munro, K.D.; Toivonen, P.M.A. Oxidative Stress: Importance for Postharvest Quality. Proc. Hort. Sci. 2004, 39, 924–929. [Google Scholar] [CrossRef] [Green Version]
- Jaimeza, R.E.; Adab, F.R.; Núñezb, C.G. The Effect of Irrigation Frequency on Water and Carbon Relations in Three Cultivars of Sweet Pepper (Capsicum Chinense Jacq), in a Tropical Semiarid Region. Sci. Hortic. 1999, 81, 301–308. [Google Scholar] [CrossRef]
- Behboudian, M.H.; Lawes, G.S.; Griffiths, K.M. The Influence of Water Deficit on Water Relations, Photosynthesis and Fruit Growth in Asian Pear (Pyrus Serotina Rehd.). Sci. Hortic. 1994, 60, 89–99. [Google Scholar] [CrossRef]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mahmood, T.; Rana, R.M.; Ahmar, S.; Saeed, S.; Gulzar, A.; Khan, M.A.; Wattoo, F.M.; Wang, X.; Branca, F.; Mora-Poblete, F.; et al. Effect of Drought Stress on Capsaicin and Antioxidant Contents in Pepper Genotypes at Reproductive Stage. Plants 2021, 10, 1286. https://doi.org/10.3390/plants10071286
Mahmood T, Rana RM, Ahmar S, Saeed S, Gulzar A, Khan MA, Wattoo FM, Wang X, Branca F, Mora-Poblete F, et al. Effect of Drought Stress on Capsaicin and Antioxidant Contents in Pepper Genotypes at Reproductive Stage. Plants. 2021; 10(7):1286. https://doi.org/10.3390/plants10071286
Chicago/Turabian StyleMahmood, Tahir, Rashid Mehmood Rana, Sunny Ahmar, Saima Saeed, Asma Gulzar, Muhammad Azam Khan, Fahad Masoud Wattoo, Xiukang Wang, Ferdinando Branca, Freddy Mora-Poblete, and et al. 2021. "Effect of Drought Stress on Capsaicin and Antioxidant Contents in Pepper Genotypes at Reproductive Stage" Plants 10, no. 7: 1286. https://doi.org/10.3390/plants10071286
APA StyleMahmood, T., Rana, R. M., Ahmar, S., Saeed, S., Gulzar, A., Khan, M. A., Wattoo, F. M., Wang, X., Branca, F., Mora-Poblete, F., Mafra, G. S., & Du, X. (2021). Effect of Drought Stress on Capsaicin and Antioxidant Contents in Pepper Genotypes at Reproductive Stage. Plants, 10(7), 1286. https://doi.org/10.3390/plants10071286