


Potential Ecological Distributions of Urban Adapters and Urban Exploiters for the Sustainability of the Urban Bird Network

 , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Bird and Vegetation Survey
2.3. Analysis
Species Distribution Modeling
3. Results
4. Discussion
4.1. Urban Bird Community
4.2. Predicted Distributions of Urban Adapters and Urban Exploiters
4.3. Improvement of Urban Ecosystem
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Setiawan, H.; Mathieu, R.; Thompson-Fawcett, M. Assessing the Applicability of the V–I–S Model to Map Urban Land Use in the Developing World: Case Study of Yogyakarta, Indonesia. Comput. Environ. Urban Syst. 2006, 30, 503–522. [Google Scholar] [CrossRef]
- Lepczyk, C.A.; Sorte, F.A.L.; Aronson, M.F.; Goddard, M.A.; MacGregor-Fors, I.; Nilon, C.H.; Warren, P.S. Global Patterns and Drivers of Urban Bird Diversity. In Ecology and Conservation of Birds in Urban Environments; Springer: Berlin/Heidelberg, Germany, 2017; pp. 13–33. [Google Scholar]
- Zain, A.F.M.; Permatasari, P.A.; Ainy, C.N.; Destriana, N.; Mulyati, D.F.; Edi, S. The Detection of Urban Open Space at Jakarta, Bogor, Depok, and Tangerang—Indonesia by Using Remote Sensing Technique for Urban Ecology Analysis. Procedia Environ. Sci. 2015, 24, 87–94. [Google Scholar] [CrossRef][Green Version]
- McKinney, M.L. Urbanization, Biodiversity, and Conservation: The Impacts of Urbanization on Native Species Are Poorly Studied, but Educating a Highly Urbanized Human Population about These Impacts Can Greatly Improve Species Conservation in All Ecosystems. Bioscience 2002, 52, 883–890. [Google Scholar] [CrossRef]
- Whitney, G.G. A Quantitative Analysis of the Flora and Plant Communities of a Representative Midwestern US Town. Urban Ecol. 1985, 9, 143–160. [Google Scholar] [CrossRef]
- Blair, R.B. Land Use and Avian Species Diversity Along an Urban Gradient. Ecol. Appl. 1996, 6, 506–519. [Google Scholar] [CrossRef]
- Lancaster, R.K.; Rees, W.E. Bird Communities and the Structure of Urban Habitats. Can. J. Zool. 1979, 57, 2358–2368. [Google Scholar] [CrossRef]
- Chace, J.F.; Walsh, J.J. Urban Effects on Native Avifauna: A Review. Landsc. Urban Plan. 2006, 74, 46–69. [Google Scholar] [CrossRef]
- Wenny, D.G.; Devault, T.L.; Johnson, M.D.; Kelly, D.; Sekercioglu, C.H.; Tomback, D.F.; Whelan, C.J. The Need to Quantify Ecosystem Services Provided by Birds. Auk 2011, 128, 1–14. [Google Scholar] [CrossRef]
- Cox, D.T.; Hudson, H.L.; Plummer, K.E.; Siriwardena, G.M.; Anderson, K.; Hancock, S.; Devine-Wright, P.; Gaston, K.J. Covariation in Urban Birds Providing Cultural Services or Disservices and People. J. Appl. Ecol. 2018, 55, 2308–2319. [Google Scholar] [CrossRef]
- Mardiastuti, A.; Mulyani, Y.A.; Rinaldi, D.; Rumblat, W.; Dewi, L.K.; Kaban, A.; Sastranegara, H. Synurbic Avian Species in Greater Jakarta Area, Indonesia. IOP Conf. Ser. Earth Environ. Sci. 2020, 457, 012001. [Google Scholar] [CrossRef]
- Kark, S.; Iwaniuk, A.; Schalimtzek, A.; Banker, E. Living in the City: Can Anyone Become an ‘urban Exploiter’? J. Biogeogr. 2007, 34, 638–651. [Google Scholar] [CrossRef]
- Seto, K.C.; Fleishman, E.; Fay, J.P.; Betrus, C.J. Linking Spatial Patterns of Bird and Butterfly Species Richness with Landsat Tm Derived Ndvi. Int. J. Remote Sens. 2004, 25, 4309–4324. [Google Scholar] [CrossRef]
- Tursilowati, L.; Tetuko Sri Sumantyo, J.; Kuze, H.; Adiningsih, E.S. Relationship between Urban Heat Island Phenomenon and Land Use/Land Cover Changes in Jakarta-Indonesia. J. Emerg. Trends Eng. Appl. Sci. 2012, 3, 645–653. [Google Scholar]
- Lim, H.C.; Sodhi, N.S. Responses of Avian Guilds to Urbanisation in a Tropical City. Landsc. Urban Plan. 2004, 66, 199–215. [Google Scholar] [CrossRef]
- Geschke, A.; James, S.; Bennett, A.F.; Nimmo, D.G. Compact Cities or Sprawling Suburbs? Optimising the Distribution of People in Cities to Maximise Species Diversity. J. Appl. Ecol. 2018, 55, 2320–2331. [Google Scholar] [CrossRef]
- Conole, L.E.; Kirkpatrick, J.B. Functional and Spatial Differentiation of Urban Bird Assemblages at the Landscape Scale. Landsc. Urban Plan. 2011, 100, 11–23. [Google Scholar] [CrossRef]
- Szulkin, M.; Garroway, C.J.; Corsini, M.; Kotarba, A.Z.; Dominoni, D.; Szulkin, M. How to Quantify Urbanization When Testing for Urban Evolution? In Urban Evolutionary Biology; Oxford University Press: Oxford, UK, 2020; pp. 13–35. [Google Scholar]
- Mildawani, I.; Rahman, A. Household Gardening as Preference Activities to Cupport Community Resilience during Pandemic COVID-19. J. Synerg. Landsc. 2021, 1, 64–79. [Google Scholar] [CrossRef]
- Santosa, E.; Susila, A.D.; Widodo, W.D.; Nasrullah, N.; Ruwaida, I.P.; Sari, R. Exploring Fruit Tree Species as Multifunctional Greenery: A Case of Its Distribution in Indonesian Cities. Sustainability 2021, 13, 7835. [Google Scholar] [CrossRef]
- Shanahan, D.F.; Miller, C.; Possingham, H.P.; Fuller, R.A. The Influence of Patch Area and Connectivity on Avian Communities in Urban Revegetation. Biol. Conserv. 2011, 144, 722–729. [Google Scholar] [CrossRef]
- Kang, W.; Minor, E.S.; Park, C.-R.; Lee, D. Effects of Habitat Structure, Human Disturbance, and Habitat Connectivity on Urban Forest Bird Communities. Urban Ecosyst. 2015, 18, 857–870. [Google Scholar] [CrossRef]
- Pradipta, E.; Kholis, N.; Lestari, F.; Lestari, R.; Bowolaksono, A. Reassessing the Benefits and Costs of Universitas Indonesia’s Urban Forest Zonation. IOP Conf. Ser. Mater. Sci. Eng. 2019, 546, 022015. [Google Scholar] [CrossRef]
- Setiowati, R.; Hasibuan, H.S.; Koestoer, R.H.; Harmain, R. Planning for Urban Green Area and Its Importance for Sustainability: The Case of Jakarta. IOP Conf. Ser. Earth Environ. Sci. 2019, 328, 012027. [Google Scholar] [CrossRef]
- Bibby, C.; Burgess, N.; Hill, D.; Mustoe, S. Bird Census Technique, 2nd ed.; Academic Press: London, UK, 2000. [Google Scholar]
- Open Data Kit. Open Data Kit Documentation (2017); Open Data Kit: Fullerton, CA, USA, 2018. [Google Scholar]
- Patrignani, A.; Ochsner, T.E. Canopeo: A Powerful New Tool for Measuring Fractional Green Canopy Cover. Agron. J. 2015, 107, 2312–2320. [Google Scholar] [CrossRef]
- Sharma, S.; Sharma, H.P.; Chaulagain, C.; Katuwal, H.B.; Belant, J.L. Estimating Occupancy of Chinese Pangolin (Manis Pentadactyla) in a Protected and Non-Protected Area of Nepal. Ecol. Evol. 2020, 10, 4303–4313. [Google Scholar] [CrossRef] [PubMed]
- Salman, I.N.; Blaustein, L. Vegetation Cover Drives Arthropod Communities in Mediterranean/Subtropical Green Roof Habitats. Sustainability 2018, 10, 4209. [Google Scholar] [CrossRef]
- Winarni, N.L.; Anugra, B.G.; Anisafitri, S.; Kaunain, N.N.; Pradana, D.H. Fieldwork during Pandemic: Backyard Bird Survey and Making Student’s Biological Field Practice Works. Biodiversitas J. Biol. Divers. 2021, 22, 4. [Google Scholar] [CrossRef]
- Lambert, F.R.; Collar, N.J. The Future for Sundaic Lowland Forest Birds: Long-Term Effects of Commercial Logging and Fragmentation. Forktail 2002, 18, 127–146. [Google Scholar]
- Cristaldi, M.A.; Giraudo, A.R.; Arzamendia, V.; Bellini, G.P.; Claus, J. Urbanization Impacts on the Trophic Guild Composition of Bird Communities. J. Nat. Hist. 2017, 51, 2385–2404. [Google Scholar] [CrossRef]
- Phillips, S.J.; Anderson, R.P.; Dudík, M.; Schapire, R.E.; Blair, M.E. Opening the Black Box: An Open-Source Release of Maxent. Ecography 2017, 40, 887–893. [Google Scholar] [CrossRef]
- Merow, C.; Smith, M.J.; Silander Jr, J.A. A Practical Guide to MaxEnt for Modeling Species’ Distributions: What It Does, and Why Inputs and Settings Matter. Ecography 2013, 36, 1058–1069. [Google Scholar] [CrossRef]
- Hof, A.R.; Jansson, R.; Nilsson, C. The Usefulness of Elevation as a Predictor Variable in Species Distribution Modelling. Ecol. Model. 2012, 246, 86–90. [Google Scholar] [CrossRef]
- Nieto, S.; Flombaum, P.; Garbulsky, M.F. Can Temporal and Spatial NDVI Predict Regional Bird-Species Richness? Glob. Ecol. Conserv. 2015, 3, 729–735. [Google Scholar] [CrossRef]
- Coxen, C.L.; Frey, J.K.; Carleton, S.A.; Collins, D.P. Species Distribution Models for a Migratory Bird Based on Citizen Science and Satellite Tracking Data. Glob. Ecol. Conserv. 2017, 11, 298–311. [Google Scholar] [CrossRef]
- Booth, T.H. Why Understanding the Pioneering and Continuing Contributions of BIOCLIM to Species Distribution Modelling Is Important. Austral Ecol. 2018, 43, 852–860. [Google Scholar] [CrossRef]
- Ermida, S.L.; Soares, P.; Mantas, V.; Göttsche, F.-M.; Trigo, I.F. Google Earth Engine Open-Source Code for Land Surface Temperature Estimation from the Landsat Series. Remote Sens. 2020, 12, 1471. [Google Scholar] [CrossRef]
- Çoban, H.O.; Örücü, Ö.K.; Arslan, E.S. MaxEnt Modeling for Predicting the Current and Future Potential Geographical Distribution of Quercus Libani Olivier. Sustainability 2020, 12, 2671. [Google Scholar] [CrossRef]
- Hasibuan, H.S.; Soemardi, T.P.; Koestoer, R.; Moersidik, S. The Role of Transit Oriented Development in Constructing Urban Environment Sustainability, the Case of Jabodetabek, Indonesia. Procedia Environ. Sci. 2014, 20, 622–631. [Google Scholar] [CrossRef]
- Nor, A.N.M.; Corstanje, R.; Harris, J.A.; Grafius, D.R.; Siriwardena, G.M. Ecological Connectivity Networks in Rapidly Expanding Cities. Heliyon 2017, 3, e00325. [Google Scholar] [CrossRef]
- Carbó-Ramírez, P.; Zuria, I. The Value of Small Urban Greenspaces for Birds in a Mexican City. Landsc. Urban Plan. 2011, 100, 213–222. [Google Scholar] [CrossRef]
- van Heezik, Y.; Smyth, A.; Mathieu, R. Diversity of Native and Exotic Birds across an Urban Gradient in a New Zealand City. Landsc. Urban Plan. 2008, 87, 223–232. [Google Scholar] [CrossRef]
- MacKinnon, J.; Phillipps, K. A Field Guide to the Birds of Borneo, Sumatra, Java, and Bali; Oxford University Press: New York, NY, USA, 1993. [Google Scholar]
- Rahman, M.A.; Moser, A.; Rötzer, T.; Pauleit, S. Comparing the Transpirational and Shading Effects of Two Contrasting Urban Tree Species. Urban Ecosyst. 2019, 22, 683–697. [Google Scholar] [CrossRef]
- Yap, C.A.; Sodhi, N.S. Southeast Asian Invasive Birds: Ecology, Impact and Management. Ornithol. Sci. 2004, 3, 57–67. [Google Scholar] [CrossRef]
- Zhou, B.; Liu, J.; Liang, W. Breeding in a Noisy World: Attraction to Urban Arterial Roads and Preference for Nest-Sites by the Scaly-Breasted Munia (Lonchura Punctulata). Glob. Ecol. Conserv. 2020, 22, e00987. [Google Scholar] [CrossRef]
- Fleming, T.H.; Williams, C.F.; Bonaccorso, F.J.; Herbst, L.H. Phenology, Seed Dispersal, and Colonization in Muntingia Calabura, a Neotropical Pioneer Tree. Am. J. Bot. 1985, 72, 383–391. [Google Scholar] [CrossRef]
- Sufian, A.S.; Ramasamy, K.; Ahmat, N.; Zakaria, Z.A.; Yusof, M.I.M. Isolation and Identification of Antibacterial and Cytotoxic Compounds from the Leaves of Muntingia Calabura L. J. Ethnopharmacol. 2013, 146, 198–204. [Google Scholar] [CrossRef]
- Sarwadi, A.; Irwan, S.N.R.; Utami, R.N.; Raya, A.B. Study on Roadside Greenery in Yogyakarta City towards Development of Productive Urban Landscape. IOP Conf. Ser. Earth Environ. Sci. 2019, 361, 012008. [Google Scholar] [CrossRef]
- Hawkeswood, T.J.; Sommung, B. Pollination of Muntingia Calabura L. (Muntingiaceae) by Native Bees in Bangkok, Thailand 2016. Calodema 2016, 421, 1–6. [Google Scholar]
- Yadav, R.; Pandey, P. Assessment of Air Pollution Tolerance Index (APTI) and Anticipated Performance Index (API) of Roadside Plants for the Development of Greenbelt in Urban Area of Bathinda City, Punjab, India. Bull. Environ. Contam. Toxicol. 2020, 105, 906–914. [Google Scholar] [CrossRef]
- Kazi, E.; Shaikh, Y.I.; Tikle, S.; Shaikh, G.B.V.S.; Kulkarni, S.; Dhulap, V.P. Effect of Air Pollutants on Urban Roadside Trees in Shivajinagar & Pashan Area, Pune City. ES Food Agrofor. 2021, 5, 29–37. [Google Scholar]
- Pradana, D.H.; Mardiastuti, A.; Yasman, Y. Utilization of Ficus Benjamina by Birds at Urban Habitat in Depok. Bioma Berk. Ilm. Biol. 2019, 20, 75–78. [Google Scholar] [CrossRef]
- Putri, F.A.; Basukriadi, A.; Pradana, D.H. Study of the Bird Community in Rubber Plantations at Universitas Indonesia, Depok. AIP Conf. Proc. 2020, 2242, 050020. [Google Scholar]
- Nugraha, B.P. Efek Gradien Urbanisasi Habitat Terhadap Komunitas Burung Di Kampus Universitas Indonesia, Depok, Jawa Barat; Skripsi, Universitas Indonesia: Depok, Indonesia, 2014. [Google Scholar]
- Melletti, M.; Penteriani, V. Nesting and Feeding Tree Selection in the Endangered White-Backed Woodpecker, Dendrocopos Leucotos Lilfordi. Wilson J. Ornithol. 2003, 115, 299–306. [Google Scholar] [CrossRef]
- Nurjaman, D.; Husodo, T.; Megantara, E.N.; Hadikusumah, H.Y.; Wulandari, I. Relationship of Bird Diversity and Plant Composition Inside The Area Campus Green Space of Universitas Padjadjaran Jatinangor, Sumedang West Java. Biost. J. Biol. Biol. Educ. 2018, 10, 500–509. [Google Scholar] [CrossRef]
- Dwyer, J.F.; McPherson, E.G.; Schroeder, H.W.; Rowntree, R.A. Assessing the Benefits and Costs of the Urban Forest. J. Arboric. 1992, 18, 227. [Google Scholar] [CrossRef]
- Fernandez-Juricic, E.; Jokimäki, J. A Habitat Island Approach to Conserving Birds in Urban Landscapes: Case Studies from Southern and Northern Europe. Biodivers. Conserv. 2001, 10, 2023–2043. [Google Scholar] [CrossRef]
- Palacio, F.X. Urban Exploiters Have Broader Dietary Niches than Urban Avoiders. Ibis 2020, 162, 42–49. [Google Scholar] [CrossRef]
- Ikin, K.; Knight, E.; Lindenmayer, D.B.; Fischer, J.; Manning, A.D. The Influence of Native versus Exotic Streetscape Vegetation on the Spatial Distribution of Birds in Suburbs and Reserves. Divers. Distrib. 2013, 19, 294–306. [Google Scholar] [CrossRef]
- Xie, S.; Marzluff, J.M.; Su, Y.; Wang, Y.; Meng, N.; Wu, T.; Gong, C.; Lu, F.; Xian, C.; Zhang, Y. The Role of Urban Waterbodies in Maintaining Bird Species Diversity within Built Area of Beijing. Sci. Total Environ. 2021, 806, 150430. [Google Scholar] [CrossRef]
- Ferenc, M.; Sedláček, O.; Fuchs, R. How to Improve Urban Greenspace for Woodland Birds: Site and Local-Scale Determinants of Bird Species Richness. Urban Ecosyst. 2014, 17, 625–640. [Google Scholar] [CrossRef]
- Pratiwi, A.D.; Wardhana, W.; Putrika, A. Community Structure of Tree at Three Location Points of Ciliwung Riparian Zone in Depok–South Jakarta. Phys. Conf. Ser. 2021, 1725, 012036. [Google Scholar] [CrossRef]
- Mardiastuti, A.; Mulyani, Y.A.; Kusrini, M.D. Bird Visit to Ficus Benjamina in Two Urbanization Gradients in the Tropics. IOP Conf. Ser. Earth Environ. Sci. 2021, 948, 012061. [Google Scholar] [CrossRef]
- Bartholomew, G.A.; Cade, T.J. The Water Economy of Land Birds. Auk 1963, 80, 504–539. [Google Scholar] [CrossRef]
- Zivanovic, A.J.; Luck, G.W. Social and Environmental Factors Drive Variation in Plant and Bird Communities across Urban Greenspace in Sydney, Australia. J. Environ. Manag. 2016, 169, 210–222. [Google Scholar] [CrossRef] [PubMed]
- Leveau, L.M.; Isla, F.I.; Bellocq, M.I. Predicting the Seasonal Dynamics of Bird Communities along an Urban-Rural Gradient Using NDVI. Landsc. Urban Plan. 2018, 177, 103–113. [Google Scholar] [CrossRef]
- Rahman, S.F.S.A.; Nor, H.M.; Burhanuddin, M. Diversity of the Insects in the Diet of Edible Nest Swiftlets in Oil Palm Plantations. J Biodivers. Env. Sci 2016, 8, 39–48. [Google Scholar]
- Sullivan, S.M.P.; Corra, J.W.; Hayes, J.T. Urbanization Mediates the Effects of Water Quality and Climate on a Model Aerial Insectivorous Bird. Ecol. Monogr. 2021, 91, e01442. [Google Scholar] [CrossRef]
- Thorburn, C. The Edible Birds’ Nest Boom in Indonesia and South-East Asia: A Nested Political Ecology. Food Cult. Soc. 2014, 17, 535–553. [Google Scholar] [CrossRef]
- Lack, D. A Review of the Genera and Nesting Habits of Swifts. Auk 1956, 73, 1–32. [Google Scholar] [CrossRef]
- Debataraja, I.B.; Pradana, D.H.; Winarni, N.L. The Impact of Landuse and the Relationship between NDVI on the Bird Species Richness in Sukmajaya District, Depok. IOP Conf. Ser. Earth Environ. Sci. 2021, 846, 012004. [Google Scholar] [CrossRef]
- Mardiastuti, A.; Putra, M.S.K.; Mulyani, Y.A. Bird Diversity Patterns along Urbanization Gradients: Some Recommendations for Landscape Planner. IOP Conf. Ser. Earth Environ. Sci. 2020, 556, 012012. [Google Scholar] [CrossRef]
- Nor, A.N.M.; Corstanje, R.; Harris, J.A.; Brewer, T. Impact of Rapid Urban Expansion on Green Space Structure. Ecol. Indic. 2017, 81, 274–284. [Google Scholar] [CrossRef]
- Doody, B.J.; Sullivan, J.J.; Meurk, C.D.; Stewart, G.H.; Perkins, H.C. Urban Realities: The Contribution of Residential Gardens to the Conservation of Urban Forest Remnants. Biodivers. Conserv. 2010, 19, 1385–1400. [Google Scholar] [CrossRef]
- Arifin, H.; Nakagoshi, N. Landscape Ecology and Urban Biodiversity in Tropical Indonesian Cities. Landsc. Ecol. Eng. 2011, 7, 33–43. [Google Scholar] [CrossRef]
- Middle, I.; Dzidic, P.; Buckley, A.; Bennett, D.; Tye, M.; Jones, R. Integrating Community Gardens into Public Parks: An Innovative Approach for Providing Ecosystem Services in Urban Areas. Urban For. Urban Green. 2014, 13, 638–645. [Google Scholar]
- Grafius, D.R.; Corstanje, R.; Siriwardena, G.M.; Plummer, K.E.; Harris, J.A. A Bird’s Eye View: Using Circuit Theory to Study Urban Landscape Connectivity for Birds. Landsc. Ecol. 2017, 32, 1771–1787. [Google Scholar] [CrossRef] [PubMed]
- Lal, R. Home Gardening and Urban Agriculture for Advancing Food and Nutritional Security in Response to the COVID-19 Pandemic. Food Secur. 2020, 12, 871–876. [Google Scholar] [CrossRef]
- Montefrio, M.J.F. Interrogating the “Productive” Home Gardener in a Time of Pandemic Lockdown in the Philippines. Food Foodways 2020, 28, 216–225. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Environmental Predictor | Description | Data Source and Reference |
|---|---|---|
| Landscape Greenness | Normalized Difference Vegetation Index (NDVI) in June 2020 | USGS Landsat 8 processed data [36] |
| Land Function | Information was developed based on Depok City Detailed Spatial Planning Documents or RDTR (Rencana Detail Tata Ruang) 2018. | Processed data were based on Detailed Spatial Planning RDTR and visually interpreted from the official website |
| Temperature | Land Surface Temperature of the research area in June 2020 | Landsat 8 processed data automated algorithm from Google Earth Engine [28] |
| Elevation | Elevation in the research area | SRTM downloaded from USGS EarthExplorer (https://earthexplorer.usgs.gov/ (accessed on 24 February 2022)) |
| Distance to Large Green Patches (LGP) | Distance to nearby green patches including urban forests, agriculture areas, and a golf course nearby the research area | Identification of Large Green Patches (LGP) was based on NDVI > 0.6 and land function green open space area [35] |
| Distance to Water | Distance to water including a river and a lake inside the research area. Processed using Euclidean Distance Tool | Processed and rasterized data using Land Function data of water body. |
| Code | Land Function | Description |
|---|---|---|
| 1 | Water Bodies | The large area of water |
| 2 | Roads | Consists of networks of artery and major roads, usually accompanied by a gradient of vegetation along the road edges |
| 3 | Business Districts + Industry and Warehousing | Includes all the business districts following the city’s major roads |
| 4 | Offices | Zoning areas representing small offices located outside business districts |
| 5 | Buffer Area | A mandatory buffer zone for at least 25 m from rivers, beaches, lakes, and water sources. |
| 6 | Settlement and Housing | Middle- to high-density settlements and housing within the research area |
| 7 | Public Service Area | Generally a zone of public services buildings and education services concentrated within Universitas Indonesia Area |
| 8 | Agriculture and Tourism | Usually tourism that uses large areas such as a golf course and estate-level agriculture such as fish ponds |
| 9 | Green Open Space | Generally a zone consisting of parks, urban forests, and cemeteries with a certain degree of vegetation/greenness planted |
| Residential | Green Open Spaces | Roadside | |
|---|---|---|---|
| Number of points | 52 | 19 | 44 |
| Number of bird species | 14 | 9 | 10 |
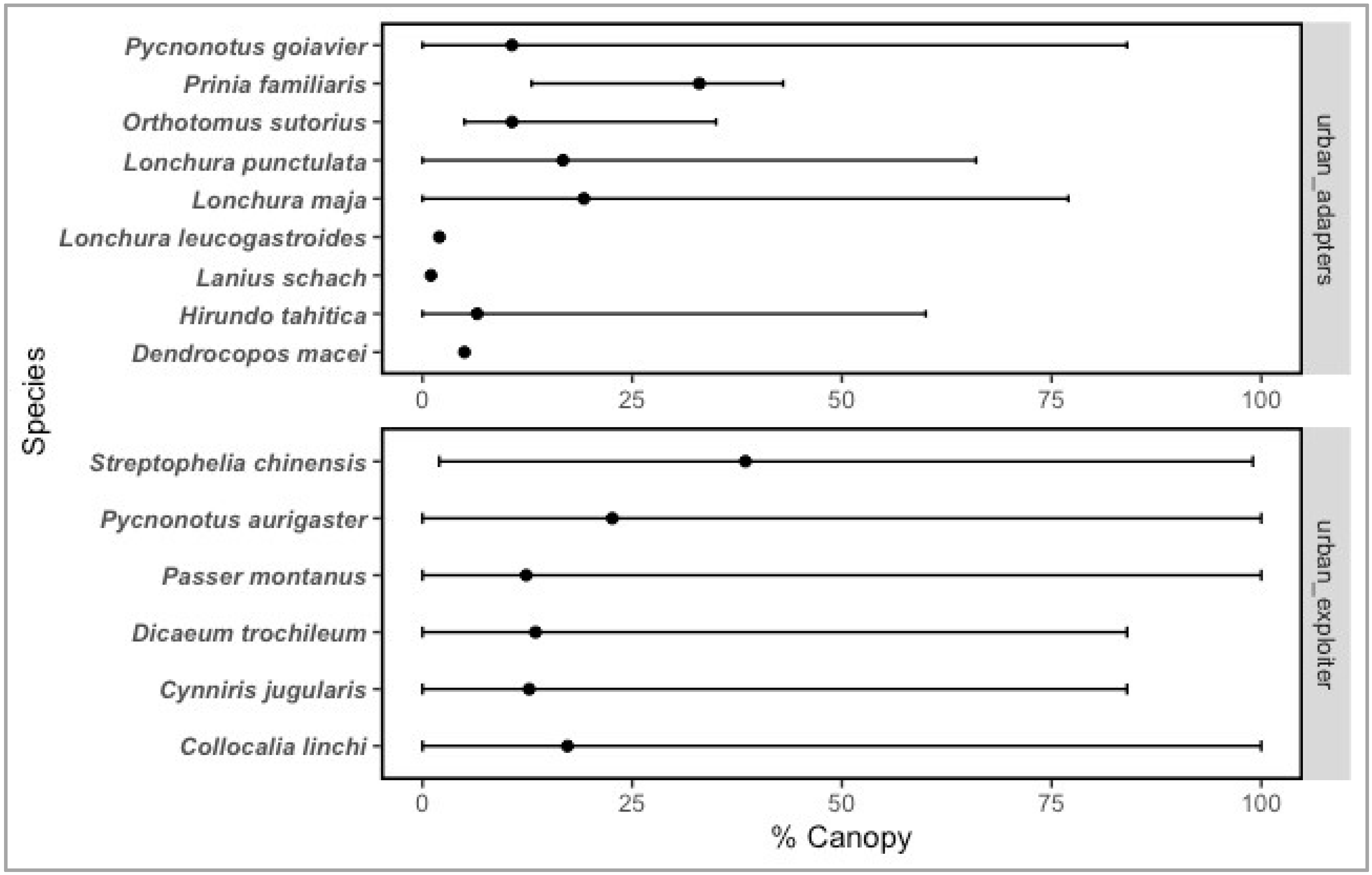
| Average canopy cover (Canopeo) | 17.52 | 20.90 | 11.13 |
| Dominant vegetation | Nephelium lappaceum, Artocarpus heterophyllus, Carica papaya | Swietenia mahagoni, Swietenia macrophylla, Cerbera manghas | Muntingia calabura, Polyalthia longifolia, Gnetum gnemon |
| Species | Trophic Guild | Houses | Green Spaces | Roadside | |||
|---|---|---|---|---|---|---|---|
| Abund. | Freq. | Abund. | Freq. | Abun. | Freq. | ||
| Urban adapters | |||||||
| Dendrocopos macei | bark insectivores | 4 | 0.06 | 0.00 | 0.00 | ||
| Hirundo tahitica | aerial insectivores | 27 | 0.42 | 2 | 0.29 | 11 | 0.29 |
| Lanius schach | carnivore/insectivore | 1 | 0.02 | 0.00 | 0.00 | ||
| Lonchura leucogastroides | foliage granivore | 1 | 0.02 | 0.00 | 0.00 | ||
| Lonchura maja | foliage granivore | 2 | 0.03 | 0.00 | 1 | 0.03 | |
| Lonchura punctulata | foliage granivore | 0.00 | 1 | 0.14 | 0.00 | ||
| Orthotomus sutorius | foliage insectivore | 6 | 0.09 | 0.00 | 0.00 | ||
| Prinia familiaris | foliage insectivore | 2 | 0.03 | 0.00 | 1 | 0.03 | |
| Pycnonotus goiavier | foliage insectivore/frugivore | 21 | 0.32 | 4 | 0.57 | 25 | 0.66 |
| Urban exploiters | |||||||
| Collocalia linchi | aerial insectivores | 83 | 0.37 | 17 | 0.39 | 74 | 0.37 |
| Cynniris jugularis | nectarivores | 11 | 0.05 | 1 | 0.02 | 1 | 0.01 |
| Dicaeum trochileum | foliage frugivore | 22 | 0.10 | 4 | 0.09 | 23 | 0.12 |
| Passer montanus | terrestrial granivore | 54 | 0.24 | 14 | 0.32 | 62 | 0.31 |
| Pycnonotus aurigaster | foliage frugivore/insectivore | 53 | 0.23 | 5 | 0.11 | 40 | 0.20 |
| Spilopelia chinensis | terrestrial granivore | 4 | 0.02 | 3 | 0.07 | 0.00 | |
| Environmental Predictor | Urban Adapter | Urban Exploiter | Mean (Min-Max) |
|---|---|---|---|
| Distance to perennial water (meter) | 3.8 | 43.30 | 193(0–589) |
| Distance to large green patches (meter) | 18.50 | 24.10 | 36(0–182) |
| Land Function (Categorical) | 28.70 | 18.00 | Categorical data, settlement and housing as dominant zone |
| Landscape greenness (Unitless) | 4.30 | 6.40 | 0.5(−0.42–0.79) |
| Elevation (masl) | 5.60 | 6.50 | 49(45–93) |
| Temperature (K) | 5.10 | 1.70 | 306(303–313) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Winarni, N.L.; Fuad, H.A.H.; Anugra, B.G.; Kaunain, N.N.; Anisafitri, S.; Atria, M.; Putrika, A. Potential Ecological Distributions of Urban Adapters and Urban Exploiters for the Sustainability of the Urban Bird Network. ISPRS Int. J. Geo-Inf. 2022, 11, 474. https://doi.org/10.3390/ijgi11090474
Winarni NL, Fuad HAH, Anugra BG, Kaunain NN, Anisafitri S, Atria M, Putrika A. Potential Ecological Distributions of Urban Adapters and Urban Exploiters for the Sustainability of the Urban Bird Network. ISPRS International Journal of Geo-Information. 2022; 11(9):474. https://doi.org/10.3390/ijgi11090474
Chicago/Turabian StyleWinarni, Nurul L., Habiburrachman A. H. Fuad, Bhisma G. Anugra, Nabilla Nuril Kaunain, Shania Anisafitri, Mega Atria, and Afiatry Putrika. 2022. "Potential Ecological Distributions of Urban Adapters and Urban Exploiters for the Sustainability of the Urban Bird Network" ISPRS International Journal of Geo-Information 11, no. 9: 474. https://doi.org/10.3390/ijgi11090474
APA StyleWinarni, N. L., Fuad, H. A. H., Anugra, B. G., Kaunain, N. N., Anisafitri, S., Atria, M., & Putrika, A. (2022). Potential Ecological Distributions of Urban Adapters and Urban Exploiters for the Sustainability of the Urban Bird Network. ISPRS International Journal of Geo-Information, 11(9), 474. https://doi.org/10.3390/ijgi11090474