

Based on Sportomics: Comparison of Physiological Status of Collegiate Sprinters in Different Pre-Competition Preparation Periods



Abstract
1. Introduction
2. Materials and Methods
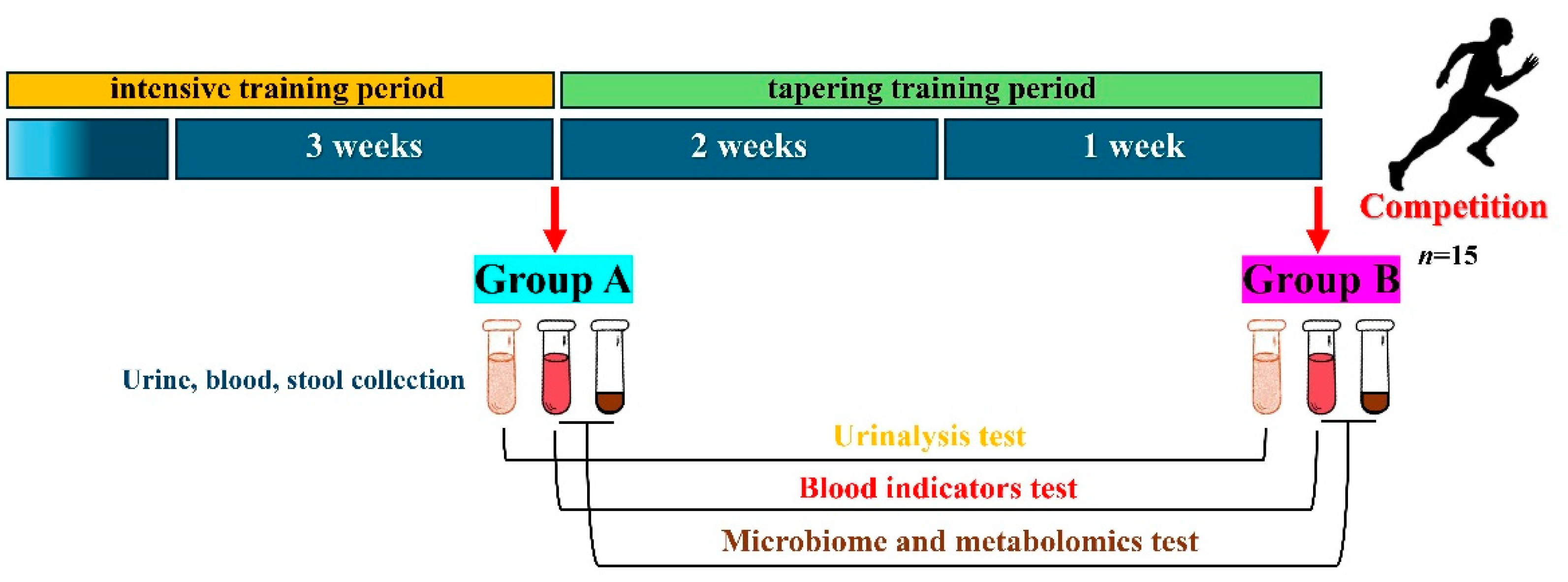
2.1. Participants and Groups
2.2. Body Composition
2.3. Dietary Survey
2.4. Energy Expenditure and Sleep Efficiency
2.5. The Test of Heart Rate and Respiratory Rate
2.6. Blood and Urine Indicators
2.7. Gut Microbiome Sequencing and Analysis
2.8. Metabolomics Sequencing and Analysis
2.9. Statistical Analysis
3. Results
3.1. Sports Performance and Body Composition
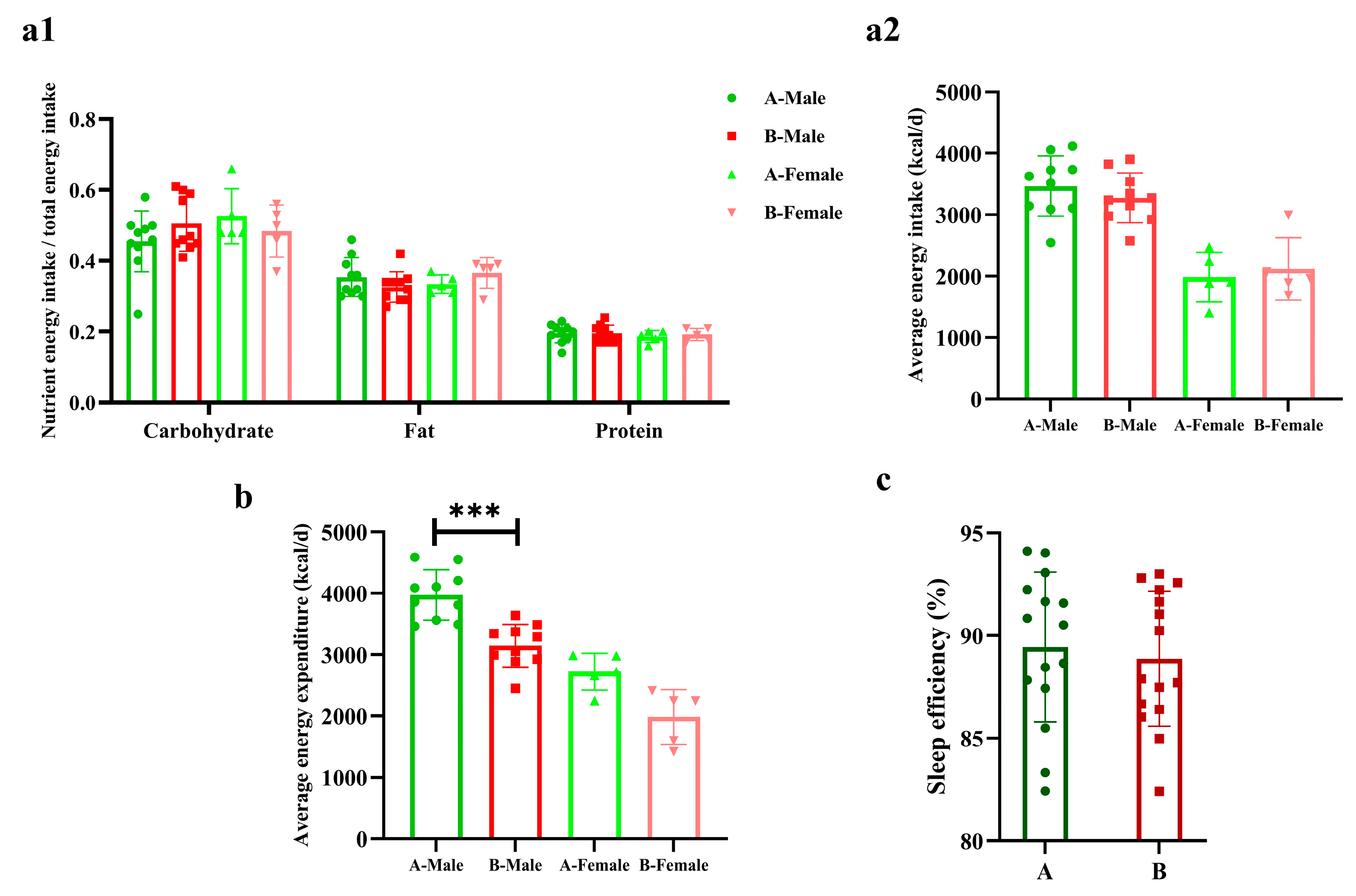
3.2. Energy Metabolism and Sleep
3.3. Heart Rate and Respiratory Rate during Training
3.4. Blood Markers
3.5. Urine Markers
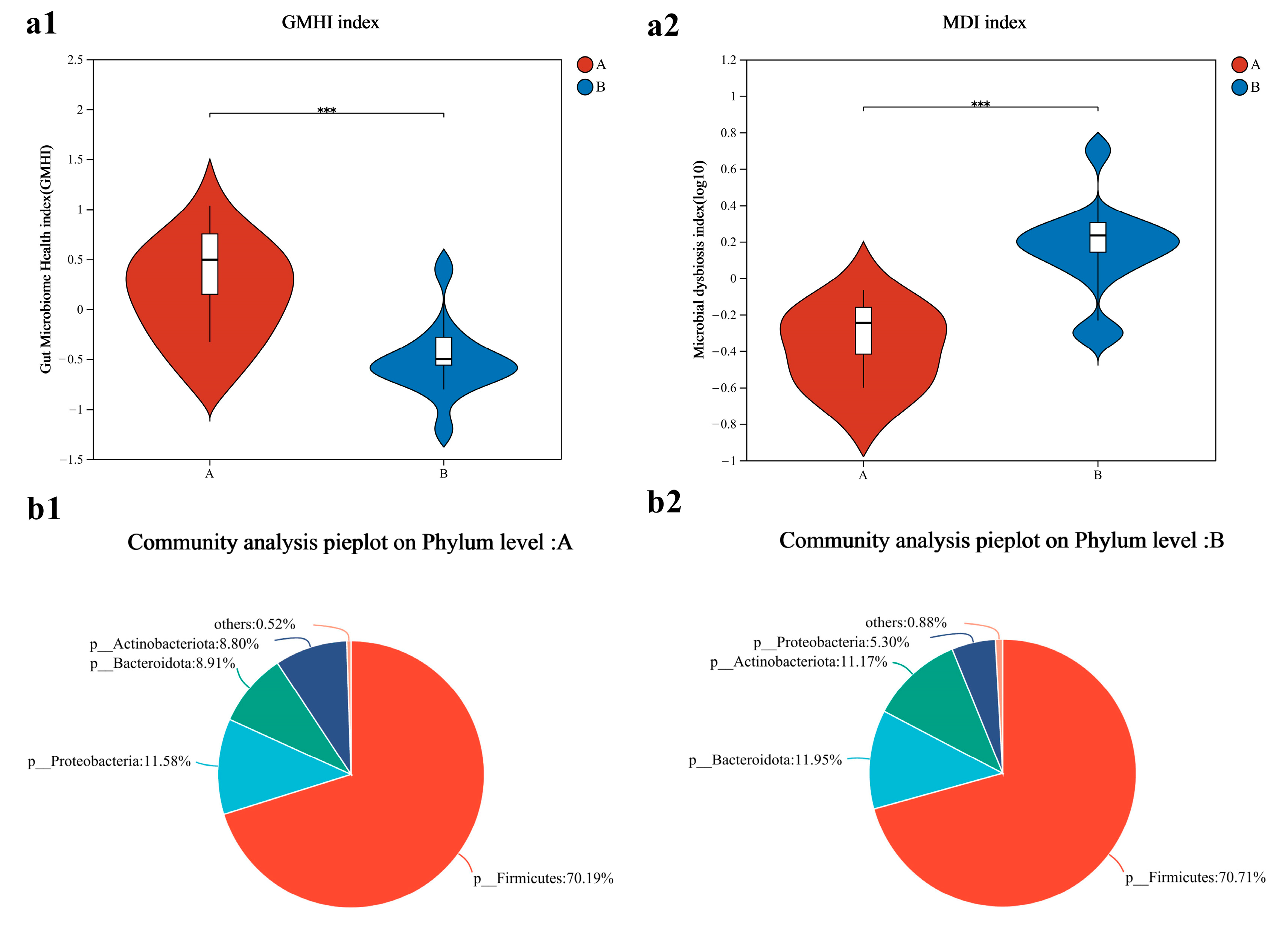
3.6. Gut Microbiome Characterization Index and Composition Analysis
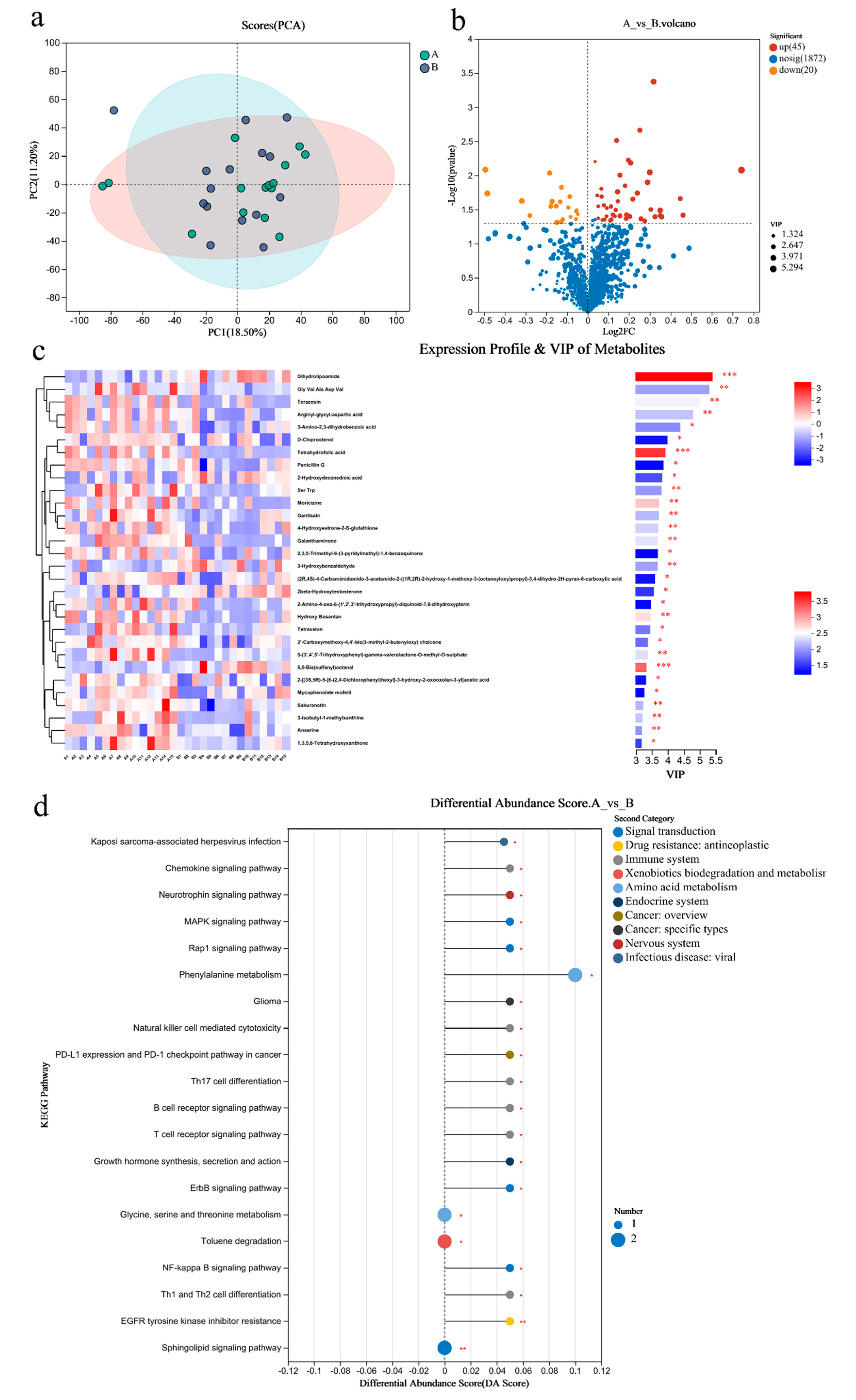
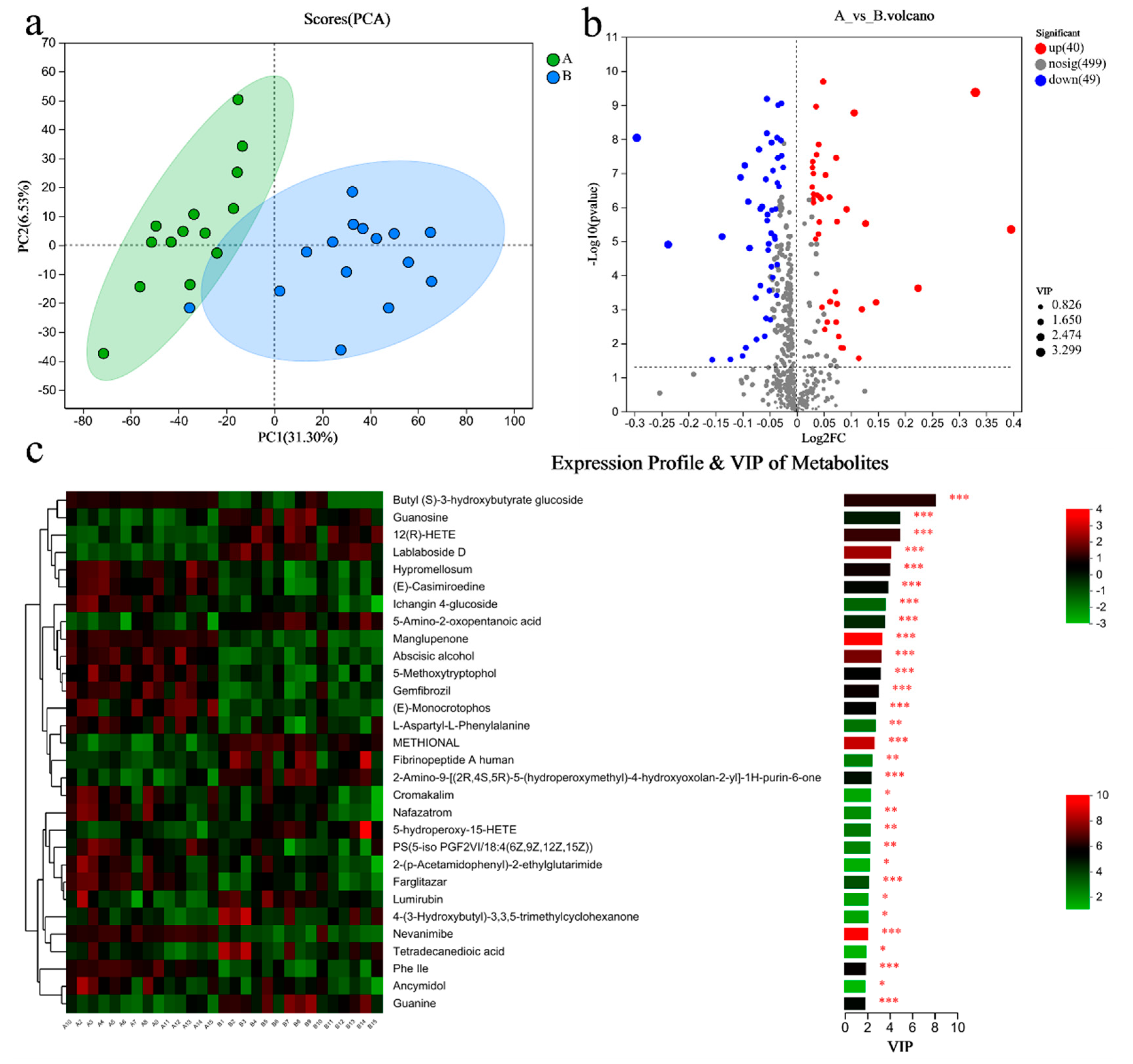
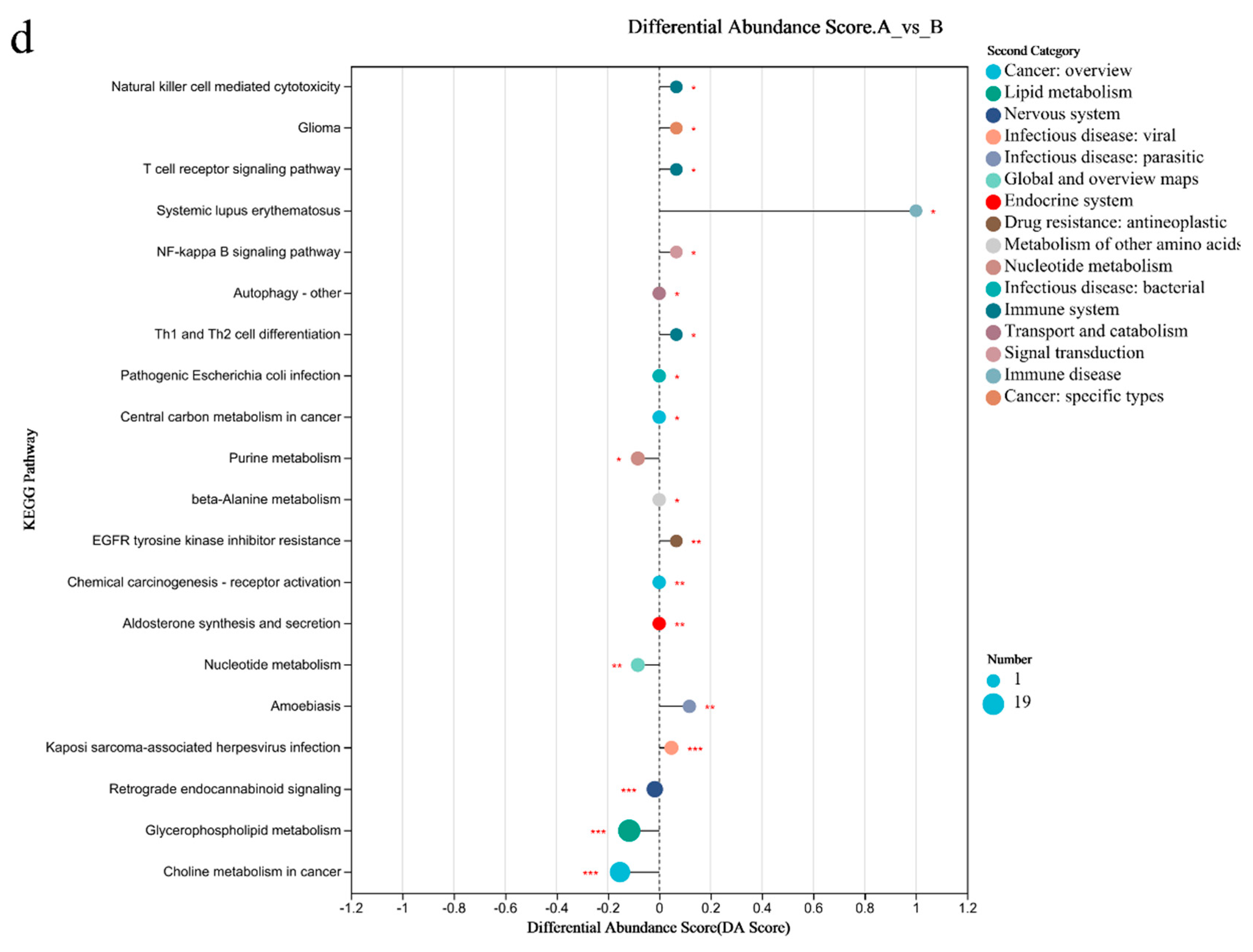
3.7. Metabolomic Analysis of the Gut Microbiome
3.8. Metabolomic Analysis of Blood
4. Discussions
4.1. Diet, Training Load Adaptation, and Sports Performance
4.2. Gut Microbiome and Sports Performance
4.3. Metabolites and Sports Performance
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Malsagova, K.A.; Kopylov, A.T.; Stepanov, A.A.; Kulikova, L.I.; Izotov, A.A.; Yurku, K.A.; Balakin, E.I.; Pustovoyt, V.I.; Kaysheva, A.L. Metabolomic and Proteomic Profiling of Athletes Performing Physical Activity under Hypoxic Conditions. Sports 2024, 12, 72. [Google Scholar] [CrossRef] [PubMed]
- Hamlin, M.J.; Wilkes, D.; Elliot, C.A.; Lizamore, C.A.; Kathiravel, Y. Monitoring Training Loads and Perceived Stress in Young Elite University Athletes. Front. Physiol. 2019, 10, 34. [Google Scholar] [CrossRef] [PubMed]
- Lee, E.C.; Fragala, M.S.; Kavouras, S.A.; Queen, R.M.; Pryor, J.L.; Casa, D.J. Biomarkers in Sports and Exercise: Tracking Health, Performance, and Recovery in Athletes. J. Strength Cond. Res. 2017, 31, 2920–2937. [Google Scholar] [CrossRef] [PubMed]
- Khoramipour, K.; Sandbakk, O.; Hassanzadeh Keshteli, A.; Gaeini, A.; Wishart, D.; Chamari, K. Metabolomics in Exercise and Sports: A Systematic Review. Sports Med. 2022, 52, 547–583. [Google Scholar] [CrossRef] [PubMed]
- Bassini, A.; Cameron, L.C. Sportomics: Building a new concept in metabolic studies and exercise science. Biochem. Biophys. Res. Commun. 2014, 445, 708–716. [Google Scholar] [CrossRef]
- Bongiovanni, T.; Lacome, M.; Fanos, V.; Martera, G.; Cione, E.; Cannataro, R. Metabolomics in Team-Sport Athletes: Current Knowledge, Challenges, and Future Perspectives. Proteomes 2022, 10, 27. [Google Scholar] [CrossRef]
- Pintus, R.; Bongiovanni, T.; Corbu, S.; Francavilla, V.C.; DessÌ, A.; Noto, A.; Corsello, G.; Finco, G.; Fanos, V.; Marincola, F.C. Sportomics in professional soccer players: Metabolomics results during preseason. J. Sports Med. Phys. Fit. 2020, 61, 324–330. [Google Scholar] [CrossRef]
- Hudson, J.F.; Phelan, M.M.; Owens, D.J.; Morton, J.P.; Close, G.L.; Stewart, C.E.J.M. “Fuel for the damage induced”: Untargeted metabolomics in elite rugby union match play. Metabolites 2021, 11, 544. [Google Scholar] [CrossRef]
- Hughes, R.L.; Holscher, H.D. Fueling Gut Microbes: A Review of the Interaction between Diet, Exercise, and the Gut Microbiota in Athletes. Adv. Nutr. 2021, 12, 2190–2215. [Google Scholar] [CrossRef]
- Li, Y.; Cheng, M.; Zha, Y.; Yang, K.; Tong, Y.; Wang, S.; Lu, Q.; Ning, K. Gut microbiota and inflammation patterns for specialized athletes: A multi-cohort study across different types of sports. mSystems 2023, 8, e0025923. [Google Scholar] [CrossRef]
- Ludbrook, C.; Clark, D. Energy expenditure and nutrient intake in long-distance runners. Nutr. Res. 1992, 12, 689–699. [Google Scholar] [CrossRef]
- Donati Zeppa, S.; Agostini, D.; Gervasi, M.; Annibalini, G.; Amatori, S.; Ferrini, F.; Sisti, D.; Piccoli, G.; Barbieri, E.; Sestili, P.; et al. Mutual Interactions among Exercise, Sport Supplements and Microbiota. Nutrients 2019, 12, 17. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Guisado, J. Athletic performance: Muscle glycogen and protein intake. Apunt. Sports Med. 2008, 43, 142–151. [Google Scholar]
- Furber, M.J.W.; Young, G.R.; Holt, G.S.; Pyle, S.; Davison, G.; Roberts, M.G.; Roberts, J.D.; Howatson, G.; Smith, D.L. Gut Microbial Stability is Associated with Greater Endurance Performance in Athletes Undertaking Dietary Periodization. mSystems 2022, 7, e0012922. [Google Scholar] [CrossRef] [PubMed]
- Naderi, A.; Gobbi, N.; Ali, A.; Berjisian, E.; Hamidvand, A.; Forbes, S.C.; Koozehchian, M.S.; Karayigit, R.; Saunders, B. Carbohydrates and Endurance Exercise: A Narrative Review of a Food First Approach. Nutrients 2023, 15, 1367. [Google Scholar] [CrossRef]
- Aragon, A.A.; Schoenfeld, B.J.; Wildman, R.; Kleiner, S.; VanDusseldorp, T.; Taylor, L.; Earnest, C.P.; Arciero, P.J.; Wilborn, C.; Kalman, D.S.; et al. International society of sports nutrition position stand: Diets and body composition. J. Int. Soc. Sports Nutr. 2017, 14, 16. [Google Scholar] [CrossRef]
- Thomas, D.T.; Erdman, K.A.; Burke, L.M. Nutrition and athletic performance. Med. Sci. Sports Exerc. 2016, 48, 543–568. [Google Scholar]
- Osterberg, K.L.; Horswill, C.A.; Baker, L.B. Pregame urine specific gravity and fluid intake by National Basketball Association players during competition. J. Athl. Train. 2009, 44, 53–57. [Google Scholar] [CrossRef]
- Wegierska, A.E.; Charitos, I.A.; Topi, S.; Potenza, M.A.; Montagnani, M.; Santacroce, L. The Connection Between Physical Exercise and Gut Microbiota: Implications for Competitive Sports Athletes. Sports Med. 2022, 52, 2355–2369. [Google Scholar] [CrossRef]
- Matijašić, M.; Meštrović, T.; Paljetak, H.; Perić, M.; Barešić, A.; Verbanac, D. Gut Microbiota beyond Bacteria-Mycobiome, Virome, Archaeome, and Eukaryotic Parasites in IBD. Int. J. Mol. Sci. 2020, 21, 2668. [Google Scholar] [CrossRef]
- Yamamura, R.; Nakamura, K.; Kitada, N.; Aizawa, T.; Shimizu, Y.; Nakamura, K.; Ayabe, T.; Kimura, T.; Tamakoshi, A. Associations of gut microbiota, dietary intake, and serum short-chain fatty acids with fecal short-chain fatty acids. Biosci. Microbiota Food Health 2020, 39, 11–17. [Google Scholar] [CrossRef] [PubMed]
- Durk, R.P.; Castillo, E.; Márquez-Magaña, L.; Grosicki, G.J.; Bolter, N.D.; Lee, C.M.; Bagley, J.R. Gut Microbiota Composition Is Related to Cardiorespiratory Fitness in Healthy Young Adults. Int. J. Sport Nutr. Exerc. Metab. 2019, 29, 249–253. [Google Scholar] [CrossRef] [PubMed]
- Han, M.; Yang, K.; Yang, P.; Zhong, C.; Chen, C.; Wang, S.; Lu, Q.; Ning, K. Stratification of athletes’ gut microbiota: The multifaceted hubs associated with dietary factors, physical characteristics and performance. Gut Microbes 2020, 12, 1842991. [Google Scholar] [CrossRef] [PubMed]
- Kakanis, M.W.; Peake, J.; Brenu, E.W.; Simmonds, M.; Gray, B.; Hooper, S.L.; Marshall-Gradisnik, S.M. The open window of susceptibility to infection after acute exercise in healthy young male elite athletes. Exerc. Immunol. Rev. 2010, 16, 119–137. [Google Scholar] [CrossRef] [PubMed]
- Baird, M.F.; Graham, S.M.; Baker, J.S.; Bickerstaff, G.F. Creatine-kinase- and exercise-related muscle damage implications for muscle performance and recovery. J. Nutr. Metab. 2012, 2012, 960363. [Google Scholar] [CrossRef]
- Ticinesi, A.; Lauretani, F.; Tana, C.; Nouvenne, A.; Ridolo, E.; Meschi, T. Exercise and immune system as modulators of intestinal microbiome: Implications for the gut-muscle axis hypothesis. Exerc. Immunol. Rev. 2019, 25, 84–95. [Google Scholar]
- San-Millán, I.; Stefanoni, D.; Martinez, J.L.; Hansen, K.C.; D’Alessandro, A.; Nemkov, T. Metabolomics of Endurance Capacity in World Tour Professional Cyclists. Front. Physiol. 2020, 11, 578. [Google Scholar] [CrossRef]
- Rowland, I.; Gibson, G.; Heinken, A.; Scott, K.; Swann, J.; Thiele, I.; Tuohy, K. Gut microbiota functions: Metabolism of nutrients and other food components. Eur. J. Nutr. 2018, 57, 1–24. [Google Scholar] [CrossRef]
- Dupuy, O.; Wafa, D.; Theurot, D.; Bosquet, L.; Dugue, B. An Evidence-Based Approach for Choosing Post-exercise Recovery Techniques to Reduce Markers of Muscle Damage, Soreness, Fatigue, and Inflammation: A Systematic Review with Meta-Analysis. Front. Physiol. 2018, 9, 403. [Google Scholar] [CrossRef]
- Gallego-Selles, A.; Galvan-Alvarez, V.; Martinez-Canton, M.; Garcia-Gonzalez, E.; Morales-Alamo, D.; Santana, A.; Gonzalez-Henriquez, J.J.; Dorado, C.; Calbet, J.A.L.; Martin-Rincon, M. Fast regulation of the NF-κB signalling pathway in human skeletal muscle revealed by high-intensity exercise and ischaemia at exhaustion: Role of oxygenation and metabolite accumulation. Redox Biol. 2022, 55, 102398. [Google Scholar] [CrossRef]
- Alkhatib, A.; Feng, W.H.; Huang, Y.J.; Kuo, C.H.; Hou, C.W. Anserine Reverses Exercise-Induced Oxidative Stress and Preserves Cellular Homeostasis in Healthy Men. Nutrients 2020, 12, 1146. [Google Scholar] [CrossRef] [PubMed]
- Kostrzewa-Nowak, D.; Nowak, R. Differential Th Cell-Related Immune Responses in Young Physically Active Men after an Endurance Effort. J. Clin. Med. 2020, 9, 1795. [Google Scholar] [CrossRef] [PubMed]
- Dudzinska, W.; Lubkowska, A.; Dolegowska, B.; Safranow, K.; Jakubowska, K. Adenine, guanine and pyridine nucleotides in blood during physical exercise and restitution in healthy subjects. Eur. J. Appl. Physiol. 2010, 110, 1155–1162. [Google Scholar] [CrossRef] [PubMed]
- Vike, N.L.; Bari, S.; Stetsiv, K.; Talavage, T.M.; Nauman, E.A.; Papa, L.; Slobounov, S.; Breiter, H.C.; Cornelis, M.C. Metabolomic response to collegiate football participation: Pre- and Post-season analysis. Sci. Rep. 2022, 12, 3091. [Google Scholar] [CrossRef]
- Wang, C.-C.; Shi, H.-H.; Zhang, L.-Y.; Ding, L.; Xue, C.-H.; Yanagita, T.; Zhang, T.-T.; Wang, Y.-M. The rapid effects of eicosapentaenoic acid (EPA) enriched phospholipids on alleviating exercise fatigue in mice. RSC Adv. 2019, 9, 33863–33871. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Times | Mon. | Tues. | Wed. | Thur. | Fri | Sat. |
|---|---|---|---|---|---|---|
| Strengthening Training Period (Group A) | ||||||
| Warm-up exercise | Warm-up and run for 10 min, cool-down for 10 min | |||||
| Formal training | 100 m run × 1 set + 60 m run × 2 sets + 30 m run × 2 sets | 300 m run × 5 sets | Upper and lower limb and back strength training | 200 m run × 4 sets + 150 m run × 4 sets | Hurdle jumping, hurdle running, hip and ankle mobility training | Full speed special athletics training |
| Cooling-down exercise | Stretching for 15 min | |||||
| Tapering training period (Group B) | ||||||
| Warm-up exercise | Warm-up and run for 10 min, cool-down for 10 min | |||||
| Formal training | 100 m run × 1 set + 60 m run × 1 set + 30 m run × 1 set | 300 m run × 2 sets | Upper and lower limb and back strength training | - | - | - |
| Cooling-down exercise | Stretching for 15 min | |||||
| Indicators (Unit) | Male (n = 10) | Female (n = 5) | ||
|---|---|---|---|---|
| Group A | Group B | Group A | Group B | |
| Height (cm) | 178.75 ± 3.19 | 169.10 ± 5.32 | ||
| Body weight (kg) | 68.52 ± 5.25 | 68.24 ± 5.59 | 56.88 ± 3.49 | 56.25±3.32 |
| Body mass index | 21.37 ± 1.18 | 21.31 ± 1.25 | 19.92 ± 0.57 | 19.68 ± 0.54 |
| Body fat percentage (%) | 10.29 ± 3.64 | 11.51 ± 2.08 | 18.60 ± 2.17 | 17.22 ± 4.32 |
| Skeletal muscle mass (kg) | 32.81 ± 2.61 | 31.50 ± 2.22 | 24.18 ± 1.58 | 24.32 ± 1.11 |
| Trunk skeletal muscle mass (kg) | 6.68 ± 0.79 | 6.96 ± 1.05 | 6.40 ± 0.48 | 6.90 ± 0.78 |
| Limb skeletal muscle mass (kg) | 26.10 ± 2.10 | 25.04 ± 1.80 ** | 17.80 ± 1.11 | 17.42 ± 0.67 |
| Indicators (Times/min) | Group A | Group B | ||
|---|---|---|---|---|
| Strength training | Heart rate | Maximum | 182 ± 29 | 164 ± 14 * |
| Average | 117 ± 17 | 113 ± 13 | ||
| Mode | 115 ± 17 | 110 ± 13 | ||
| Respiratory rate | Maximum | 423 ± 11 | 37 ± 6 | |
| Average | 23 ± 3 | 21 ± 2 | ||
| Mode | 24 ± 5 | 20 ± 3 ** | ||
| Specific athletics training | Heart rate | Maximum | 185 ± 15 | 188 ± 20 |
| Average | 120 ± 9 | 128 ± 9 | ||
| Mode | 110 ± 19 | 124 ± 12 * | ||
| Respiratory rate | Maximum | 43 ± 5 | 46 ± 5 | |
| Average | 23 ± 7 | 24 ± 3 | ||
| Mode | 24 ± 5 | 20 ± 3 ** | ||
| Blood Indicators (Unit) (Reference Values) | Implication | Group A | Group B | ||
|---|---|---|---|---|---|
| Male (n = 10) | Female (n = 5) | Male (n = 10) | Female (n = 5) | ||
| WBC (109/ L) (3.69~9.16) | Immune status | 5.24 ± 0.89 | 5.35 ± 1.04 | ||
| Lymphocyte ratio (%) (24~48.4) | 46.43 ± 6.85 | 39.20 ± 7.63 * | |||
| IgA (g/L) (0.72~4.29) | 2.24 ± 0.70 | 2.26 ± 0.73 | |||
| RBC (1012/L) (M: 4.3~5.8; F: 3.8~5.1) | Aerobic capacity | 5.11 ± 0.39 | 4.52 ± 0.26 | 5.08 ± 0.39 | 4.64 ± 0.25 |
| Ferritin (ng/mL) (M: 21.81~274.66; F: 4.63~204.00) | Aerobic capacity Nutritional status | 101.02 ± 29.04 | 25.33 ± 12.68 | 123.30 ± 22.43 | 33.27 ± 16.59 |
| HGB (g/L) (M: 130~175; F: 115~150) | Aerobic capacity Training loads | 153.30 ± 8.29 | 129.20 ± 10.11 | 152.50 ± 8.72 | 132.60 ± 9.96 |
| MCHC (g/L) (310~370) | 350.10 ± 8.79 | 336.40 ± 12.10 | 355.00 ± 9.10 | 339.40 ± 10.78 | |
| Testosterone (ng/mL) (M: 2.49~8.36; F: 0.029~0.481) | Training loads Recovery ability | 7.39 ± 1.32 | 0.48 ± 0.23 | 6.91 ± 1.56 | 0.52 ± 0.15 |
| Cortisol (μg/dL) (4.26~24.85) | 19.27 ± 4.09 | 17.66 ± 4.49 | 17.43 ± 3.35 | 17.47 ± 2.79 | |
| T/C | 0.40 ± 0.09 | 0.03 ± 0.02 | 0.40 ± 0.10 | 0.03 ± 0.01 | |
| CK (U/L) (M: 38~174; F: 26~140) | Training intensity Muscle injury and recovery | 191.76 ± 68.26 | 217.19 ± 53.62 | 286.65 ± 63.59 * | 370.32 ± 59.07 ** |
| BUN (mmol/L) (2.9~8.2) | Training volume | 5.45 ± 1.08 | 5.11 ± 0.94 | ||
| LDH (U/L) (109~245) | Anaerobic capacity | 163.23 ± 19.29 | 167.45 ± 25.49 | ||
| IL-6 (pg/mL) (≤7) | Inflammatory response | 9.29 ± 5.29 | 12.87 ± 9.28 | ||
| IL-1β (pg/mL) (≤5) | 17.28 ± 9.24 | 20.62 ± 13.20 | |||
| Urine Indicators | Group A | Group B | |
|---|---|---|---|
| WBC | The detection rates | 13.33% | 35.71% |
| GLU | (-) | (-) | |
| BLD | (-) | (-) | |
| PRO | 6.67% | 35.71% | |
| NIT | (-) | (-) | |
| UBG | (-) | (-) | |
| BIL | (-) | (-) | |
| KET | (-) | (-) | |
| pH (3~7.5) | The test value (Reference values) | 5.63 ± 0.74 | 5.62 ± 0.74 |
| SG (1.010~1.025) | 1.028 ± 0.003 | 1.027 ± 0.003 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fu, P.; Duan, X.; Zhang, Y.; Dou, X.; Gong, L. Based on Sportomics: Comparison of Physiological Status of Collegiate Sprinters in Different Pre-Competition Preparation Periods. Metabolites 2024, 14, 527. https://doi.org/10.3390/metabo14100527
Fu P, Duan X, Zhang Y, Dou X, Gong L. Based on Sportomics: Comparison of Physiological Status of Collegiate Sprinters in Different Pre-Competition Preparation Periods. Metabolites. 2024; 14(10):527. https://doi.org/10.3390/metabo14100527
Chicago/Turabian StyleFu, Pengyu, Xiaomin Duan, Yuting Zhang, Xiangya Dou, and Lijing Gong. 2024. "Based on Sportomics: Comparison of Physiological Status of Collegiate Sprinters in Different Pre-Competition Preparation Periods" Metabolites 14, no. 10: 527. https://doi.org/10.3390/metabo14100527
APA StyleFu, P., Duan, X., Zhang, Y., Dou, X., & Gong, L. (2024). Based on Sportomics: Comparison of Physiological Status of Collegiate Sprinters in Different Pre-Competition Preparation Periods. Metabolites, 14(10), 527. https://doi.org/10.3390/metabo14100527