
Evidence of 11-Hydroxy-hexahydrocannabinol and 11-Nor-9-carboxy-hexahydrocannabinol as Novel Human Metabolites of Δ9-Tetrahydrocannabinol


Abstract
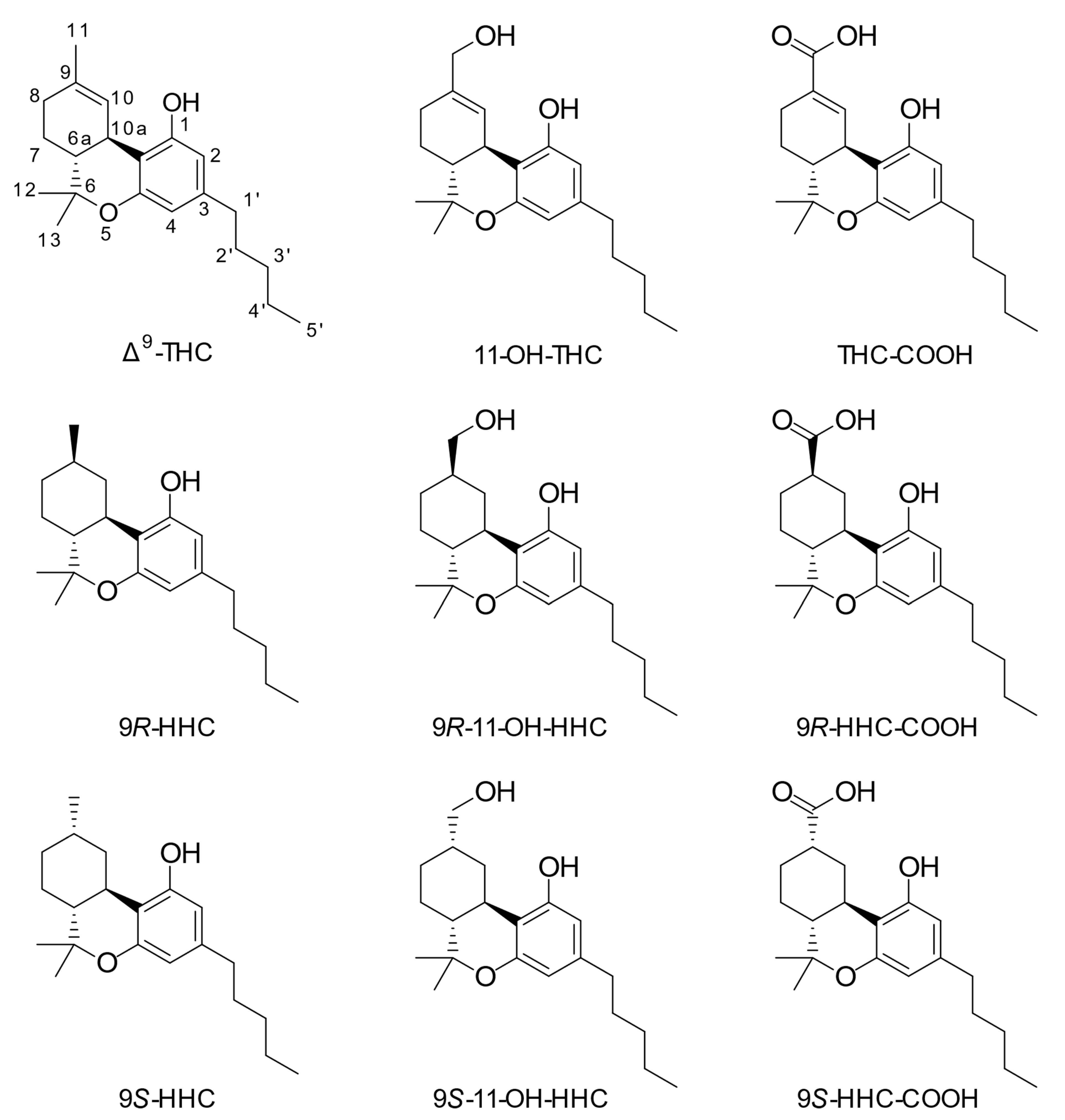
1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Sample Preparation
2.3. Liquid Chromatography–Mass Spectrometry Analysis
2.4. Incubation in Human Liver Microsomes
2.5. Forensic Traffic Cases
2.6. Data Analysis
3. Results
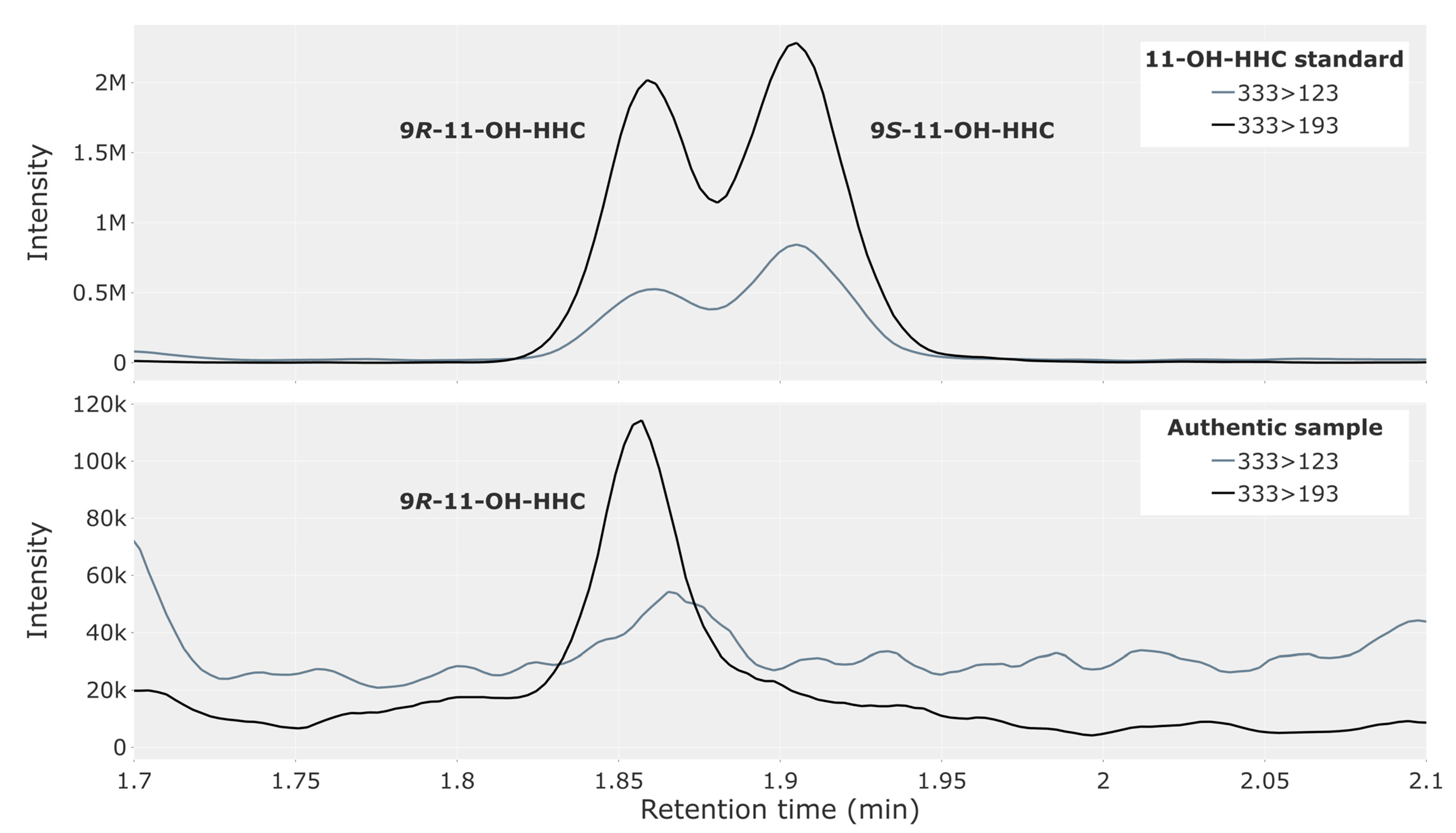
3.1. Traffic Cases
3.2. Incubation with Human Liver Microsomes
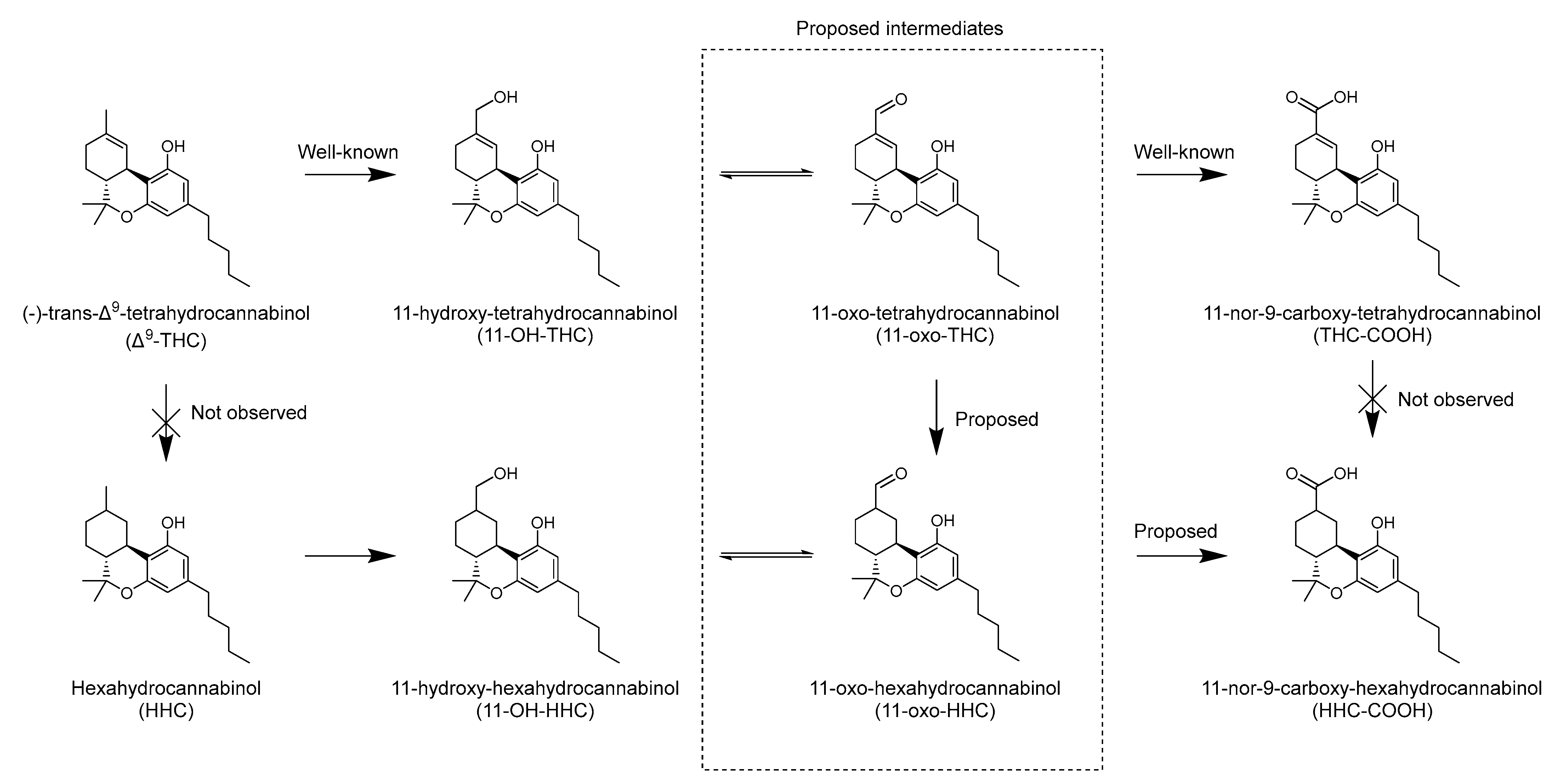
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Huestis, M.A. Human Cannabinoid Pharmacokinetics. Chem. Biodivers. 2007, 4, 1770–1804. [Google Scholar] [CrossRef] [PubMed]
- Agurell, S.; Halldin, M.; Lindgren, J.E.; Ohlsson, A.; Widman, M.; Gillespie, H.; Hollister, L. Pharmacokinetics and Metabolism of Δ1-Tetrahydrocannabinol and Other Cannabinoids with Emphasis on Man. Pharmacol. Rev. 1986, 38, 21–43. [Google Scholar]
- Bornheim, L.M.; Lasker, J.M.; Raucy, J.L. Human Hepatic Microsomal Metabolism of Delta 1-Tetrahydrocannabinol. Drug Metab. Dispos. 1992, 20, 241–246. [Google Scholar] [PubMed]
- Halldin, M.M.; Andersson, L.K.; Widman, M.; Hollister, L.E. Further Urinary Metabolites of Delta 1-Tetrahydrocannabinol in Man. Arzneimittelforschung 1982, 32, 1135–1138. [Google Scholar] [PubMed]
- Halldin, M.M.; Carlsson, S.; Kanter, S.L.; Widman, M.; Agurell, S. Urinary Metabolites of Delta 1-Tetrahydrocannabinol in Man. Arzneimittelforschung 1982, 32, 764–768. [Google Scholar] [PubMed]
- Halldin, M.M.; Widman, M.; Bahr, C.V.; Lindgren, J.E.; Martin, B.R. Identification of in Vitro Metabolites of Delta 1-Tetrahydrocannabinol Formed by Human Livers. Drug Metab. Dispos. 1982, 10, 297–301. [Google Scholar]
- Watanabe, K.; Yamaori, S.; Funahashi, T.; Kimura, T.; Yamamoto, I. Cytochrome P450 Enzymes Involved in the Metabolism of Tetrahydrocannabinols and Cannabinol by Human Hepatic Microsomes. Life Sci. 2007, 80, 1415–1419. [Google Scholar] [CrossRef]
- Williams, P.L.; Moffat, A.C. Identification in Human Urine of Delta 9-Tetrahydrocannabinol-11-Oic Acid Glucuronide: A Tetrahydrocannabinol Metabolite. J. Pharm. Pharmacol. 1980, 32, 445–448. [Google Scholar] [CrossRef]
- European Monitoring Centre for Drugs and Drug Addiction (EMCDDA). Technical Report: Hexahydrocannabinol (HHC) and Related Substances; European Monitoring Centre for Drugs and Drug Addiction (EMCDDA): Lisbon, Portugal, 2023. [Google Scholar]
- Harvey, D.J.; Brown, N.K. Comparative in Vitro Metabolism of the Cannabinoids. Pharmacol. Biochem. Behav. 1991, 40, 533–540. [Google Scholar] [CrossRef]
- Schirmer, W.; Auwärter, V.; Kaudewitz, J.; Schürch, S.; Weinmann, W. Identification of Human Hexahydrocannabinol Metabolites in Urine. Eur. J. Mass Spectrom. 2023, 14690667231200139. [Google Scholar] [CrossRef]
- Manier, S.K.; Angert Valdiviezo, J.; Vollmer, A.C.; Eckstein, N.; Meyer, M.R. Analytical Toxicology of the Semi-Synthetic Cannabinoid Hexahydrocannabinol Studied in Human Samples, Pooled Human Liver S9 Fraction, Rat Samples and Drug Products Using HPLC-HRMS-MS. J. Anal. Toxicol. 2023, bkad079. [Google Scholar] [CrossRef] [PubMed]
- Harvey, D.J.; Martin, B.R.; Paton, W.D. Identification of Metabolites of Delta1- and Delta1(6)-Tetrahydrocannabinol Containing a Reduced Double Bond. J. Pharm. Pharmacol. 1977, 29, 495–497. [Google Scholar] [CrossRef] [PubMed]
- Brown, N.K.; Harvey, D.J. In Vivo Metabolism of the Methyl Homologues of Delta-8-Tetrahydrocannabinol, Delta-9-Tetrahydrocannabinol and Abn-Delta-8-Tetrahydrocannabinol in the Mouse. Biomed. Env. Mass Spectrom. 1988, 15, 389–398. [Google Scholar] [CrossRef]
- Brown, N.K.; Harvey, D.J. In Vivo Metabolism of the Ethyl Homologues of Delta-8-Tetrahydrocannabinol and Delta-9-Tetrahydrocannabinol in the Mouse. Biol. Mass Spectrom. 1991, 20, 324–328. [Google Scholar] [CrossRef] [PubMed]
- Andersen, D.; Rasmussen, B.; Linnet, K. Validation of a Fully Automated Robotic Setup for Preparation of Whole Blood Samples for LC-MS Toxicology Analysis. J. Anal. Toxicol. 2012, 36, 280–287. [Google Scholar] [CrossRef][Green Version]
- Team, T. The pandas development team Pandas-Dev/Pandas: Pandas. Zenodo 2022, 21, 1–9. [Google Scholar]
- Plotly Technologies Inc. Collaborative Data Science. Available online: https://plot.ly (accessed on 22 November 2023).
- Harris, C.R.; Millman, K.J.; van der Walt, S.J.; Gommers, R.; Virtanen, P.; Cournapeau, D.; Wieser, E.; Taylor, J.; Berg, S.; Smith, N.J.; et al. Array Programming with NumPy. Nature 2020, 585, 357–362. [Google Scholar] [CrossRef]
- Virtanen, P.; Gommers, R.; Oliphant, T.E.; Haberland, M.; Reddy, T.; Cournapeau, D.; Burovski, E.; Peterson, P.; Weckesser, W.; Bright, J.; et al. SciPy 1.0: Fundamental Algorithms for Scientific Computing in Python. Nat. Methods 2020, 17, 261–272. [Google Scholar] [CrossRef]
- Huang, M.; Hu, H.; Ma, L.; Zhou, Q.; Yu, L.; Zeng, S. Carbon-Carbon Double-Bond Reductases in Nature. Drug Metab. Rev. 2014, 46, 362–378. [Google Scholar] [CrossRef]
- Penning, T.M. The Aldo-Keto Reductases (AKRs): Overview. Chem. Biol. Interact. 2015, 234, 236–246. [Google Scholar] [CrossRef]
- Ben-Zvi, Z.; Burstein, S. 7-Oxo-Delta1-Tetrahydrocannabinol: A Novel Metabolite of Delta1-Tetrahydrocannabinol. Res. Commun. Chem. Pathol. Pharmacol. 1974, 8, 223–229. [Google Scholar] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Monitored m/z Transitions | Cone Voltage (V) | Collision Energy (eV) | Retention Time (min) |
|---|---|---|---|---|
| Δ9-tetrahydrocannabinol | 315.22 > 193.12 * 315.22 > 123.04 | 42 42 | 22 32 | 3.60 |
| 9R-Hexahydrocannabinol | 317.25 > 193.12 * 317.25 > 123.04 | 20 20 | 20 40 | 4.08 |
| 9S-Hexahydrocannabinol | 317.25 > 193.12 * 317.25 > 123.04 | 20 20 | 20 40 | 4.00 |
| 9R-11-hydroxy-hexahydrocannabinol | 333.24 > 193.12 * 333.24 > 123.04 | 20 20 | 25 35 | 1.86 |
| 9S-11-hydroxy-hexahydrocannabinol | 333.24 > 193.12 * 333.24 > 123.04 | 20 20 | 25 35 | 1.91 |
| 9R-11-nor-carboxy-hexahydrocannabinol | 347.22 > 193.12 * 347.22 > 123.04 | 20 20 | 25 35 | 1.80 |
| 9S-11-nor-carboxy-hexahydrocannabinol | 347.22 > 193.12 * 347.22 > 123.04 | 20 20 | 25 35 | 1.97 |
| 11-hydroxy-tetrahydrocannabinol | 331.22 > 193.12 * 331.22 > 201.20 | 20 20 | 29 25 | 1.71 |
| 11-nor-9-carboxy-tetrahydrocannabinol | 345.20 > 193.12 * 345.20 > 299.20 | 42 42 | 26 20 | 1.90 |
| Δ9-tetrahydrocannabinol-d3 | 318.22 > 196.12 | 42 | 22 | 3.58 |
| 11-hydroxy-tetrahydrocannabinol-d3 | 334.27 > 196.23 | 20 | 29 | 1.89 |
| 11-nor-9-carboxy-tetrahydrocannabinol-d3 | 348.20 > 196.12 | 34 | 20 | 1.70 |
| Group | Δ9-THC | HHC | Number of Cases (n = 308) | 9R-HHC-COOH | 9S-HHC-COOH | 9R-11-OH-HHC | 9S-11-OH-HHC | THC- COOH | 11-OH- THC |
|---|---|---|---|---|---|---|---|---|---|
| A | + | − | 222 | 186 (84%) | 112 (50%) | 33 (15%) | 0 (0%) | 204 (92%) | 178 (80%) |
| B | + | + | 10 | 10 (100%) | 3 (30%) | 1 (10%) | 1 (10%) | 10 (100%) | 7 (70%) |
| C | − | − | 76 | 2 (3%) | 0 (0%) | 0 (0%) | 0 (0%) | 1 (1%) | 0 (0%) |
| Compound | Median Concentration (ng/mL) | Maximum Concentration (ng/mL) |
|---|---|---|
| Δ9-THC | 2.8 | 35 |
| 11-OH-THC | 1.1 | 14 |
| THC-COOH | 21 | 269 |
| 9R-HHC-COOH a | 1.4 | 16 |
| 9S-HHC-COOH a | 0 | 4.4 |
| Δ9-THC | 11-OH-THC | THC-COOH | |||||||
|---|---|---|---|---|---|---|---|---|---|
| 20 min | 120 min | 180 min | 20 min | 120 min | 180 min | 20 min | 120 min | 180 min | |
| 9R-11-OH-HHC a | − | (+) | (+) | − | − | − | |||
| 9S-11-OH-HHC a | − | − | − | − | − | − | |||
| 9R-HHC-COOH | − | + | + | − | + | + | − | − | − |
| 9S-HHC-COOH | − | − | − | − | + | + | − | − | − |
| 11-OH-THC | + | + | + | − | − | − | |||
| THC-COOH | − | + | + | − | + | + | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Falck Jørgensen, C.; Schou Rasmussen, B.; Linnet, K.; Thomsen, R. Evidence of 11-Hydroxy-hexahydrocannabinol and 11-Nor-9-carboxy-hexahydrocannabinol as Novel Human Metabolites of Δ9-Tetrahydrocannabinol. Metabolites 2023, 13, 1169. https://doi.org/10.3390/metabo13121169
Falck Jørgensen C, Schou Rasmussen B, Linnet K, Thomsen R. Evidence of 11-Hydroxy-hexahydrocannabinol and 11-Nor-9-carboxy-hexahydrocannabinol as Novel Human Metabolites of Δ9-Tetrahydrocannabinol. Metabolites. 2023; 13(12):1169. https://doi.org/10.3390/metabo13121169
Chicago/Turabian StyleFalck Jørgensen, Christian, Brian Schou Rasmussen, Kristian Linnet, and Ragnar Thomsen. 2023. "Evidence of 11-Hydroxy-hexahydrocannabinol and 11-Nor-9-carboxy-hexahydrocannabinol as Novel Human Metabolites of Δ9-Tetrahydrocannabinol" Metabolites 13, no. 12: 1169. https://doi.org/10.3390/metabo13121169
APA StyleFalck Jørgensen, C., Schou Rasmussen, B., Linnet, K., & Thomsen, R. (2023). Evidence of 11-Hydroxy-hexahydrocannabinol and 11-Nor-9-carboxy-hexahydrocannabinol as Novel Human Metabolites of Δ9-Tetrahydrocannabinol. Metabolites, 13(12), 1169. https://doi.org/10.3390/metabo13121169