1H-NMR Based Serum Metabolomics Identifies Different Profile between Sarcopenia and Cancer Cachexia in Ageing Walker 256 Tumour-Bearing Rats

 , ,
, , 
Abstract
1. Introduction
2. Results
2.1. Morphometric Parameters
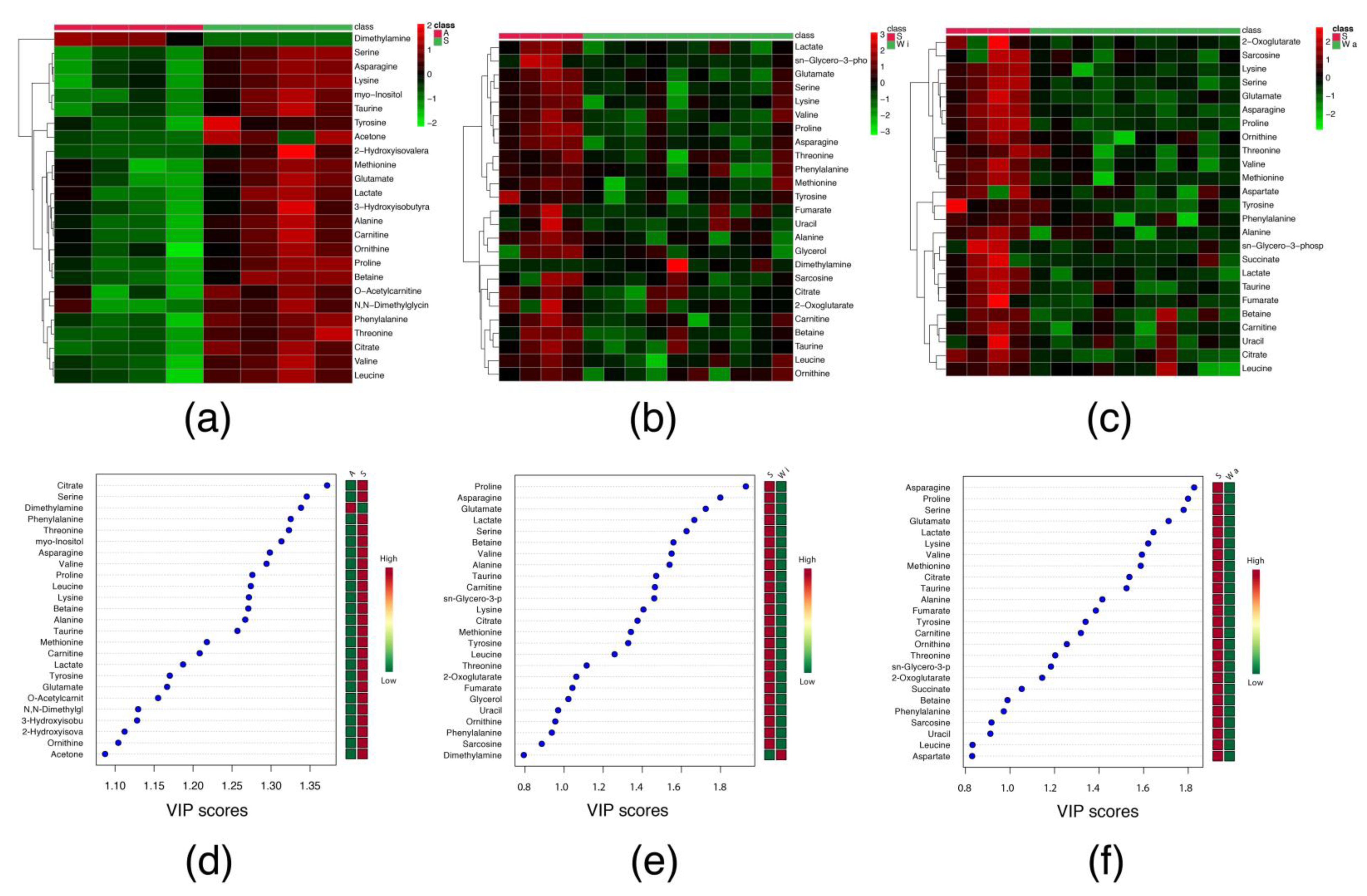
2.2. Serum Metabolomic Profile
2.2.1. Sarcopenia
2.2.2. Tumour Evolution
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Experimental Protocol
4.3. Metabolomic Analyses
4.3.1. Serum Preparation for 1H-NMR Acquisition
4.3.2. H-NMR Spectra Acquisition and Metabolic Quantification
4.3.3. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Baracos, V.E.; Martin, L.; Korc, M.; Guttridge, D.C.; Fearon, K.C.H. Cancer-associated cachexia. Nat. Rev. Dis. Prim. 2018, 4, 17105. [Google Scholar] [CrossRef]
- Dunne, R.F.; Loh, K.P.; Williams, G.R.; Jatoi, A.; Mustian, K.M.; Mohile, S.G. Cachexia and sarcopenia in older adults with cancer: A comprehensive review. Cancers 2019, 11, 1861. [Google Scholar] [CrossRef] [PubMed]
- Williams, G.R.; Rier, H.N.; McDonald, A.; Shachar, S.S. Sarcopenia & aging in cancer. J. Geriatr. Oncol. 2019, 10, 374–377. [Google Scholar]
- Loumaye, A.; Thissen, J.P. Biomarkers of cancer cachexia. Clin. Biochem. 2017, 50, 1281–1288. [Google Scholar] [CrossRef] [PubMed]
- Sakuma, K.; Aoi, W.; Yamaguchi, A. Molecular mechanism of sarcopenia and cachexia: Recent research advances. Pflug. Arch. 2017, 469, 573–591. [Google Scholar] [CrossRef] [PubMed]
- Cala, M.P.; Agulló-Ortuño, M.T.; Prieto-García, E.; González-Riano, C.; Parrilla-Rubio, L.; Barbas, C.; Díaz-García, C.V.; García, A.; Pernaut, C.; Adeva, J.; et al. Multiplatform plasma fingerprinting in cancer cachexia: A pilot observational and translational study. J. Cachexia Sarcopenia Muscle 2018, 9, 348–357. [Google Scholar] [CrossRef]
- Yang, Q.J.; Zhao, J.R.; Hao, J.; Li, B.; Huo, Y.; Han, Y.L.; Wan, L.L.; Li, J.; Huang, J.; Lu, J.; et al. Serum and urine metabolomics study reveals a distinct diagnostic model for cancer cachexia. J. Cachexia Sarcopenia Muscle 2018, 9, 71–85. [Google Scholar] [CrossRef]
- Boguszewicz, Ł.; Bieleń, A.; Mrochem-Kwarciak, J.; Skorupa, A.; Ciszek, M.; Heyda, A.; Wygoda, A.; Kotylak, A.; Składowski, K.; Sokół, M. NMR-based metabolomics in real-time monitoring of treatment induced toxicity and cachexia in head and neck cancer: A method for early detection of high risk patients. Metabolomics 2019, 15, 110. [Google Scholar] [CrossRef]
- Miller, J.; Alshehri, A.; Ramage, M.I.; Stephens, N.A.; Mullen, A.B.; Boyd, M.; Ross, J.A.; Wigmore, S.J.; Watson, D.G.; Skipworth, R.J.E. Plasma metabolomics identifies lipid and amino acid markers of weight loss in patients with upper gastrointestinal cancer. Cancers 2019, 11, 1594. [Google Scholar] [CrossRef]
- Quanjun, Y.; Genjin, Y.; Lili, W.; Bin, L.; Jin, L.; Qi, Y.; Yan, L.; Yonglong, H.; Cheng, G.; Junping, Z. Serum metabolic profiles reveal the effect of formoterol on cachexia in tumor-bearing mice. Mol. Biosyst. 2013, 9, 3015–3025. [Google Scholar] [CrossRef]
- Lautaoja, J.H.; Lalowski, M.; Nissinen, T.A.; Hentilä, J.; Shi, Y.; Ritvos, O.; Cheng, S.; Hulmi, J.J. Muscle and serum metabolomes are dysregulated in colon-26 tumor-bearing mice despite amelioration of cachexia with activin receptor type 2B ligand blockade. Am. J. Physiol. Endocrinol. Metab. 2019, 316, E852–E865. [Google Scholar] [CrossRef] [PubMed]
- Pin, F.; Barreto, R.; Couch, M.E.; Bonetto, A.; O’Connell, T.M. Cachexia induced by cancer and chemotherapy yield distinct perturbations to energy metabolism. J. Cachexia Sarcopenia Muscle 2019, 10, 140–154. [Google Scholar] [CrossRef] [PubMed]
- Viana, L.R.; Canevarolo, R.; Luiz, A.C.; Soares, R.F.; Lubaczeuski, C.; Zeri, A.C.; Gomes-Marcondes, M.C. Leucine-rich diet alters the 1H-NMR based metabolomic profile without changing the Walker-256 tumour mass in rats. BMC Cancer 2016, 16, 764. [Google Scholar] [CrossRef] [PubMed]
- Viana, L.R.; Tobar, N.; Busanello, E.N.B.; Marques, A.C.; de Oliveira, A.G.; Lima, T.I.; Machado, G.; Castelucci, B.G.; Ramos, C.D.; Brunetto, S.Q.; et al. Leucine-rich diet induces a shift in tumour metabolism from glycolytic towards oxidative phosphorylation, reducing glucose consumption and metastasis in Walker-256 tumour-bearing rats. Sci. Rep. 2019, 9, 15529. [Google Scholar] [CrossRef] [PubMed]
- Cruz, B.; Oliveira, A.; Gomes-Marcondes, M.C.C. L-leucine dietary supplementation modulates muscle protein degradation and increases pro-inflammatory cytokines in tumour-bearing rats. Cytokine 2017, 96, 253–260. [Google Scholar] [CrossRef]
- Cruz, B.; Oliveira, A.; Ventrucci, G.; Gomes-Marcondes, M.C.C. A leucine-rich diet modulates the mTOR cell signalling pathway in the gastrocnemius muscle under different Walker-256 tumour growth conditions. BMC Cancer 2019, 19, 349. [Google Scholar] [CrossRef]
- Modi, H.R.; Katyare, S.S.; Patel, M.A. Ageing-induced alterations in lipid/phospholipid profiles of rat brain and liver mitochondria: Implications for mitochondrial energy-linked functions. J. Membr. Biol. 2008, 221, 51–60. [Google Scholar] [CrossRef]
- Siu, P.M.; Pistilli, E.E.; Alway, S.E. Age-dependent increase in oxidative stress in gastrocnemius muscle with unloading. J. Appl. Physiol. 2008, 105, 1695–1705. [Google Scholar] [CrossRef]
- Nistiar, F.; Racz, O.; Lukacinova, A.; Hubkova, B.; Novakova, J.; Lovasova, E.; Sedlakova, E. Age dependency on some physiological and biochemical parameters of male Wistar rats in controlled environment. J. Environ. Sci. Health A Toxic Hazard. Subst. Environ. Eng. 2012, 47, 1224–1233. [Google Scholar] [CrossRef]
- Evans, W.J. Skeletal muscle loss: Cachexia, sarcopenia, and inactivity. Am. J. Clin. Nutr. 2010, 91, 1123S–1127S. [Google Scholar] [CrossRef]
- Dodson, S.; Baracos, V.E.; Jatoi, A.; Evans, W.J.; Cella, D.; Dalton, J.T.; Steiner, M.S. Muscle wasting in cancer cachexia: Clinical implications, diagnosis, and emerging treatment strategies. Annu. Rev. Med. 2011, 62, 265–279. [Google Scholar] [CrossRef]
- Ali, S.; Garcia, J.M. Sarcopenia, cachexia and aging: Diagnosis, mechanisms and therapeutic options—A mini-review. Gerontology 2014, 60, 294–305. [Google Scholar] [CrossRef]
- Fazelzadeh, P.; Hangelbroek, R.W.; Tieland, M.; de Groot, L.C.; Verdijk, L.B.; van Loon, L.J.; Smilde, A.K.; Alves, R.D.; Vervoort, J.; Müller, M.; et al. The muscle metabolome differs between healthy and frail older adults. J. Proteome Res. 2016, 15, 499–509. [Google Scholar] [CrossRef] [PubMed]
- Calvani, R.; Picca, A.; Marini, F.; Biancolillo, A.; Gervasoni, J.; Persichilli, S.; Primiano, A.; Coelho-Junior, H.J.; Bossola, M.; Urbani, A.; et al. A distinct pattern of circulating amino acids characterizes older persons with physical frailty and sarcopenia: Results from the BIOSPHERE study. Nutrients 2018, 10, 1691. [Google Scholar] [CrossRef] [PubMed]
- Kouchiwa, T.; Wada, K.; Uchiyama, M.; Kasezawa, N.; Niisato, M.; Murakami, H.; Fukuyama, K.; Yokogoshi, H. Age-related changes in serum amino acids concentrations in healthy individuals. Clin. Chem. Lab. Med. 2012, 50, 861–870. [Google Scholar] [CrossRef] [PubMed]
- Nyengaard, J.R.; Bendtsen, T.F. Glomerular number and size in relation to age, kidney weight, and body surface in normal man. Anat. Rec. 1992, 232, 194–201. [Google Scholar] [CrossRef]
- O’Sullivan, E.D.; Hughes, J.; Ferenbach, D.A. Renal Aging: Causes and Consequences. J. Am. Soc. Nephrol. 2017, 28, 407–420. [Google Scholar] [CrossRef]
- Li, R.; Dai, J.; Kang, H. The construction of a panel of serum amino acids for the identification of early chronic kidney disease patients. J. Clin. Lab. Anal. 2018, 32, e22282. [Google Scholar] [CrossRef]
- Zheng, H.; Dong, B.; Ning, J.; Shao, X.; Zhao, L.; Jiang, Q.; Ji, H.; Cai, A.; Xue, W.; Gao, H. NMR-based metabolomics analysis identifies discriminatory metabolic disturbances in tissue and biofluid samples for progressive prostate cancer. Clin. Chim. Acta 2020, 501, 241–251. [Google Scholar] [CrossRef]
- Malaguarnera, M.; Risino, C.; Gargante, M.P.; Oreste, G.; Barone, G.; Tomasello, A.V.; Costanzo, M.; Cannizzaro, M.A. Decrease of serum carnitine levels in patients with or without gastrointestinal cancer cachexia. World J. Gastroenterol. 2006, 12, 4541–4545. [Google Scholar] [CrossRef]
- Dando, I.; Pozza, E.D.; Ambrosini, G.; Torrens-Mas, M.; Butera, G.; Mullappilly, N.; Pacchiana, R.; Palmieri, M.; Donadelli, M. Oncometabolites in cancer aggressiveness and tumour repopulation. Biol. Rev. Camb. Philos. Soc. 2019, 94, 1530–1546. [Google Scholar] [CrossRef] [PubMed]
- QuanJun, Y.; Genjin, Y.; Lili, W.; Yang, Q.; Yonglong, H.; Jin, L.; Jie, L.; Jinlu, H.; Cheng, G. Integrated analysis of serum and intact muscle metabonomics identify metabolic profiles of cancer cachexia in a dynamic mouse model. RSC Adv. 2015, 5, 92438–92448. [Google Scholar] [CrossRef]
- Alborghetti, M.R.; Correa, M.E.P.; Whangbo, J.; Shi, X.; Aricetti, J.A.; da Silva, A.A.; Miranda, E.C.M.; Sforca, M.L.; Caldana, C.; Gerszten, R.E.; et al. Clinical metabolomics identifies blood serum branched chain amino acids as potential predictive biomarkers for chronic graft vs. host disease. Front. Oncol. 2019, 9, 141. [Google Scholar] [CrossRef] [PubMed]
- Altman, D.G.; Bland, J.M. Standard deviations and standard errors. BMJ 2005, 331, 903. [Google Scholar] [CrossRef]
- Chong, J.; Wishart, D.S.; Xia, J. Using MetaboAnalyst 4.0 for comprehensive and integrative metabolomics data analysis. Curr. Protoc. Bioinform. 2019, 68, e86. [Google Scholar] [CrossRef]
- Benjamini, Y.; Hochberg, Y. Controlling the false discovery rate: A practical and powerful approach to multiple testing. J. R. Stat. Soc. Ser. B 1995, 57, 449–518. [Google Scholar] [CrossRef]
- Castro, A.; Duft, R.G.; Ferreira, M.L.V.; Andrade, A.L.L.; Gáspari, A.F.; Silva, L.M.; Oliveira-Nunes, S.G.; Cavaglieri, C.R.; Ghosh, S.; Bouchard, C.; et al. Association of skeletal muscle and serum metabolites with maximum power output gains in response to continuous endurance or high-intensity interval training programs: The TIMES study—A randomized controlled trial. PLoS ONE 2019, 14, e0212115. [Google Scholar] [CrossRef]
- Xia, J.; Wishart, D.S. Web-based inference of biological patterns, functions and pathways from metabolomic data using MetaboAnalyst. Nat. Protoc. 2011, 6, 743–760. [Google Scholar] [CrossRef]


{kind=link}
| Morphometric Parameters | A | S | Wi | Wa | ||||
|---|---|---|---|---|---|---|---|---|
| Mean ± SD (Absolute) | Mean ± SD (Relative) | Mean ± SD (Absolute) | Mean ± SD (Relative) | Mean ± SD (Absolute) | Mean ± SD (Relative) | Mean ± SD (Absolute) | Mean ± SD (Relative) | |
| Initial body weight (g) | 245.100 ± 33.310 | - | 673.100 ± 71.110 | - | 642.700 ± 81.380 | - | 598.900 ± 56.990 | - |
| Final body weight (g) | 314.100 ± 24.250 | - | 705.000 ± 71.110 | - | 624.200 ± 93.180 | - | 570.900 ± 100.400 | - |
| Delta body weight (g) | 75.000 ± 16.800 | - | 31.700 ± 12.100 | - | −31.200 ± 31.020 | - | −51.500 ± 74.550 b | - |
| Gastrocnemius muscle weight (g) | 2.194 ± 0.189 | 0.009 ± 0.001 | 2.780 ± 0.565 | 0.004 ± 0.001 a | 2.729 ± 0.900 | 0.004 ± 0.001 | 2.638 ± 0.884 | 0.004 ± 0.001 |
| Liver weight (g) | 9.442 ± 1.214 | 0.038 ± 0.003 | 18.770 ± 2.434 | 0.027 ± 0.004 | 15.710 ± 3.482 | 0.024 ± 0.005 | 15.080 ± 4.538 | 0.025 ± 0.007 |
| Tumour weight (g) | - | - | - | - | 12.600 ± 5.825 | 0.020 ± 0.010 | 23.560 ± 9.083 | 0.040 ± 0.017 c |
| Metabolite | S versus A | Wi versus S | Wa versus S | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Mean ± SD (mM) | Mean ± SD (mM) | P-Value | Mean ± SD (mM) | Mean ± SD (mM) | P-Value | Mean ± SD (mM) | Mean ± SD (mM) | P-Value | |
| Alanine | 0.245 ± 0.047 | 0.090 ± 0.049 | <0.001 | 0.182 ± 0.037 | 0.245 ± 0.047 | 0.041 | 0.151 ± 0.029 | 0.245 ± 0.047 | 0.001 |
| Asparagine | 0.041 ± 0.007 | 0.016 ± 0.004 | <0.001 | 0.020 ± 0.008 | 0.041 ± 0.007 | <0.001 | 0.017 ± 0.004 | 0.041 ± 0.007 | <0.001 |
| Aspartate | 0.019 ± 0.003 | 0.009 ± 0.005 | 0.001 | 0.013 ± 0.003 | 0.019 ± 0.003 | 0.040 | 0.010 ± 0.003 | 0.019 ± 0.003 | 0.001 |
| Betaine | 0.112 ± 0.027 | 0.035 ± 0.020 | 0.005 | 0.066 ± 0.024 | 0.112 ± 0.027 | 0.059 | 0.072 ± 0.036 | 0.112 ± 0.027 | 0.119 |
| Carnitine | 0.027 ± 0.006 | 0.010 ± 0.006 | 0.001 | 0.018 ± 0.004 | 0.027 ± 0.006 | 0.019 | 0.016 ± 0.005 | 0.027 ± 0.006 | 0.006 |
| Citrate | 0.147 ± 0.020 | 0.038 ± 0.020 | <0.001 | 0.097 ± 0.036 | 0.147 ± 0.020 | 0.044 | 0.080 ± 0.029 | 0.147 ± 0.020 | 0.005 |
| Dimethylamine | 0.001 ± 0.0001 | 0.027 ± 0.008 | <0.001 | 0.002 ± 0.002 | 0.001 ± 0.0001 | 0.946 | 0.002 ± 0.002 | 0.001 ± 0.0001 | 0.923 |
| Fumarate | 0.003 ± 0.002 | 0.002 ± 0.002 | 0.930 | 0.001 ± 0.001 | 0.003 ± 0.002 | 0.185 | 0.001 ± 0.0005 | 0.003 ± 0.002 | 0.032 |
| Glutamate | 0.087 ± 0.023 | 0.036 ± 0.022 | 0.007 | 0.056 ± 0.025 | 0.087 ± 0.023 | 0.063 | 0.037 ± 0.010 | 0.087 ± 0.023 | 0.001 |
| Glutamine | 0.374 ± 0.065 | 0.107 ± 0.061 | <0.001 | 0.238 ± 0.066 | 0.374 ± 0.065 | 0.014 | 0.192 ± 0.076 | 0.374 ± 0.065 | 0.001 |
| Glycine | 0.141 ± 0.027 | 0.052 ± 0.027 | <0.001 | 0.086 ± 0.017 | 0.141 ± 0.027 | 0.008 | 0.095 ± 0.032 | 0.141 ± 0.027 | 0.029 |
| Lactate | 5.588 ± 1.510 | 1.720 ± 1.541 | <0.001 | 3.192 ± 0.856 | 5.588 ± 1.510 | 0.002 | 2.404 ± 0.499 | 5.588 ± 1.510 | <0.001 |
| Leucine | 0.063 ± 0.007 | 0.027 ± 0.013 | 0.005 | 0.047 ± 0.011 | 0.063 ± 0.007 | 0.239 | 0.051 ± 0.017 | 0.063 ± 0.007 | 0.500 |
| Lysine | 0.163 ± 0.027 | 0.088 ± 0.056 | 0.008 | 0.117 ± 0.027 | 0.163 ± 0.027 | 0.069 | 0.093 ± 0.018 | 0.163 ± 0.027 | 0.003 |
| Methionine | 0.030 ± 0.004 | 0.014 ± 0.007 | <0.001 | 0.021 ± 0.006 | 0.030 ± 0.004 | 0.019 | 0.018 ± 0.002 | 0.030 ± 0.004 | 0.001 |
| Phenylalanine | 0.034 ± 0.003 | 0.013 ± 0.006 | <0.001 | 0.029 ± 0.007 | 0.034 ± 0.003 | 0.480 | 0.026 ± 0.004 | 0.034 ± 0.003 | 0.127 |
| Proline | 0.127 ± 0.025 | 0.046 ± 0.023 | <0.001 | 0.062 ± 0.019 | 0.127 ± 0.025 | <0.001 | 0.058 ± 0.011 | 0.127 ± 0.025 | <0.001 |
| Pyruvate | 0.165 ± 0.022 | 0.022 ± 0.017 | <0.001 | 0.102 ± 0.032 | 0.165 ± 0.022 | 0.002 | 0.100 ± 0.022 | 0.165 ± 0.022 | 0.001 |
| Serine | 0.124 ± 0.022 | 0.028 ± 0.019 | <0.001 | 0.073 ± 0.020 | 0.124 ± 0.022 | <0.001 | 0.060 ± 0.013 | 0.124 ± 0.022 | <0.001 |
| Taurine | 0.304 ± 0.088 | 0.128 ± 0.082 | 0.022 | 0.170 ± 0.076 | 0.304 ± 0.088 | 0.040 | 0.148 ± 0.078 | 0.304 ± 0.088 | 0.014 |
| Threonine | 0.164 ± 0.034 | 0.052 ± 0.019 | 0.002 | 0.119 ± 0.037 | 0.164 ± 0.034 | 0.248 | 0.092 ± 0.047 | 0.164 ± 0.034 | 0.024 |
| Tyrosine | 0.049 ± 0.014 | 0.020 ± 0.009 | <0.001 | 0.035 ± 0.007 | 0.049 ± 0.014 | 0.052 | 0.031 ± 0.007 | 0.049 ± 0.014 | 0.011 |
| Valine | 0.093 ± 0.014 | 0.039 ± 0.017 | <0.001 | 0.062 ± 0.018 | 0.093 ± 0.014 | 0.018 | 0.052 ± 0.015 | 0.093 ± 0.014 | 0.001 |
| Pathway | Metabolite | S versus A | Wi versus S | Wa versus S | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Match Status | Regulation | Adjusted P-Value * | Match Status | Regulation | Adjusted P-Value * | Match Status | Regulation | Adjusted P-Value * | ||
| Aminoacyl-tRNA biosynthesis | Alanine | 15/48 | ↑ | <0.001 | 9/48 | ↓ | <0.001 | 13/48 | ↓ | <0.001 |
| Asparagine | ↑ | ↓ | ↓ | |||||||
| Aspartate | ↑ | ↓ | ↓ | |||||||
| Glutamate | ↑ | ns | ↓ | |||||||
| Glutamine | ↑ | ↓ | ↓ | |||||||
| Glycine | ↑ | ↓ | ↓ | |||||||
| Leucine | ↑ | ns | ns | |||||||
| Lysine | ↑ | ns | ↓ | |||||||
| Methionine | ↑ | ↓ | ↓ | |||||||
| Phenylalanine | ↑ | ns | ns | |||||||
| Proline | ↑ | ↓ | ↓ | |||||||
| Serine | ↑ | ↓ | ↓ | |||||||
| Threonine | ↑ | ns | ↓ | |||||||
| Tyrosine | ↑ | ns | ↓ | |||||||
| Valine | ↑ | ↓ | ↓ | |||||||
| Alanine, aspartate and glutamate metabolism | Alanine | 7/48 | ↑ | <0.001 | 6/28 | ↓ | <0.001 | 8/28 | ↓ | <0.001 |
| Asparagine | ↑ | ↓ | ↓ | |||||||
| Aspartate | ↑ | ↓ | ↓ | |||||||
| Citrate | ↑ | ↓ | ↓ | |||||||
| Fumarate | ns | ns | ↓ | |||||||
| Glutamate | ↑ | ns | ↓ | |||||||
| Glutamine | ↑ | ↓ | ↓ | |||||||
| Pyruvate | ↑ | ↓ | ↓ | |||||||
| Glyoxylate and dicarboxylate metabolism | Citrate | 6/32 | ↑ | <0.001 | 5/32 | ↓ | <0.001 | 6/32 | ↓ | <0.001 |
| Glutamate | ↑ | ns | ↓ | |||||||
| Glutamine | ↑ | ↓ | ↓ | |||||||
| Glycine | ↑ | ↓ | ↓ | |||||||
| Pyruvate | ↑ | ↓ | ↓ | |||||||
| Serine | ↑ | ↓ | ↓ | |||||||
| Glycine, serine and threonine metabolism | Betaine | 5/34 | ↑ | 0.001 | 3/34 | ns | 0.054 | 4/34 | ns | 0.011 |
| Glycine | ↑ | ↓ | ↓ | |||||||
| Pyruvate | ↑ | ↓ | ↓ | |||||||
| Serine | ↑ | ↓ | ↓ | |||||||
| Threonine | ↑ | ns | ↓ | |||||||
| Valine, leucine and isoleucine biosynthesis | Leucine | 3/8 | ↑ | 0.002 | 1/8 | ns | 0.403 | 2/8 | ns | 0.033 |
| Threonine | ↑ | ns | ↓ | |||||||
| Valine | ↑ | ↓ | ↓ | |||||||
| Arginine biosynthesis | Aspartate | 3/14 | ↑ | 0.012 | 2/14 | ↓ | 0.095 | 4/14 | ↓ | <0.001 |
| Fumarate | ns | ns | ↓ | |||||||
| Glutamate | ↑ | ns | ↓ | |||||||
| Glutamine | ↑ | ↓ | ↓ | |||||||
| Phenylalanine, tyrosine and tryptophan biosynthesis | Phenylalanine | 2/4 | ↑ | 0.014 | 0/4 | ns | ns | 1/4 | ns | 0.244 |
| Tyrosine | ↑ | ns | ↓ | |||||||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Viana, L.R.; Lopes-Aguiar, L.; Rossi Rosolen, R.; Willians dos Santos, R.; Cintra Gomes-Marcondes, M.C. 1H-NMR Based Serum Metabolomics Identifies Different Profile between Sarcopenia and Cancer Cachexia in Ageing Walker 256 Tumour-Bearing Rats. Metabolites 2020, 10, 161. https://doi.org/10.3390/metabo10040161
Viana LR, Lopes-Aguiar L, Rossi Rosolen R, Willians dos Santos R, Cintra Gomes-Marcondes MC. 1H-NMR Based Serum Metabolomics Identifies Different Profile between Sarcopenia and Cancer Cachexia in Ageing Walker 256 Tumour-Bearing Rats. Metabolites. 2020; 10(4):161. https://doi.org/10.3390/metabo10040161
Chicago/Turabian StyleViana, Laís Rosa, Leisa Lopes-Aguiar, Rafaela Rossi Rosolen, Rogerio Willians dos Santos, and Maria Cristina Cintra Gomes-Marcondes. 2020. "1H-NMR Based Serum Metabolomics Identifies Different Profile between Sarcopenia and Cancer Cachexia in Ageing Walker 256 Tumour-Bearing Rats" Metabolites 10, no. 4: 161. https://doi.org/10.3390/metabo10040161
APA StyleViana, L. R., Lopes-Aguiar, L., Rossi Rosolen, R., Willians dos Santos, R., & Cintra Gomes-Marcondes, M. C. (2020). 1H-NMR Based Serum Metabolomics Identifies Different Profile between Sarcopenia and Cancer Cachexia in Ageing Walker 256 Tumour-Bearing Rats. Metabolites, 10(4), 161. https://doi.org/10.3390/metabo10040161