
Thermal Characterization and Interaction of the Subunits from the Multimeric Bacteriophage Endolysin PlyC
 ,
,
Abstract
:Simple Summary
Abstract
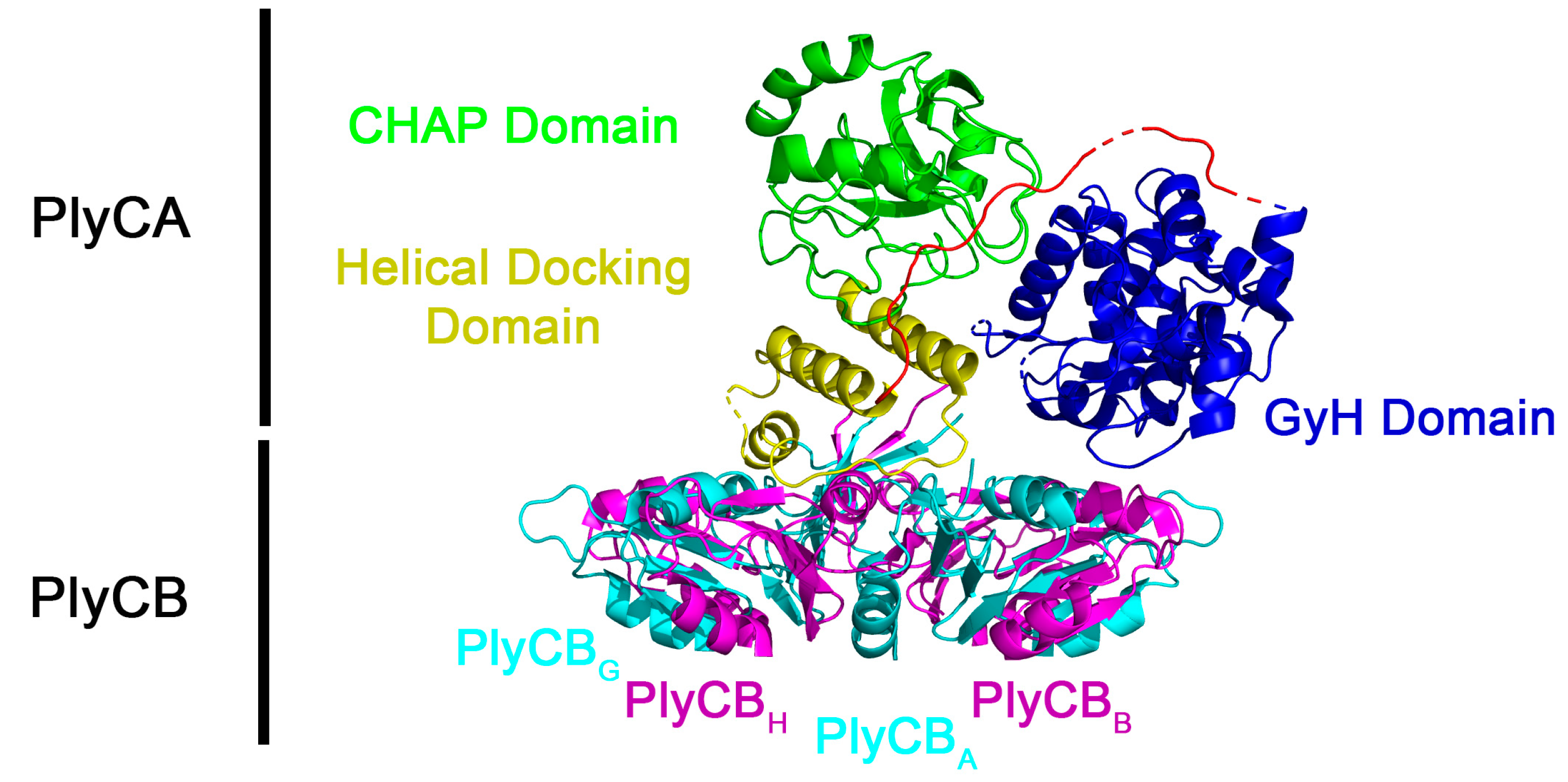
1. Introduction
2. Materials and Methods
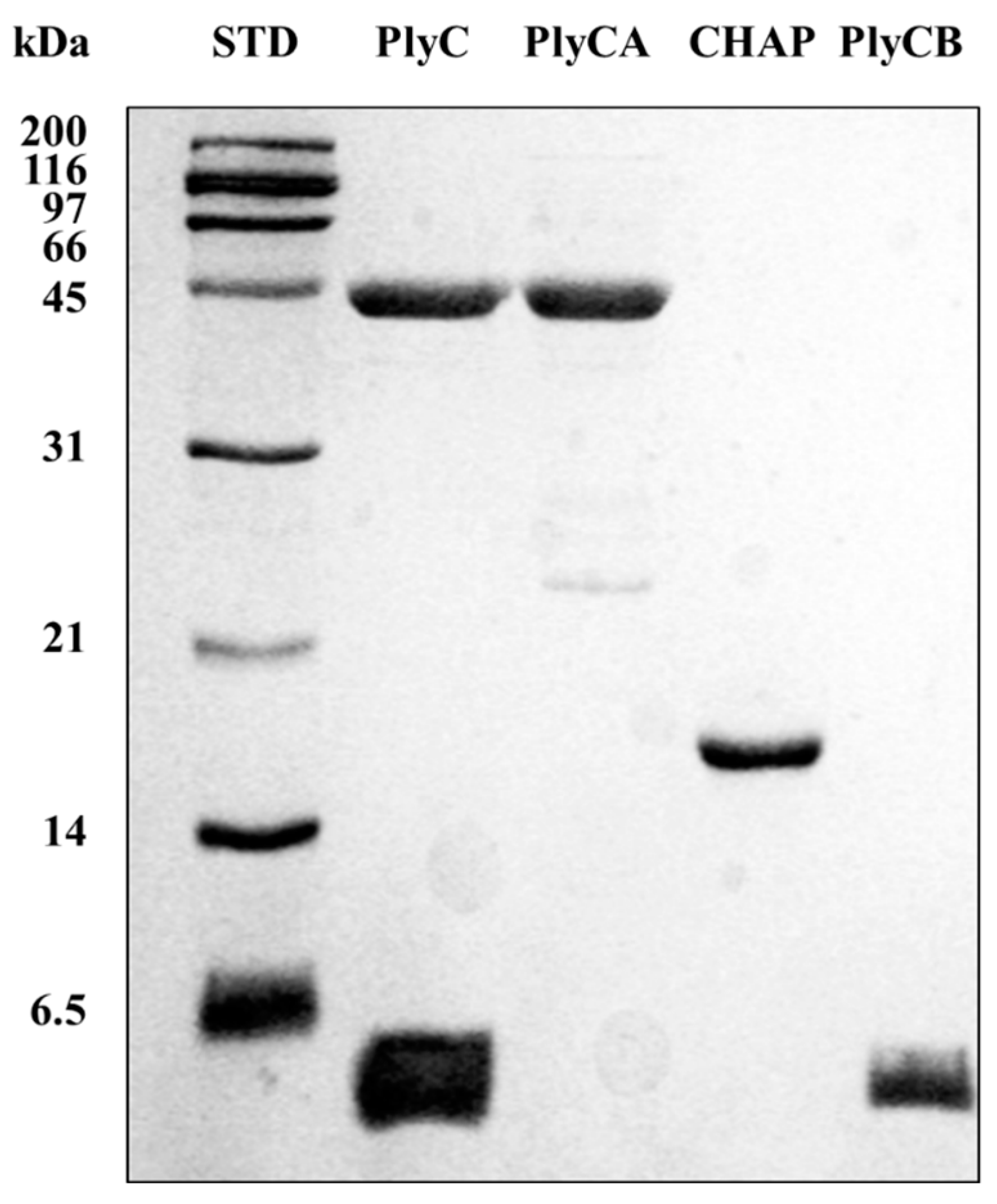
2.1. Bacterial Strains, Culture Conditions, Expression, and Purification
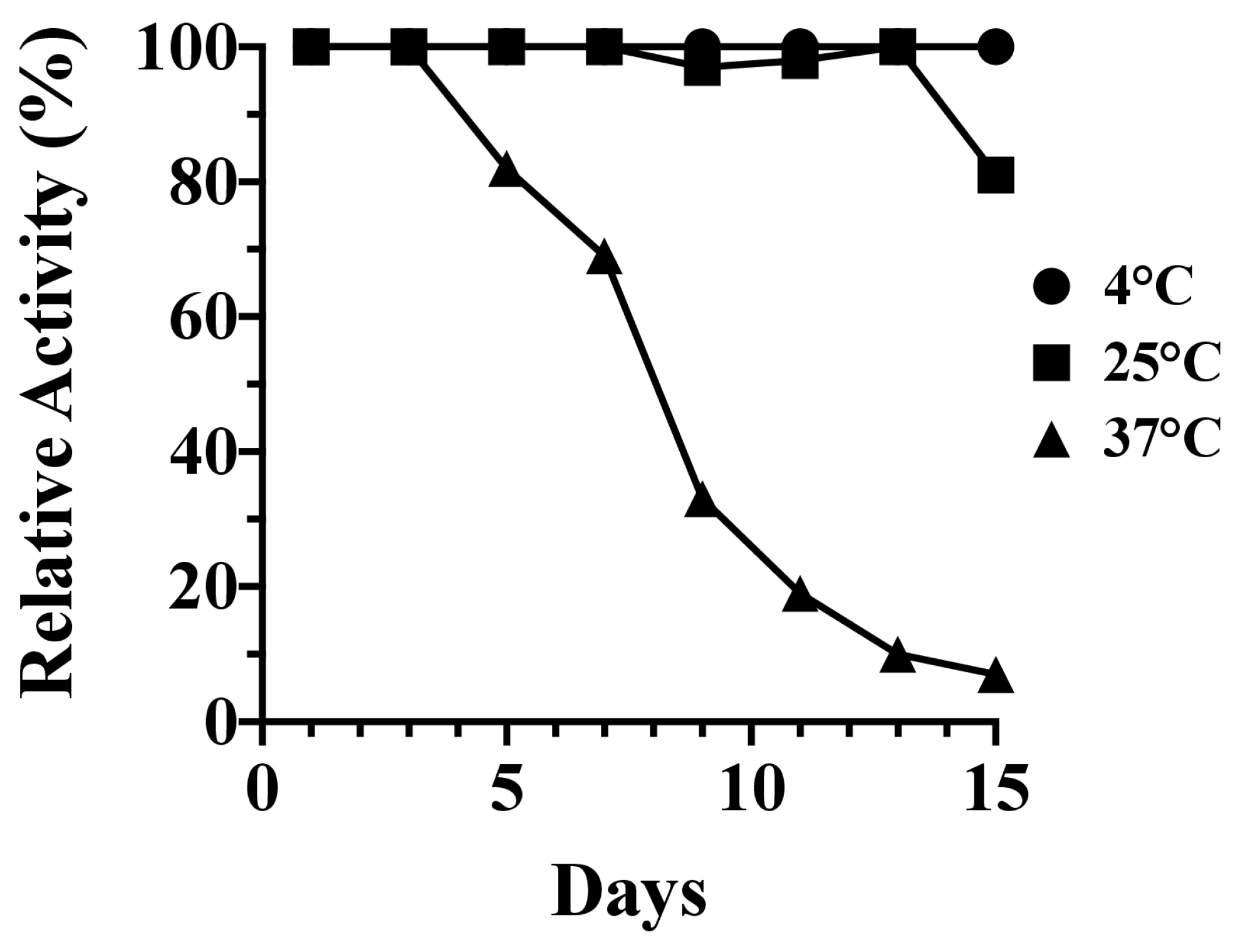
2.2. Fifteen-Day Kinetic Stability Assay
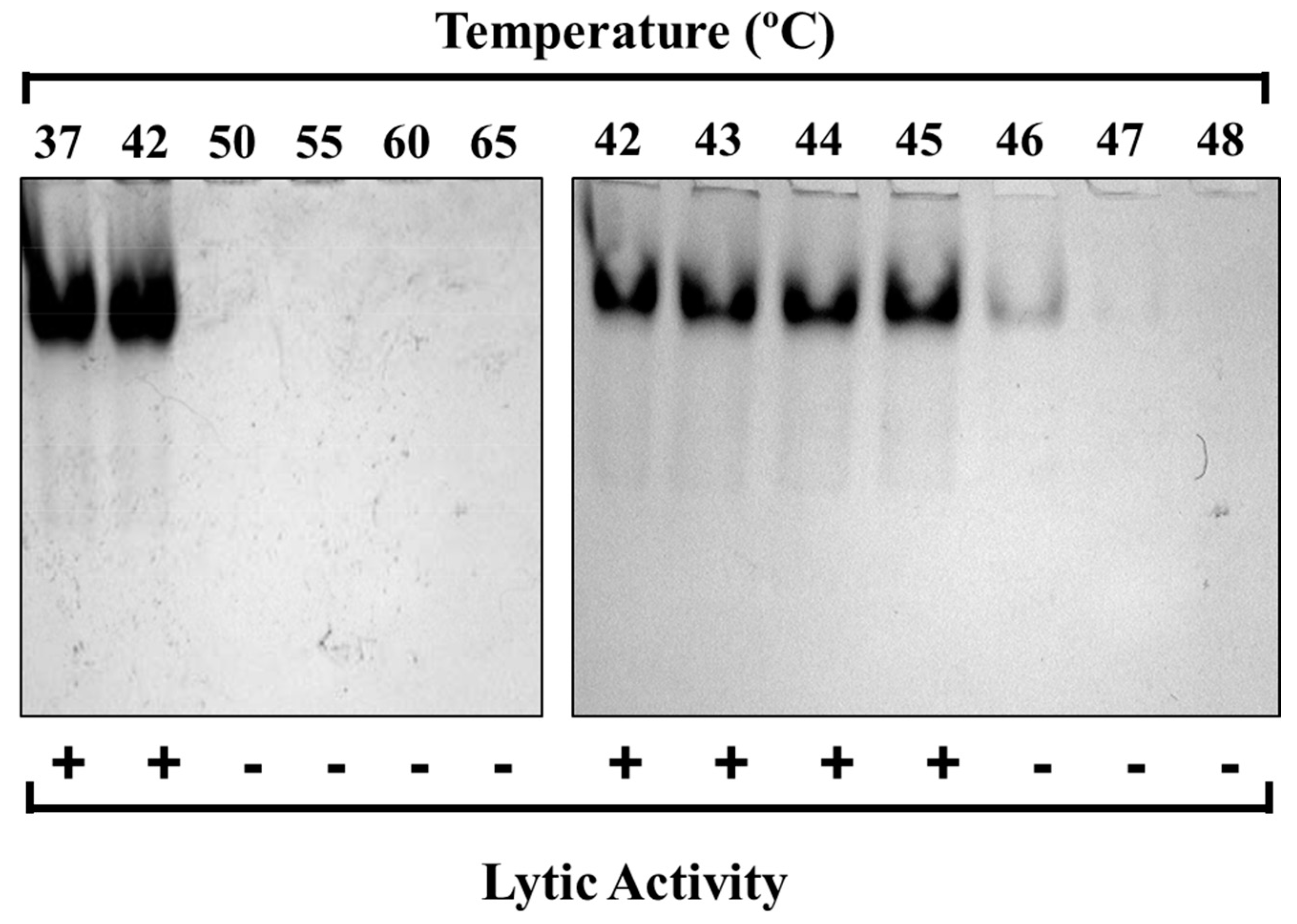
2.3. Thermal Denaturation of PlyC
2.4. Differential Scanning Calorimetry
2.5. PlyCB Cross-Linking
2.6. Bright Field and Fluorescence Microscopy
2.7. Thermal Regeneration of Native PlyC from Its Constituent Subunits
3. Results
3.1. Kinetic Stability and Thermal Denaturation of PlyC
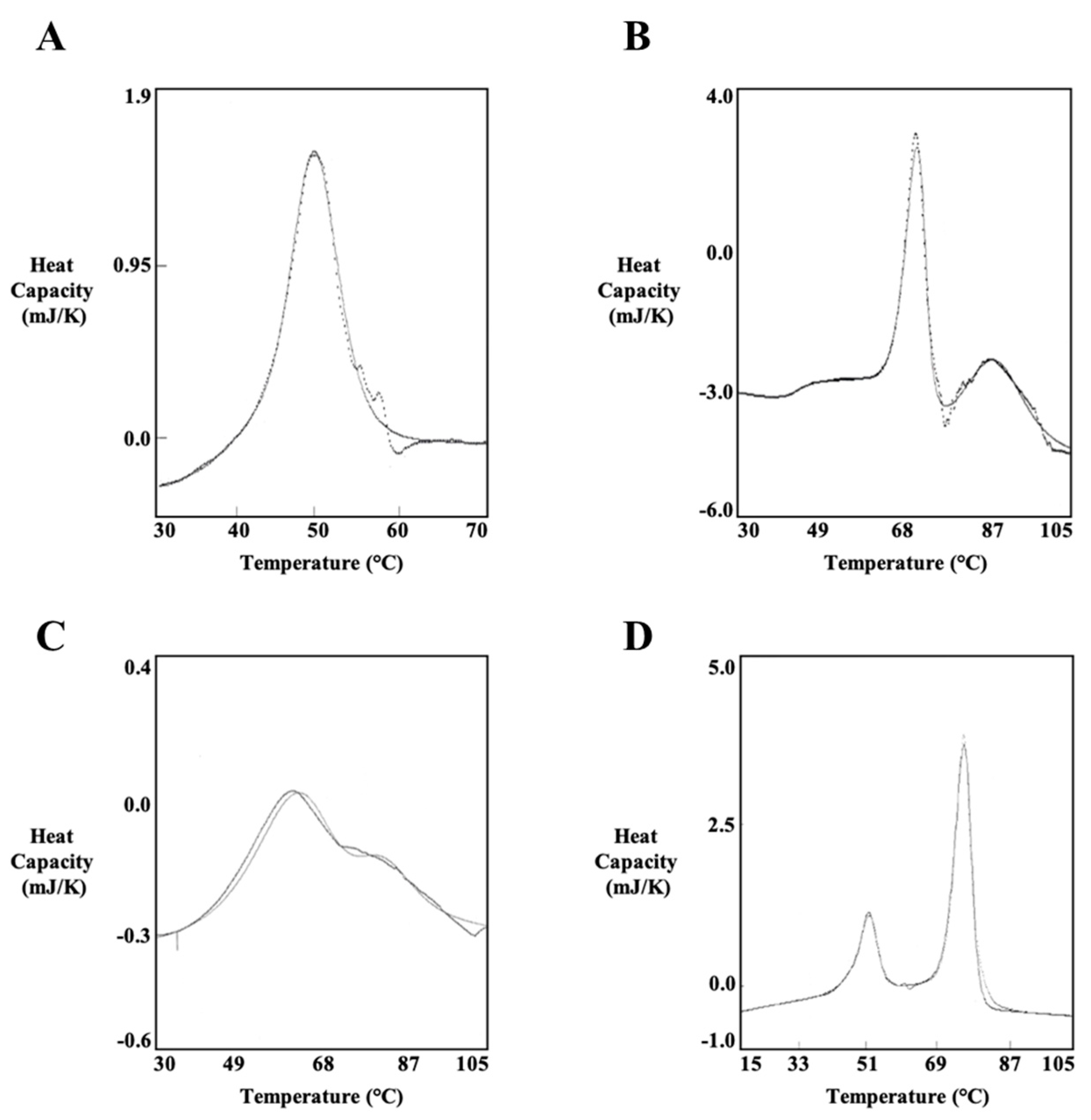
3.2. Thermodynamic Characterization of PlyC
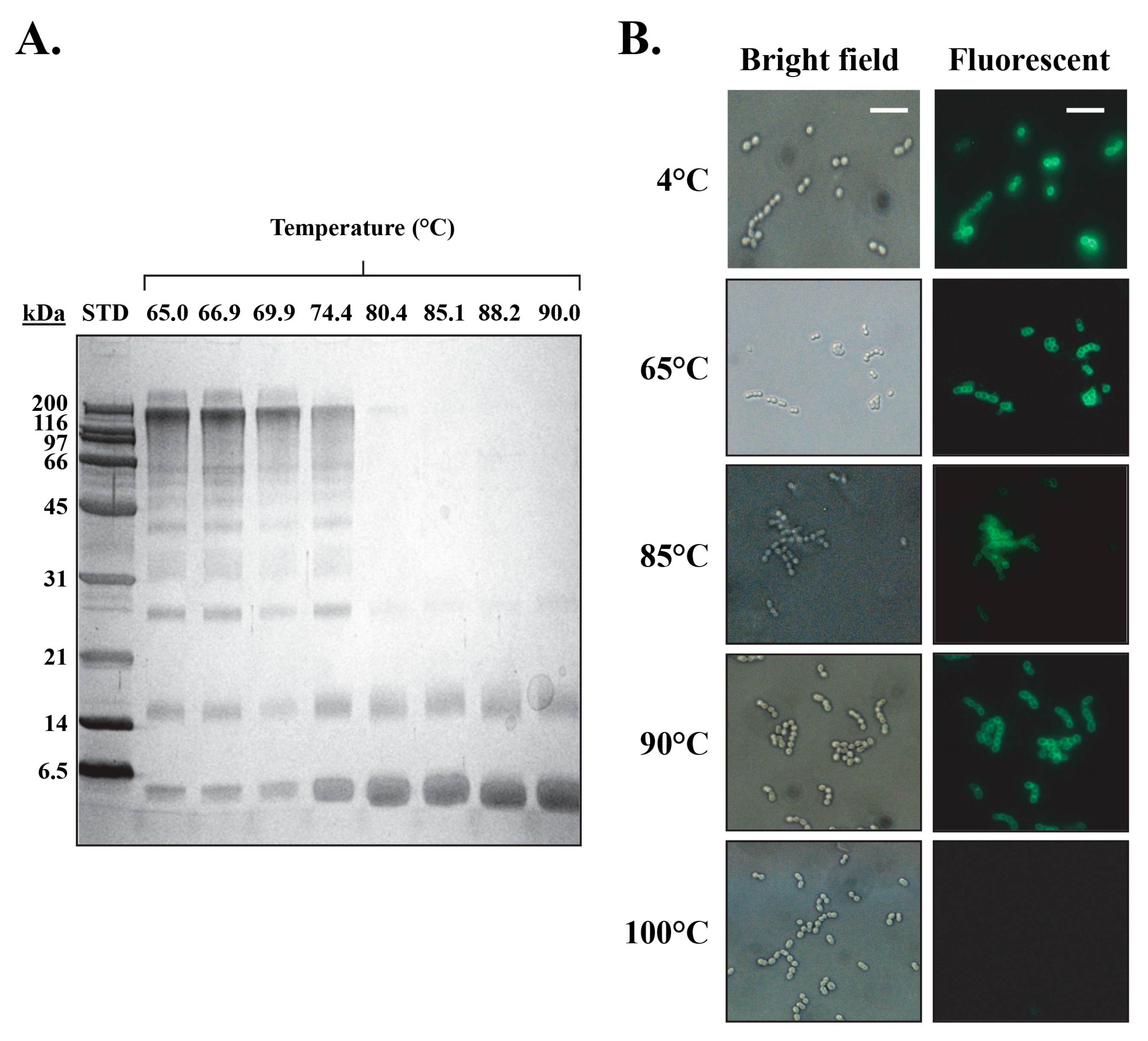
3.3. PlyCB Octamer Structural Integrity and Kinetic Stability as a Function of Temperature
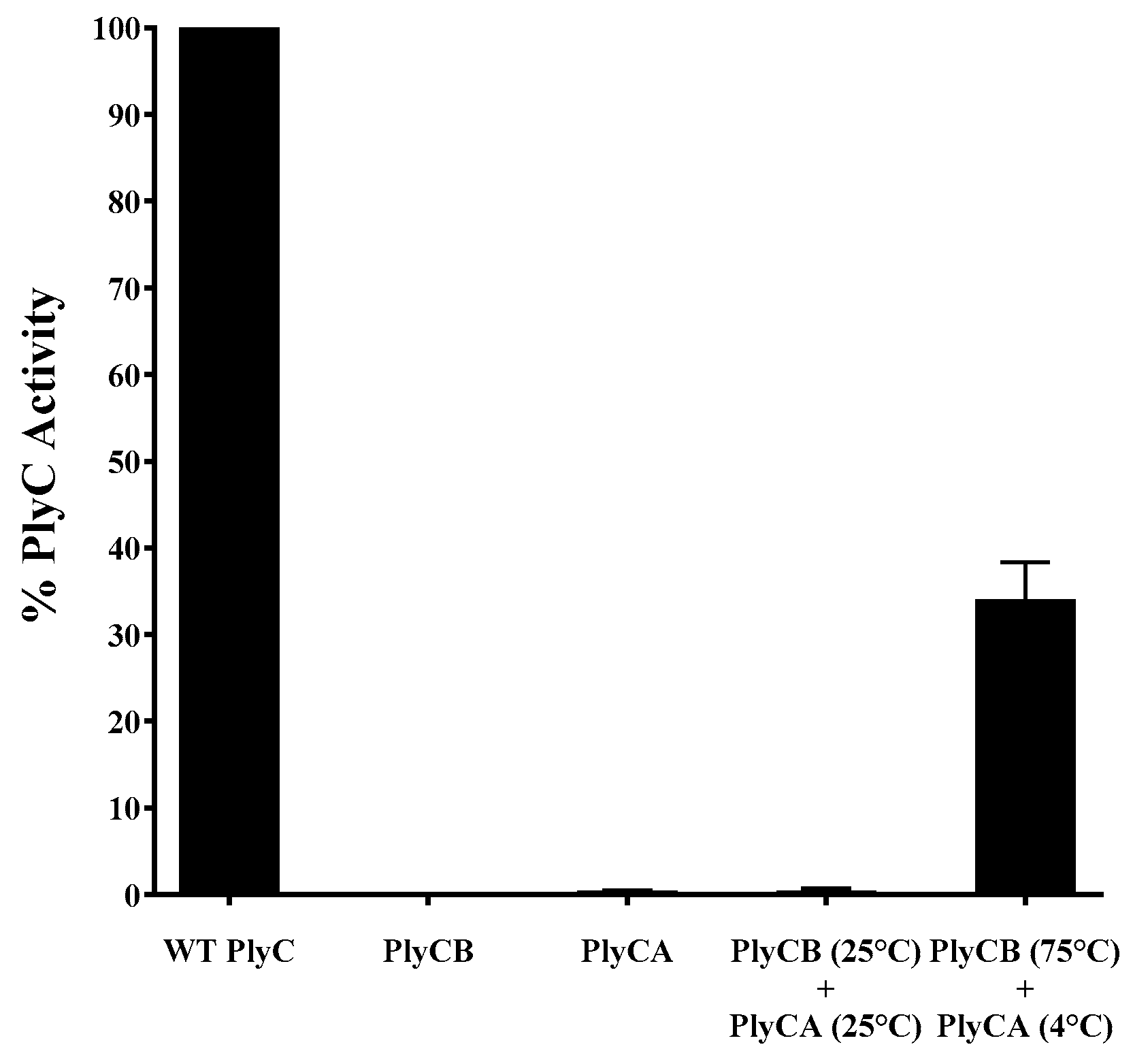
3.4. Refolding of the PlyC Holoenzyme
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- CDC. Antibiotic Resistance Threats in the United States, 2019; U.S. Department of Health and Human Services: Atlanta, GA, USA, 2019.
- Ikuta, K.S.; Swetschinski, L.R.; Aguilar, G.R.; Sharara, F.; Mestrovic, T.; Gray, A.P.; Weaver, N.D.; Wool, E.E.; Han, C.; Hayoon, A.G.; et al. Global mortality associated with 33 bacterial pathogens in 2019: A systematic analysis for the Global Burden of Disease Study 2019. Lancet 2022, 400, 2221–2248. [Google Scholar] [CrossRef] [PubMed]
- Czaplewski, L.; Bax, R.; Clokie, M.; Dawson, M.; Fairhead, H.; Fischetti, V.A.; Foster, S.; Gilmore, B.F.; Hancock, R.E.; Harper, D.; et al. Alternatives to antibiotics-a pipeline portfolio review. Lancet Infect. Dis. 2016, 16, 239–251. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Rivers, J.; Mathis, S.; Li, Z.; McGee, L.; Chochua, S.; Metcalf, B.J.; Fleming-Dutra, K.E.; Nanduri, S.A.; Beall, B. Continued increase of erythromycin nonsusceptibility and clindamycin nonsusceptibility among invasive Group A Streptococci driven by genomic clusters, United States, 2018–2019. Clin. Infect. Dis. 2023, 76, e1266–e1269. [Google Scholar] [CrossRef] [PubMed]
- Abdelkader, K.; Gerstmans, H.; Saafan, A.; Dishisha, T.; Briers, Y. The preclinical and clinical progress of bacteriophages and their lytic enzymes: The parts are easier than the whole. Viruses 2019, 11, 96. [Google Scholar] [CrossRef]
- Schmelcher, M.; Loessner, M.J. Bacteriophage endolysins—Extending their application to tissues and the bloodstream. Curr. Opin. Biotechnol. 2021, 68, 51–59. [Google Scholar] [CrossRef]
- Danis-Wlodarczyk, K.M.; Wozniak, D.J.; Abedon, S.T. Treating bacterial infections with bacteriophage-based enzybiotics: In vitro, in vivo and clinical application. Antibiotics 2021, 10, 1497. [Google Scholar] [CrossRef]
- Ho, M.K.Y.; Zhang, P.; Chen, X.; Xia, J.; Leung, S.S.Y. Bacteriophage endolysins against gram-positive bacteria, an overview on the clinical development and recent advances on the delivery and formulation strategies. Crit. Rev. Microbiol. 2022, 48, 303–326. [Google Scholar] [CrossRef]
- Murray, E.; Draper, L.A.; Ross, R.P.; Hill, C. The advantages and challenges of using endolysins in a clinical setting. Viruses 2021, 13, 680. [Google Scholar] [CrossRef]
- Wong, K.Y.; Megat Mazhar Khair, M.H.; Song, A.A.; Masarudin, M.J.; Chong, C.M.; In, L.L.A.; Teo, M.Y.M. Endolysins against streptococci as an antibiotic alternative. Front. Microbiol. 2022, 13, 935145. [Google Scholar] [CrossRef]
- Liu, B.; Guo, Q.; Li, Z.; Guo, X.; Liu, X. Bacteriophage endolysin: A powerful weapon to control bacterial biofilms. Protein J. 2023, 42, 463–476. [Google Scholar] [CrossRef]
- Shen, Y.; Barros, M.; Vennemann, T.; Gallagher, D.T.; Yin, Y.; Linden, S.B.; Heselpoth, R.D.; Spencer, D.J.; Donovan, D.M.; Moult, J.; et al. A bacteriophage endolysin that eliminates intracellular streptococci. eLife 2016, 5, e13152. [Google Scholar] [CrossRef] [PubMed]
- Nelson, D.; Loomis, L.; Fischetti, V.A. Prevention and elimination of upper respiratory colonization of mice by group A streptococci by using a bacteriophage lytic enzyme. Proc. Natl. Acad. Sci. USA 2001, 98, 4107–4112. [Google Scholar] [CrossRef] [PubMed]
- McGowan, S.; Buckle, A.M.; Mitchell, M.S.; Hoopes, J.T.; Gallagher, D.T.; Heselpoth, R.D.; Shen, Y.; Reboul, C.F.; Law, R.H.; Fischetti, V.A.; et al. X-ray crystal structure of the streptococcal specific phage lysin PlyC. Proc. Natl. Acad. Sci. USA 2012, 109, 12752–12757. [Google Scholar] [CrossRef] [PubMed]
- Broendum, S.S.; Williams, D.E.; Hayes, B.K.; Kraus, F.; Fodor, J.; Clifton, B.E.; Geert Volbeda, A.; Codee, J.D.C.; Riley, B.T.; Drinkwater, N.; et al. High avidity drives the interaction between the streptococcal C1 phage endolysin, PlyC, with the cell surface carbohydrates of Group A Streptococcus. Mol. Microbiol. 2021, 116, 397–415. [Google Scholar] [CrossRef] [PubMed]
- Anderson, G.; Scott, M. Determination of product shelf life and activation energy for five drugs of abuse. Clin. Chem. 1991, 37, 398–402. [Google Scholar] [CrossRef]
- Blanco, I.; Abate, L.; Antonelli, M.L. The regression of isothermal thermogravimetric data to evaluate degradation Ea values of polymers: A comparison with literature methods and an evaluation of lifetime prediction reliability. Polym. Degrad. Stab. 2011, 96, 1947–1954. [Google Scholar] [CrossRef]
- Kirchhoff, W.H. Exam: A two-state thermodynamic analysis program. NIST Tech. Note 1993, 1401, 1–103. [Google Scholar]
- Manly, S.P.; Matthews, K.S.; Sturtevant, J.M. Thermal denaturation of the core protein of lac repressor. Biochemistry 1985, 24, 3842–3846. [Google Scholar] [CrossRef]
- Fischetti, V.A.; Gotschlich, E.C.; Bernheimer, A.W. Purification and physical properties of group C streptococcal phage-associated lysin. J. Exp. Med. 1971, 133, 1105–1117. [Google Scholar] [CrossRef]
- Heselpoth, R.D.; Yin, Y.; Moult, J.; Nelson, D.C. Increasing the stability of the bacteriophage endolysin PlyC using rationale-based FoldX computational modeling. Protein Eng. Des. Sel. 2015, 28, 85–92. [Google Scholar] [CrossRef]
- Heselpoth, R.D.; Nelson, D.C. A new screening method for the directed evolution of thermostable bacteriolytic enzymes. J. Vis. Exp. 2012. [Google Scholar] [CrossRef]
- Kim, K.S.; Kim, S.; Yang, H.J.; Kwon, D.Y. Changes of glycinin conformation due to pH, heat and salt determined by differential scanning calorimetry and circular dichroism. Int. J. Food Sci. Technol. 2004, 39, 385–393. [Google Scholar] [CrossRef]
- Yamasaki, M.; Yano, H.; Aoki, K. Differential scanning calorimetric studies on bovine serum albumin: I. Effects of pH and ionic strength. Int. J. Biol. Macromol. 1990, 12, 263–268. [Google Scholar] [CrossRef] [PubMed]
- Richard, A.J.; Liu, C.C.; Klinger, A.L.; Todd, M.J.; Mezzasalma, T.M.; LiCata, V.J. Thermal stability landscape for Klenow DNA polymerase as a function of pH and salt concentration. Biochim. Biophys. Acta 2006, 1764, 1546–1552. [Google Scholar] [CrossRef] [PubMed]
- Bhat, R.; Timasheff, S.N. Steric Exclusion Is the Principal Source of the Preferential Hydration of Proteins in the Presence of Polyethylene Glycols. Protein Sci. 1992, 1, 1133–1143. [Google Scholar] [CrossRef]
- Nelson, D.; Schuch, R.; Chahales, P.; Zhu, S.; Fischetti, V.A. PlyC: A multimeric bacteriophage lysin. Proc. Natl. Acad. Sci. USA 2006, 103, 10765–10770. [Google Scholar] [CrossRef]
- Kumar, S.; Tsai, C.J.; Nussinov, R. Factors enhancing protein thermostability. Protein Eng. 2000, 13, 179–191. [Google Scholar] [CrossRef]
- Niu, C.; Zhu, L.; Xu, X.; Li, Q. Rational design of disulfide bonds increases thermostability of a mesophilic 1,3-1,4-beta-glucanase from Bacillus terquilensis. PLoS ONE 2016, 11, e0154036. [Google Scholar] [CrossRef]
- Liu, L.; Deng, Z.; Yang, H.; Li, J.; Shin, H.D.; Chen, R.R.; Du, G.; Chen, J. In silico rational design and systems engineering of disulfide bridges in the catalytic domain of an alkaline alpha-amylase from Alkalimonas amylolytica to improve thermostability. Appl. Environ. Microbiol. 2014, 80, 798–807. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Conc. (mg/mL) | Scan Rate (K/h) | Model | TG (°C) | ∆HvH (kJ/mol) | ∆Hcal (kJ/mol) | Ratio a |
|---|---|---|---|---|---|---|
| 1.0–2.0 | 15 | A ⟷ I | 41.4 ± 1.7 | 845 ± 154 | 1633 ± 163 | 1.08 ± 0.25 |
| I ⟷ B | 44.6 ± 0.5 | 674 ± 194 | ||||
| 1.0–2.0 | 60 | A ⟷ I | 42.0 ± 2.3 | 302 ± 76 | 922 ± 205 | 1.09 ± 0.26 |
| I ⟷ B | 48.6 ± 0.2 | 545 ± 81 |
| Conc. (mg/mL) | Scan Rate (K/h) | Model | TG (°C) | ∆HvH (kJ/mol) | ∆Hcal (kJ/mol) | Ratio a |
|---|---|---|---|---|---|---|
| 1.0 | 15 | A ⟷ I | 38.5 ± 0.5 | 464 ± 2 | 439 ± 44 | 0.42 ± 0.04 |
| I ⟷ B | 44.3 ± 0.5 | 583 ± 58 | ||||
| 1.0–2.0 | 60 | A ⟷ I | 39.1 ± 0.4 | 430 ± 70 | 394 ± 3 | 0.40 ± 0.07 |
| I ⟷ B | 46.1 ± 0.6 | 554 ± 6 |
| Conc. (mg/mL) | Scan Rate (K/h) | Model | TG (°C) | ∆HvH (kJ/mol) | ∆Hcal (kJ/mol) | Ratio a | |
|---|---|---|---|---|---|---|---|
| 1.0 | 60 | 2A ⟷ 2I | 73.7 ± 0.5 | 1265 ± 53 | 2371 ± 237 | 1.87 ± 0.21 | |
| 2I ⟷ 2B | 91.8 ± 0.1 | 368 ± 37 | |||||
| Rescan to 95 °C | 1.0 | 60 | 2A ⟷ 2I | 49.4 ± 0.1 | 275 ± 4 | 93 ± 10 | |
| 2I ⟷ 2B | 67.0 ± 0.1 | 230 ± 23 | 275 ± 28 | ||||
| Rescan to 105 °C | 1.0 | 60 | 2A ⟷ 2I | 68.2 ± 0.1 | 263 ± 26 | 148 ± 15 | |
| 2I ⟷ 2B | 91.5 ± 0.1 | 650 ± 65 | 57 ± 6 |
| Conc. (mg/mL) | Scan Rate (K/h) | Model | TG (°C) | ∆HvH (kJ/mol) | ∆Hcal (kJ/mol) | Ratio a | |
|---|---|---|---|---|---|---|---|
| 1.0–2.0 | 60 | 2A ⟷ 2I | 70.4 ± 1.4 | 1160 ± 119 | 2349 ± 235 | 2.03 ± 0.28 | |
| 2I ⟷ 2B | 83.5 ± 2.7 | 373 ± 84 | |||||
| Rescan to 75 °C | 1.0 | 60 | 2A ⟷ 2B | 68.8 ± 0.1 | 945 ± 95 | 1870 ± 187 | |
| Rescan to 95 °C | 2.0 | 60 | 2A ⟷ 2I | 63.0 ± 0.1 | 427 ± 43 | 281 ± 28 | |
| 2I ⟷ 2B | 82.4 ± 0.1 | 405 ± 13 | |||||
| Rescan to 105 °C | 1.0 | 60 | 2A ⟷ 2I | 58.1 ± 0.1 | 241 ± 24 | 555 ± 56 | |
| 2I ⟷ 2B | 82.1 ± 0.1 | 227 ± 3 |
| Conc. (mg/mL) | Scan Rate (K/h) | Model | TG (°C) | ∆HvH (kJ/mol) | ∆Hcal (kJ/mol) | Ratio a | |
|---|---|---|---|---|---|---|---|
| 0.81–3.82 | 15 | A ⟷ I | 44.5 ± 0.5 | 677 ± 42 | 501 ± 72 | 0.33 ± 0.04 | |
| I ⟷ B | 48.4 ± 0.2 | 862 ± 105 | |||||
| 2A ⟷ 2B | 73.7 ± 0.4 | 1238 ± 76 | 1857 ± 979 | 1.5 ± 0.6 | |||
| 1.0–2.25 | 60 | A ⟷ I | 46.2 ± 1.3 | 472 ± 92 | 637 ± 41 | 0.57 ± 0.11 | |
| I ⟷ B | 50.6 ± 0.5 | 637 ± 41 | |||||
| 2A ⟷ 2B | 75.0 ± 0.1 | 1082 ± 33 | 2075 ± 168 | 1.92 ± 0.17 | |||
| Rescan to 95 °C | 1.0–2.25 | 60 | 2A ⟷ 2B | 68.9 ± 1.8 | 486 ± 149 | 143 ± 12 | |
| Rescan to 105 °C | 1.0–2.25 | 60 | 2A ⟷ 2B | 68.3 ± 0.5 | 430 ± 10 | 62 ± 6 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hoopes, J.T.; Heselpoth, R.D.; Schwarz, F.P.; Nelson, D.C. Thermal Characterization and Interaction of the Subunits from the Multimeric Bacteriophage Endolysin PlyC. Biology 2023, 12, 1277. https://doi.org/10.3390/biology12101277
Hoopes JT, Heselpoth RD, Schwarz FP, Nelson DC. Thermal Characterization and Interaction of the Subunits from the Multimeric Bacteriophage Endolysin PlyC. Biology. 2023; 12(10):1277. https://doi.org/10.3390/biology12101277
Chicago/Turabian StyleHoopes, J. Todd, Ryan D. Heselpoth, Frederick P. Schwarz, and Daniel C. Nelson. 2023. "Thermal Characterization and Interaction of the Subunits from the Multimeric Bacteriophage Endolysin PlyC" Biology 12, no. 10: 1277. https://doi.org/10.3390/biology12101277
APA StyleHoopes, J. T., Heselpoth, R. D., Schwarz, F. P., & Nelson, D. C. (2023). Thermal Characterization and Interaction of the Subunits from the Multimeric Bacteriophage Endolysin PlyC. Biology, 12(10), 1277. https://doi.org/10.3390/biology12101277