
Evolution of the Family Equidae, Subfamily Equinae, in North, Central and South America, Eurasia and Africa during the Plio-Pleistocene

 ,
,  , , ,
, , ,  ,
,  , ,
, ,  ,
,  ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
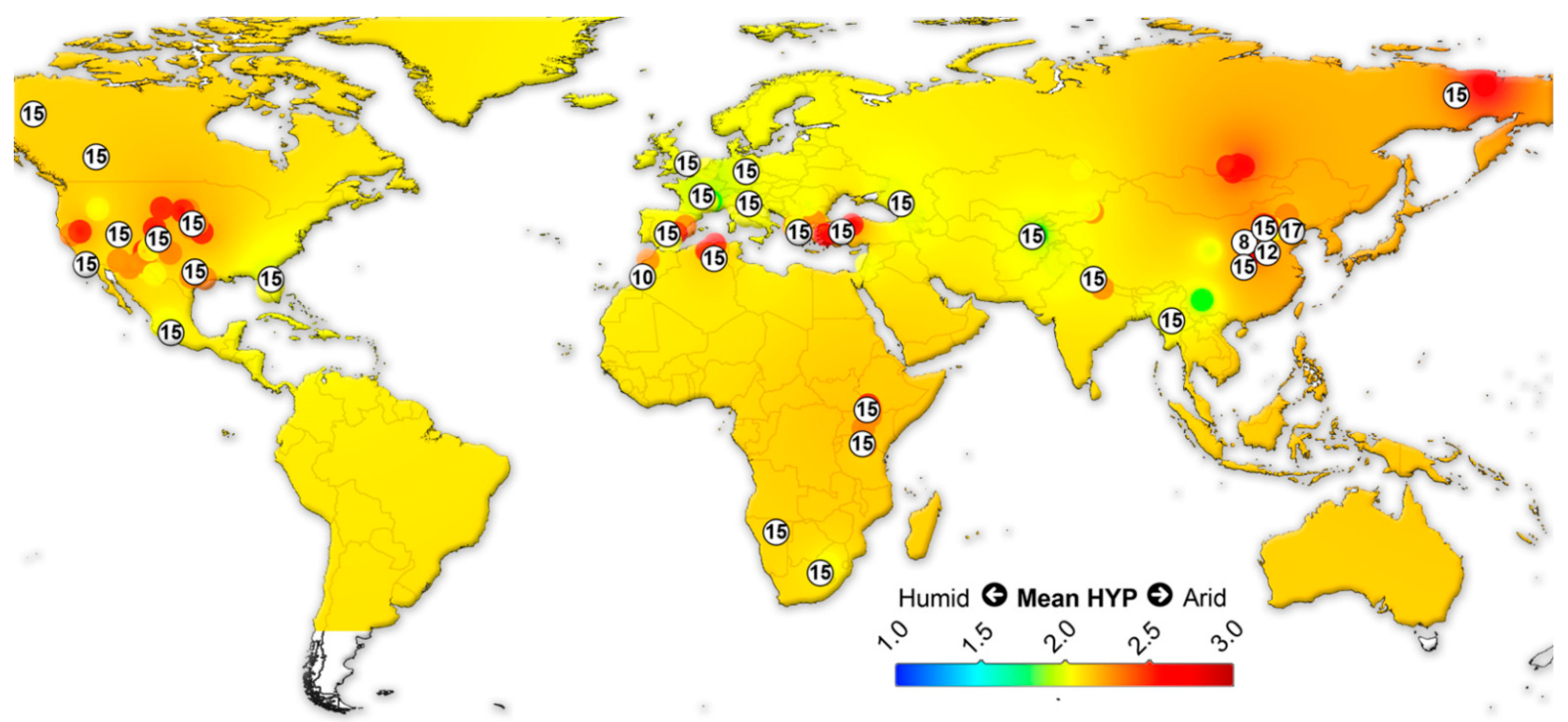{kind=link}
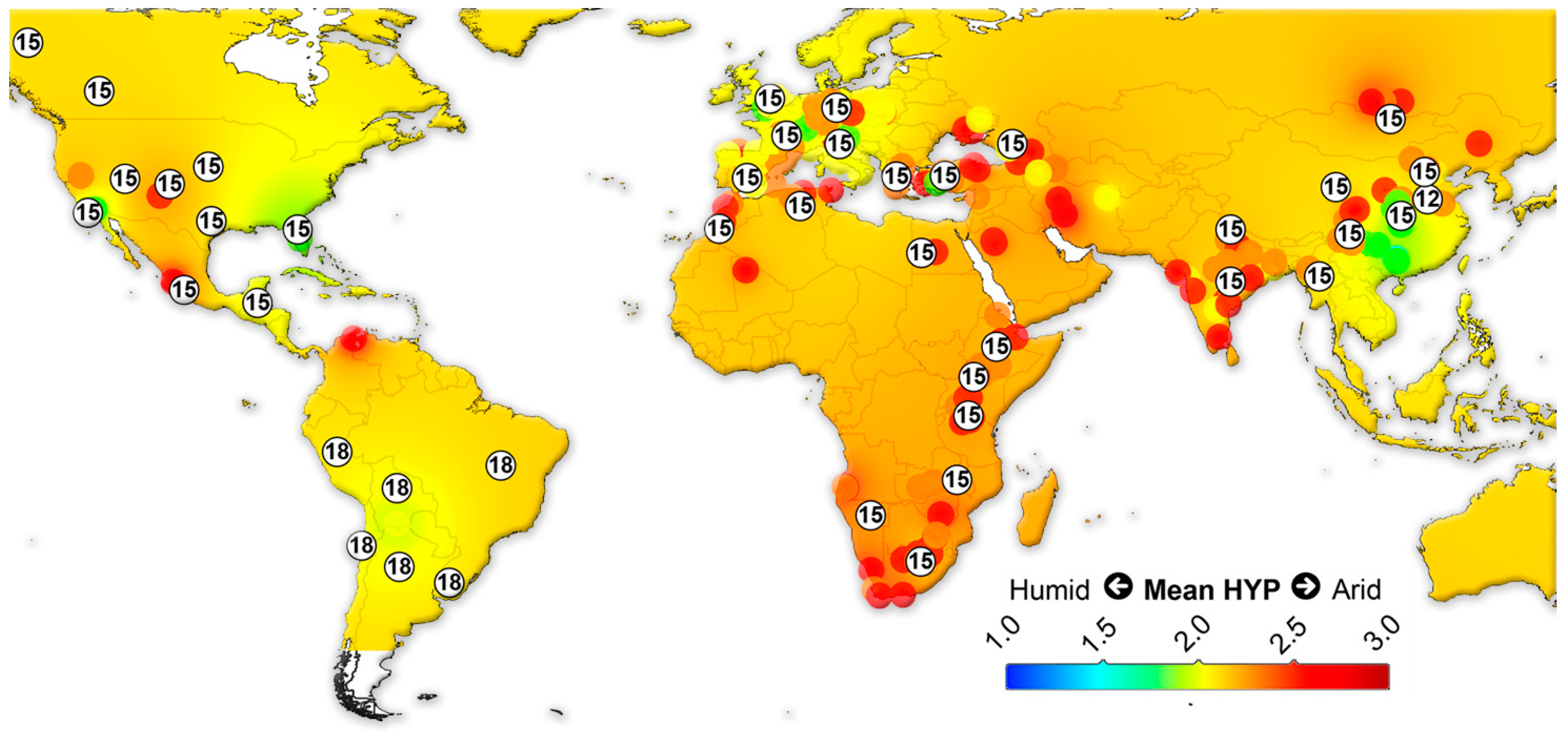{kind=link}
Abstract
Simple Summary
Abstract
1. Introduction
- (1)
- Provide a systematic revision of tridactyl and monodactyl horses from 5.3 Ma to 10 ka. These taxa are discussed in chronological order, from the oldest to the youngest occurrence in each region starting from North and Central America, South America, Eastern Asia (i.e., Central Asia, China, Mongolia and Russia), Indian Subcontinent, Europe and Africa, with their temporal ranges, geographic distributions, time of origins and extinctions. Part of this information is also reported in Table S1;
- (2)
- Integrate the distribution of the fossil record with paleoclimate and paleoecological data in order to provide new insights into the evolution of Equinae and their associated paleoenvironments;
- (3)
- Compare and discuss the latest morphological and genetic-based phylogenetic hypotheses on the emergence of the genus Equus. Recently, Barrón–Ortiz et al. [9] and Cirilli et al. [10] provided phylogenies of Equus, including fossil and extant species, with different resulting hypotheses. We compare and discuss the results of these two competing hypotheses here;
- (4)
- Provide a summary synthesis of the major patterns in the evolution, adaptation and extinction of Equidae 5.3 Ma–10 ka.
2. Materials and Methods
3. Systematics of the Equinae since 5.3 Ma in North, Central and South America
3.1. North and Central America
- 1. Calippus elachistus, Hulbert, 1988 [30] (Right Mandibular Fragment with m2–m3, UF342139). This species seems to be restricted to Florida (USA) from the Late Miocene to the Late Hemphillian. The occlusal dimensions of its cheeck teeth are much smaller than any other species of Calippus, except Ca. regulus, with slightly smaller occlusal dimensions in the early to middle wear stages and significantly smaller basal crown lengths than Ca. regulus.
- 2. Calippus hondurensis, Olson and McGrew, 1941 [31] (Partial Skull Containing Left P2–M3 and Right P2–4, WM 1769). This species has been reported in Puntarenas (Costa Rica), Gracias (Honduras), Mexico (Guanajuato, Hidalgo, Jalisco and Zacatecas) and in the USA (Florida). It may be distinguished by its small size and relatively small protocone.
- 3. Dinohippus leardi, Drescher, 1941 [32] (M1, CIT 2645). This species has been recorded from the Late Miocene in California (USA). The size of the molars is similar to that of Pliohippus nobilis or larger in unworn teeth.
- 4. Dinohippus spectans, Cope, 1880 [33] (Left M2 with Associated or Referred P2, AMNH 8183). This species has been recorded in Oregon, Nevada, California, Texas and Idaho (USA) dating to the Late Miocene, with molar teeth of larger size than those of any of the extinct American horses, except Equus excelsus, approximately equal to those of Hippidion principale.
- 5. Astrohippus ansae, Matthew and Stirton, 1930 [34] (Partial Left Maxilla with P2-M3, UC30225). This species is a Hemphilian–Blancan species recorded in Zacatecas (Mexico), New Mexico, Oklahoma and Texas (USA).
- 6. Astrohippus stockii, Lance, 1950 [35] (Palate with P2-M2, Front Portion of M3 on the Right Maxilla and P2-M2 on the Left One, CIT3576). This species has been recorded in Chihuahua, Guanajuato and Jalisco (Mexico) and Florida, New Mexico and Texas (USA), from the latest Hemphilian to Blancan NALMAs. Astrohippus stockii is smaller than A. ansae, but it possesses higher-crowned cheek teeth [35]. In recent phylogenetic analyses, A. stockii was recovered as the sister group to the clade composed of “Dinohippus” mexicanus plus Equus sensu lato [8,36] or the sister group to the clade composed of successive species of “Dinohippus” (i.e., “Dinohippus” leardi, “Dinohippus” interpolatus, Dinohippus leidyanus and “Dinohippus” mexicanus) plus Equus sensu lato [37].
- 7. Dinohippus interpolatus, Cope, 1893 [38] (First and Second Upper Molars, Plate XII, Figures 3 and 4). This is a late Hemphillian-Blancan species that has been recorded in Hidalgo and Zacatecas (Mexico) and in California, Kansas, New Mexico and Texas (USA).
- 8. Dinohippus leiydianus, Osborn, 1918 [3] (Skull, Jaws, Vertebrae, Fore and Hind Limbs, Considerable Portions of the Ribs and Other Parts of the Skeleton of One Individual, AMNH 17224). This species comes from the late Hemphillian–Blancan with records in Alberta (Canada) and in Arizona, California, Kansas, Nebraska and Oklahoma (USA).
- 9. Dinohippus mexicanus, Lance, 1950 [35] (Partial Left Maxilla with P2-M3 and Part of the Zygomatic Arch, LACM-CIT 3697). This species has been found in Chihuahua, Guanajuato, Jalisco, Hidalgo, Nayarit and Zacatecas (Mexico) and in California, Florida, New Mexico and Texas (USA) from Hemphillian to Blancan NALMAs. It is a medium-sized monodactyl equine horse.
- 10. Cormohipparion occidentale, Leidy, 1856 [39] (Four Left and One Right Upper Cheek Teeth, ANSP 11287). This species is a Hemphillian–Blancan species recorded in California, Florida, Nebraska, New Mexico and Oklahoma (USA). It is a large and hypsodont North American hipparion.
- 11. Nannippus aztecus, Mooser, 1968 [40] (Fragmented Right Maxillary with P3–M3, FO 873). This species has been recorded in Mexico (i.e., Chihuahua, Guanajuato and Jalisco) and in the USA (i.e., Alabama, Florida, Louisiana, Mississippi, Oklahoma and Texas) from the latest Hemphillian to Blancan NALMAs. It is a small-sized horse.
- 12. Nannippus lenticularis, Cope, 1893 [3] (Two Upper Cheek Teeth). This species has been recorded from Hemphillian to Blancan NALMAs in Alberta (Canada) and in Alabama, Kansas, Nebraska, Oklahoma and Texas (USA).
- 13. Nannippus peninsulatus, Cope, 1885 [41] (M2, AMNH8345). This is a Hemphillian–Blancan species with records in Guanajuato, Hidalgo, Jalisco and Michoacan (Mexico) and in Arizona, Florida, Kansas, Nebraska, New Mexico and Texas (USA). Nannippus peninsulatus was a highly cursorial equid that appears to have been functionally monodactyl [42,43]. It had an estimated body mass of 59.6 kg [44].
- 14. Neohipparion eurystyle, Cope, 1893 [38] (TMM 40289-1). This species has been recorded in Alberta (Canada); Guanajuato, Hidalgo, Jalisco and Zacatecas (Mexico); Alabama, California, Florida, Kansas, Nebraska, Oklahoma and Texas (USA) from the Hemphillian to Blancan NALMAs. It is a very hypsodont medium-sized hipparion.
- 15. Neohipparion leptode, Merriam, 1915 [45] (Lower Molar, UCMP 19414). This species is a Hemphillian–Blancan species recorded in California, Kansas, Nebraska, Nevada, Oklahoma and Oregon (USA). It is a large hipparion.
- 16. Hipparionini genus and Species Indeterminate. Hulbert and Harington [46] reported a remarkable specimen of a Hipparionini equid from the Canadian Arctic, which represents the northernmost fossil record of an equid reported to date. It was found in an Early Pliocene deposit (~3.5–4 Ma) from the Strathcona Fiord, Beaver Pond locality, Ellesmere Island, Canada [46]. The specimen consists of associated maxillae and premaxillae with the right dI1 and dP2–dP4 and the left dP1–dP4 of a young foal (approximately 6–10 months of age) [46]. It is a relatively large hipparionine equid (estimated adult tooth row length of 150 mm), with deciduous premolars that have low crowns; complex enamel plications; oval, isolated protocones; a facial region that shows a reduced preorbital fossa located posterior to the infraorbital foramen [46]. This combination of traits is not known in any contemporaneous North American hipparionines, but it is found in some Asiatic hipparionines, particularly Plesiohipparion, indicating possible affinities with this group and suggesting a previously unrecognized dispersal event from Asia into North America [8,46]. Alternatively, the Ellesmere Island hipparionine could represent a previously unknown autochthonous lineage of high-latitude North American hipparionines that potentially evolved from the mid-Miocene North American Cormohipparion [46].
- 17. Neohipparion gidley, Merriam, 1915 [45] (Left M3, UCMP 21382). This species is a Hemphillian species recorded in California and Oklahoma (USA). It is the largest of the North American hipparions.
- 18. Boreohippidion galushai, MacFadden and Skinner, 1979 [47] (Partial Skull with Well-Preserved Dentition, AMNH 100077). This is a late Hemphillian horse from Arizona (USA).
- 19. Cormohipparion emsliei, Hulbert, 1987 [48] (partial skull with most of the right maxilla including dP1, P2-M3; right and left premaxillae with I1–I3; edentulous fragment of the left maxilla with alveoli for dP1 and P2). UF 94700. All elements possibly belong to the same individual as they present similar stages of tooth wear and preservation. It is a species recorded in the latest Hemphillian to Blancan NALMAs in Alabama, Florida and Louisiana (USA). It is a medium-sized species of Cormohipparion.
- 20. Pseudohipparion simpsoni, Webb and Hulbert, 1986 [49] (Associated P3-M1, UF 12943). This is a latest Hemphillian species recorded in Florida, Kansas, Oklahoma and Texas (USA).
- 21. Pliohippus coalingensis, Merriam, 1914 [50] (UCMVP 21341). This species is a Pliocene horse from California (USA).
- 22. Nannippus beckensis, Dalquest and Donovan, 1973 [51] (Partial Skull with Right and Left P2–M3, TMM41452-1). This is a Blancan species from Texas (USA). This species is a medium-sized and moderately hypsodont hipparion.
- 23. Equus simplicidens, Cope, 1892 [52] (Left M1, TMM 40282-6). This species is interpreted to have been a medium- to large-sized equid with primitive dentition [53], recorded in Baja California (Mexico) and Arizona, California, Idaho, Kansas, Nebraska and Texas (USA) from Blancan to Irvingtonian. The species was initially based upon fragmented molars, with sizes comparable to E. occidentalis and E. caballus [52]. According to Skinner [54], E. simplicidens shows great similarities in the skull and dentition with the modern Equus grevyi, and the differences in the skull are small and expected in temporal and geographic separation. Gidley [55] described the Hagerman horses based upon characters common to all zebrine horses, with taxonomically significant differences from non-zebrines, but the characters used to distinguish it from other zebrines are of doubtful validity [56]. Comparing E. simplicidens with the East African Grevy’s zebra, E. grevyi, Skinner [54] included both in the subgenus Dolichohippus [57]. This proposal was questioned by Forsten and Eisenmann [57], as the cranial similarities found by Skinner [54] might be allometrically related to the large skull size [57]. Furthermore, Skinner [54] did not compare the basicranium, missing the comparison of Franck’s Index (i.e., the distance from the staphylioin to the hormion and from the hormion to the basion). The index was considered phylogenetically important, as the lengthening of the hormion to the basion distance seems to have led to a decrease in the index during Equus evolution, with a high index being related to a more primitive character than a lower derived index [57]. In Forsten and Eisenmann’s [57] analysis, both E. simplicidens and Pliohippus (Dinohippus), considered the generic ancestor of Equus, presented a high index, while E. grevyi and the other extant species presented lower indices. Following Matthew [4], Forsten and Eisenmann [57] also suggested E. simplicidens should be included in the subgenus Plesippus [4,58]. Equus simplicidens has long been considered the earliest common ancestor of Equus [57], but a recent analysis of the genus Equus suggested that Plesippus and Allohippus should be elevated to a generic rank, indicating Allohippus stenonis as the sister taxon to Equus and Plesippus simplicidens and P. idahoensis as the sister taxa to the Allohippus plus Equus clades [9]. On the other hand, recent cladistic analyses combined with morphological and morphometrical comparisons of skulls suggest E. simplicidens as the ancestor of Equus, not endorsing Plesippus and Allohippus at the genus or subgenus level [10].
- 24. Equus idahoensis, Merriam, 1918 [59] (Upper Left Premolar, UCMP 22348). This is a Blancan and early Irvingtonian species recorded in Arizona, California, Idaho and Nevada (USA). The type locality is Locality 3036C, in the beds of the Idaho locality, near Froman Ferry on the Snake River, 8 mi SW Caldwell, Idaho. According to Winans [56], none of the traits from the original description of E. idahoensis are unique to this species. However, large samples of specimens (e.g., Grandview, Idaho; 111 Ranch, Arizona) have been referred to as E. idahoensis, which have distinctive morphological features [9,60] that indicate that this is a potentially valid species. The cheek teeth are large and heavily cemented.
- 25. Equus enormis, Downs and Miller, 1994 [61] (The holotype, IVCM 32, is a partial skull and right and left mandibles, with the right distal humerus, right radius-ulna, MCIII, unciform, magnum, trapezoid and MCIII, phalanges 1, 2, and 3 of the manus; partial pelvis, right femur, MTIII with MTII and MTIV and phalanx 3 of the pes from Vallecito Creek, Anza-Borrego Desert State Park, San Diego County, California, USA). This species is known primarily from the late Blancan – Irvingtonian of California (USA). Equus enormis is a large-sized monodactyl horse with an estimated height at the withers of 1.5 m.
- 26. Equus cumminsii, Cope, 1893 [38] (Fragmentary Upper Molar, TMM 40287-14). This small species has been recorded in Kansas and Texas (USA) from Blancan to early Irvingtonian (NALMAs). Although it is poorly represented by fossils and the type of specimen is a single damaged tooth, some authors consider this species as an early ass based on dental morphology [60,62,63].
- 27. Equus calobatus, Troxell, 1915 [64] (Left MTIII, YPM 13470). This species is a large stilt-legged horse reported from the late Blancan to early Rancholabrean NALMAs, with records in Alberta (Canada); Aguascalientes (Mexico); Colorado, Kansas, Nebraska, New Mexico, Oklahoma and Texas (USA). The original discovery consisted of “unusually long and slender” limb bones [64] from Rock Creek, Texas, but no single holotype specimen was designated. Hibbard [65], therefore, selected YPM 13460 as the lectotype. Because the lectotype and cotypes are limb bones with no distinctive characters other than the large size and relative slenderness of the metapodials, there are few morphological criteria available for evaluating this species. Multiple studies [29,56] have synonymized E. calobatus with Equus (or Haringtonhippus) francisci, but other studies consider it a valid species [66,67].
- 28. Equus scotti, Gidley, 1900 [68] (Associated Skeleton with Skull, Mandible, Complete Feet and Forelimb Bones, One Complete Foot and Hindlimb and All the Cervical, Several Dorsal and Lumbar Vertebrae, AMNH 10606). This species is recorded from the late Blancan to Rancholabrean NALMAs in Alberta, Ontario, Saskatchewan and Yukon (Canada) and in California, Florida, Idaho, Kansas, Nebraska, New Mexico, Oklahoma and Texas (USA). Winans [54] also interpreted E. scotti to be on average slightly smaller than E. simplicidens, but the measurements provided in that study actually indicate the opposite, and subsequent review confirms that E. scotti was of a larger form than E. simplicidens.
- 29. Equus stenonis anguinus, Azzaroli and Voorhies, 1993 [69] (Complete Skull and Jaw, USNM 23903). This is a late Blancan species recorded in Arizona and Idaho (USA). It is described as similar to E. stenonis from the Early Pleistocene of Italy, with skull dimensions falling within the size range of this latter species [69]; however, the limb bones are, on average, more elongated. Equus stenonis anguinus possesses a preorbital pit and a deep narial notch as do the European E. stenonis samples.
- 30. Equus conversidens, Owen, 1869 [70] (Fragmentary Right and Left Maxilla with All Cheek Teeth, IGM4008, Old Catalog Number MNM-403). This is a widespread species reported to have ranged from the Irvingtonian to Rancholabrean with a geographic distribution encompassing North and Central America: Alberta (Canada); Aguacaliente de Cartago (Costa Rica); Apopa Municipality (El Salvador); Aguascalientes, Chiapas, Hidalgo, Jalisco, Michoacán, Nuevo Leon, Puebla, Oaxaca, San Luis Potosi, Sonora, Estado de Mexico, Tlaxcala, Yucatán and Zacatecas (Mexico); Azuero Peninsula and El Hatillo (Panama); Arizona, California, Florida, Kansas, Nebraska, New Mexico, Oklahoma, Texas and Wyoming (USA). The holotype specimen from the Tepeyac Mountain was described by Owen [70] based upon photos [71]. Owen considered the species to be almost identical to E. curvidens (South American E. neogeus) but with cheek tooth rows converging towards their anterior ends. Cope [72] interpreted the anterior convergence of the cheek tooth rows to be an artifact of the restoration compounded by the photography and assigned the specimen to E. tau, albeit without any stated justification. Gidley [69] interpreted the two sides of the maxilla to be from different individuals, since they were found separately and with missing broken edges. Hibbard [65] provided a reconsideration of the specimen and confirmed that Cope [68] was correct regarding the distortion of the palate and that Gidley [73] was incorrect regarding the two sides deriving from different individuals. Azzaroli [74] described a fragmentary skull (LACM 308/123900) from Barranca del Muerto near Tequixquiac, Mexico, in which the “two tooth rows converge rostrally, giving evidence that the palate of the holotype (of E. conversidens) was correctly mounted and that Owen’s name is after all appropriate”.
- 31. Equus lambei, Hay 1915 [75] (~200 ka–~10 ka) (nearly complete skull from a female, USNM8426, collected from Gold Run Creek in the Klondike Region, Yukon Territory, Canada). This species inhabited the steppe–tundra grasslands of Beringia, with remains having been recovered from Siberia, Alaska, and the Yukon (extending slightly into the adjacent Northwest Territories). Recent genomic evidence suggests that E. lambei and E. ferus may represent a single species [29,76,77,78], although further research is required before this phylogeny can be resolved (see Supplementary Materials, Section 3.4 supplementary text for a discussion on E. ferus in North America).
- 32. Equus (or Haringtonhippus) francisci, Hay, 1915 [76] (Complete Cranium, Mandible and MTIII, TMM 34–2518). This species has been recorded from Irvingtonian to Rancholabrean localities in the Yukon (Canada); Aguascalientes, Estado de Mexico, Jalisco, Puebla, Sonora, and Zacatecas (Mexico); Alaska, Arizona, Florida, Kansas, Nebraska, New Mexico, Texas and Wyoming (USA). It is the oldest name assigned to the stilt-legged group and was first described as being similar to E. tau but with different P3-M1 proportions, which is expected in teeth at different stages of wear and, therefore, probably not a significant taxonomical difference (56). Eisenmann et al. [67] reassigned E. francisci to the genus Amerhippus, as Amerhippus francisci. Heintzman et al. [29] assigned stilt-legged, non-caballine specimens from Gypsum Cave, Natural Trap Cave, the Yukon and elsewhere to their new genus Haringtonhippus under the species Ha. francisci, based upon complete mitochondrial and partial nuclear genomes as well as morphological data and a crown group definition of the genus Equus. Barrón-Ortiz et al. [9] considered Haringtonhippus to be a synonym of Equus and regarded both E. francisci and E. conversidens as distinct taxa based upon morphological criteria.
- 33. Equus fraternus, Leidy, 1860 [79] (Upper Left P2, AMNH 9200). This species has been recorded in Alberta (Canada) and in Florida, Illinois, Mississippi, Nebraska, Pennsylvania, South Carolina and Texas (USA) from Irvintonian to Rancholabrean. Winans [56] considered the dental characteristics used to identify the species to vary with wear and that the specimens used in its diagnosis represented more than one individual, making it uncertain whether to attribute it to one species. Azzaroli [74,80] referred several complete skulls, mandibles and other bones from the southeastern USA to this species.
- 34. Equus pseudaltidens, Hulbert, 1995 [67] (right maxillary with worn DP2, DP3, DP4, M1 and M2 and unerupted P2, P3 and P4 (BEG 31186-35); right and left mandibles with worn i1, di2, dp2, dp3, dp4, m1 and m2 and unerupted p2, p3 and p4 (BEG 31186-36); cranium lacking occiput (BEG 31186-37); right third metacarpal (BEG 31186-3); right and left femora (BEG 31186-2, 34); right and left tibiae (BEG 31186-1, 10); right and left third metatarsals (BEG 31186-4, 7); first phalanx (BEG 31186-24), all thought to belong to the same individual and estimated to have been approximately 3 years old [81]). This species was originally described as Onager altidens by Quinn [81]. The use of Onager instead of Hemionus by Quinn [81] was invalid [67]. Referral of this species to the genus Equus makes it a homonym of Equus altidens, von Reichenau, 1915 [53,67,82]. Therefore, Hulbert [67] proposed the replacement name E. pseudaltidens for E. altidens (Quinn). Also referred to this species are a pair of maxillae (BEG 31186-23) and a right mandible (BEG 31186-22) of an animal approximately 1 year old, and 24 deciduous and permanent upper and lower teeth recovered from the type locality [81]. Equus pseudaltidens is known from the Irvingtonian–Rancholabrean and it has been reported from the Gulf Coastal Plain of Texas [67,81] and possibly from Coleman, Florida [67]. It is a stilt-legged equid with metapodial dimensions that are similar to extant hemionines [67,76]. Compared to other stilt-legged equids discussed here, Equus pseudaltidens is smaller than E. calobatus but larger than both E. francisci and E. cedralensis [67,76,83]. Kurtén and Anderson [84] synonymized E. (Hemionus) pseudaltidens with Equus (Hemionus) hemionus. Winans [85] assigned it to her E. francisci species group. Hulbert [67] considered that E. pseudaltidens and E. francisci were distinct species and hypothesized that they are sister taxa. Azzaroli [74] synonymized E. pseudaltidens (as Onager altidens) with E. semiplicatus. Eisenmann et al. [67] considered E. pseudaltidens distinct from E. semiplicatus and E. francisci and assigned it along with the latter species to Amerhippus. Heintzman et al. [29] considered E. altidens (=E. pseudaltidens) a junior synonym of Haringtonhippus francisci.
- 35. Equus verae, Sher, 1971 [86] (Holotype Mandible with a Full Row of Teeth, GIN 835-123/21, River Bolshaja Chukochya Exp. 21, Kolyma Lowland, Northeast Yakutia). This species is a large-bodied, stout-legged Early Pleistocene (Olyorian)–Rancholabrean species recorded in Northeastern Siberia (Russia) and the Yukon (Canada). E. verae is much larger than the E. stenonis species and similarities in the teeth and the size of limb bones with E. suessenbornensis, suggesting a subspecies position for E. verae as well as for E. coliemensis (see below). Eisenmann [87,88] suggested that E. verae may belong to the subgenus Sussemionus, but this has not been substantiated by other authors.
- 36. Equus occidentalis, Leidy, 1865 [89] (Lectotype Left P3, VPM 9129). It is a Rancholabrean NALMA horse with records in Mexico (i.e., Baja California and Sonora) and the USA (i.e., Arizona, California, Nevada, New Mexico and Oregon). Leidy [89] named E. occidentalis from two upper premolars and one lower molar from two widely separated geographic localities but did not designate a holotype. Gidley [73] selected a left P3 from Tuolumne County, California, as the lectotype. Merriam [90] referred to E. occidentalis thousands of bones of a large and stout-limbed equid recovered from Rancho La Brea. Savage [91] and Miller [92] believed that the equid from Rancho La Brea, identified as E. occidentalis, did not conform to the lectotype designated by Gidley [73], but neither of these authors proposed a new name. Azzaroli [74] decided to retain the name E. occidentalis sensu Merriam [90] and selected the skull figured by Merriam [90] as his lectotype. However, since Gidley [73] had already designated a lectotype for the species, according to ICZN Article 74, no subsequent lectotype designations can be made. Brown et al. [93] concluded that some of Leidy’s original fossils of E. occidentalis (exclusive of the Tuolumne County tooth) most likely come from the McKittrick asphalt deposits; this locality was not named by Leidy [89], because the town of McKittrick, California, was not named until 1900. These authors also confirmed that many specimens from the original type series of E. occidentalis closely resemble the large Pleistocene horses from McKittrick and Rancho La Brea [93]. Barrón-Ortiz et al. [94] recognized the presence of this species outside of the North American Western Interior during the Late Pleistocene. Barrón-Ortiz et al. [9] recognized it as a valid species closely related to E. neogeus.
- 37. Equus cedralensis, Alberdi et al., 2014 [83] (fragment of a mandibular ramus formed by two specimens: one p2-m3 right row (DP-2675 I-2 15) and a second fragment of the symphysis with the anterior dentition (DP-2674 I-2 8), articulated together from Rancho La Amapola, Cedral, San Luis Potosí, Mexico, and stored at the Paleontological Collection (DP-INAH) of the Laboratorio de Arqueozoología “M. en C. Ticul Álvarez Solórzano” Subdirección de Laboratorios y Apoyo Académico, INAH in Mexico City). This species is primarily known from the Rancholabrean of Mexico (i.e., Aguascalientes, Chihuahua, Estado de Mexico, Michoacán, Puebla and San Luis Potosí). Equus cedralensis is an equid with a small body mass (estimated mean mass of 138 kg) [83,95]. Equus cedralensis was diagnosed as stout-legged, but a recent analysis placed it within stilt-legged horses and with its dental morphology being similar to Ha. francisci. Jimenez-Hidalgo and Diaz-Sibaja [96] considered it a junior synonym of Ha. francisci. Equus cedralensis differs from the holotype of Ha. francisci, as it is smaller in size and the lower first and second incisors possess enamel cups.
- 38. Equus mexicanus, Hibbard, 1955 [65] (cranium lacking the LM3 (No. 48 (HV-3)) from Tajo de Tequixquiac, Estado de México, Mexico, and stored at the Museo Nacional de Historia Natural; the specimen is cataloged as IGM4009). This species is known from the Rancholabrean of Mexico (i.e., Aguascalientes, Chiapas, Estado de Mexico, Jalisco, Michoacán, Oaxaca, Puebla, San Luis Potosi and Zacatecas) and the USA (i.e., California, Oregon and Texas). Equus mexicanus is a large body sized species (estimated mean mass of 458 kg) [83,95]. Winans [85] placed E. mexicanus in her Equus laurentius species group, but as with other species groups in this study, this was not a strict synonymy. Azzaroli [74] recognized E. mexicanus as a valid taxon, noting that previous investigations had proposed synonymy with and tentatively identified as E. pacificus but rejecting this, since the latter species was initially based upon a single tooth. Barrón-Ortiz [97] assigned specimens identified as E. mexicanus to E. ferus scotti. Barrón-Ortiz et al. [94] assigned specimens identified as E. mexicanus to E. ferus. Barrón-Ortiz et al. [9] recognized E. mexicanus as a valid taxon distinct from E. ferus.
3.2. South America
- 1. Equus neogeus, Lund, 1840 [101] (MTIII, 866 Zoologisk Museum). This is a Middle–Late Pleistocene species (Ensenandan and Lujanian SALMA) and the only representative of the genus in the South American continent [9,98,99,100]. Most records are from the Late Pleistocene, but its earliest appearance is recorded in the Middle Pleistocene in Tarija, Bolivia, dated at approximately 1.0–0.8 Ma [102,103]. It has a wide geographic range distribution, encompassing all of South America, except for the Amazon basin and latitudes below 40° [100]. The species probably became extinct sometime during the Late Pleistocene–Holocene transition as suggested by the youngest direct radiocarbon date of 11,700 BP (Río Quequén Salado, Argentina [104]).
- 2. Hippidion saldiasi, Roth, 1899 [105] (p2, Museo Nacional de La Plata). This is a Late Pleistocene species, dated between 12,000 and 10,000 years BP, mostly known from Argentinian and Chilean Patagonia, with records in Central Chile and the Atacama Desert [98,106]. The last records for the species were radiocarbon dated between 12,110 and 9870 BP in Southern Patagonia (Cerro Bombero, Argentina [107]) and Cueva Lago Sofía, Chile [108].
- 3. Hippidion principale, Lund, 1846 [109] (M2, Peter W. Lund Collection, ZMK). This is a Late Pleistocene species (Lujanian SALMA), with records in Argentina, Bolivia, Brazil and Uruguay [98]. This species represents the largest Hippidion. There are few radiocarbon-dated records for this species, with the youngest situated at approximately 16,130 BP (Arroyo La Carolina, Argentina) [104]; however, remains found in archaeological contexts dating close to 13,200 BP in Tagua, Central Chile [110], suggest a later presence for this taxon.
- 4. Hippidion devillei, Gervais, 1855 [111] (P2–P3 Row and Fragmented Astragalus, IPMNHN). This species has been reported in Uquia (Argentina, Late Pliocene–Early Pleistocene), Tarija (Bolivia) and Buenos Aires (Argentina) from the Middle Pleistocene (Ensenadan SALMA) to the Late Pleistocene (Lujanian SALMA) and in Brazil [98]. This taxon has been directly radiocarbon dated only from cave contexts in the high Andes of Peru, with the youngest record of 12,860 BP [112,113].
4. Systematics of the Equinae since 5.3 Ma in Eurasia and Africa
4.1. Eastern and Central Asia (China, Mongolia, Russia, Uzbekistan, Kazakhstan and Tajikistan)
- 1. Plesiohipparion houfenense, Teilhard de Chardin and Young, 1931 [116] (MN13–MN15; 6–3.55Ma). The lectotype RV 31031 includes the right p3–m3 from Jingle, Shanxi. The earliest P. houfenense first occurs in the Late Miocene Khunuk Formation, Kholobolchi Nor, Mongolia [8,117,118,119] and the Late Miocene/Early Pliocene Goazhuan Formation of the Yushe Basin (5.8–4.2 Ma) [8,120]. It also occurs into the Pliocene of China.
- 2. Proboscidipparion pater, Matsumoto, 1927 [121] (MN14–MN15; 5–3.5 Ma). The lectotype THP14321 is a skull with a mandible estimated to be 4–3.55 Ma [122]. This species is reported from the Yushe Basin (China), and it may be the original source for the evolution of the Pliocene European species Proboscidipparion crassum and Proboscidipparion heintzi.
- 3. Plesiohipparion huangheense, Qiu et al., 1987 [122] (MN15; 5.0–3.55). The lectotype THP 10097 is a lower jaw fragment, including the cheek teeth, from the Yushe Basin [122]. It is a Chinese species reported from Inner Mongolia at 3.9 Ma [7,119,122] and more broadly from the MN15 of China and India [8]. Ultimately, Pl. aff. huangheense has been reported from the Early Pleistocene in Gulyazi, Turkey [123].
- 6. Plesiohipparion shanxiense, Bernor et al., 2015 [124] (MNQ17; 2.5–1.8 Ma). The holotype is F:AM111820, a complete skull with the mandible [124]. This species, previously recognized as Plesiohipparion cf. P. houfenense [117], is the largest and, at the same, the time youngest member of the genus in Eastern Eurasia. It is believed to be 2.0 Ma in age [124]. It may represent the last evolutionary stage of the genus Plesiohipparion in China. The absence of the POF suggest an evolutionary relationship with Pl. houfenense.
- 7. Proboscidipparion sinense, Sefve, 1927 [125] (MN17-MQ1; 2.5–1.0 Ma). The holotype is PMU M3925, a complete cranium from Henan Province, China. Proboscidipparion sinense occurs later in the record and is approximately one-seventh larger than P. pater. Proboscidipparion sinense is the latest occurring hipparion in China extending its range up to 1.0 Ma [126].
- 8. Equus eisenmannae, Qiu et al., 2004 [127] (2.55–1.86 Ma). The holotype is IVPP V13552, a complete cranium with and the mandible from Longdan [127]. It is a large-sized horse, mostly known from the Early Pleistocene locality of Longdan (China), similar in size to E. livenzovensis (see below), and the primitive features of the cranial morphology suggest a close evolutionary relationship with E. simplicidens [10,13]. At the present time, its evolutionary linkage with other Chinese Equus is not known.
- 9. Equus sanmeniensis, Teilhard de Chardin and Piveteau, 1930 [128] (2.5–0.8 Ma). The lectotype is NIH 002 (Paris), a complete cranium with the mandible from Nihewan, Hebei Province. It is a large-sized, Early Pleistocene species from North and northwest China, Siberia (Aldan River, Bajakal Lake area), Kazakhstan and Tajikistan [114,129]. Sun and Deng [114] suggested a morphological similarity between E. sanmeniensis and E. simplicidens, although diversified from E. stenonis. This evolutionary hypothesis has also been supported by Cirilli et al. [10,13] through morphometric studies on crania. Equus sanmeniensis has been reported from the Early to Middle Pleistocene [114,130].
- 10. Equus huanghoensis, Chow and Liu, 1959 [131] (2.5–1.7 Ma). The holotype is IVPP V2385–2389, with three upper premolars and two molars from Huanghe, Shanxi (Sun and Deng, 2019). It is a large-sized, Early Pleistocene species from the localities of Nihewan (Hebei), Linyi (Shanxi), Sanmenxia Pinglu (Shanxi), Xunyi (Shaanxi) and Nanjing (Jiangsu). Sun and Deng [114] supported the hypothesis provided by Deng and Xue [130] that E. huanghoensis is a stenonid horse, considering the Nihewan sample as one of the Equus species with the largest palatal length with E. eisenmannae. The morphometric analyses of Cirilli et al. [10,13] show a primitive morphology of the cranium, similar to E. simplicidens and distinct from E. stenonis. Ao et al. [132] indicated the age of 1.7 Ma as the youngest record of this species in China.
- 11. Equus yunnanensis, Colbert, 1940 [133] (2.5–0.01 Ma). The lectotype is IVPP V 4250.1, an almost complete but deformed cranium. It is a medium-sized, Early Pleistocene species from the Chinese localities of Yuanmou (Yunnan), Liucheng (Guangxi), Jianshi and Enshi (Hubei), Hanzhong (Shaanxi) and Huili (Sichuan) and from Irrawaddy in Myanmar. The species was initially described by Colbert [133] on isolated cheek teeth, while better knowledge of this species came with the new discoveries from the Yuanmou locality. Deng and Xue [130] proposed a close evolutionary relationship with E. wangi, whereas Sun and Deng [114] suggested a close evolutionary relationship with E. teilhardi, suggesting that these species are distinct from all other Chinese stenonid horses [114].
- 12. Equus teilhardi, Eisenmann, 1975 [134] (2.0–1.0 Ma). The holotype is NIH001, an incomplete mandible. It is a medium-sized, Early Pleistocene species from northwestern and North China. Sun et al. [135] proposed a close evolutionary relationship between E. teilhardi and E. yunnanensis, later supported by the cladistic analysis of Sun and Deng [114].
- 13. Equus qyingyangensis, Deng and Xue, 1999 [130] (2.1–1.2 Ma). The holotype is NWUV 1128, an incomplete cranium. It is a medium-sized, Early Pleistocene species from northwestern and North China. Eisenmann and Deng [136] recognized some close anatomical features between E. simplicidens and E. qyingyangensis, suggesting a close evolutionary relationship between these two species. The latest phylogenetic results of Sun and Deng [114] and Cirilli et al. [10] support this last hypothesis. A new E. qyingyangensis sample has recently been described from Jinyuan Cave [115], with an FAD of 2.1 Ma.
- 14. Equus wangi, Deng and Xue, 1999 [130] (2.0–ca. 1.0 Ma). The holotype is NWUV 1170, a complete upper and lower cheek teeth rows from Gansu Province, Early Pleistocene [130]. Sun and Deng [114] reported a large size for E. wangi, similar to E. eisenmanne, E. sanmeniensis and E. huanghoensis. The phylogenetic position of E. wangi is not well defined, although Sun and Deng [114] highlighted a possible closer relationship with E. eisenmannae than any other stenonine Equus.
- 15. Equus pamirensis, Sharapov, 1986 [137] (Early Pleistocene). The holotype is IZIP 1-438 (Institute of Zoology and Parasitology, Uzbekistan), a complete upper tooth row from Kuruksai [137], approximately 2 Ma (MN17), possibly earlier. It is a large species of stenonine horse described from the Kuruksai, 18 km NE of Baldzuan, Tajikistan, in the Kuruksai River valley in the Afghan–Tajik Depression [138]. The site has been correlated to the middle Villafranchian. The taxonomy of this horse is contentious [138], with it being referred to variously as Equus (Allohippus) aff. sivalensis [137] and E. stenonis bactrianus [139]. We currently regard this as a distinct species.
- 16. Central Asian Small Equus sp. from Kuruksai [138] (Early Pleistocene). This is a small species of horse that co-occurs with the larger E. pamirensis at Kuruksai. Metrically, the metapodials fall within the range of variation of E. stehlini [138]. Similar small horse remains have also been found in the Early–Middle Pleistocene Lakhuti 1 locality in the Afghan–Tajik Depression [138].
- 17. Equus (Hemionus) nalaikhaensis, Kuznetsova and Zhegallo, 1996 [140]. This species was found in the late Early Pleistocene and early Middle Pleistocene (approximately 1 my, Jaramillo paleomagnetic episode) in Mongolia. The lectotype PIN 3747/500 is represented by the incomplete skull of an old male from Nalajkha [141].
- 18. Equus coliemensis, Lazarev, 1980 [142]. The holotype is a skull with very worn teeth (col. IA 1741). The type locality is the river Bolshaja Chukochya, Kolyma lowland, northeast Yakutia, Siberia, Russia. It is reported from the late Early Pleistocene in northeastern Siberia (Russia). Recently, Eisenmann [87] included E. coliemensis in the subgenus Sussemonius.
- 19. Equus lenensis, Rusanov, 1968 [143]. The holotype skull comes from the Lena River delta (GIN Yakutia col. 33). The type locality is the river Bolshaja Chukochya, Kolyma lowland, northeast Yakutia, Siberia (Russia). It is reported from the Middle Pleistocene from northeastern Siberia (Russia). Lazarev [144] considered E. lenensis to be close to the North American E. lambei, even larger and more heavily built than the latter. It is also known from the Middle Pleistocene in Yakutia Equus orientalis (Equus caballus/ferus orientalis) and Equus nordostensis (Equus ferus/caballus nordostensis) [143,144,145]. Equus nordostensis is characterized by a large size based on the skull, with low plication of the “marks” of the upper teeth and a long protocone [144]. According to Kuzmina [129], it is junior synonym of E. mosbachensis. Equus orientalis has a large skull and a long snout with an elongated teeth row, flat protocone and rare plication of the upper teeth [144].
- 20. Equus beijingensis, Liu, 1963 [146] (late Middle Pleistocene). The holotype is V2573-2574, a palate and jaw from Zhoukoudian, China [146,147]. It is a Chinese caballine horse recovered mostly form locality 21 of Zhoukoudian. Unfortunately, the species is not well represented, although Forsten [147] indicated a similar size with E. sanmeniensis. Liu [146] also indicated E. sanmeniensis as the possible ancestor for E. beijingensis; nevertheless, this hypothesis was discarded by Fortsen [147] and Deng and Xue [130], who identified E. beijingensis as a caballine horse, characterized by a U-shaped linguaflexid. Its evolutionary position is not well defined, although Forsten [147] and Deng and Xue [130] proposed that E. beijingensis is a relative of European E. ferus (their E. mosbachensis) or of a North Americam caballine horse [130].
- 21. Equus valeriani, Gromova, 1946 [148] (Late Middle–Late Pleistocene). The hypodigm includes upper and lower cheek teeth, figured in Eisenmann et al. [149]. It is an enigmatic taxon, described by Gromova [148], from Samarkand, Uzbekistan. According to Gromova [148] and Eisenmann et al. [149], it shows a stenonine metaconid-metastylid in the lower cheek teeth but with a long protocone in the upper cheek teeth. Its possible occurrence has also been proposed in Syrie (Kéberien Géométrique d’Umm el Tlel), although this identification still remains uncertain [149].
- 22. Equus dalianensis, Zhow et al., 1985 [150] (Late Pleistocene). The holotype is V821966, an incomplete mandible preserving two lower cheek teeth rows. It is a Chinese caballine horse described from Gulongshan Cave, Liaoning. Forsten [147] demonstrated close morphological and morphometrical similarities with E. ferus gemanicus, E. ferus orientalis and E. ferus chosaricus, suggesting that they all represent individual populations of a single widespread species, E. ferus. Deng and Xue [130] suggested a common origin for E. dalianensis and E. przewalskii, with no ancestor-descendant relationships between them. Nevertheless, a recent genomic analysis by Yuan et al. [151] revealed that E. dalianensis is a separate clade of caballine horses, distinct from E. przewalskii.
- 23. Equus ovodovi, Eisenmann and Vasiliev, 2011 [152] (0.04–0.01 Ma). The holotype is IAES 21, a fragmentary palate from Proskuriakova Cave [152]. It was described in the Late Pleistocene site of Proskuriakova Cave (Khakassia, southwestern Siberia, Russia). It was first considered a species related to E. hydruntinus and modern hemiones, although the genomic analyses by Orlando et al. [153] suggest a relationship with wild asses, representing a new separated fossil clade with no extant relatives [150]. For this reason, Orlando et al. [153] and Eisenmann and Vasiliev [152] included this species in the subgenus Sussemionus. Molecular studies suggest that E. ovodovi is a sister to extant zebras and is nested within the clade that includes both extant zebra and asses [154]. Equus ovodovi has been recognized in the Late Pleistocene of southern and eastern Russia and more recently in China [152,154,155].
- 24. Equus hemionus, Pallas, 1774 [156] (0.0 Ma). Pallas [156] did not refer to any holotype or lectotype but gave a detailed description of the anatomical features of the species, associated with an illustration of an animal located near Lake Torej-Nur, Transbaikal area [156] (V.19, pp. 394–417, Pl. VII). Equus hemionus is known as the Asiatic wild ass, distributed in China, India, Iran, Mongolia and Turkmenistan. Historically, it has also been reported in Afghanistan, Armenia, Azerbaijan, Georgia, Iraq, Jordan, Kuwait, Kyrgyzstan, Russia, Saudi Arabia, Syria, Tajikistan, Turkey and Ukraine. Four different subspecies are identified, mostly describing the present areal distribution: E. hemionus hemionus (Mongolia), E. hemionus khur (India), E. hemionus kulan (Turkmenistan) and E. hemionus onager (Iran). Another extinct subspecies was recognized in Syria, E. hemionus hemippus. The fossil record is not well studied, but crania and mandibles of the species have been reported from Narmada Valley in Central India [157], the Indian state of Gujarat [158], and the Son Valley in Northern India [159]. This species is distinguished from E. namadicus by its smaller size and smaller protocones on the premolars. Radiocarbon dates from these deposits suggest that this species entered South Asia during the last glacial period, most likely from West Asia [160,161]. The most recent genetic analyses suggest that the onager and kiang populations diverged evolutionarily ca. 0.4–0.2 Ma [28].
- 25. Equus kiang, Moorcroft, 1841 [162] (0.0 Ma). Moorcroft [162] did not designate any holotype or lectotype but provided a general description of the species [162]. Equus kiang is known as the Tibetan ass, with a distribution in China, Pakistan, India, Nepal and, possibly, Bhutan. Three subspecies have been identified, E. kiang kiang, E. kiang holdereri and E. kiang polyodont, with different authors pointing out their subspecific statuses [162,163,164,165]. At the present time, no paleontological information is available for E. kiang. Jonsson et al. [28] distinguished E. kiang from E. hemionus as a valid taxon. The most recent morphological cladistic analysis found E. kiang and E. hemionus to be stenonine horses [10].
- 26. Equus przewalskii, Poliakov, 1881 [166] (~0.1–0.0 Ma). The holotype, a skull (number 512) and skin (number 1523), are found in the collection of the Laboratory of Evolutionary Morphology, Moscow (museum exposition), originally obtained by N. M. Polyakov in Central Asia, southern Dzhungaria, in 1878 [129]. For a complete description of the species see Groves [167] and Grubb and Groves [168]. It is an extant species of caballine horse that is found in small geographic areas of Central Asia, although, historically, it once ranged from Eastern Europe to eastern Russia [169]. In China, E. przewalskii is common in the Late Pleistocene (~0.1–0.012 Ma) sites in the northern and central regions of the country, but it is absent in the Holocene, except in northwestern China [170]. It shows close morphological similarities with Equus ferus (see below), and both are members of caballine horses [10,28,153]. While genomic analyses have shown that the Przewalski’s horses are the descendants of the first domesticated horses from the Botai culture in Central Asia (Kazakhstan) around 5.5 ka [171,172], subsequent morphological studies have shown that Botai horses are not domestic horses but harvested wild Prezwalski’s horses [173].
4.2. Indian Subcontinent
- 1. “Hippotherium” antelopinum, Falconer and Cautley, 1849 [179] (3.6–2.6 Ma). The lectotype is NHMUK PV M.2647, a subadult right maxilla fragment with P2-M3. It is a species of hipparionine horse from the late Pliocene age deposits between the rivers Yamuna and Sutlej in India. This taxon has been the subject of much nomenclatural confusion. Lydekker named the lectotype and along with the hypodigm, placed it within the genus Hipparion. Later authors [180,181,182] have referred Miocene hipparionine material collected on the Potwar Plateau in Pakistan to this táxon and reassigned the species to the genus Cremohipparion. However, given that the hypodigm of “Hippotherium” antelopinum comes from the Late Pliocene and does not preserve any apomorphies of Cremohipparion; we refer to the late Pliocene specimens as “Hippotherium” antelopinum, separate from the Potwar Plateau specimens from the Dhok Pathan Formation, which can still be taxonomically referred to as Cremohipparion, but a formal description of the species with a new type of specimen is required. A more comprehensive study currently in preparation will attempt to resolve this issue
- 3. Eurygnathohippus sp. (3.6–2.6 Ma). Four mandibular cheek teeth from the Late Pliocene of the Siwaliks from the Potwar Plateau in Pakistan and the Siwalik Hills in India were identified by Jukar et al. [175] as Eurygnathohippus sp. These specimens all bear the characteristic single pli-caballinid and ectostylid.
- 4. Equus sivalensis, Falconer and Cautley, 1849 [179] (2.6–0.6 Ma). The lectotype is NHMUK PV M.16160, an incomplete cranium from the Siwaliks [183]. It is a large species of stenonine horse found in the Siwaliks of the Indian Subcontinent, ranging from the Potwar Plateau in the west to the Nepal Siwaliks in the east. The exact temporal distribution is unknown; however, based on paleomagnetic dating of the Pinjor Formation where the species was found, it likely ranges in age from 2.6 to 0.6 Ma [176,184]. However, some potentially older occurrences from just below the Gauss–Matuyama boundary (>2.6 Ma) have also been reported [185,186].
- 5. Equus sp. (~2.2–1.2 Ma). This is small species of Equus with smaller slender metapodials has been reported from the Pinjor Formation of the Upper Siwaliks. This species has been referred to as Equus sivalensis minor [187], Equus sp. A [188] or Equus cf. E. sivalensis [189]. A set of postcranial remains, including metapodials, astragali and phalanges, which were formerly tentatively referred to as “Hippotherium” antelopinum, are now believed to belong to this small species of Equus [138,184]. Based on specimens collected from the Mangla–Samwal Anticline and the Pabbi Hills in Pakistan, this species likely ranges in age from ~2.2 to 1.2 Ma [176]. Geographically, it ranges from the Pabbi Hills to the river Yamuna in the east.
- 6. Equus namadicus, Falconer and Cautley, 1849 [179] (~0.5–0.015 Ma). The lectotype is NHMUK PV M.2683, an incomplete cranium from the Siwaliks [183]. It is a large-sized stenonine horse from the Middle and Late Pleistocene of the Indian Subcontinent. The stratigraphic range includes the Middle Pleistocene Surajkund Formation in Central India [190] and Late Pleistocene deposits throughout peninsular India [177]. However, Lydekker [191] reported some specimens from the uppermost Upper Siwaliks, which might suggest that the species extends back to the early Middle Pleistocene.
4.3. Europe
- 1. Plesiohipparion longipes, Gromova, 1952 [193] (7–3.0 Ma). The holotype is PIN2413/5030, a complete mt3 from Pavlodar [193]. It has been identified from Pavlodar (Kazhakstan), Akkasdagi and Calta (Turkey) [194] and Baynunah (UAE) [8,195]. In all cases, Pl. longipes was recognized by its extreme length dimensions of mc3s and mt3s. None of these attributions of Plesiohipparion display the characteristics of extremely angled and pointed metaconids and metastylids of Pl. houfenense, Pl. huangheense, Pl. rocinantis or Pl. shanxiense. If all these taxa are referable to Plesiohipparion, the chronologic range would be 7 Ma to the Early Pleistocene and the geographic range being from China to Spain.
- 2. “Cremohipparion” fissurae, Crusafont and Sondaar, 1971 [196] (MN14-15). The holotype is an mt3 from Layna, Spain, figured in Crusafont and Sondaar [196] (Pl. 1). It was originally described as “Hipparion” fissurae. The species is recorded from the MN15 Pliocene localities in Spain [197] and more recently from the MN14 of Puerto de la Cadena [198]. The most recent analyses provided by Cirilli et al. [192] suggest an attribution to the genus Cremohipparion, yielding this species as the possible last representative of the genus in Europe. Its evolutionary framework is not yet defined.
- 3. Proboscidipparion crassum, Gervais, 1859 [199] (ca. 4.0–2.7 Ma; MN14-16). Deperet [200] described and figured the sample from Roussillon, France, without assigning a holotype or lectotype. The sample figured by Deperet [200] (Pl. V, Figures 6–10; Pl. VI) represents the hypodigm for the species. The sample is a small-sized equid with remarkable similarities with Pr. heintzi [192,194]. No complete crania are known from this species, whereas it is well documented by isolated upper and lower cheek teeth and postcranial elements. Bernor and Sen [194] showed that Pr. crassum also has a very short mc3, whereas Cirilli et al. [192] gave substantial indications in cranial and postcranial elements for its attribution to the genus Proboscidipparion. This species is mostly known from the Pliocene of France (Perpignan, Montpellier) but also from Dorkovo (Bulgaria) and Reg Crag (England) [197,201,202].
- 4. Proboscidipparion heintzi, Eisenmann and Sondaar, 1998 [203] (MN15). The holotype is MNHN.F.ACA49A, a complete mc3 from Calta, Turkey [203]. It was originally identified and described “Hipparion” heintzi [203], whereas Bernor and Sen [194] restudied and allocated this taxon to Pr. heintzi, recognizing the similarity of the Calta juvenile skull MNHN.F.ACA336 to Chinese Pr. pater in the retracted and anteriorly broadly open nasal aperture accompanied by very elongate anterostyle of dP2 [190]. It has only been reported from the locality of Calta.
- 5. Plesiohipparion rocinantis, Hernández-Pacheco, 1921 [204] (3.0–2.58 Ma). The lectotype is a p3/4 figured in Alberdi [205] (Pl. 6, Figure 4). It is the largest three-toed horse from Europe. Qiu et al. [122], followed by Bernor et al. [124,206] and Bernor and Sun [207], recognized this species as being a member of the Plesiohipparion clade by cranial and postcranial morphological features. Plesiohipparion rocinantis is reported between 3.0 and 2.6 Ma [208,209] from La Puebla de Almoradier, Las Higuerelas and Villaroya (Spain); Roca-Neyra (France); Red Crag (England); Kvabebi (Georgia). This species may include “Hipparion” moriturum from Ercsi (Hungary) and the sample from Sèsklo (Greece) previously ascribed to Plesiohipparion cf. Pl. shanxiense [210]. It represents the last occurrence of Plesiohipparion in Europe at the Plio–Pleistocene transition [8,192].
- 6. Equus livenzovensis, Bajgusheva, 1978 [212] (2.6–2.0 Ma). The holotype is POMK L-4, a fragmentary skull from Liventsovka [212] in the Early Pleistocene. The species represents the Equus Datum in Western Eurasia, Early Pleistocene localities dated at the Plio–Pleistocene boundary (2.58 Ma) such as Liventsovka (Russia), Montopoli (Italy) and El Rincón–1 (Spain) [184,212,213,214,215,216,217,218,219]. Recent research [214,216,220] suggests the occurrence of E. livenzovensis in Eastern European localities, dated between 2.58 and 2.0 Ma. The species shows the typical stenonine morphology, even though it is a large-sized horse.
- 7. Equus major, Delafond and Depéret, 1893 [221] (?2.6–1.9 Ma). The hypodigm is represented by a P2-M1 and a 1ph3 figured by Delafond and Depéret [221] from Chagny, France. It is poorly represented in the Early Pleistocene of Europe, being represented by few remains and localities. It was described by an incomplete upper cheek tooth row and postcranial elements from the Early Pleistocene locality of Chagny (Central France). Following the ICZN guidelines, Alberdi et al. [216] established that E. major has priority over Equus robustus Pomel, 1853 [222]; Equus stenonis race major Boule, 1891 [223]; Equus bressanus Viret, 1954 [224]; Equus major euxinicus Samson, 1975 [225]. Equus major has been reported from the European sites of Senèze, Chagny, Pardines and Le Coupet (France); Tegelen (the Netherlands); East Reunion and Norfolk (England) [216], and, as reported by Forsten [214], it may also be present at Liventsovka (Russia). It is a large-sized monodactyl horse, the largest species of the European Early Pleistocene.
- 8. Equus stenonis, Cocchi, 1867 [226] (2.45–ca. 1.6 Ma). The holotype is IGF560, a complete cranium from the Upper Valdarno Basin, Italy [13]. It was the most widespread Equus species during the Early Pleistocene (MNQ17 and MNQ18). In the last century, E. stenonis samples were identified under several subspecies as E. stenonis vireti, E. stenonis senezensis, E. stenonis stenonis, E. stenonis granatensis, E. stenonis guthi, E. stenonis mygdoniensis, E. stenonis anguinus, E. stenonis pueblensis and E. stenonis olivolanus. Recently, Cirilli et al. [13] reevaluated these subspecies, considering most of them to be ecomorphotypes of the same species, resulting in recognizing that E. stenonis is a monotypic, polymorphic species. At the present time, the species is reported as having occurred from Georgia to Spain in the circum-Mediterranean area as well as the Levant.
- 9. Equus senezensis, Prat, 1964 [227] (2.2–2.0 Ma). The lectotype includes a left P2-M3 and two mc3s, figured in Prat [227] (Pl.1 Figure C; Pl.2 Figure D,E). It is a medium-sized horse, distributed in France and Italy. Earlier referred to as E. stenonis senezensis [227], it has been recognized as a different species by Alberdi et al. [216] and Cirilli et al. [13]. It is morphologically similar to the European E. stenonis but with a reduced size. It is reported from the type locality of Senèze and possibly from Italy between 2.2 and 2.1 [228,229].
- 10. Equus stehlini, Azzaroli, 1964 [230] (1.9–1.78 Ma). The holotype is IGF563, an incomplete cranium from the Upper Valdarno Basin. It is the smallest Early Pleistocene Equus species. Over the last decades, it was considered both either a species or a subspecies of E. senezensis Azzaroli [216,230]. Cirilli [229] found that it can be considered a different Early Pleistocene species that probably evolved from E. senezensis. At the present time, E. stehlini is known only from Italy [228,229].
- 11. Equus altidens, von Reichenau, 1915 [82] (1.8–0.78 Ma). The lectotype is a right p2 figured in von Reichenau [82] (Pl. 6, Figure 17) from Sussenborn, Germany. It is a medium-sized horse, intermediate in size between E. stehlini and E. stenonis, occurring in Western Eurasia in the late Early to early Middle Pleistocene. Originally described from the Middle Pleistocene in Süssenborn [82], over the last decades its chronologic range was extended to the late Early Pleistocene, representing the most widespread species after 1.8 Ma until the Middle Pleistocene [231,232]. Recently, Bernor et al. [14] reported its first occurrence in Dmanisi (Georgia, 1.85–1.76 Ma), supporting the hypothesis of a dispersion of this species from east to west, being part of a faunal turnover that included several other mammalian species during this time frame [14,233,234]. It was also identified in Moldova (Tiraspol, layer 5) [129]. Several populations of E. altidens have been identified as different subspecies (i.e., E. altidens altidens and E. altidens granatensis). Nevertheless, within the nomen, E. altidens may also be included in other European taxa such as Equus marxi von Reichenau, 1915 [82]; Equus hipparionoides Vekua, 1962 [235]; E. stenonis mygdoniensis Koufos, 1992 [236]; Equus granatensis Eisenmann, 1995 [13,216,232,233,237,238]. Its origin remains controversial. Guerrero Alba and Palmqvist [239] proposed a possible African origin, claiming it to be part of the E. numidicus–E. tabeti evolutionary lineage. This latter hypothesis was also reported by Belmaker [240]. Eisenmann [87] included E. altidens in the new subgenus Sussemionus, with other Early and Middle Pleistocene species. More recently, Bernor et al. [14] proposed a new evolutionary hypothesis, considering that E. altidens originated in Western Asia and is potentially related to living E. hemionus and E. grevyi.
- 12. Equus suessenbornensis, Wüst, 1901 [241] (1.5–0.6 Ma). The lectotype is IQW1964/1177, a P2-M3 from Süssenborn, Germany. It is a large horse, larger than E. stenonis and E. livenzovensis but smaller than E. major. As for E. altidens, the species has been described from the Middle Pleistocene in Süssenborn [241], even if its best-known sample comes from the Georgian locality of Akhalkalaki (ca. 1.0 Ma) [242]. In addition to Akhalkalaki and Süssenborn, it has been reported in Central European localities such as Stránská Skála (Czech Republic) and Ceyssaguet and Solilhac (France) [216,243,244]. Over the last decades, its biochronologic range was extended to the late Early Pleistocene, with its earliest occurrence in the Italian localities of Farneta and Pirro Nord [216,231] and in the Spanish sites of Barranco León 5 and Fuente Nueva 3 [245].
- 13. Equus apolloniensis, Koufos et al., 1997 [246] (1.2–0.9 Ma). The holotype is LGPUT-APL-148, a nearly complete cranium from Apollonia, Greece [246]. It is a peculiar species of the late Early Pleistocene, mostly recorded from the locality of Apollonia–1 (Mygdonia Basin, Greece) and, possibly, from other localities of the Balkans and Anatolia [244,246,247]. As reported by Gkeme et al. [247], this species differs from E. stenonis and other European Early Pleistocene Equus, with a distinct cranial morphology, and its size is intermediate between E. stenonis and E. suessenbornensis. Koufos et al. [246] interpreted E. apolloniensis as an intermediate species between E. stenonis and E. suessenbornensis, whereas Eisenmann and Boulbes [248] considered E. apolloniensis as “a step within the lineage of asses”.
- 14. Equus wuesti, Musil, 2001 [249] (1.1–0.9 Ma). The holotypes are IQW1980/17067 and IQW1981/17619, two fragmentary mandibles with p2-m3 from Untermassfeld, Germany [249]. It has been established from the Epivillafranchian locality of Untermassfeld (Germany). Musil [249] reported isolated teeth, mandibles and long bones with primitive and derivate characteristics, and a larger size when compared with the widespread E. altidens. This evidence was also reported by Palombo and Alberdi [232], highlighting a more robust morphology of the postcranial elements. However, scholars disagree on its possible origin. Forsten [250] considered E. wuesti close to and derived from E. altidens, whereas others [249,251] recognized E. wuesti as the possible source for E. altidens. Nevertheless, considering the latest E. altidens discoveries [14,231,232,238], this last hypothesis seems to not be well supported. Palombo and Alberdi [232] suggest also that it can represent an ecomorphotype of E. altidens.
- 15. Equus petralonensis, Tsoukala, 1989 [252] (ca. 0.4 Ma). The holotype is PEC-500, an mc3 from Petralona Cave, Greece [252]. It is a slender and gracile horse from Greece (Petralona Cave). However, the taxonomic status of this species has been actively debated. Forsten [250] considered E. petralonensis as being a member of the E. altidens group, together with the Early Pleistocene equids from Libakos, Krimini and Gerakaou. Eisenmann et al. [67] synonymized E. petralonensis with Equus hydruntinus, within the subspecies E. hydruntinus petralonensis. Despite the taxonomic controversy surrounding this species, E. petralonensis may be considered a stenonine horse because of its mandibular cheek tooth morphology [246].
- 16. Equus graziosii, Azzaroli, 1969 [253] (MIS 6). No access number is available for the holotype, which is figured in Azzaroli [254] (Pl. XLV, Figure 1a,b) as a complete cranium from Val di Chiana, Arezzo. It is an enigmatic species from the late Middle–Late Pleistocene. It was described as a different species by Azzaroli [254] based on a partial cranium, mandibles and postcranial elements. According to Azzaroli [254], the cranium and maxillary and mandibular cheek teeth have typical asinine features, although some other authors have highlighted the morphological similarities with E. hydruntinus [244,255,256]. The evolutionary and phylogenetic position of E. graziosii is still questionable.
- 17. Equus hydruntinus, Regalia, 1907 [257] (0.6–0.01 Ma). There is no holotype. The hypodigm includes isolated upper and lower molars and the fragments of radius and tibia from Grotta Castello (Sicily, Italy). It is known also as the European wild ass, which is a small-sized and slender horse from the Middle and Late Pleistocene in Europe. Its geographic range spans across Europe and is documented in numerous localities [244]. Apparently, the first specimens of this species were also found in Central Asia in several Uzbekistan Late Weichselian localities associated with the Late Paleolithic [129,258]. Its evolutionary history has been debated by many authors, who have proposed different scenarios such as a direct origin from E. altidens [150,259,260], Equus tabeti [237,239] or, more recently, as a new taxon arrival from Asia [228,238]. Nevertheless, the most recent DNA analyses relate E. hydruntinus as a morphotype of the modern E. hemionus, proposing the subspecies E. hemionus hydruntinus [261]. However, several morphological features distinguish E. hydruntinus from E. hemionus (for a detailed discussion, see [244]), allowing for the consideration of E. hydruntinus as a still valid name for fossil identification. The oldest remains of E. hydruntinus were reported from the Middle Pleistocene levels of Vallparadìs, ca. 0.6 Ma [262,263], although the species has been documented in Western Eurasia until the Holocene [244]. Different subspecies have been suggested including E. hydruntinus minor (Lunel Viel, France), E. hydruntinus danubiensis (Romania), E. hydruntinus petralonensis (Petralona Cave, Greece) and E. hydruntinus davidi (Saint-Agneau, France). As reported by Boulbed and Van Asperen [244], E. hydruntinus adapted to semiarid, steppe conditions with a preference for temperate climates, although it could tolerate limited cold conditions.
- 18. Equus ferus, Boddaert, 1758 [264] (ca. 0.7–0.6 Ma). Boddaert [264] did not refer to any holotype or lectotype but gave a detailed description of the anatomical features of the species [264] (p. 159). It first appears in Western Eurasia in the early Middle Pleistocene, although a precise age is not available at the present time. The species has been questioned mostly from a taxonomic viewpoint, wherein many subspecies or different species have been erected to identify the Middle and Late Pleistocene fossil samples of the caballine horses as Equus mosbachensis, E. mosbachensis tautavelensis, E. mosbachensis campdepeyri, E. mosbachensis micoquii, E. mosbachensis palustris, Equus steinheimensis, Equus torralbae, Equus achenheimensis, E. ferus taubachensis, E. ferus piveteaui, E. ferus germanicus, E. ferus antunesi, E. ferus gallicus, E. ferus latipes, E. ferus arcelini and Equus caballus. These taxa are based on the size or morphological differences among the different fossil samples, representing an interesting case of morphological variability within the same lineage. These different subspecies have been considered to be chrono species by Eisenmann and Kuznetsova [265]. Nevertheless, van Asperen [266] noted that differences in the size and morphology of Middle and Late Pleistocene caballine horses can be observed, although they are not more variable than modern ponies or highly homogeneous groups such as Arabian horses or E. przewalskii [244]. Moreover, no unidirectional or evolutionary trend in size and shape can be identified, whereas morphology and size fluctuate over time [19,266]. For this reason, the proposal of Boulbes and van Asperen [244] to consider these species/subspecies as ecomorphological variants of the same species, E. ferus, seems the most parsimonious position. Moreover, as reported by van Asperen [267] following the ICZN, the correct species to indicate the wild caballine horses should be E. ferus and not E. caballus, which refers to domesticated forms. In addition, the genomic studies of Weinstock et al. [77] and Orlando et al. [153] support the genetic variation of the Middle and Late Pleistocene caballine horses.
4.4. Africa
- 1. Cremohipparion periafricanum, Villalta and Crusafont, 1957 [272] (6.8–4.0 Ma). The lectotype is a P2-M3 figured in Alberdi [205] (Pl.3, Figure 3) from Vadecebro II, Spain. It is a small (dwarf) hipparion that is a close relative (or senior synonym) of Cr. nikosi from the Quarry 5 levels of Samos, Greece, dated 6.8 Ma [206]. Fragmentary remains of Cr. aff. periafricanum have been reported from Tizi N’Tadderth, Morocco [273], and Sahabi, Libya [274].
- 2. Eurygnathohippus feibeli, Bernor and Harris, 2003 [275] (6.8–4.0 Ma). The holotype is KNM-LT139, a partial right forelimb including a fragmentary radius, mt3, a1ph3, a2ph3, partial mc2, a1ph2, a2ph2, a2ph3 and a partial mc4 [275]. There were additional dental and postcranial elements from Lower and Upper Nawata that were referred to this species. Bernor and Harris [275] suggested that the Ekora 4 cranium, ca. 4.0 Ma, was a late surviving member of Eu. feibeli. Whereas Churcher and Richardson [268] recognized “Hipparion” sitifense in the North African Late Miocene–Pliocene horizons, Bernor and Harris [275] and Bernor and Scott [276] noted that the type material described by Pomel [277] could not be located and could potentially be confused either with Cr. periafricanum or Eu. feibeli.
- 3. Sivalhippus turkanensis, Hooijer and Maglio, 1973 [278] (6.5–4.0 Ma). The holotype is KNM-LT136, an adult female cranium. Bernor and Harris [275] assigned this as being a species of Eurygnathohippus, Eu. turkanense. Subsequent studies of the Sivalhippus clade [174] and by Sun et al. [279] demonstrate the close identity of cranial, dental and, in particular, postcranial anatomy of Si. turkanensis and Si. perimensis and the extension of this genus into China and Africa in the Late Miocene.
- 4. Eurygnathohippus hooijeri, Bernor and Kaiser, 2006 [280] (5.0 Ma). The holotype is SAMPQ-L22187, a complete adult female skull with associated dentition, mandible and dentition and postcranial characteristics from the earliest Pliocene Langebaanweg E Quarry, South Africa [280]. Hooijer [281] originally described the specimen under the nomen “Hipparion” cf. H. baardi.
- 5. Eurygnathohippus woldegabrieli, Bernor et al., 2013 [282] (4.4–4.2 Ma). The holotype is ARA-VP-3/21, an incomplete mandible [282]. The hypodigm include also 156 dental and postcranial specimens from 14 localities at Aramis (Middle Awash), Ethiopia. The type specimen is ARA-VP-3/21 a mandible including symphysis, right partial ramus with p2 and p3, left ramus with p2 and 3 preserved and pr-m3 poorly preserved. Mandibular incisor teeth and canines, if originally present are lacking.
- 6. Eurygnathohippus afarense Eisenmann, 1976 [283] (KH3, Hadar, ca. 3.0 Ma). The holotype is AL363-18, a partial cranium from the Kada Hadar Member [283]. “Hipparion” afarense was nominated for skeletal material originating from the Kada Hadar 3 horizon, Hadar, Ethiopia. Eisenmann [283] also referred a mandible, AL177-2, to E. afarense.
- 7. Eurygnathohippus hasumense, Eisenmann, 1983 [284] (3.8–3.2 Ma). The holotype is KNM-ER 2776, a p4-m2 from zones B and C of the Kubi Algi Formation [284]. She included cheek teeth of common morphology from the Chemeron Formation, Kenya and the Denen Dora Member of the Hadar Formation, Ethiopia. A cranium with associated mandible was included in this hypodigm, AL340-8 [269] (Figure 13 in Bernor et al. [269]), and a partial skeleton including cheek teeth and complete postcranial elements, AL155-6 from DD2, ca. 3.2 Ma. Bernor et al. [281,285] analyzed a series of Ethiopian Eurygnathohippus establishing a phyletic relationship that currently includes Eu. feibeli, Eu. woldegabrieli, Eu. “afarense”, Eu. hasumense and Eu. cornelianus.
- 8. Eurygnathohippus pomeli Eisenmann and Geraads, 2006, [286] (ca. 2.5 Ma). The holotype is AaO-3647, an almost complete, but transversely crushed skull [286]. It was originally described as “Hipparion” pomeli. Reference [286] reported a well-preserved assemblage of hipparionini Ahl al Oughlam near Casablanca. Eisenmann and Geraads [286] argued that the sample is homogeneous and biochronologically correlative with eastern African faunas that are ca. 2.5 Ma, roughly contemporaneous with Omo Shungura D. Bernor and Harris [275] recognized this as a species of Eurygnathohippus. The northward extension of Plio–Pleistocene Eurygnathohippus into North Africa is remarkable as was its extension into India at this time [175].
- 9. Eurygnathohippus cornelianus, van Hoepen, 1930 [287] (ca. 2.6–1.0 Ma). The hypodigm includes a mandibular dentition from Cornelia, Orange Free State, with hypertrophied i1s and i2s and atrophied i3s placed immediately posterior to the i2s [287] (plates 20–22). Leakey [288] (pl. 20, 4 figures) reported the occurrence of “Stylohipparion albertense” (=Eu. cornelianus) from Bed II, Olduvai Gorge, Tanzania based on premaxillae and mandibular symphyses with identical incisor morphology. Hooijer [281] reported an adult skull from Olduvai BKII which he referred to Hipparion cf. H. ethiopicum which is likely a member of the Eu. cornelianus lineage. Eisenmann [284] did not recognize the existence of Eu. cornelianus at Olduvai, but referred an immature cranium, KNM-ER3539 to Eu. cornelianus. Armour-Chelu et al. [289] and Armour-Chelu and Bernor [290] argued that the first evidence of this clade may be from the Upper Ndolanya Beds, Tanzania, circa 2.6 Ma. It is also likely present in the Omo Shungura F, dated 2.36. Bernor et al. [8,269] advanced the hypothesis that Eu. cornelianus is a member of an evolving lineage that occurred in East and South Africa between ca. 2.4 and less than 1 Ma. The specimen/locality content of Eurygnathohippus is currently under investigation.
- 10. Hipparion (Eurygnathohippus) steytleri, van Hoepen, 1930 [287]. Van der Made et al. [291] argued that the author named Hipparion steytleri based on a right M1/1, left M3 and left m1-2 that formed a type series. Van Hoepen [287] also named Eu. cornelianus on the basis of a mandibular symphysis. Van der Made et al. [291] considered the nomen H. stytleri to have priority over Eu. cornelianus, but the cheek teeth of H. stytleri are of insufficient diagnostic value in themselves to define a valid species as stipulated by Article 75.5 of the ICZN. On the other hand, Van Hoepen [287] exercised considerable foresight in recognizing the highly derived state of the type specimen mandible of Eu. cornelianus because of its very wide mandibular symphysis with hugely hypertrophied i1 and i2 with very reduced, peg-like i3 situated immediately posterior along the mid-line of i2. Moreover, Leakey [288] illustrated a series of Eu. cornelianus mandibular symphyses and premaxillae from Olduvai Bed 1. Bernor et al. [292] described a juvenile skull of Eu. cornelianus (RMNH67) from Olduvai Gorge [293] which has a long preorbital bar, faint preorbital fossa and a dP2 with an extended anterostyle. Eurygnathohippus cornelianus sensu strictu has a known chronologic range of 2–1 Ma, but the lineage apparent extends lower in time.
- 11. Hipparion (Eurygnathohippus) libycum Pomel, 1897 [273]. The hypodigm includes a left p3/4 figured by Pomel ([273], pl. 1, Figures 5–7), two lower cheek teeth from “carriers des gres ouvertes a la campagne Brunie” in Oran (pl. 1, Figures 1–7) and a distal epiphysis of a third metatarsal from “carriers de grès du quartier”, St-Pierre, Oran. Hopwood [289] assigned the left p3/4 figured by Pomel ([273], pl. 1, Figures 5–7) as a lectotype [287]. It shows gracile metapodials. Van der Made et al. [287] reported that the original type specimens from Oran are in the Central Faculty of Algiers (MGFCA) but have provided no accession numbers for the holotype.
- 12. Hipparion (Eurygnathohippus) ambiguum, Pomel, 1897 [277]. The hypodigm includes a right P2 from Beni Fouda (Ain Boucherit) [277] (pl. 2, Figures 2–4). Previously the repository was unknown. Van der Made et al. [291] (Figures 2a,b and 3), redrafted an image by Pomel [277] and reported that the specimen is currently maintained in the Central Faculty of Algiers (MGFCA) but offered no photographic images or measurements of this specimen. Van der Made et al. [291] have provided no accession number for the holotype. The type locality of Aïn Boucherit has a magnetostratigraphic date of 2.44 [291] whereas the East African localities referred to by Van der Made et al. [291] have an age range of 3.8–1.2 Ma [269].
- 13. Hipparion (Eurygnatohippus) massoesylium, Pomel [277]. The hypodigm includes five teeth from “puits Carouby” and “aux portes d’Oran” (also Puits Karoubi), left P4, M1-3 and right M3 (pl. 1, Figures 8–10). Van der Made et al. [291] considered these specimens to be the holotype of the species. Van der Made et al. [291] have reported that the holotype is kept in the Central Faculty of Algiers (MGFCA) but have provided no accession numbers for the associated cheek teeth.
- 14. “Hipparion” sitifense, Pomel, 1897 [277]. The hypodigm includes four specimens including two teeth from St. Arnaud, a calcaneum from a nearby locality and a tooth figured by Thomas [294]. Pomel figured an M1/2 (Pl. 1, Figures 13–16), a right P4 (Pl. 1, Figures 11 and 12) and a calcaneum (Pl.2, Figures 9 and 10). The M1/2 figured by Pomel represents the lectotype [291]. The latter authors re-figured the original illustrations from Pomel [277], but this figure does not have a scale bar, and the specimens do not have formal institutional accession numbers. Eisenmann [295] stated that it is not known where the “type specimens” from Saint Arnaud et al. are, whereas Van der Made et al. [291] report that the original material is in the Central Faculty of Algiers (MGFCA; p. 44). Van der Made et al. [291] nominated a lectotype citing a specimen figured by Pomel [277] (pl. 1, Figures 13–15) without specifically designating a specimen accession number, institution, element and providing a redrafted original figure of the specimen without the benefit of a scale. It cannot be known if “H. sitifense” is referable to the genus Hipparion or other small hipparionins Cremohipparion or Eurygnathohippus (for which mandibular cheek teeth are needed). Arambourg [296,297] reported a specimen “from the type locality (for which we cannot be certain)” described and figured a mandibular specimen “with rounded metaconid-metastylid and no ectostylid”. It should be noted that this material is not a legitimate sample of “the original type series of Pomel, 1897”. Arambourg [296,297] assigned material from Ain el Hadj Baba, Mascara, Saint Donat and Aïn el Bey to “H. sitifense”. These are all small sized.
- 15. Equus numidicus, Pomel, 1897 [277] (2.44–1.2 Ma). The hypodigm includes a right P2 [273] (Pl. 2, Figure 2). Arambourg reported cranial and postcranial elements from Aïn Boucherit and Aïn Jourdel (Pl. 18, Figures 6 and 7; Pl. 19–20; Figures 58 and 62). It is a medium-sized horse approximately the size of a large zebra that originated from Aïn Boucherit and Aïn Hanech, Algeria, ca. 2.44–2.0 Ma [270,271,295]. No complete crania are known, although incomplete cranial remains associated with postcranial elements have been reported [296]. The evolutionary relationships of E. numidicus are not well defined. Azzaroli [298] pointed out a possible relationship to E. stenonis, although it was no longer investigated. However, some anatomical features of the upper and lower cheek teeth and postcranial elements resemble those of E. stenonis [297,298].
- 16. Equus tabeti, Arambourg, 1970 [297] (2.44–1.2 Ma). The holotype is a partial palate (1949.2:773) figured in Arambourg [297] (Pl. 21, Figure 3). It is a medium-small sized species of Equus with “asinine” maxillary cheek teeth, stenonine mandibular cheek teeth and slender third metapodials and phalanges [284,297]. No complete crania are known, even if some incomplete crania and mandibles have been reported by Arambourg [297]. The type material originates from Aïn Hanech, Algeria [297]. Geraads [299] estimated the age of Aïn Hanech to be 1.2 Ma. Eisenmann [284] reported the possible presence of Equus cf. tabeti at Koobi Fora, Kenya believing that it is a primitive ass and may have been derived from E. numidicus. The recent analyses of Duval et al. [270] suggest an earlier occurrence of E. tabeti and E. numidicus at 2.44 Ma in the lowermost levels of Aïn Boucherit. Beside North Africa, E. tabeti is reported from Early Pleistocene sites of “Ubeidiya”, Bizat Ruhama and Qafzed (Levantine corridor [300,301,302]) and from East Africa [183].
- 17. Equus koobiforensis, Eisenmann, 1983 [284] (2.1–1.0 Ma). The holotype specimen is KNM-ER 1484, a complete skull recovered from the Notochoerus scotti Zone, Area 130, just below the KBS Tuff, ca. 1.9 Ma. Eisenmann [284] reported a number of close dental similarities shared by E. koobiforensis and European E. stenonis but did not suggest a direct phylogenetic relationship between these taxa. Azzaroli [258] stated that E. koobiforensis was essentially a Grevy’s zebra. Bernor et al. [183] cited the likely evolutionary relationship between North American Pliocene E. simplicidens, European E. stenonis and E. koobiforensis. Cirilli et al. [10] demonstrated the explicit cladistic relationships between the E. simplicidens -E. stenonis-E. koobiforensis-E. grevyi clade, whereas Cirilli et al. [13] reinforced this result on robust statistical grounds. No precise age is available for its last occurrence in the fossil record.
- 18. Equus oldowayensis, Hopwood, 1937 [293] (1.9–1.0 Ma). The holotype is a lower jaw from an animal approximately 2 years old [293] (Figures 1 and 2; Catalogue Number VIII, 353, in the Bayerische Paläontologische Staatssammlung, Munich). Hopwood [293] also designated a lower incisive region with the left incisors and right first incisor (BMNH M14199) as the paratype. The original Olduvai collection deposited in Munich, which included the type of E. oldowayensis, was destroyed together with its catalogue, during WW II (K. Heissig personal communication with Churcher and Hooijer; [269]). Equus oldowayensis is usually reported from the type locality of Olduvai (1.8 Ma, Tanzania), but no precise ages are available for its chronologic range. Recently, Bernor et al. [183] reported an incomplete cranium from Olorgesailie (ca. 1.0 Ma, Kenya).
- 19. Equus capensis, Broom, 1909 [303] (ca. 2.0?–? Ma). It was a large-bodied horse estimated to be 150 cm at the withers, with a body mass of approximately 400 kg [304]. It originated from South Africa. Churcher [305] synonymized E. helmei, E. cawoodi, E. kubmi, E. zietsmani and specimens of E. harrisi and E. plicatus into E. capensis. Equus capensis was widely distributed in the Plio–Pleistocene of South Africa, although no information is known about its first and last occurrence in the fossil record.
- 20. Equus mauritanicus, Pomel, 1897 [277] (1.0 Ma). The hypodigm include isolated teeth and postcranial elements, figured in Pomel [277] (Pl.3–8). It is reported from a large sample from Tighenif (Algeria). Churcher and Richardson [268] referred E. mauritanicus to a subspecies of E. burchellii (=E. quagga mauritanicus). Eisenmann [284,295] recognized E. mauritanicus as a distinct species of Hippotigris and claimed similarities to E. stenonis in the dentition. Eisenmann [284] further introduced the notion of cross-breeding between E. mauritanicus and Quaggas. Churcher and Richardson [268] reported an extensive distribution of E. (Hippotigris) “burchelli” (=quagga) from North, East and South Africa. The possible relationships of E. mauritanicus with plain zebras have been suggested by Eisenmann [295] and more recently from Bernor et al. [14] by morphological analyses. Beside the type locality of Tighenif, E. mauritanicus is reported also in Oumm Qatafa (Juden Desert, Egypt, Middle Pleistocene [306]).
- 21. Equus melkiensis Bagtache et al. 1984 [307] (Late Pleistocene). The holotype is a short mc3 (I.P.H. Allo. 61–1314) recovered from Allobroges, Algeria and of latest Pleistocene age. Eisenmann [236] reported E. melkiensis also from Morocco. This species has been identified also at Gesher Benot Ya’akov and Nahal Hesi (Israel; [87,301]) and Oumm Qatafa (Egypt, Middle Pleistocene, [306]).
- 22. Equus algericus, Bagtache et al. 1984 [307] (Late Pleistocene). The holotype is IPH61-103, a m2 from Allobroges, Algeria [307] (Figure 1). It is reported to be a caballine species with a withers height of approximately 1.44 m. Equus algericus was also reported from Morocco [308], which are purported to have the characteristic caballine metaconid-metastylid (=double knot) morphology.
- 23. Equus grevyi, Oustalet, 1882 [309] (0.5–0.0 Ma). Oustalet [309] (v.10, pp. 12–14) described the anatomical features of the species, with an associated illustration (Figures 1 and 2). He also reported that the living animal was donated to the Museum National d’Histoire Naturelle in Paris, where the holotype should be kept. Equus grevyi is the largest living wild equid with a withers height of 140–160 cm. Azzaroli [259], Bernor et al. [14,183] and Cirilli et al. [10,13] all have cited the close relationship between European E. stenonis, E. koobiforensis and E. grevyi. Equus grevyi is currently distributed in the arid regions of Ethiopia and Northern Kenya and has recently vanished from Somalia, Djibouti and Eritrea [269]. Eisenmann [284] recognized Equus cf. E. grevyi from the Metridiochoerus compactus zone, the Guomde Formation and Galana Boi beds of Kenya based on both cheek tooth and postcranial remains. Bernor et al. [183] and Cirilli et al. [10,13] have recognized E. grevyi as a terminal member of the E. simplicidens–E. stenonis–E. koobiforensis–E. grevyi clade. Recently, O’Brien et al. [310] reported an incomplete cranium ascribed to E. grevyi from the Kapthurin Formation (Kenya) dated between 547 and 392.6 ka. This age represents the best dated E. grevyi FAD, at the present time.
- 24. Equus quagga, Boddaert, 1785 [264] (?1.0–0.0 Ma). Boddaert [264] did not identify a holotype or lectotype for E. quagga but gave a description of the anatomical features of the species (p. 160). It has a shoulder height ranging from a mean of 128 cm in males and 123 cm in females. Equus quagga is one of the most widely distributed African ungulates ranging from southern Sudan and southern Ethiopia to northern Nambia and northern South Africa. Several subspecies have been recognized including E. quagga crawshaii, E. quagga borensis, E. quagga boehmi, E. quagga chapmani, E. quagga burchellii and E. quagga quagga. Fossil remains have been reported from North to South Africa. Leonard et al. [311] suggest that the various subspecies of E. quagga differentiated between 120 and 290 ka. Fossil remains of quagga have been reported from South African Plio–Pleistocene karst deposits, but their certain identity in North and East Africa are somewhat elusive [269]. Pedersen et al. [312] have identified a South African region as the likely source for the origin of the plain zebras from which all extant populations expanded from at approximately 370 ka. Moreover, the genetic analyses of Pedersen et al. [312] have reported a remarkable gene flow in the extant E. quagga subspecies, highlighting the challenge of identifying the subspecific designation only by morphology, identifying at least four genetic clusters for E. quagga boehmi and E. quagga crawshayi.
- 25. Equus zebra, Linnaeus, 1758 [313] (?0.5–0.0 Ma). Linnaeus [313] does not refer any holotype or lectotype but gives a description of the anatomical features of the species (p. 101). It is a medium-sized, long-legged zebra with a mean shoulder height ranging from 124–127 cm. There are two recognized sub-species, E. zebra zebra (Cape Mountain Zebra) and E. zebra hartmannae (Hartmann’s Mountain Zebra). Churcher and Richardson [264] report a relatively small sample from the Middle Pleistocene to recent fossil remains of E. zebra in South Africa.
- 26. Equus (Asinus) africanus, Heuglin and Fitzinger, 1866 [314]. The lectotype is designated as a skull of an adult female collected by von Heuglin near Atbarah River, Sudan, and in Stuttgart (SMNS32026). Equus (Asinus) africanus are equines of small size and stocky build. Churcher [315] reported the earliest occurrence of this taxon from the middle of Bed II, Olduvai Gorge (>1.2 Ma). This identification was based on a single third metatarsal, which was short (231 mm) and slender, although not to the extent of E. tabeti. Two subspecies are recognized, E. (A.) africanus africanus von Heuglin and Fitzinger, 1866 [314] (Nubian wild ass) and E. (A.) africanus somaliensis Noak, 1884 [316] (Somali wild ass) [317,318,319]. The African wild ass E. africanus is widely believed to have been the ancestor of the domestic donkey [269] and, more recently, believed to be the descendant of the Early Pleistocene species E. tabeti [320]. Nevertheless, this last evidence has only been suggested and not yet proven.
5. Biochronology and Biogeography
5.1. NALMA Timescale and Equinae Evolution
5.2. SALMA Timescale and Equinae Evolution
5.3. ALMA Timescale and Equinae evolution
5.4. ELMA Timescale and Equinae Evolution
5.5. AFLMA Timescale and Equinae Evolution
5.6. General Remarks about Equinae Biochronology
6. Paleoecology
6.1. Relationships of Diet, Habitats and Body Size in Equines, with Pleistocene Equus from Eurasia and Africa as a Particular Example
6.2. North America
6.3. Eurasia
6.4. Africa
7. Climate and Evolution
8. Phylogeny
8.1. What Is Equus? Paradigms, Phylogeny, and Taxonomy
8.1.1. What Is a Genus?
8.1.2. Morphological Phylogenetic Analysis of Derived Equini
8.1.3. Phylogentic Gaps and the Concept of Equus
8.1.4. Uniqueness of Adaptive Zone and the Concept of Equus
8.1.5. Crown Group and the Concept of Equus
8.1.6. Divergence Time and the Concept of Equus
8.1.7. Taxonomic Implications
8.2. Cirilli et al. [10] Phylogeny: Equus Modeled as a Sigle Monophyletic Clade
8.2.1. Phylogenetic Gaps, Crown Group, Adaptive Zone, and Divergence Time Applied to the Phylogeny of Cirilli et al. [10]
8.2.2. Living and Fossil Equids
8.3. The Contribution from the Molecular Phylogeny
9. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Marsh, C.O. Polydactyle Horses, Recent and Extinct. Am. J. Sci. 1879, 17, 499–505. [Google Scholar] [CrossRef]
- Gidley, J.W. Revision of the Miocene and Pliocene Equidae in North America. Bull. Am. Mus. Nat. Hist. 1907, 23, 865–934. [Google Scholar]
- Osborn, H.F. Equidae of the Oligocene, Miocene, and Pliocene of North America. Iconographic type revision. Mem. Am. Mus. Nat. Hist. New Ser. 1918, 2, 1–326. [Google Scholar]
- Matthew, W.D. A new link in the ancestry of the horse. Am. Mus. Novit. 1924, 131, 130–131. [Google Scholar]
- Stirton, R.A. Phylogeny of North American Equidae. Univ. Calif. Publ. Geol. Sci. 1940, 25, 165–198. [Google Scholar]
- Simpson, G.G. Horses: The Story of the Horse Family in the Modern World and through Sixty Million Years of History; Oxford University Press: New York, NY, USA, 1951; pp. 1–247. [Google Scholar]
- MacFadden, B.J. Fossil Horses—Evidence for Evolution. Science 2005, 307, 1728–1730. [Google Scholar] [CrossRef] [PubMed]
- Bernor, R.L.; Kaya, F.; Kaakinen, A.; Saarinen, J.; Fortelius, M. Old World hipparion evolution, biogeography, climatology and ecology. Earth Sci. Rev. 2021, 221, 103748. [Google Scholar] [CrossRef]
- Barrón-Ortiz, C.I.; Avilla, L.S.; Jass, C.N.; Bravo-Cuevas, V.M.; Machado, H.; Mothé, D. What is Equus? Reconciling taxonomy and phylogenetic analyses. Front. Ecol. Evol. 2019, 7, 343. [Google Scholar] [CrossRef]
- Cirilli, O.; Pandolfi, L.; Rook, L.; Bernor, R.L. Evolution of Old World Equus and origin of the zebra-ass clade. Sci. Rep. 2021, 11, 10156. [Google Scholar] [CrossRef]
- Eisenmann, V.; Alberdi, M.T.; De Giuli, C.; Staesche, U. Methodology. In Studying Fossil Horses; Woodburne, M., Sondaar, P.Y., Eds.; E.J. Brill Press: Leiden, The Netherlands, 1988; pp. 1–71. [Google Scholar]
- Bernor, R.L.; Tobien, H.; Hayek, L.A.C.; Mittmann, H.W. Hippotherium primigenium (Equidae, Mammalia) from the late Miocene of Höwenegg (Hegau, Germany). Andrias 1997, 10, 1–230. [Google Scholar]
- Cirilli, O.; Saarinen, J.; Pandolfi, L.; Rook, L.; Bernor, R.L. An updated review on Equus stenonis (Mammalia, Equidae): New implications for the European Early Pleistocene Equus taxonomy and paleoecology, and remarks on the Old World Equus evolution. Quat. Sci. Rev. 2021, 269, 107155. [Google Scholar] [CrossRef]
- Bernor, R.L.; Cirilli, O.; Buskianidze, M.; Lordkipanidze, D.; Rook, L. The Dmanisi Equus: Systematics, biogeography and paleoecology. J. Hum. Evol. 2021, 158, 103051. [Google Scholar] [CrossRef] [PubMed]
- Fortelius, M.; Solounias, N. Functional characterization of ungulate molars using the abrasion-attrition wear gradient: A new method for reconstructing paleodiets. Am. Mus. Novit. 2000, 3301, 1–36. [Google Scholar] [CrossRef]
- Solounias, N.; Semprebon, G. Advances in the reconstruction of ungulate ecomorphology with application to early fossil equids. Am. Mus. Novit. 2002, 3366, 1–49. [Google Scholar] [CrossRef]
- Oksanen, O.; Žliobaitė, I.; Saarinen, J.; Lawing, A.M.; Fortelius, M. A Humboldtian approach to life and climate of the geological past: Estimating palaeotemperature from dental traits of mammalian communities. J. Biogeogr. 2019, 46, 1760–1776. [Google Scholar] [CrossRef]
- Scott, K.M. Postcranial dimensions of ungulates as predictors of body mass. In Body Size in Mammalian Palaeobiology; Damuth, J., MacFadden, B.J., Eds.; Cambridge University Press: New York, NY, USA, 1990; pp. 301–355. [Google Scholar]
- Saarinen, J.; Cirilli, O.; Strani, F.; Meshida, K.; Bernor, R.L. Testing equid body mass estimate equations on modern zebras—with implications to understanding the relationship of body size, diet and habitats of Equus in the Pleistocene of Europe. Front. Ecol. Evol. 2021, 9, 622412. [Google Scholar] [CrossRef]
- Saarinen, J.; Eronen, J.; Fortelius, M.; Seppä, H.; Lister, A.M. Patterns of diet and body mass of large ungulates from the Pleistocene of Western Europe, and their relation to vegetation. Palaeontol. Electron. 2016, 19, 1–58. [Google Scholar] [CrossRef]
- Mihlbachler, M.C.; Rivals, F.; Solounias, N.; Semprebon, G.M. Dietary change and evolution of horses in North America. Science 2011, 331, 1178–1181. [Google Scholar] [CrossRef]
- The NOW Community. New and Old World Database of Fossil Mammals (NOW). Licensed under CC BY 4.0. 2022. Available online: https://nowdatabase.org/now/database/ (accessed on 4 April 2022).
- Fortelius, M.; Eronen, J.T.; Jernvall, J.; Liu, L.; Pushkina, D.; Rinne, J.; Tesakov, A.; Vislobokova, I.A.; Zhang, Z.; Zhou, L. Fossil mammals resolve regional patterns ofEurasian climate change during 20 million years. Evol. Ecol. Res. 2002, 4, 1005–1016. [Google Scholar]
- Eronen, J.T.; Evans, A.S.; Fortelius, M.; Jernvall, J. The impact of regional climate on the evolution of mammals: A case study using fossil horses. Evolution 2010, 64, 398–408. [Google Scholar] [CrossRef]
- Eronen, J.T.; Polly, P.D.; Fred, M.; Damuth, J.; Frank, D.C.; Mosbrugger, V.; Scheidegger, C.; Stenseth, N.C.; Fortelius, M. Ecometrics: The grains that bind the past and present together. Integr. Zool. 2010, 5, 88–101. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Puolamäki, K.; Eronen, J.T.; Mirzaie Ataabadi, M.; Hernesniemi, E.; Fortelius, M. Dentalfunctional traits of mammals resolve productivity in terrestrial ecosystemspast and present. Proc. R. Soc. Lond. Biol. Sci. 2012, 279, 2793–2799. [Google Scholar]
- Orlando, L.; Vilstrup, L.; Ginolhac, A.; Zhang, G.; Froese, D.; Albrechtsen, A.; Stiller, M.; Schubert, M.; Cappellini, E.; Petersen, B.; et al. Recalibrating Equus evolution using the genome sequence of an early Middle Pleistocene horse. Nature 2013, 499, 74–80. [Google Scholar] [CrossRef]
- Jónsson, H.; Schubert, M.; Seguin-Orlando, A.; Ginolhac, A.; Petersen, L.; Fumagalli, M.; Albrechtsen, A.; Petersen, B.; Korneliussen, T.S.; Vilstrup, J.T.; et al. Speciation with gene flow in equids despite extensive chromosomal plasticity. Proc. Natl. Acad. Sci. USA 2014, 111, 18655–18660. [Google Scholar] [CrossRef] [PubMed]
- Heintzman, P.D.; Zazula, G.D.; MacPhee, R.D.; Scott, E.; Cahill, J.A.; McHorse, B.K.; Kapp, J.D.; Stiller, M.; Wooller, M.J.; Orlando, L.; et al. A new genus of horse from Pleistocene North America. eLife 2017, 6, e29944. [Google Scholar] [CrossRef]
- Hulbert, R.C. Calippus and Protohippus (Mammalia, Perissodactyla, Equidae) from the Miocene (Barstovian-early Hemphillian) of the Gulf Coastal Plain. Bull. Fla. State Mus. Biol. Sci. 1988, 32, 221–340. [Google Scholar]
- Olson, E.C.; McGrew, P.O. Mammalian fauna from the Pliocene of Honduras. Bullettin Geol. Soc. Am. 1941, 52, 1219–1244. [Google Scholar] [CrossRef]
- Drescher, A.B. Later Tertiary Equidae from the Tejon Hills, California. Carneg. Inst. Wash. Pub. 1941, 530, 1–23. [Google Scholar]
- Cope, E.D. A new Hippidium. Am. Nat. 1880, 14, 223. [Google Scholar]
- Matthew, W.D.; Stirton, R.A. Equidae from the Pliocene of Texas. Bullettin 1930, 19, 349–396. [Google Scholar]
- Lance, J.F. Palaeontología y estratigrafía del Pliocene de Yepómera, estado de Chihuahua 1a Parte: Equidos, excepto Neohipparion. Inst. Geol. Univ. Nac. Autónoma México 1950, 54, 83–94. [Google Scholar]
- Prado, J.E.; Alberdi, M.T. A cladistic analysis of the horses of the tribe Equini. J. Paleontol. 1996, 39, 663–680. [Google Scholar]
- Kelly, T.S. New Middle Miocene equid crania from California and their implications for the phylogeny of the Equini. Nat. Hist. Mus. Los Angel. Cty. Contrib. Sci. 1998, 473, 1–43. [Google Scholar] [CrossRef]
- Cope, E.D. A Preliminary Report on the Vertebrate Paleontology of the Llano Estacado; Geological Survey of Texas: Austin, TX, USA, 1893; pp. 1–136. [Google Scholar]
- Leidy, J. Notices of some remains of extinct Mammalia, recently discovered by Dr. F.V. Hayden, in the badlands of Nebraska. Proc. Acad. Nat. Sci. Phila. 1856, 8, 59. [Google Scholar]
- Mooser, O. Fossil Equidae from the middle Pliocene of the Central Plateau of Mexico. Southwest. Nat. 1968, 13, 1–12. [Google Scholar] [CrossRef]
- Cope, E.D. Pliocene horses of southwestern Texas. Am. Nat. 1885, 19, 1208–1209. [Google Scholar]
- Sondaar, P.Y. The osteology ofthe manus of fossil and Recent Equidae. Ned. Akad. Wettenschappen 1968, 25, 1–76. [Google Scholar]
- MacFadden, B.J. Systematics and phylogeny of Hipparion, Neohipparion, Nannippus, and Cormohipparion (Mammalia, Equidae), from the Miocene and Pliocene of the New World. Bull. Am. Mus. Nat. Hist. 1984, 179, 196. [Google Scholar]
- MacFadden, B.J. Fossil Horses: Systematics, Paleobiology, and Evolution of the Family Equidae; Cambridge University Press: Cambridge, UK, 1992. [Google Scholar]
- Merriam, J.C. New Species of the Hipparion Group from the Pacific Coast and Great Basin Provinces of North America; University of California Press: Oakland, CA, USA, 1915; Volume Voume 9, pp. 1–8. [Google Scholar]
- Hulbert, R.C.; Harrington, R. An early Pliocene hipparionine horse from the Canadian Arctic. Paleontology 1999, 42, 1017–1025. [Google Scholar] [CrossRef]
- MacFadden, B.J.; Skinner, M. Diversification and biogeography of the one-toed horses Onohippidium and Hippidion. Yale Peabody Mus. Nat. Hist. Postilla 1979, 175, 1–9. [Google Scholar]
- Hulbert, R.C. Late Neogene Neohipparion (Mammalia, Equidae) from the Gulf Coastal Plain of Florida and Texas. J. Paleontol. 1987, 61, 809–830. [Google Scholar] [CrossRef]
- Webb, S.D.; Hulbert, R.C. Systematics and evolution of Pseudhipparion (Mammalia, Equidae) from the late Neogene of the Gulf Coastal Plain and the Great Plains. In Vertebrates, Phylogeny, and Philosophy; Flanagan, K.M., Lillegraven, J.A., Eds.; University of Wyoming: Laramie, WY, USA, 1986; pp. 237–272. [Google Scholar]
- Merriam, J.C. Correlation between the Tertiary of the Great Basin and That of the Marginal Marine Province in California. Science 1914, 40, 643–645. [Google Scholar] [CrossRef] [PubMed]
- Dalquest, W.W.; Donovan, T.J. A new three-toed horse (Nannippus) from the late Pliocene of Scurry County, Texas. J. Paleont. 1973, 47, 34–45. [Google Scholar]
- Cope, E.D. A contribution to the vertebrate paleontology of Texas. Proc. Am. Philos. Soc. 1892, 30, 123–131. [Google Scholar]
- Dalquest, W.W.; Schultz, G.E. Ice Age Mammals of Northwestern Texas; Midwestern State University Press: Wichita Falls, TX, USA, 1992. [Google Scholar]
- Orlando, M.F. Order Perissodactyla. In Early Pleistocene Preglacial and Glacial Rocks and Faunas of North-Central Nebraska; Skinner, M.F., Hibbard, C.W., Eds.; American Museum of Natural History: New York, NY, USA, 1972; Volume 148, pp. 117–125. [Google Scholar]
- Gidley, J.W. A new Pliocene horse from Idaho. J. Mammal. 1930, 16, 52–60. [Google Scholar] [CrossRef]
- Winans, M.C. Revision of North American fossil species of the genus Equus (Mammalia: Perissodactyla: Equidae). Ph.D. Thesis, University of Texas, Austin, TX, USA, 1985; p. 264. [Google Scholar]
- Forsten, A.; Eisenmann, V. Equus (Plesippus) simplicidens (Cope), not Dolichohippus. Mammalia 1995, 59, 85–89. [Google Scholar] [CrossRef]
- Gazin, C.L. A study of the fossil horses remains from the Upper Pliocene of Idaho. Proc. U.S. Natl. Mus. 1936, 83, 281–320. [Google Scholar] [CrossRef]
- Merriam, J.C. New Mammalia from the Idaho Formation. Bull. Dep. Geol. 1918, 10, 523–530. [Google Scholar]
- Scott, E. Equus idahoensis from the Pliocene of Arizona, and Its Role in Plesippine Evolution in the American Southwest. 2005. Available online: http://www.sbcounty.gov/museum/discover/divisions/geo/pdf/SVP_2005b_EquusIdahoensis.pdf (accessed on 10 March 2022).
- Downs, T.; Miller, G.J. Late Cenozoic equids from the Anza-Borrego Desert of California. Nat. Hist. Mus. Los Angel. Co. Contrib. Sci. 1994, 440, 1–90. [Google Scholar] [CrossRef]
- Forsten, A. Mitochondrial-DNA time-table and ethe evolution of Equus: Comparison of molecular and paleontological evidence. Ann. Zool. Fenn. 1992, 28, 301–309. [Google Scholar]
- Azzaroli, A. The genus Equus in Europe. In European Neogene Mammal Chronology; Lindsay, E.H., Fahlbusch, V., Mein, P., Eds.; Plenum Press: New York, NY, USA, 1990; pp. 339–355. [Google Scholar]
- Troxell, E.L. The Vertebrate Fossils of Rock Creek, Texas. Am. J. Sci. 1915, 39, 613–638. [Google Scholar] [CrossRef]
- Hibbard, C.W. Pleistocene vertebrates from the Upper Becerra (Becerra Superior) formation, Valley of Tequixquiac, Mexico, with notes on other Pleistocene forms. Contrib. Mus. Paleontol. Univ. Mich. 1955, 12, 47–96. [Google Scholar]
- Eisenmann, V.; Howe, J.; Pichardo, M. Old World Hemiones and New World slender species (Mammalia, Equidae). Palaeovertebrata 2008, 36, 159–233. [Google Scholar] [CrossRef]
- Hulbert, R.C. Equus from Leisey Shell Pit 1A and other Irvingtonian localities from Florida. Bull. Fla. Mus. Nat. Hist. 1995, 37, 553–602. [Google Scholar]
- Gidley, J.W. A new species of Pleistocene horse from the Staked Plains of Texas. Bull. Am. Mus. Nat. Hist. 1900, 13, 111–116. [Google Scholar]
- Azzaroli, A.; Voorhies, M. The genus Equus in North America: The Blancan species. Palaeontogr. Ital. 1993, 80, 175–198. [Google Scholar]
- Owen, R. On Fossil Remains of Equines from Centrai and South America. Philos. Trans. Lond. 1869, 159, 559–573. [Google Scholar]
- Mooser, O.; Dalquest, W.W. Pleistocene mammals from Aguascalientes, Central Mexico. J. Mammal. 1975, 56, 781–820. [Google Scholar] [CrossRef]
- Cope, E.D. Extinct Mammalia of the Valley of Mexico. Proc. Am. Philos. Soc. 1884, 22, 1–15. [Google Scholar]
- Gidley, J.W. Tooth characters and revision of the North American species of the genus. Bull. Am. Mus. Nat. Hist. 1901, 14, 91–142. [Google Scholar]
- Azzaroli, A. The genus Equus in North America—The Pleistocene species. Paleontogr. Ital. 1998, 85, 1–60. [Google Scholar]
- Hay, O.P. Description of a new species of Pleistocene horse, Equus lambei, from the Pleistocene of Yukon Territory. Proc. U. S. Natl. Mus. 1917, 53, 435–443. [Google Scholar] [CrossRef][Green Version]
- Hay, O.P. Contributions to the knowledge of the mammals of the Pleistocene of North America. Proc. U. S. Natl. Mus. 1915, 48, 515–575. [Google Scholar] [CrossRef]
- Weinstock, J.; Willerslev, E.; Sher, A.; Tong, W.; Ho, S.; Rubenstein, D.; Storer, J.; Burns, J.; Martin, L.; Bravi, C.; et al. Evolution, systematics, and phylogeography of Pleistocene horses in the New World: A molecular perspective. PLoS Biol. 2005, 3, e241. [Google Scholar] [CrossRef]
- Vershinina, A.O.; Heintzman, P.D.; Froese, D.G.; Zazula, G.; Cassatt-Johnstone, M.; Dalen, L.; Der Sarkissian, C.; Dunn, S.G.; Ermini, L.; Gamba, C.; et al. Ancient horse genomes reveal the timing and extent of dispersals across the Bering Land Bridge. Mol. Ecol. 2021, 23, 6144–6161. [Google Scholar] [CrossRef]
- Leidy, J. Description of Vertebrate Fossils. In Post-Pleiocene Fossils of South-Carolina; Holmes, F.S., Ed.; Russel and Jones: Charleston, SC, USA, 1860; pp. 99–122. [Google Scholar]
- Azzaroli, A. A Synopsis of the Quaternary species of Equus in North America. Boll. Soc. Paleont. Ital. 1995, 34, 205–221. [Google Scholar]
- Quinn, J.H. Pleistocene Equidae of Texas. Econ. Geol. Rep. Investig. 1957, 33, 1–51. [Google Scholar]
- von Reichenau, W. Beitrage zur naheren Kenntnis fossiler Pferde aus deutschem Pleistozan, insbesondere über die Entwicklung und die Abkaustadiem del Gebisses von Hochterrassenpferde (Equus mosbachensis v. R.). Abh. Gross. Hess. Geolog. Darmstadt 1915, 7, 1–55. [Google Scholar]
- Alberdi, M.T.; Arroyo-Cabrales, J.; Marin-Leyva, A.H.; Polaco, O.J. Study of Cedral Horses and their place in the Mexican Quaternary. Rev. Mex. Cienc. Geológicas 2014, 31, 221–237. [Google Scholar]
- Kurten, B.; Anderson, E. Pleistocene Mammals of North America; Columbia University Press: New York, NY, USA, 1980; pp. 1–442. [Google Scholar]
- Winans, M.C. A quantitative study of the North American fossil species of the genus Equus. Evol. Perissodactyls 1989, 72, 262–297. [Google Scholar]
- Sher, A.V. Mlekopitaiushchie i Stratigrafia Pleistotsena Krainego Severo-Vostoka SSSR i Severnoi Ameriki; Nauka: Moskva, Russia, 1971. [Google Scholar]
- Eisenmann, V. Sussemionus, a new subgenus of Equus (Perissodactyla, Mammalia). Comp. Rendus. Biol. 2010, 333, 235–240. [Google Scholar] [CrossRef] [PubMed]
- Eisenmann, V. The equids from Liventsovka and other localities of the Khaprovskii Faunal Complex, Russia: A revision. Geobios 2022, 70, 17–33. [Google Scholar] [CrossRef]
- Leidy, J. Bones and teeth of horses from California and Oregon. Proc. Acad. Nat. Sci. Phila. 1865, 17, 94. [Google Scholar]
- Merriam, J.C. Preliminary report on the horses of Rancho La Brea. Bull. Dep. Geol. Sci. 1913, 7, 397–418. [Google Scholar]
- Savage, D.E. Late Cenozoic vertebrates of the San Francisco Bay Region. Bull. Dep. Geol. Sci. 1951, 28, 215–314. [Google Scholar]
- Miller, W.E. Pleistocene vertebrates of the Los Angeles Basin and vicinity (exclusive of Rancho La Brea). Bull. Los Angel. Cty. Mus. 1971, 10, 1–124. [Google Scholar]
- Brown, K.E.; Akersten, W.A.; Scott, E. Equus occidentalis Leidy from “Asphalto”, Kern County, California. In La Brea and Beyond: The Paleontology of Asphalt-Preserved Biotas; Harris, J.M., Ed.; Natural History Museum of Los Angeles County: Los Angeles, CA, USA, 2015; pp. 81–89. [Google Scholar]
- Barrón-Ortiz, C.I.; Rodrigues, A.T.; Theodor, J.M.; Kooyman, B.P.; Yang, D.Y.; Speller, C.F. Cheek tooth morphology and ancient mitochondrial DNA of late pleistocene horses from the western interior of North America: Implications for the taxonomy of North American Late Pleistocene. PLoS ONE 2017, 12, e0183045. [Google Scholar] [CrossRef]
- Marín-Leyva, A.H.; Arroyo-Cabrales, J.; García-Zepeda, M.L.; Ponce-Saavedra, J.; Schaaf, P.; Pérez-Crespo, V.P.; Pedro Morales-Puente, P.; Cienfuegos-Alvarado, E.; Alberdi, M.T. Feeding ecology and habitat of Late Pleistocene Equus horses fromwest-central Mexico using carbon and oxygen isotopes variation. Rev. Mex. Cienc. Geol. 2016, 33, 157–169. [Google Scholar]
- Jimenez-Hidalgo, E.; Diaz-Sibaja, R. Was Equus cedralensis a non-stilt legged horse? Taxonomical implication for the Mexican Pleistocene horses. Ameghiniana 2020, 57, 284–288. [Google Scholar] [CrossRef]
- Barrón-Ortiz, C. The Late Pleistocene Extinction in North America: An Investigation of Horse and Bison Fossil Material and Its Implication for Nutritional Extinction Models. Ph.D. Thesis, University of Calgary, Calgary, AB, Canada, 2016. [Google Scholar] [CrossRef]
- Prado, J.L.; Alberdi, M.T. Fossil Horses of South America; Springer: Berlin/Heidelberg, Germany, 2017; pp. 1–150. [Google Scholar]
- Machado, H.; Grillo, O.; Scott, E.; Avilla, L. Following the footsteps of the South American Equus: Are autopodia taxonomically informative? J. Mammal. Evol. 2018, 25, 397–405. [Google Scholar] [CrossRef]
- Machado, H.; Avilla, L. The Diversity of South American Equus. Did Size Really Matter? Front. Ecol. Evol. 2019, 7, 235. [Google Scholar] [CrossRef]
- Lund, P.W. Nouvelles Recherches sur la Faune fossile du Brésil. Ann. Sci. Nat. 1840, 13, 310–319. [Google Scholar]
- MacFadden, B.J.; Siles, O.; Zeitler, P.; Johnson, N.M.; Campbell, K.E. Magnetic polarity stratigraphy of the middle Pleistocene (Ensenadan) Tarija Formation of southern Bolivia. Quat. Res. 1983, 19, 172–187. [Google Scholar] [CrossRef]
- MacFadden, B.J.; Zeitler, P.K.; Anaya, F.; Cottle, J.M. Confirmation of the middle Pleistocene age of the sedimentary sequence from Tarija, Bolivia. Quat. Res. 2013, 79, 268–273. [Google Scholar] [CrossRef]
- Prado, J.L.; Martinez-Maza, C.; Alberdi, M.T. Megafauna extinction in South America: A new chronology for theArgentine Pampas. Palaeogeogr. Palaeoclim. Palaeoecol. 2015, 425, 41–49. [Google Scholar] [CrossRef]
- Roth, S. El mamífero misterioso de la Patagonia Grypotherium domesticum. II. Descripción de los restos encontrados en la caverna de Última Esperanza. Rev. Mus. Plata 1899, 9, 421–453. [Google Scholar]
- Labarca, R.; Caro, F.J.; Villavicencio, N.A.; Capriles, J.M.; Briones, E.; Latorre, C.; Santoro, C.M. A Partially Complete Skeleton of Hippidion saldiasi Roth, 1899 (Mammalia: Perissodactyla) from the Late Pleistocene of the High Andes in Northern Chile. J. Vertebr. Paleontol. 2021, 40, e1862132. [Google Scholar] [CrossRef]
- Paunero, R.S.; Rosales, G.; Prado, J.L.; Alberdi, M.T. Cerro bombero: Registro de hippidion saldiasi roth, 1899 (equidae, perissodactyla) en el holoceno temprano de Patagonia (Santa Cruz, Argentina). Estud. Geol. 2008, 64, 89–98. [Google Scholar] [CrossRef][Green Version]
- Massone, M.; Prieto, A. Evaluacion de la modalidad cultural Fell 1 en Magallanes. Chungara 2004, 1, 303–315. [Google Scholar] [CrossRef]
- Lund, P.W. Meddelelse af det ubbytte de i 1844 undersögte Knoglehuler have avgivet til Kundskaben om Brasiliens Dyreverden för sidste Jordomvaeltning. Ebenda 1846, 12, 1–94. [Google Scholar]
- Montané, J. Paleo-Indian remains from Lagunade Tagua Tagua, Central Chile. Science 1968, 161, 1137–1138. [Google Scholar] [CrossRef] [PubMed]
- Gervais, P. Recherches sur les Mammiferes Fossiles de l’Amerique Meridionale; Bertrand, P., Ed.; Librarie-Editeur: Paris, France, 1855; pp. 1–63. [Google Scholar]
- Shockey, B.J.; Salas-Gismondi, R.; Baby, P.; Guyot, J.-L.; Baltazar, M.C.; Huamán, L.; Clack, A.; Stucchi, M.; Pujos, F.; Emerson, J.M.; et al. New Pleistocene Cave Faunas of the Andes of Central Perú: Radiocarbon Ages and the Survival of Low Latitude, Pleistocene DNA. Palaeontol. Electron. 2009, 12, 15. [Google Scholar]
- Villavicencio, N.A.; Werdelin, L. The casa del diablo cave (puno, peru) and the late pleistocene demise of megafauna in the andean altiplano. Quat. Sci. Rev. 2018, 195, 21–31. [Google Scholar] [CrossRef]
- Sun, B.; Deng, T. The Equus Datum and the Early Radiation of Equus in China. Front. Ecol. Evol. 2019, 7, 429. [Google Scholar] [CrossRef]
- Sun, B.; Liu, W.; Liu, J.; Liu, L.; Jin, C. Equus qiyngyangensis in Jinyan Cave and its paleozoographic significance. Quat. Int. 2020, 591, 35–46. [Google Scholar] [CrossRef]
- Teilhard de Chardin, P.; Young, C.C. Fossil mammals from the late Cenozoic of northern China. Palaeontol. Sin. 1931, 9, 1–67. [Google Scholar]
- Flynn, L.J.; Bernor, R.L. Late Tertiary mammals from the Mongolian People’s Republic. Am. Mus. Novit. 1987, 2872, 1–16. [Google Scholar]
- Wang, X.; Li, Q.; Xie, G.; Saylor, J.E.; Tseng, Z.J.; Takeuchi, G.T.; Deng, T.; Wang, Y.; Hou, S.; Liu, J.; et al. Mio-Pleistocene Zanda Basin biostratigraphy and geochronology, pre-Ice Age fauna, and mammalian evolution in western Himalaya. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2013, 374, 81–95. [Google Scholar] [CrossRef]
- Jukar, A.; Sun, B.; Bernor, R.L. The first occurrence of Plesiohipparion huangheense (Qiu, Huang & Guo, 1987) (Equidae, Hipparionini) from the late Pliocene of India. Boll. Soc. Paleo. Ital. 2018, 57, 125–132. [Google Scholar]
- Qiu, Z.X.; Qiu, Z.D.; Deng, T.; Li, C.K.; Zhang, Z.Q.; Wang, B.Y.; Wang, X. Neogene land mammal stages/ages of China. In Fossil Mammals of Asia: Neogene Biostratigraphy and Chronology; Wang, X., Flynn, L.J., Fortelius, M., Eds.; Columbia University Press: New York, NY, USA, 2013; pp. 29–90. [Google Scholar]
- Matsumoto, H. On Hipparion richthofeni Koken. Sci. Rep. Tohoku Univ. 1927, 10, 59–75. [Google Scholar]
- Qiu, Z.; Huang, W.; Guo, Z. The Chinese hipparionine fossils. Palaeontol. Sin. New Ser. 1987, 175, 1–250. [Google Scholar]
- Bernor, R.L.; Lipscomb, D. The systematic position of “Plesiohipparion” aff. huangheense (Equidae, Hipparionini) from Gülyazi, Turkey. Mitt. Bayer. Staatssamml. Paläontol. Hist. Geol. 1991, 31, 107–123. [Google Scholar]
- Bernor, R.L.; Sun, B.; Chen, Y. Plesiohipparion shanxiense n. sp. from the Early Pleistocene (Nihowanian) of E Shanxi, China. Boll. Soc. Paleontol. Ital. 2015, 54, 197–210. [Google Scholar]
- Sefve, I. Die Hipparionen Nord-Chinas. Palaeo. Sin. Ser. 1927, 4, 1–54. [Google Scholar]
- Liu, P.; Lovlie, R. Magnetostratigraphic age of Pleistocene loess/paleosol sections at Kehe, Shanxi. J. Stratigr. 2007, 31, 240–246. [Google Scholar]
- Qiu, Z.; Deng, T.; Banyue, W. Early Pleistocene mammalian fauna from Longdan, Dongxiang, Gansu, China. Palaeontol. Sin. New Ser. 2004, 191, 1–245. [Google Scholar]
- Teilhard de Chardin, P.; Piveteau, J. Les mammifères fossiles de Nihowan (Chine). Ann. Paléont. 1930, 19, l–134. [Google Scholar]
- Kuzmina, I.E. Horses of North Eurasia from the Pliocene till the Present Time; Vereschagin, N.K., Ed.; Russian academy of Sciences: Saint-Petersburg, Russia, 1997; Volume 273, pp. 1–224. [Google Scholar]
- Deng, T.; Xue, X. Chinese Fossil Horses of Equus and Their Environment; China Ocean Press: Beijing, China, 1999. [Google Scholar]
- Chow, M.; Liu, H. Fossil equine teeth from Shansi. Vert. PalAsiat. 1959, 1, 133–136. [Google Scholar]
- Ao, H.; An, Z.; Dekkers, M.J.; Li, Y.; Xiao, G.; Zhao, H.; Qiang, X. Pleistocene magnetochronology of the fauna and Paleolithic sites in the Nihewan Basin: Significance for environmental and hominin evolution in North China. Quat. Geochronol. 2013, 18, 78–92. [Google Scholar] [CrossRef][Green Version]
- Colbert, E. Pleistocene mammals from the Ma Kai Valley of northern Yunnan, China. Am. Mus. Novit. 1940, 1099, 1–10. [Google Scholar]
- Eisenmann, V. Nouvelles interpretations des restes d’équidés (Mammalia, Perissodactyla) de Nihowan (Pléistocène inférieur de la Chine du Nord): Equus teilhardi nov. sp. Geobios 1975, 8, 125–134. [Google Scholar] [CrossRef]
- Sun, B.; Deng, T.; Liu, Y. Early Pleistocene Equus (Equidae, Perissodactyla) from Andersson Loc. 32 in Qixian, Shanxi, China. Hist. Biol. 2017, 31, 211–222. [Google Scholar] [CrossRef]
- Eisenmann, V.; Deng, T. Equus qingyangensis (Equidae, Perissodactyla) of the Upper Pliocene of Bajiazui, China: Evidence for the North American origin of an Old World lineage distinct from E. stenonis. Quaternaire 2005, 2, 113–122. [Google Scholar]
- Sharapov, S. Kuruksaiskii Kompleks Pozdnepliotsenovykh Mlekopitajushchikh Afgano-Adzhikistanskoi Depressii; Izdatelstvo Donish: Dushanbe, Tajikistan, 1986; p. 270. [Google Scholar]
- Forsten, A.; Sharapov, S. Fossil equids (Mammalia, Equidae) from the Neogene and Pleistocene of Tadzhikistan. Geodiversitas 2000, 22, 293–314. [Google Scholar]
- Vangengeim, E.A.; Sotnikova, M.V.; Alekseeva, L.I.; Vislobokova, I.A.; Zhegallo, V.I.; Zazhigin, V.S.; Shevyreva, N.S. Biostratigrafiya Pozdnego Pliotsena—Rannego Pleistotsena Adzhikistana; Nuaka: Moscow, Russia, 1988; p. 125. [Google Scholar]
- Kuznetsova, T.V.; Zhegallo, V.I. Taxonomic Diversity of Equids from the Nalaikha Locality (Mongolia). In Proceedings of the International Conference on the Conditions of the Theriofauna in Russia and Adjacent Countries, Moscow, Russia, 1–3 February 1995; pp. 174–178. [Google Scholar]
- Kuznetsova, T.V.; Zhegallo, V.I. Equus (Hemionus) nalaikhaensis from the Pleistocene of Mongolia, the Earliest Kulan of Central Asia. Paleontol. J. 2009, 43, 574–583. [Google Scholar] [CrossRef]
- Lazarev, P.A. Anthropogenic Horses of Yakutia; Nauka: Moscow, Russia, 1980; p. 190. (In Russian) [Google Scholar]
- Rusanov, B.S. Biostratigraphy of Cenoizoic Deposits of Southern Yakutia; Nauka: Moscow, Russia, 1968; p. 459. [Google Scholar]
- Lazarev, P.A. Large mammals of Anthropogene in Yakutia; Nauka: Moscow, Russia, 2008; p. 160. (In Russian) [Google Scholar]
- Spasskaya, N.; Pavlinov, I.; Sharko, F.; Boulygina, E.; Tsygankova, S.; Nedoluzhko, A.; Mashchenko, E. Morphometric and genetic analyses of diversity of the Lena horse (Equus lenensis Russanov, 1968; Mammalia: Equidae). Russ. J. Theriol. 2021, 20, 82–95. [Google Scholar] [CrossRef]
- Liu, H.Y. A new species of Equus from Locality 21 of Zhoukoudian. Vert PalAsiat 1963, 7, 318–322. [Google Scholar]
- Forsten, A. Chinese fossil horses of the genus Equus. Acta Zool. Fenn. 1986, 181, 1–40. [Google Scholar]
- Gromova, V.I. O novoj iskopaiemoj loshadi iz srednej Azii. Dokl. Akad. Nauk SSSR 1946, 54, 349–351. [Google Scholar]
- Eisenmann, V.; Helmer, D.; Segui, M.S. The big Equus from Geometric Kebaran of Umm el Tlel, Syria: Equus valeriani, Equus capensis or Equus caballus? Proc. Fifth Int. Symp. Archaeozoology Southwest. Asia Adjac. Areas 2002, 1, 62–73. [Google Scholar]
- Zhou, X.; Sun, Y.; Xu, Q.; Li, Y. Note on a new Late Pleistocene Equus from Dalian. Vertebr. Palasiat. 1985, 23, 69–76. [Google Scholar]
- Yuan, J.; Hou, X.; Barlow, A.; Preick, M.; Taron, U.H.; Alberti, F.; Basler, N.; Deng, T.; Lai, X.; Hofreiter, M.; et al. Molecular identification of Late and terminal Pleistocene Equus ovodovi from northeastern China. PLoS ONE 2019, 14, e0216883. [Google Scholar] [CrossRef] [PubMed]
- Eisenmann, V.; Vasiliev, S. Unexpected finding of a new Equus species (Mammalia, Perissodactyla) belonging to a supposedly extinct subgenus in Late Pleistocene deposits of Khakassia (southwestern Siberia). Geodiversitas 2011, 33, 519–530. [Google Scholar] [CrossRef]
- Orlando, L.; Metcalf, J.L.; Alberdi, M.T.; Telles-Antunes, M.; Bonjean, D.; Otte, M.; Martin, F.; Eisenmann, V.; Mashkour, M.; Morello, F.; et al. Revising the recent evolutionary history of equids using ancient DNA. Proc. Natl. Acad. Sci. USA 2009, 106, 21754–21759. [Google Scholar] [CrossRef] [PubMed]
- Yuan, J.; Sheng, G.; Preick, M.; Sun, B.; Hou, X.; Chen, S.; Taron, U.H.; Barlow, A.; Wang, L.; Hu, J.; et al. Mitochondrial genomes of Late Pleistocene caballines horses from China belong to a separate clade. Quat. Sci. Rev. 2020, 250, 106691. [Google Scholar] [CrossRef]
- Plasteeva, N.; Vasiliev, S.; Kosintsev, P. Equus (Sussemionus) ovodovi Eisenmann et Vasiliev, 2011 from the Laye Pleistocene of Western Siberia. Russ. J. Theriol. 2015, 14, 187–200. [Google Scholar] [CrossRef]
- Pallas, P.S. Equus hemionus. Novi. Comment. Acad. Sci. Imp. Petropol. 1774, 19, 394–417. [Google Scholar]
- Biswas, S.K. Regional Tectonic Framework, Structure and Evolution of the Western Marginal Basins of India. Tectonophysics 1987, 135, 302–327. [Google Scholar]
- Costa, A.G. A new Late Pleistocene fauna from arid coastal India: Implications for inundated coastal refugia and human dispersals. Quat. Int. 2017, 436, 253–269. [Google Scholar] [CrossRef]
- Dassarma, D.C.; Biswas, S. Newly discovered Late Quaternary vertebrates from the terraced alluvial fills of the Son Valley. Indian J. Earth Sci. 1977, 4, 122–136. [Google Scholar]
- Turvey, S.T.; Sathe, V.; Crees, J.J.; Jukar, A.M.; Chakraborty, P.; Lister, A.M. Late Quaternary megafaunal extinctions in India: How much do we know? Quat. Sci. Rev. 2021, 252, 106740. [Google Scholar] [CrossRef]
- Stewart, M.J.; Louys, G.J.; Price, N.A.; Drake, H.S.; Petraglia, M.D. Middle and Late Pleistocene mammal fossils of Arabia and surrounding regions: Implications for biogeography and hominin dispersals. Quat. Int. 2019, 515, 12–29. [Google Scholar] [CrossRef]
- Moorcroft, W. In Ladakh and Kashmir; in Peshawar, Kabul, Kunduz, and Bokhara. In Travels in the Provinces of Hindustan and the Panjab; John Murray: London, UK, 1841; p. 312. [Google Scholar]
- Shah, N. Status and action plan for the kiang (Equus kiang). In Equids: Zebras, Asses and Horses. Status Survey and Conservation Action Plan; Moehlman, P.D., Ed.; IUCN: Gland, Switzerland, 2002; pp. 72–81. [Google Scholar]
- Wang, Y.X. A Complete Check-List of Mammal Species and Subspecies in China: A Taxonomic and Geographic Reference; China Forestry Publishing House: Beijing, China, 2002. [Google Scholar]
- Schaller, G.B. Wildlife of the Tibetan Steppe; University of Chicago Press: Chicago, IL, USA, 1998. [Google Scholar]
- Poliakov, I.S. Przewalski horse (Equus Przevalskii n. sp.). In Izvestija Russkogo Geograficheskogo Obshchestva; Russian Academy of Sciences: Saint Petersburg, Russia, 1881; Volume 17, pp. 1–19. (In Russian) [Google Scholar]
- Groves, C. Morphology, habitat, and taxonomy. In Przewalski’s Horse: The History and Biology of an Endangered Species; Boyd, L., Houpt, K.A., Eds.; State University of New York Press: Albany, NY, USA, 1994; pp. 39–59. [Google Scholar]
- Groves, C.; Grubb, P. Ungulate Taxonomy; Johns Hopkins University Press: Baltimore, MD, USA, 2011. [Google Scholar]
- Heptner, V.; Nasimovich, A.A.; Bannikov, A.G. Mammals of the Soviet Union. In Artiodactyla and Perissodactyla; Vysshaya Shkola: Moscow, Russia, 1961; Volume 1. [Google Scholar]
- Deng, T. Ages of some Late Pleistocene faunas based on the presence of Equus przewalskii (Perissodactyla, Equidae). Vertebr. Palasiat. 1999, 23, 51–56. [Google Scholar]
- Gaunitz, C.; Fages, A.; Hanghøj, K.; Albrechtsen, A.; Khan, N.; Schubert, M.; Seguin-Orlando, A.; Owens, I.J.; Felkel, S.; Bignon-Lau, O.; et al. Ancient genomes revisit the ancestry of domestic and Przewalski’s horses. Science 2018, 360, 111–114. [Google Scholar] [CrossRef]
- Librado, P.; Orlando, L. Genomics and Evolutionary History of Equids. Annu. Rev. Anim. Biosci. 2020, 7, 16–41. [Google Scholar] [CrossRef]
- Taylor, W.T.T.; Barrón-Ortiz, C.I. Rethinking the evidence for early horse domestication at Botai. Sci. Rep. 2021, 11, 7440. [Google Scholar] [CrossRef]
- Wolf, D.; Bernor, R.L.; Hussain, S.T. A systematic, biostratigraphic, and paleobiogeographic reevaluation of the Siwalik hipparionine horse assemblage from the Potwar Plateau, Northern Pakistan. Palaeontogr. Abt. 2013, 300, 1–115. [Google Scholar] [CrossRef]
- Jukar, A.M.; Sun, B.; Nanda, A.C.; Bernor, R.L. The first occurrence of Eurygnathohippus (Mammalia, Perissodatyla, Equidae) outside Africa and its biogeographic significance. Boll. Soc. Paleo. Ital. 2019, 58, 171–179. [Google Scholar]
- Rook, L.; Bernor, R.L.; Avilla, L.D.; Cirilli, O.; Flynn, L.; Jukar, A.M.; Sanders, W.; Scott, E.; Wang, X. Mammal Biochronology (Land Mammal Ages) around the World from the Late Miocene to Middle Pleistocene and Major Events in Horse Evolutionary History. Front. Ecol. Evol. 2019, 7, 278. [Google Scholar] [CrossRef]
- Jukar, A.M.; Lyons, S.K.; Wagner, P.J.; Uhen, M.D. Late Quaternary extinctions in the Indian Subcontinent. Paleogeol. Paleoclim. Paleoecol. 2021, 562, 110137. [Google Scholar] [CrossRef]
- Thomas, P.K.; Joglekar, P. Holocene Faunal Studies. Man Environ. 1994, 19, 179–203. [Google Scholar] [CrossRef]
- Falconer, H.; Cautley, P.T. Illustrations-Part IX Equidae, Camelidae, Sivatherium. In Fauna Antiqua Sivalensis, Being the Fossil Zoology of the Sewalik Hills in the North of India; Smith, Elder and Co.: London, UK, 1849. [Google Scholar]
- Hussain, S.T. Revision of Hipparion (Equidae, Mammalia) from the Siwalik Hills of Pakistan and India. Geol. Mijnb. 1971, 147, 80–81. [Google Scholar]
- MacFadden, B.J.; Woodburne, M.O. Systematics of the Neogene Siwalik Hipparions (Mammalia, Equidae) based on cranial and dental morphology. J. Vert. Paleo 1982, 2, 185–218. [Google Scholar] [CrossRef]
- Bernor, R.L.; Hussain, S.T. An Assessment of the Systematic, Phylogenetic and Biogeographic Relationships of Siwalik Hipparionine horses. J. Vert. Paleo 1985, 5, 32–87. [Google Scholar] [CrossRef]
- Lydekker, R. Containing the order Ungulata, suborders Perissodactyla, Toxodontia, Condylartha, and Ambylopoda. In Catalogue of the Fossil Mammalia in the British Museum (Natural History) Part III; Taylor Francis: London, UK, 1886. [Google Scholar]
- Bernor, R.L.; Cirilli, O.; Jukar, A.M.; Potts, R.; Buskianidze, M.; Rook, L. Evolution of Early Equus in Italy, Georgia, the Indian Subcontinent, East Africa, and the Origins of African Zebras. Front. Ecol. Evol. 2019, 7, 166. [Google Scholar] [CrossRef]
- Patel, R.; Nainwal, H.C.; Sharma, T.; Rana, R.S. National Workshop on Indian Siwalik: Recent Advances and Future Research. In Equus cf. sivalensis from the Tatrot Formation (Upper Pliocene) of Jhil-Bankabara Area Sirmaur District, Himachal Pradesh, India; Geological Survey of India: Lucknow, India, 2017; pp. 51–53. [Google Scholar]
- Moigne, A.M.; Dambricourt Malassé, A.; Singh, M.; Kaur, A.; Gaillard, C.; Karir, B.; Pal, S.; Bhardwaj, V.; Abdessadok, S.; Chapon Sao, C.; et al. The faunal assemblage of the paleonto-archeological localities of the Late Pliocene Quranwala Zone, Mosol Formation, Siwalik Range, NW India. Palevol 2016, 15, 359–378. [Google Scholar] [CrossRef]
- Gaur, R.; Chopra, S.R.K. Taphonomy, fauna, environment and ecology of Upper Sivaliks (Plio-Pleistocene) near Chandigarh, India. Nature 1984, 308, 353–355. [Google Scholar] [CrossRef]
- Hussain, S.T.; van den Bergh, G.; Steensma, K.J.; de Visser, J.A.; de Vos, J.; Mohammad, A.; van Dam, J.; Sondaar, P.; Malik, S.B. Biostratigraphy of the Plio-Pleistocene continental sediments (Upper Siwaliks) of the Mangla-Samwal anticline, Azad Kashmir, Pakistan. Proc. K. Ned. Akad. Wet. 1992, 95, 65–80. [Google Scholar]
- Dennell, R.; Coard, R.; Turner, A. The biostratigraphy and magnetic polarity zonation of the Pabbi Hills, northern Pakistan: An Upper Siwalik (Pinjor Stage) Upper Pliocene–Lower Pleistocene fluvial sequence. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2006, 234, 168–185. [Google Scholar] [CrossRef]
- Sonakia, A.; Biswas, S. Fossil mammals including early man from the Quaternary deposits of the Narmada and Son basins of Madhya Pradesh, India. Palaeontol. Indica 2011, 53, 1–84. [Google Scholar]
- Lydekker, R. Siwalik and Narbada Equidae. Mem. Geol. Soc. India Palaeontol. Indica Ser. 1882, 2, 67–98. [Google Scholar]
- Cirilli, O.; Bernor, R.L.; Rook, L. New insights on the Early Pleistocene equids from Roca-Neyra (France, central Europe): Implications for the Hipparion LAD and the Equus FAD in Europe. J. Paleontol. 2021, 95, 406–425. [Google Scholar] [CrossRef]
- Gromova, V. Le Genre Hipparion; Bureau de Recherches Geologiques et Minieres: Orléans, France, 1952; Volume 12, pp. 1–288. [Google Scholar]
- Bernor, R.L.; Sen, S. The Early Pliocene Plesiohipparion and Proboscidipparion (Equidae, Hipparionini) from Çalta, Turkey (Ruscinian Age, c. 4.0 Ma). Geodiversitas 2017, 39, 285–314. [Google Scholar] [CrossRef]
- Bernor, R.L.; Beech, M.; Bibi, F. Equidae from the Baynunah Formation. In Sands of Time; Springer: Berlin/Heidelberg, Germany, 2022; pp. 259–280. [Google Scholar]
- Crusafont, P.M.; Sondaar, P. Una nouvelle espece d’Hipparion du Pliocéne terminal d’Espagne. Palaeovertebrata 1971, 4, 59–66. [Google Scholar] [CrossRef][Green Version]
- Alberdi, M.T.; Alcalà, L. A study of the new samples of the Pliocene Hipparion (Equidae, Mammalia) from Spain and Bulgaria. Trans. R. Soc. Edinb. Earth Sci. 1999, 89, 167–186. [Google Scholar] [CrossRef]
- Piñero, P.; Agustí, J.; Oms, O.; Fierro, I.; Montoya, P.; Mansino, S.; Ruiz-Sánchez, F.; Alba, D.M.; Alberdi, M.T.; Blain, H.A.; et al. Early Pliocene continental vertebrate Fauna at Puerto de la Cadena (SE Spain) and its bearing on the marine-continental correlation of the Late Neogene of Eastern Betics. Paleogeogr. Palaeoclim. Palaeoecol. 2017, 479, 102–114. [Google Scholar] [CrossRef]
- Gervais, P. Zoologie et Paléontologie Française (Animaux Vertébrés): Deuxième Edition; Arthus Bertrand: Paris, France, 1859; pp. 1–544. [Google Scholar]
- Depéret, C. Les animaux pliocènes du Roussilon. Mem. Soc. Geol. Fr. 1890, 3, 1–198. [Google Scholar]
- Forsten, A. The hipparions (Mammalia, Equidae) of Suffolk, England. Trans. R. Soc. Edinb. Earth Sci. 2001, 92, 115–120. [Google Scholar] [CrossRef]
- Forsten, A. Latest Hipparion Cristol, 1832 in Europe. A review of the Pliocene Hipparion crassum Gervais Group and other finds (Mammalia, Equidae). Geodiversitas 2002, 24, 465–486. [Google Scholar]
- Eisenmann, V.; Sondaar, P. Pliocene vertebrate locality of Calta, Ankara, Turkey. Hipparion. Geodiversitas 1998, 20, 409–439. [Google Scholar]
- Hernández-Pacheco, E. La llanura manchega y sus mamíferos fósiles: Yacimiento de La Puebla de Almoradier. Com. Investig. Pleontol. Mem. 1921, 28, 1–41. [Google Scholar]
- Alberdi, M.T. El género Hipparion en España, revision e historia evolutive. Trab. Sobre Neógeno Cuatern. 1974, 1, 1–139. [Google Scholar]
- Bernor, R.L.; Koufos, G.D.; Woodburne, M.O.; Fortelius, M. The evolutionary history and biochronology of European and southwestern Asian late Miocene and Pliocene hipparionine horses. In The Evolution of Western Eurasian Neogene Mammal Faunas; Bernor, R.L., Fahlbusch, V., Mittman, H.W., Eds.; Columbia University Press: New York, NY, USA, 1996; pp. 307–338. [Google Scholar]
- Bernor, R.L.; Sun, B.Y. Morphology through ontogeny of Chinese Proboscidipparion and Plesiohipparion and observations on their Eurasian and African relatives. Vertebr. Palasiat. 2015, 53, 73–92. [Google Scholar]
- Agustì, J.; Oms, O. On the age of the last hipparionine faunas in western Europe. Comptes Rendus Acad. Sci. Paris Sci. 2001, 332, 291–297. [Google Scholar] [CrossRef]
- Azanza, B.; Morales, M.; Andrés, M.; Cerdeño, E.; Alberdi, M.T. Los macromamíferos y la datación del yacimiento clásico de Villarroya. In Villarroya, Yacimiento Clave de la Paleontologìa Riojana; Alberdi, M.T., Azanza, B., Cervante, E., Eds.; Instituto de Estudios Riojanos: La Rioja, Spain, 2016; pp. 267–280. [Google Scholar]
- Athanassiou, A. A Villafranchian Hipparion-bearing mammal fauna from Sésklo (E. Thessaly, Greece): Implications for the question of Hipparion-Equus sympatry in Europe. Quaternary 2018, 12, 12. [Google Scholar] [CrossRef]
- Rook, L.; Cirilli, O.; Bernor, R.L. A late occurring “Hipparion” from the middle Villafranchian of Montopoli, Italy (Early Pleistocene; MN16b; ca. 2.5 Ma). Boll. Soc. Paleontol. Ital. 2017, 56, 333–339. [Google Scholar]
- Bajgusheava, V.S. Krupnaja Loshad Khaprovskogo Kompleksa is alluvija Severo Vostochnoto Priazovija. Invesija Servo Kavk. Nauchnogo Zentra Vychei Shkoly 1978, 1, 98–102. [Google Scholar]
- Gromova, V.I. Istorija loshadej (roda Equus) v Starom Svete. Chast’ 1. Obzor i opisanie form. Tr. Paleontol. Inst. 1949, 17, 1–373. [Google Scholar]
- Forsten, A. The fossil horses (Equidae, Mammalia) from the Plio–Pleistocene of Liventsovka near Rostov–Don, Russia. Geobios 1997, 31, 645–657. [Google Scholar] [CrossRef]
- Alberdi, M.T.; Cerdeño, E.; López–Martínez, N.; Morales, J.; Soria, M.D. La Fauna Villafranquiense de El Rincon–1 (Albacete, Castilla—La Mancha). Estud. Geol. 1997, 53, 69–93. [Google Scholar] [CrossRef]
- Alberdi, M.T.; Ortiz–Jaureguizar, E.; Prado, J.L. A quantitative review of European stenonoid horses. J. Paleontol. 1998, 72, 371–387. [Google Scholar] [CrossRef]
- Azzaroli, A. On Equus livenzovensis Baigusheva 1978 and the “stenonid” lineage of Equids. Paleontogr. Ital. 2000, 87, 1–17. [Google Scholar]
- Bernor, R.L.; Cirilli, O.; Wang, S.Q.; Rook, L. Equus cf. livenzovenzis from Montopoli, Italy (early Pleistocene; MN16b; ca. 2.6 Ma). Boll. Soc. Paleontol. Ital. 2018, 57, 203–216. [Google Scholar]
- Cherin, M.; Cirilli, O.; Azzarà, B.; Bernor, R.L. Equus stenonis (Equidae, Mammalia) from the Early Pleistocene of Pantalla (Italy) and the dispersion of stenonines horses in Europe. Boll. Soc. Paleontol. Ital. 2021, 60, 1–18. [Google Scholar]
- Terhune, C.; Curran, S.; Croitor, R.; Drăguşin, V.; Gaudin, T.; Petrulescu, A.; Robinson, C.; Robu, M.; Werdelin, L. Early Pleistocene fauna of the Olteţ Valley of Romania: Biochronological and biogeographical implications. Quat. Int. 2020, 553, 14–33. [Google Scholar] [CrossRef]
- Delafond, F.; Depéret, C. Etude des Gites Minéraux de la France. In Les Terrains Tertiaires de la Bresse et Leurs Gites de Lignites et de Minerais de Fer; Imprimerie National: Paris, France, 1893; pp. 1–332. [Google Scholar]
- Pomel, A. Catalogue Méthodique et Descriptif des Vertebres Fossiles Découverts dans le Bassin Hydrographique Supérieur de la Loire et Surtout Dans la Vallée de Son Affluent Principal; L’Allirée Chez, J., Baillière, B., Eds.; Hachette: Paris, France, 1853; pp. 1–123. [Google Scholar]
- Boule, M. Bulletin de la Société Géologique de France; BSGF—Earth Sciences Bulletin: Vienne, France, 1891; Volume 18, p. 801. [Google Scholar]
- Viret, J. Le loess à bancs durcis de Saint Vallier (Drome) et sa faune de mammifères villagranchiens. Nouv. Arch. Mus. D’histoire Nat. Lyon 1954, 4, 1–200. [Google Scholar]
- Samson, P. Les équidés fossiles de Roumanie (Pliocène moyen—Pléistocène supérieur). Geol. Romana 1975, 14, 165–354. [Google Scholar]
- Cocchi, I. L’uomo fossile dell’Italia Centrale. Mem. Della Soc. Ital. Sci. Nat. 1867, 2, 1–180. [Google Scholar]
- Prat, F. Les equidés villafranchiens en France, genre Equus. Cah. Quat. 1980, 2, 1–290. [Google Scholar]
- Palombo, M.R.; Alberdi, M.T.; Bellucci, L.; Sardella, R. An intriguing middle–sized horse from Coste San Giacomo (Anagni Basin, central Italy). Quat. Res. 2017, 87, 347–362. [Google Scholar] [CrossRef]
- Cirilli, O. Equus stehlini Azzaroli, 1964 (Perissodactyla, Equidae). A revision of the most enigmatic horse from the Early Pleistocene of Europe, with new insights on the evolutionary history of European medium- and small-sized horses. Riv. Paleont. Strat. 2022, 128, 241–265. [Google Scholar] [CrossRef]
- Azzaroli, A. The two Villafranchian Horses of the Upper Valdarno. Palaeontogr. Ital. 1964, 59, 1–12. [Google Scholar]
- Alberdi, M.T.; Palombo, M.R. The late Early to early Middle Pleistocene stenonoid horses from Italy. Quat. Int. 2013, 288, 25–44. [Google Scholar] [CrossRef]
- Palombo, M.R.; Alberdi, M.T. Light and shadows in the evolution of South European stenonoid horses. Foss. Impr. 2017, 73, 115–140. [Google Scholar] [CrossRef]
- Rook, L.; Martínez–Navarro, B. Villafranchian: The long story of a Plio–Pleistocene European large mammal biochronologic unit. Quat. Int. 2010, 219, 134–144. [Google Scholar] [CrossRef]
- Bartolini-Lucenti, S.; Cirilli, O.; Pandolfi, L.; Bernor, R.L.; Bukhsianidze, M.; Carotenuto, F.; Lordkipanidze, D.; Tsikaridze, N.; Rook, L. Zoogeographic significance of Dmanisi large mammal assemblage. J. Hum. Evol. 2022, 163, 103125. [Google Scholar] [CrossRef]
- Vekua, A.K. The Lower Pleistocene Mammalian Fauna of Akhalkalaki. Tbilisi. 1962, pp. 1–191. (In Georgian). Available online: http://www.rhinoresourcecenter.com/index.php?s=1&act=pdfviewer&id=1435388432&folder=143 (accessed on 16 July 2022).
- Koufos, G.D. Early Pleistocene equids from Mygdonia basin (Macedonia, Greece). Palaeontogr. Ital. 1992, 79, 167–199. [Google Scholar]
- Eisenmann, V. Equus granatensis of Venta Micena and evidence for primitive non–stenonid horses in the Lower Pleistocene. In The Hominids and Their Environment during the Lower and Middle Pleistocene of Eurasia; Gilbert, J., Sanchez, F., Gilbert, L., Ribot, F., Eds.; Museo de Prehistoria y Paleontología: Orce, Spain, 1995; pp. 175–189. [Google Scholar]
- Koufos, G.D.; Vlachou, T.; Gkeme, A. The Fossil Record of Equids (Mammalia: Perissodactyla: Equidae) in Greece. In Fossil Vertebrates of Greece; Vlachos, E., Ed.; Springer: Berlin/Heidelberg, Germany, 2021; Volume 2, pp. 351–401. [Google Scholar]
- Guerrero-Alba, S.; Palmqvist, P. Estudio morfometrico del caballo de Venta Micena (Orce, Granada) y su comparacion con los equidos modernos y del Plio- Pleistoceno en el viejo mundo. Paleontol. Evol. 1997, 30, 93–148. [Google Scholar]
- Belmaker, M. Early Pleistocene faunal connections between Africa and Eurasia: An ecological perspective. In Out of Africa I: The First Hominin Colonization of Eurasia; Fleagle, J.G., Shea, J.J., Grine, F.E., Baden, A.L., Leakey, R.E., Eds.; Springer: Berlin/Heidelberg, Germany, 2010; pp. 183–205. [Google Scholar]
- Wüst, E. Untersuchungen über das Pliozan und das Alteste Pleistozan Thüringens, nordlich vom Thüringer Walde und westlich von der Saale. Abh. Nat. Ges. Halle 1900, 23, 1–352. [Google Scholar]
- Vekua, A.K. The Lower Pleistocene mammalian fauna of Akhalkalaki (Southern Georgia, USSR). Palaeontogr. Ital. 1986, 74, 63–96. [Google Scholar]
- Musil, R. Die Pferdefunde der lokalität Stránská Skála. Anthropos 1972, 20, 185–192. [Google Scholar]
- Boulbes, N.; Van Asperen, E.N. Biostratigraphy and palaeoecology of European Equus. Front. Ecol. Evol. 2019, 7, 301. [Google Scholar] [CrossRef]
- Alberdi, M.T. Estudio de los caballos de los yacimientos de Fuente Nueva–3 y Barranco León–5 (Granada). In Ocupaciones Humanas en el Pleistoceno Inferior Y Medio de la Cuenca de Guadix–Baza; Toro, I., Martínez–Navarro, B., Agustí, J., Eds.; Arqueología Monografías, Junta de Andalucía, Consejería de Cultura: Andalucía, Spain, 2010; pp. 291–306. [Google Scholar]
- Koufos, G.D.; Kostopoulos, D.S.; Sylvestrou, I.A. Equus apolloniensis n. sp. (Mammalia, Equidae) from the latest Villafranchian locality of Apollonia, macedonia, Greece. Paleontol. Evol. 1997, 30, 49–76. [Google Scholar]
- Gkeme, A.G.; Koufos, G.D.; Kostopoulos, D.S. Reconsidering the Equids from the Early Pleistocene Fauna of Apollonia 1 (Mygdonia Basin, Greece). Quaternary 2021, 4, 12. [Google Scholar] [CrossRef]
- Eisenmann, V.; Boulbes, N. New results on equids from the Early Pleistocene site of Untermassfeld. In The Pleistocene of Untermassfeld Near Meiningen (Thütingen, Germany); Kahlke, R.D., Ed.; Romisch Germanischen Zentralmuseums: Weimar, Germany, 2020; pp. 1295–1321. [Google Scholar]
- Musil, R. Die Equiden–Reste aus dem Unterpleistozän von Untermassfeld. In Das Pleistozän von Untermassfeld bei Meiningen (Thüringen); Kahlke, R.D., Ed.; Monographien, Römisch–Germanisches Zentralmuseum, Forschungsinstitut für vor und Frühgeschichte: Liebniz, Germany, 2001; pp. 557–587. [Google Scholar]
- Forsten, A. A review of Equus stenonis Cocchi (Perissodactyla, Equidae) and related forms. Quat. Sci. Rev. 2000, 18, 1373–1408. [Google Scholar] [CrossRef]
- Lister, A.M.; Parfitt, S.A.; Owen, F.J.; Collinge, S.E.; Breda, M. Metric analysis of ungulate mammals in the early Middle Pleistocene of Britain, in relation to taxonomy and biostratigraphy: II: Cervidae, Equidae and Suidae. Quat. Int. 2010, 228, 157–179. [Google Scholar] [CrossRef]
- Tsoukala, E. Contribution to the Study of the Pleistocene Fauna of Large Mammals (Carnivora, Perissodactyla, Artiodactyla) from Petralona Cave Chalkidiki (N. Greece). Ph.D. Thesis, Aristotle University of Thessaloniki, Thessaloniki, Serres, 1989. [Google Scholar]
- Azzaroli, A. On a Late Pleistocene ass from Tuscany with notes on thehistory of ass. Palaeontol. Ital. 1979, 71, 27–47. [Google Scholar]
- Azzaroli, A. Pleistocene and living Horses of the old World. Palaeontogr. Ital. 1966, 61, 1–15. [Google Scholar]
- Caloi, L. Il genere Equus nell’Italia centrale. Studi Geol. Camerti 1995, 469–486. [Google Scholar]
- Eisenmann, V. Pliocene and Pleistocene Equids: Palaeontology versus molecular biology. Cour. Forsch. Senckenberg 2006, 256, 71–89. [Google Scholar]
- Regalia, E. Sull’Equus (Asinus) hydruntinus Regalia della grotta di Romanelli (Castro, Lecce). Arch. Antropol. Etnol. 1907, 37, 375–390. [Google Scholar]
- Vishnyatsky, L.B. The Palaeolithic of Central Asia. J. World Prehistory 1999, 13, 69–122. [Google Scholar] [CrossRef]
- Musil, R. Die Equiden–Reste aus dem Pleistozän von Süssenborn bei Weimar. Paläontologische Abh. Paläozool. 1969, 3, 617–666. [Google Scholar]
- Azzaroli, A. Ascent and decline of monodactyl equids: A case for prehistoric overkill. Ann. Zool. Fenn. 1992, 28, 151–163. [Google Scholar]
- Bennett, E.A.; Champlot, S.; Peters, J.; Arbuckle, B.S.; Guimaraes, S.; Pruvost, M.; Bar-David, S.; Davis, S.J.M.; Gautier, M.; Kaczensky, P.; et al. Taming the late Quaternary phylogeography of the Eurasiatic wild ass through ancient and modern DNA. PLoS ONE 2017, 12, e0174216. [Google Scholar]
- Aurell-Garrido, J.; Madurell-Malapeira, J.; Alba, D.M.; Moyà-Solà, S. The stenonian and caballoid equids from the Pleistocene section of Vallparadís (Terrassa, Catalonia, Spain). Cidaris 2010, 30, 61–66. [Google Scholar]
- Madurell-Malapeira, J.; Minwer-Barakat, R.; Alba, D.M.; Garcés, M.; Gómez, M.; Aurell-Garrido, J.; Ros-Montoya, S.; Moya-Sola, S.; Berastegui, X. The Vallparadís section (Terrassa, Iberian peninsula) and the latest Villafranchian faunas of Europe. Quat. Sci. Rev. 2010, 29, 2972–2982. [Google Scholar] [CrossRef]
- Boddaert, P. Elenchus Animalium, Volumen 1: Sistens Quadrupedia Huc Usque Nota, Erorumque Varietates; C. R. Hake: Rotterdam, The Netherlands, 1785; p. 174. [Google Scholar]
- Eisenmann, V.; Kuznetsova, T.V. Early Pleistocene equids (Mammalia, Perissodactyla) of Nalaikha (Mongolia) and the emergence of modern Equus Linnaeus, 1758. Geodiversitas 2004, 26, 535–561. [Google Scholar]
- van Asperen, E.N. Implications of age variation and sexual dimorphism in modern equids forMiddle Pleistocene equid taxonomy. Int. J. Osteoarchaeol. 2013, 23, 1–12. [Google Scholar] [CrossRef]
- van Asperen, E.N. Ecomorphological adaptations to climate and substrate in late middle Pleistocene caballoid horses. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2010, 297, 584–596. [Google Scholar] [CrossRef]
- Churcher, C.S.; Richardson, M.L. Equidae. In Evolution of African Mammals; Maglio, V.J., Cooke, H.S.B., Eds.; Harvard University Press: Cambridge, MA, USA, 1978; pp. 379–422. [Google Scholar]
- Bernor, R.L.; Armour-Chelu, M.; Gilbert, H.; Kaiser, T.; Schulz, E. Equidae. In Cenozoic Mammals of Africa; Werdelin, L., Sanders, W., Eds.; University of California Press: Berkeley, CA, USA, 2010; pp. 685–721. [Google Scholar]
- Duval, M.; Sahnouni, M.; Parés, J.P.; van der Made, J.; Abdessadok, S.; Harichane, Z.; Cheheb, R.C.; Boulaghraif, K.; Pérez-Gonzalez, A. The Plio-Pleistocene sequesnce of Oued Boucherti (Algeria): A unique chronologically-constrained archeological and paleontological record in North Africa. Quat. Sci. Rev. 2021, 271, 107116. [Google Scholar] [CrossRef]
- Sahnouni, M.; Pares, J.M.; Duval, M.; Caceres, I.; Harichane, Z.; Van der Made, J.; Perez-Gonzalez, A.; Abdessadok, S.; Kandi, N.; Derradji, A.; et al. 1.9-million- and 2.4 million-year-old artifacts and stone toolcutmarked bones from Aïn Boucherit, Algeria. Science 2017, 362, 1297–1307. [Google Scholar] [CrossRef] [PubMed]
- Villalta, J.F.; Crusafont, M. Les gisements de mammifères du Néogène espagnol. Compt. Rendus Soc. Geol. Fr. 1957, 1, 9–10. [Google Scholar]
- Cirilli, O.; Zouhri, S.; Boughabi, S.; Benvenuti, M.; Bernor, R.L.; Papini, M.; Rook, L. The hipparionine horses (Perissodactyla: Mammalia) from the Late Miocene of Tizi N’Tadderht (southern Ouarzazate basin; Central High Atlas; Morocco). Riv. Paleontol. Strat. 2020, 126, 1–12. [Google Scholar]
- Bernor, R.L.; Boaz, N.T.; Cirilli, O.; El-Shawaihdi, M.; Rook, L. Sahabi Eurygnathohippus feibeli; its systematic, stratigraphic, chronologic and biogeographic contexts. Riv. Paleont. Strat. 2020, 126, 561–581. [Google Scholar]
- Bernor, R.L.; Harris, J. Systematics and Evolutionary Biology of the Late Miocene and Early Pliocene Hipparionine Horses from Lothagam, Kenya. In Lothagam: The Dawn of Humanity in Eastern Africa; Leakey, M., Harris, J., Eds.; Columbia University Press: New York, NY, USA, 2003; pp. 387–438. [Google Scholar]
- Bernor, R.L.; Scott, R.S. New Interpretations of the Systematics, Biogeography and Paleoecology of the Sahabi Hipparions (latest Miocene), Libya. Geodiversitas 2003, 25, 297–319. [Google Scholar]
- Pomel, A. Les equidés: Carte géologie de l’Algérie. Monogr. Paléontol. 1897, 12, 1–44. [Google Scholar]
- Hooijer, D.A.; Maglio, V.J. The earliest Hipparion south of the Sahara, in the late Miocene of Kenya. Proc. K. Ned. Akad. Wet. 1973, 76, 311–315. [Google Scholar]
- Sun, B.; Zhang, X.; Liu, Y.; Bernor, R.L. Sivalhippus ptychodus and Sivalhippus platyodus (Perissodactyla, Mammalia) from the late Miocene of China. Riv. Ital. Paleo. Strat. 2018, 124, 1–22. [Google Scholar]
- Bernor, R.L.; Kaiser, T. Systematics and Paleoecology of the Earliest Pliocene Equid, Eurygnathohippus hooijeri n. sp. from Langebaanweg, South Africa. Mitt. Aus Hamburischen Zool. Mus. Inst. 2006, 103, 147–183. [Google Scholar]
- Hooijer, D.A. The late Pliocene Equidae of Langebaanweg, Cape Province, South Africa. Zool. Verh. 1976, 148, 3–39. [Google Scholar]
- Bernor, R.L.; Gilbert, H.; Semprebon, G.; Simpson, S.; Semaw, S. Eurygnathohippus woldegabrieli sp. nov. (Perissodactyla: Mammalia) from the Middle Pliocene of Aramis, Ethiopia (4.4 Ma.). J. Vert. Paleo. 2013, 33, 1472–1485. [Google Scholar] [CrossRef]
- Eisenmann, V. Le protostylide: Valeur systematique et signification phylétique chez les espèces actuelles et fossiles du genre Equus (Perissodactyla, Mammalia). Z. Saugetierkd. 1976, 41, 349–365. [Google Scholar]
- Eisenmann, V. Family Equidae. The Fossil Ungulates: Proboscidea, Perissodactyla, and Suidae. In Koobi Fora Research Project; Harris, J.M., Ed.; Clarendon Press: London, UK, 1983; Volume 2, pp. 156–214. [Google Scholar]
- Bernor, R.L.; Scott, R.S.; Haile-Selassie, Y. A Contribution to the Evolutionary History of Ethiopian Hipparionine Horses: Morphometric Evidence from the Postcranial Skeleton. Geodiversitas 2005, 27, 133–158. [Google Scholar]
- Eisenmann, V.; Geraads, D. Hipparion pomeli sp. nov. from the late Pliocene of Ahl al Oughlam, Morocco, and a revision of the relationships of Pliocene and Pleistocene African hipparions. Palaeo Afr. 2006, 42, 51–98. [Google Scholar]
- van Hoepen, E.C.N. Fossiele Pferde van Cornelia. Bloemfontein 1930, 2, 13–24. [Google Scholar]
- Leakey, L.S.B. A Preliminary Report on the Geology and Fauna. In Olduvai Gorge 1951–1961; Cambridge University Press: Cambridge, UK, 1965; Volume 1. [Google Scholar]
- Armour-Chelu, M.; Bernor, R.L.; Mittmann, H.-W. Hooijer’s hypodigm for “Hipparion” cf. ethiopicum (Equidae, Hipparioninae) from Olduvai, Tanzania and comparative material from the East African Plio-Pleistocene. Abh. Bayer. Akad. Wiss. 2006, 30, 15–24. [Google Scholar]
- Armour-Chelu, M.; Bernor, R.L. Equidae. In Geology and Paleontology of Laetoli; Harrison, T., Ed.; Springer: Berlin/Heidelberg, Germany, 2011; pp. 295–326. [Google Scholar]
- van der Made, J.; Boulaghraif, K.; Chelli Cheheb, R.; Caceres, I.; Harichane, Z.; Sahnouni, M. The last North African hipparions —Hipparion decline and extinction follows a common pattern. N. Jb. Geol. Palaont. Abh. 2022, 303, 39–97. [Google Scholar] [CrossRef]
- Bernor, R.L.; Coillot, T.; Wolf, D. Phylogenetic Signatures in the Juvenile Skull and Dentition of Olduvai Gorge Eurygnathohippus cornelianus (Mammalia: Equidae). Riv. Ital. Paleontol. Stratigr. 2014, 120, 243–252. [Google Scholar]
- Hopwood, A.T. Die fossilen Pferde von Oldoway. Wiss. Ergeb. Oldoway Exped. 1937, 4, 112–136. [Google Scholar]
- Thomas, P. Recherches stratigraphiques et paleontologiques sur quelques formations d’eau douce de l’Algerie. Mem. Soc. Geol. Fr. 1884, 3, 1–50. [Google Scholar]
- Eisenmann, V. Les Chevaux (Equus Sensu Lato) Fossiles et Actuels: Crânes et Dents Jugales Supérieures; Editions du Centre National de la Recherche Scientifique: Paris, France, 1980. [Google Scholar]
- Arambourg, C. Vertébrés continentaux du Miocène supérieur de l’Afrique du Nord. Publ. Serv. Cart. Geol. L’algerie 1959, 4, 1–161. [Google Scholar]
- Arambourg, C. Les Vertébrés du Pléistocène de l’Afrique du Nord. Arch. Mus. Natl. D’histoiry Nat. 1970, 10, 1–127. [Google Scholar]
- Azzaroli, A. On Villafranchian Palaearctic Equids and their allies. Palaeontogr. Ital. 1982, 72, 74–97. [Google Scholar]
- Geraads, D.; Raynal, J.-P.; Eisenmann, V. The earliest human occupation of North Africa: A reply to Sahnouni et al. (2002). J. Hum. Evol. 2004, 46, 751–761. [Google Scholar] [CrossRef] [PubMed]
- Tchernov, E. Memoires et Travaux du Centre de Recherche Francais de Jerusalem 5. In The Lower Pleistocene Mammals of ‘Ubeidiya (Jordan Valley); Association Paleorient: Paris, France, 1986. [Google Scholar]
- Yeshurun, R.; Zaidner, Y.; Eisenmann, V.; Martinez-Navarro, B.; Bar-Oz, G. Lower Paleolithic hominin ecology at the fringe of the desert: Faunal remains from Bizat Ruhama and Nahal Hesi, Northern Negev, Israel. J. Hum. Evol. 2011, 60, 492–507. [Google Scholar] [CrossRef]
- Tchernov, E. The Faunal Sequence of the Southwest Asian Middle Paleolithic in Relation to Hominid Dispersal Events. In Neandertals and Modern Humans in Western Asia; Akazawa, T., Aoki, K., Bar-Yosef, O., Eds.; Springer: Berlin/Heidelberg, Germany, 2002; pp. 77–94. [Google Scholar]
- Broom, R. On evidence of a large horse recently extinct in South Africa. Ann. S. Afr. Mus. 1909, 7, 281–282. [Google Scholar]
- Eisenmann, V. Equus capensis (Mammalia, Perissodactyla) from Elandsfontein. Palaeontol. Afr. 2000, 36, 91–96. [Google Scholar]
- Churcher, C.S. The fossil equidae from the Krugersdorp Caves. Ann. Transvaal Mus. 1970, 26, 145–168. [Google Scholar]
- Morom, N.; Lazagabaster, I.A.; Shafir, R.; Natalio, F.; Eisenmann, V.; Kolsa Horwitz, L. The Middle Pleistocene mammalian fauna of Oumm Qatafa Cave, Judean Desert: Taxonomy, taphonomy and paleoenvironment. J. Quat. Sci. 2022, 37, 612–638. [Google Scholar] [CrossRef]
- Bagtache, B.; Hadjouis, D.; Eisenmann, V. Presence d’un Equus caballine (E. algericus n. sp.) et d’une autre espece nouvelle d’Equus (E. melkiensis n. sp.) dans l’Atérien des Allbroges, Algerie. Comptes Rendus Séances L’académie Sci. Paris 1984, 298, 609–612. [Google Scholar]
- Aouraghe, H.; Debenath, A. Les Equides du Pleistocene Superior de la grotte Zourah a el Harhoura, Maroc. Quaternarie 1999, 10, 283–292. [Google Scholar] [CrossRef]
- Oustalet, E. Une nouvelle espèce de zèbre, le zèbre de Grévy (Equus grevyi). Nature 1882, 10, 12–14. [Google Scholar]
- O’Brien, K.; Tryon, C.A.; Blegen, N.; Kimeu, B.; Rowan, J.; Faith, J.T. First appearance of Grévy’s zebra (Equus grevyi), from the Middle Pleistocene of the Kapthurin Formation, Kenya, sheds lights on the evolution and paleoecology of large zebras. Quat. Sci. Rev. 2021, 256, 106835. [Google Scholar] [CrossRef]
- Leonard, J.A.; Rohland, N.; Glaberman, S.; Fleischer, R.C.; Caccone, A.; Hofreiter, M. A rapid loss of stripes: The evolutionary history of the extinct quagga. Biol. Lett. 2005, 1, 291–295. [Google Scholar] [CrossRef]
- Pedersen, C.E.T.; Albrechtsen, A.; Etter, P.D.; Johnson, E.A.; Orlando, L.; Chikhi, L.; Siegismund, H.R.; Heller, R. A southern African origin and cryptic structure in highly mobile plains zebras. Nat. Ecol. Evol. 2018, 2, 491–498. [Google Scholar] [CrossRef]
- Linnaeus, C. Systema Naturae per Regna Tria Naturae, Secundum Classes, Ordines, Genera, Species cum Characteribus, Differentiis, Synonimis, Locis: Editio Decima, Reformata; Laurentii Salvii: Stockholm, Sweden, 1758; pp. 1–824. [Google Scholar]
- von Heuglin, T. Systematische Übersicht der Vögel Nord-Ost-Afrika’s: Mit Einschluss der Arabischen Küste des Rothen Meeres und der Nil-Quellen-Länder Südwärts bis zum IV. Grade Nördl. Breite; K. Akademie der Wissenschaften: Berlin, Germany, 1866. [Google Scholar]
- Churcher, C.S. Oldest ass recovered from Olduvai Gorge, Tanzania, and the origin of asses. J. Paleontol. 1982, 56, 1124–1132. [Google Scholar]
- Noack, N. Neues aus der Tierhandlung von Karl Hagenbeck, sowie aus dem Zoologischen Garten in Hamburg. Zool. Gart. 1884, 25, 100–115. [Google Scholar]
- Groves, C.P. The taxonomy, distribution, and adaptations of recent equids. In Equids in the Ancient World; Meadow, R.H., Uerpmann, H.-P., Eds.; Reichert: Wiesbaden, Germany, 1986; pp. 11–65. [Google Scholar]
- Denzau, G.; Denzau, H. (Eds.) Wildesel; Thorbecke: Stuttgart, Germany, 1999; pp. 1–221. [Google Scholar]
- Groves, C.P.; Smeenk, C. The nomenclature of the African wild ass. Zool. Meded. 2007, 81, 121–135. [Google Scholar]
- Sam, Y. African origins of modern asses as seen from paleontology and DNA: What about the Atlas wild ass? Geobios 2020, 58, 73–84. [Google Scholar] [CrossRef]
- Berggren, W.A.; Van Couvering, J.A. The late Neogene: Biostratigraphy, geochronology and paleoclimatology of the last 15 million years in marine and continental sequences. Palaeogeogr. Palaeoclimatol. Palaeoecol. 1974, 16, 1–216. [Google Scholar]
- Bell, C.J.; Lundelius, E.L., Jr.; Barnosky, A.D.; Graham, R.W.; Lindsay, E.H.; Ruez, D.R., Jr.; Semken, H.A.; Webb, S.D.; Zakrzwski, R.J. The Blancan, Irvingtonian, and Rancholabrean Mammal Ages. In Late Cretaceous and Cenozoic Mammals of North America: Biostratigraphy and Geochronology; Woodburne, M.O., Ed.; Columbia University Press: New York, NY, USA, 2004; pp. 232–314. [Google Scholar]
- Woodburne, M.O. The great American biotic interchange: Dispersals, tectonics, climate, sea level and holding pens. J. Mammal. Evol. 2010, 17, 245–264. [Google Scholar] [CrossRef] [PubMed]
- Avilla, L.S.; Bernardes, C.; Mothé, D. A new genus for Onohippidium galushai Macfadden and Skinner, 1979 (Mammalia, Equidae), from the late Hemphillian of North America. J. Vertebr. Paleontol. 2015, 35, e925909. [Google Scholar] [CrossRef]
- Coltorti, M.; Abbazzi, L.; Ferretti, M.P.; Iacumin, P.; Paredes Rios, F.; Pellegrini, M.; Pieruccini, P.; Rustioni, M.; Rook, L. Last Glacial mammals in South America: A new scenario from the Tarija Basin (Bolivia). Naturwissenschaften 2007, 94, 288–299. [Google Scholar] [CrossRef] [PubMed]
- Bernor, R.L.; Golich, U.B.; Harzahauser, M.; Semprebon, G.M. The Pannonian C hipparions from the Vienna Basin. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2017, 476, 28–41. [Google Scholar] [CrossRef]
- Barry, J.C.; Behrensmeyer, A.K.; Badgley, C.E.; Flynn, L.J.; Peltonen, H.; Cheema, I.U.; Pilbeam, D.; Lindsay, E.H.; Raza, S.M.; Rajpar, A.R.; et al. The Neogene Siwaliks of the Potwar Plateau, Pakistan. In Fossil Mammals of Asia; Wang, X., Flynn, L.J., Fortelius, M., Eds.; Columbia University Press: New York, NY, USA, 2013; pp. 373–399. [Google Scholar]
- Opdyke, N.D.; Lindsay, E.; Johnson, G.D.; Johnson, N.; Tahirkheli, R.A.K.; Mirza, M.A. Magnetic polarity stratigraphy and vertebrate paleontology of the upper siwalik subgroup of northern Pakistan. Palaeogeogr. Palaeoclimatol. Palaeoecol. 1979, 27, 1–34. [Google Scholar] [CrossRef]
- Patnaik, R. Indian Neogene Siwalik mammalian biostratigraphy: An overview. In Fossil Mammals of Asia: Neogene Biostratigraphy and Chronology; Wang, X., Flynn, L.J., Fortelius, M., Eds.; Columbia University Press: New York, NY, USA, 2013; pp. 423–444. [Google Scholar]
- Gaur, R.; Chopra, S.R.K. On a new subspecies of Equus from Pinjor Formation of Upper Sivaliks—With remarks on Sivalik Equus. J. Palaeontol. Soc. India 1984, 29, 19–25. [Google Scholar]
- Zhu, R.X.; Potts, R.; Pan, Y.X.; Yao, H.T.; Lü, L.Q.; Zhao, X. Early evidence of the genus Homo in East Asia. J. Hum. Evol. 2008, 55, 1075–1085. [Google Scholar] [CrossRef]
- Chauhan, P.R. Large mammal fossil occurrences and associated archaeological evidence in Pleistocene contexts of peninsular India and Sri Lanka. Quarter. Int. 2008, 192, 20–42. [Google Scholar] [CrossRef]
- Van Couvering, J.; Delson, E. African Land Mammal Ages. J. Vert. Pal. 2020, 40, 5. [Google Scholar] [CrossRef]
- Werdelin, L. Chronology of Neogene Mammal localities. In Cenozoic Mammals of Africa; Werdelin, L., Sanders, W.L., Eds.; University of California Press: Berkeley, CA, USA, 2010; pp. 27–43. [Google Scholar]
- Pickford, M. Preliminary Miocene mammalian biostratigraphy for Western Kenya. J. Hum. Evol. 1981, 10, 73–97. [Google Scholar] [CrossRef]
- Gilbert, H.; Bernor, R.L. Equidae. In Homo Erectus—Pleistocene Evidence from the Middle Awash, Ethiopia; Gilbert, H., Asfaw, B., Eds.; University of California Press: Berkeley, CA, USA, 2008; pp. 133–166. [Google Scholar]
- Geist, V. The relation of social evolution and dispersal in ungulates during the Pleistocene, with emphasis on Old World deer and the genus Bison. Quat. Res. 1971, 1, 285–315. [Google Scholar] [CrossRef]
- Guthrie, D.R. Frozen Fauna of the Mammoth Steppe—The Story of Blue Babe; University of Chicago Press: Chicago, IL, USA, 1990. [Google Scholar]
- McNab, B.K. Geographic and temporal correlations of mammalian size reconsidered: A resource rule. Oecologia 2010, 164, 13–23. [Google Scholar] [CrossRef] [PubMed]
- Saarinen, J. Ecometrics of Large Herbivorous Land Mammals in Relation to Climatic and Environmental Changes during the Pleistocene. Ph.D. Thesis, University of Helsinki, Helsinki, Finland, 2014. [Google Scholar]
- Damuth, J.; Janis, C. On the relationships between hypsodonty and feeding ecology in ungulate mammals, and its utility in paleoecology. Biol. Rev. Camb. Philos. Soc. 2011, 86, 733–758. [Google Scholar] [CrossRef]
- Strömberg, C. Using phytolith assemblages to reconstruct the origin and spread of grass-dominated habitats in the great plains of North America during the late Eocene to early Miocene. Palaeogeo. Palaeoclim. Palaeoecol. 2004, 207, 239–275. [Google Scholar] [CrossRef]
- Strömberg, C. Decoupled taxonomic radiation and ecological expansion of open-habitat grasses in the Cenozoic of North America. Proc. Natl. Acad. Sci. USA 2005, 102, 11980–11984. [Google Scholar] [CrossRef]
- Strömberg, C. Evolution of Grasses and Grassland Ecosystems. Annu. Rev. Earth Planet. Sci. 2011, 39, 517–544. [Google Scholar] [CrossRef]
- Janis, C.; Sahney, S.; Benton, M. Grit not grass: Concordant patterns of early origin of hypsodonty in Great Plains ungulates and Glires. Palaeogeo. Palaeoclim. Palaeoecol. 2012, 366, 1–10. [Google Scholar]
- Semprebon, G.M.; Rivals, F.; Janis, C.M. The role of grass vs. exogenous abrasives in the paleodietary patterns of North American ungulates. Front. Ecol. Evol. 2019, 7, 65. [Google Scholar] [CrossRef]
- Strömberg, C.; McInerney, F. The Neogene transition from C3 to C4 grasslands in North America: Assemblage analysis of fossil phytoliths. Paleobiology 2011, 37, 50–71. [Google Scholar] [CrossRef]
- Pérez-Crespo, V.A.; Bravo-Cuevas, V.M.; Arroyo-Cabrales, J. Feeding habits of Equus conversidens and Haringtonhippus francisci from Valsequillo, Puebla, México. Hist. Biol. 2021, 34, 1252–1259. [Google Scholar] [CrossRef]
- Hulbert, R.C. Taxonomic evolution in North American Neogene horses (Subfamily Equinae): The rise and fall of an adaptive radiation. Paleobiology 1993, 19, 216–234. [Google Scholar] [CrossRef]
- Hulbert, R.C.; MacFadden, B.J. Morphological transformation and cladogenesis at the base of the adaptive radiation of Miocene hypsodont horses. Am. Mus. Novit. 1991, 3000, 1–61. [Google Scholar]
- Radinsky, L. Allometry and reorganization of horse skull proportions. Science 1983, 221, 1189–1191. [Google Scholar] [CrossRef] [PubMed]
- Radinsky, L. Ontogeny and phylogeny in horse skull evolution. Int. J. Org. Evol. 1984, 38, 1–15. [Google Scholar] [CrossRef]
- Hulbert, R.C. Phylogenetic interrelationships and evolution of North American late Neogene Equidae. In The Evolution of Perissodactyls; Prothero, D.R., Schoch, R.M., Eds.; Clarendon Press: Oxford, UK, 1989; pp. 176–196. [Google Scholar]
- Osborn, H.F. The Age of Mammals; MacMillan Company: New York, NY, USA, 1910. [Google Scholar]
- Scott, W.B. A History of Land Mammals in the Western Hemisphere; MacMillian Company: New York, NY, USA, 1937. [Google Scholar]
- Simpson, G.G. Tempo and Mode in Evolution; Columbia University Press: New York, NY, USA, 1944. [Google Scholar]
- Stirton, R.A. Observation of evolutionary rates in hypsodonty. Evolution 1947, 1, 32–41. [Google Scholar] [CrossRef]
- Webb, S.D. A history of savanna vertebrates in the New World. Part I: North America. Annu. Rev. Ecol. Syst. 1977, 8, 355–380. [Google Scholar] [CrossRef]
- Webb, S.D. The rise and fall of the late Miocene ungulate fauna in North America. In Coevolution; Nitecki, M.H., Ed.; University of Chicago Press: Chicago, IL, USA, 1983; pp. 267–306. [Google Scholar]
- Janis, C.M. The significance of fossil ungulate communities as indicators of vegetation structure and climate. In Fossils and Climate; Brenchley, P.J., Ed.; John Wiley & Sons: New York, NY, USA, 1984; pp. 85–104. [Google Scholar]
- Janis, C.M. Tertiary mammal evolution in the context of changing climates, vegetation, and tectonic events. Annu. Rev. Ecol. Syst. 1993, 24, 467–500. [Google Scholar] [CrossRef]
- Webb, S.D.; Opdyke, N.D. Global climatic influence on Cenozoic land mammal faunas. National Research Council Panel on Effects of Past Global Change on Life. In Effects of Past Global Change on Life; National Academies Press: Washington, DC, USA, 1995. [Google Scholar]
- Cohen, J.; DeSantis, L.R.G.; Lindsay, E.L.; Meachen, J.A.; O’Keefe, F.R.; Southon, J.R.; Binder, W.J. Dietary stability inferred from dental mesowear analysis in large ungulates from Racho La Brea and opportunistic feeding during the late Pleistocene. Palaeogeo. Palaeoclim. Palaeoecol. 2021, 570, 110360. [Google Scholar] [CrossRef]
- Jones, D.B.; DeSantis, L.R.G. Dietary ecology od ungulates from La Brea tar pits in southern California: A multi-proxy approach. Palaeogeo. Palaeoclim. Palaeoecol. 2017, 466, 110–117. [Google Scholar] [CrossRef]
- Kaiser, T.M.; Solounias, N.; Fortelius, M.; Bernor, R.L.; Schrenk, F. Tooth mesowear analysis on Hippotherium primigenium from the Vallesian Dinotheriensande (Germany). A blind test study. Carolinea 2000, 58, 103–114. [Google Scholar]
- Kaiser, T.M.; Bernor, R.L.; Scott, R.S.; Franzen, J.L.; Solounias, N. New Interpretations of the Systematics and Palaeoecology of the Dorn Dürkheim 1 Hipparions (Late Miocene, Turolian Age [MN11]) Rheinhessen, German. Senckenberg Lethea 2003, 83, 103. [Google Scholar] [CrossRef]
- Rivals, F.; Lister, A.M. Dietary flexibility and niche partitioning of large herbivores through the Pleistocene of Britain. Quat. Sci. Rev. 2016, 146, 116–133. [Google Scholar] [CrossRef]
- Head, M.J. Pollen and dinoflagellates from the Red Crag at Walton-on-the-Naze, Essex: Evidence for a mild climatic phase during the early Late Pliocene of eastern England. Geol. Mag. 1998, 135, 803–817. [Google Scholar] [CrossRef]
- Zalasiewicz, J.A.; Mathers, S.J.; Hughes, M.J.; Gibbard, P.L.; Peglar, S.M.; Harland, R.; Nicholson, R.A.; Boulton, G.S.; Cambridge, P.; Wealthall, G.P. Stratigraphy and Palaeoenvironments of the Red Crag and Norwich Crag Formations between Aldeburgh and Sizewell, Suffolk, England. Philos. Trans. R. Soc. Lond. 1988, 322, 221–272. [Google Scholar]
- Rivals, F.; Athanassiou, A. Dietary adaptations in an ungulate community from the late pliocene of greece. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2008, 265, 134–139. [Google Scholar] [CrossRef]
- Strani, F.; DeMiguel, D.; Bellucci, L.; Sardella, R. Dietary response of early pleistocene ungulate communities to the climate oscillations of the Gelasian/Calabrian transition in central Italy. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2018, 499, 102–111. [Google Scholar] [CrossRef]
- Strani, F.; DeMiguel, D.; Sardella, R.; Bellucci, L. Paleoenvironments and climatic changes in the italian peninsula during the early pleistocene: Evidence from dental wear patterns of the ungulate community of Coste San Giacomo. Quat. Sci. Rev. 2015, 121, 28–35. [Google Scholar] [CrossRef]
- Strani, F.; Pushkina, D.; Bocherens, H.; Bellucci, L.; Sardella, R.; DeMiguel, D. Dietary adaptations of early and middle pleistocene equids from the anagni basin (Frosinone, Central Italy). Front. Ecol. Evol. 2019, 7, 176. [Google Scholar] [CrossRef]
- Saarinen, J.; Oksanen, O.; Zliobaite, I.; Fortelius, M.; DeMiguel, D.; Azanza, B.; Bocherens, H.; Luzón, C.; Solano-Garcia, J.; Yravedra, J.; et al. Pliocene to Middle Pleistocene climate history in the Guadix-Baza Basin, and the environmental conditions of early Homo in Europe. Quat. Sci. Rev. 2021, 268, 107132. [Google Scholar] [CrossRef]
- Strani, F.; DeMiguel, D.; Alba, D.; Moyà-Solà, S.; Bellucci, L.; Sardella, R.; Madurell-Malapeira, J. The effects of the “0.9 Ma event” on the Mediterranean ecosystems during the Early-Middle Pleistocene transition as revealed by dental wear patterns of fossil ungulates. Quat. Sci. Rev. 2019, 210, 80–89. [Google Scholar] [CrossRef]
- Blain, H.-A.; Lozano-Fernández, I.; Agustí, J.; Bailon, S.; Menéndez, L.; Patrocinio, M.; Ros-Montoya, S.; Manuel, J.; Toro-Moyano, I.; Martínez-Navarro, B.; et al. Refining upon the climatic background of the early Pleistocene hominid settlement in western Europe: Barranco León and Fuente Nueva-3 (Guadix- Baza Basin, SE Spain). Quat. Sci. Rev. 2016, 144, 132–144. [Google Scholar] [CrossRef]
- Sánchez-Bandera, C.; Oms, O.; Blain, H.-A.; Lozano-Fernández, I.; Bispal-Chinesta, J.F.; Agustí, J.; Saarinen, J.; Fortelius, M.; Titton, S.; Serrano-Ramos, A.; et al. New stratigraphically constrained palaeoenvironmental reconstructions for the first human settlement in Western Europe: The Early Pleistocene herpetofaunal assemblages from Barranco León and Fuente Nueva 3 (Granada, SE Spain). Quat. Sci. Rev. 2020, 243, 106466. [Google Scholar] [CrossRef]
- Kahlke, R.D.; García, N.; Kostopoulos, D.S.; Lacombat, F.; Lister, A.M.; Mazza, P.P.; Spassov, N.; Titov, V.V. Western Palaearctic palaeoenvironmental conditions during the Early and early Middle Pleistocene inferred from large mammal communities, and implications for hominin dispersal in Europe. Quat. Sci. Rev. 2011, 30, 1368–1395. [Google Scholar] [CrossRef]
- Rivals, F.; Julien, M.-A.; Kuitems, M.; Van Kolfschoten, T.; Serangeli, J.; Drucker, D.G.; Bocherens, H.; Conard, N.J. Investigation of equid paleodiet from Schöningen 13 II-4 through dental wear and isotopic analyses: Archaeological implications. J. Hum. Evol. 2015, 89, 129–137. [Google Scholar] [CrossRef] [PubMed]
- Roberts, M.B. Excavation of the Lower Paleolithic site at Amey’s Eartham Pit, Boxgrove, West Sussex: A preliminary report. Proc. Prehist. Soc. 1986, 52, 215–245. [Google Scholar] [CrossRef]
- Urban, B.; Bigga, G. Environmental reconstruction and biostratigraphy of late Middle Pleistocene lakeshore deposits at Schöningen. J. Hum. Evol. 2015, 89, 57–70. [Google Scholar] [CrossRef]
- Strani, F.; DeMiguel, D.; Bona, F.; Sardella, R.; Biddittu, I.; Bruni, L.; De Castro, A.; Guadagnoli, F.; Bellucci, L. Ungulate dietary adaptations and palaeoecology of the Middle Pleistocene site of Fontana Ranuccio (anagni, central italy). Palaeogeogr. Palaeoclimatol. Palaeoecol. 2018, 496, 238–247. [Google Scholar] [CrossRef]
- Leroi-Gourhan, A. Pollen analysis of sediment samples from Gough’s Cave, Cheddar. Proc. Univ. Bristol. Spelaeol. Soc. 1986, 17, 141–144. [Google Scholar]
- Bates, M.R.; Bates, C.R.; Gibbard, P.L.; Macphail, R.I.; Owen, F.J.; Parfitt, S.A.; Preece, R.C.; Roberts, M.B.; Robinson, J.E.; Whittaker, J.E.; et al. Late Middle Pleistocene deposits at Norton Farm on the West Sussex coastal plain, southern England. J. Quat. Sci. 2000, 15, 61–89. [Google Scholar] [CrossRef]
- West, R.G.; Lambert, C.A.; Sparks, B.W.; Dickson, J.H. Interglacial deposits at Ilford, Essex. Philos. Trans. R. Soc. 1964, 247, 185–212. [Google Scholar]
- Gibbard, P.L. Pleistocene History of the Lower Thames Valley; Cambridge University Press: Cambridge, UK, 1994. [Google Scholar]
- Sedov, S.N.; Khokhlova, O.S.; Sinitsyn, A.A.; Korkka, A.V.; Ortega, B.; Solleiro, E.; Rozanova, M.S.; Kuznetsova, A.M. Kazdym Late Pleistocene paleosol sequences as an instrument for the local paleographic reconstruction of the Kostenki 14 key section (Voronezh oblast) as an example. Eurasian Soil Sc. 2010, 43, 876–892. [Google Scholar] [CrossRef]
- Ukraintseva, V.V. Mammoths and the Environment; Cambridge University Press: Cambridge, UK, 2013. [Google Scholar]
- van Asperen, E.N.; Stefaniak, K.; Proskurnyak, I.; Ridush, B. Equids from Emine-Bair-Khosar Cave (Crimea, Ukraine): Co-occurrence of the stenonid Equus hydruntinus and the caballoid E. ferus latipes based on skull and postcranial remains. Palaeontol. Electron. 2012, 15, 28. [Google Scholar] [CrossRef]
- Bernor, R.L.; Kaiser, T.M.; Nelson, S.V. The Oldest Ethiopian Hipparion (Equinae, Perissodactyla) from Chorora: Systematics, Paleodiet and Paleoclimate. Cour. Forsch. Inst. Senckenberg 2004, 246, 213–226. [Google Scholar]
- Bernor, R.L. The Latest Miocene Hipparionine (Equidae) from Lemundong’o, Kenya. Kirtlandia 2007, 56, 148–151. [Google Scholar]
- Bernor, R.L.; Kaiser, T.M.; Wolf, D. Revisiting Sahabi Equid Species Diversity, Biogeographic Patterns and Diet Preferences. Gharyounis Bull. 2008, 5, 159–167. [Google Scholar]
- Bernor, R.L.; Haile Selassie, Y. Equidae. In Ardipithecus Kadabba: Late Miocene Evidence from the Middle Awash, Ethiopia; Haile-Selassie, Y., Woldegabriel, G., Eds.; University of California Press: Berkeley, CA, USA, 2009; pp. 397–428. [Google Scholar]
- Bernor, R.L.; Boaz, N.T.; Rook, L. Eurygnathohippus feibeli (Perissodactyla: Mammalia) from the late Miocene of Sahabi (Libya) and its evolutionary and Biogeographic Significance. Bolletino Soc. Paleontol. Ital. 2012, 51, 39–48. [Google Scholar]
- Schulz, E.; Kaiser, T.M. Historical distribution, habitat requirements and feeding ecology of the genus Equus (Perissodactyla). Mammal. Rev. 2013, 43, 111–123. [Google Scholar] [CrossRef]
- Goloboff, P.; Farris, J.; Nixon, K. TNT, a free program for phylogenetic analysis. Cladistics 2008, 24, 774–786. [Google Scholar] [CrossRef]
- Bennett, D.K. Stripes do not a zebra make, Part I: A cladistic analysis of Equus. Syst. Zool. 1980, 29, 272–287. [Google Scholar] [CrossRef]
- Antoine, P.O. Phylogénie et évolution des Elasmotheriina (Mammalia, Rhinocerotidae). Mémoir. Mus. Natl. Hist. 2002, 188, 1–359. [Google Scholar]
- Antoine, P.O.; Downing, K.F.; Crochet, J.Y.; Duranthon, F.; Flynn, L.J.; Marivaux, L.; Métais, G.; Rajpar, A.R.; Roohi, G. A revision of Aceratherium blanfordi Lydekker, 1884 (Mammalia: Rhinocerotidae) from the Early Miocene of Pakistan: Postcranials as a key. Zool. J. Linn. Soc. Lond. 2010, 160, 139–194. [Google Scholar] [CrossRef]
- Pandolfi, L. Persiatherium rodleri, gen. et sp. nov. (Mammalia, Rhinocerotidae), from the Late Miocene of Maragheh (Northwestern Iran). J. Vert Paleontol. 2015, 36, e1040118. [Google Scholar] [CrossRef]
- de Queiroz, K.; Gauthier, J. Phylogenetic taxonomy. Annu. Rev. Ecol. Syst. 1992, 23, 449–480. [Google Scholar] [CrossRef]
- Ereshefsky, M. The Poverty of the Linnaean hierarchy. A Philosophical Study of Biological Taxonomy; Cambridge University Press: Cambridge, UK, 2001. [Google Scholar]
- Papavero, N.; Llorente-Bousquets, J.; Minoro Abel, J. Proposal of a new system of nomenclature for phylogenetic systematics. Arq. Zool. 2001, 36, 1–145. [Google Scholar] [CrossRef][Green Version]
- Béthoux, O. Propositions for a character-state-based biological taxonomy. Zool. Scr. 2007, 36, 409–416. [Google Scholar] [CrossRef]
- Cantino, P.D.; de Queiroz, K. PhyloCode: International Code of Phylogenetic Nomenclature (Version 4c). 2010. Available online: http://www.ohio.edu/phylocode (accessed on 5 June 2022).
- Zachos, F.E. Linnaean ranks, temporal banding, and time-clipping: Why not slaughter the sacred cow? Biol. J. Linn. Soc. 2011, 103, 732–734. [Google Scholar] [CrossRef]
- Vences, M.; Guayasamin, J.M.; Miralles, A.; de la Riva, I. To name or not to name: Criteria to promote economy of change in Linnaean classification schemes. Zootaxa 2013, 3636, 201–244. [Google Scholar] [CrossRef]
- Hennig, W. Phylogenetic Systematics; University Illinois Press: Champaign, IL, USA, 1966. [Google Scholar]
- Mayr, E. Principles of Systematic Zoology; McGraw-Hill: New York, NY, USA, 1969. [Google Scholar]
- Groves, C.P. Primate Taxonomy; Smithsonian Institution Press: Washington, DC, USA, 2001. [Google Scholar]
- Groves, C.P. The what, why, and how of primate taxonomy. Int. J. Primatol. 2004, 25, 1105–1126. [Google Scholar] [CrossRef]
- Simpson, G.G. The Major Features of Evolution; Columbia University Press: New York, NY, USA, 1953. [Google Scholar]
- Mayr, E. Taxonomic categories in fossil hominids. Cold Spring Harb. Symp. Quant. Biol. 1950, 15, 109–118. [Google Scholar] [CrossRef]
- de Queiroz, K. Replacement of an essentialistic perspective on taxonomic definitions as exemplified by the definition of “Mammalia”. Syst. Biol. 1994, 43, 497–510. [Google Scholar] [CrossRef]
- Janis, C.M.; Bernor, R.L. The Evolution of Equid Monodactyly: A Review Including a New Hypothesis. Front. Ecol. Evol 2019, 7, 119. [Google Scholar] [CrossRef]
- Schultz, J.R. Plesippus francescana (Frick) from the late Pliocene, Coso Mountains, California, with a review of the genus Plesippus. In Contributions to Palæontology. Publications of the Carnegie Institution of Washington; Carnegie institution of Washington: Washington, DC, USA, 1936; Volume 1, pp. 1–13. [Google Scholar]
- Eisenmann, V.; Baylac, M. Extant and fossil Equus (Mammalia, Perissodactyla) skulls: A morphometric definition of the subgenus Equus. Zool. Scr. 2000, 29, 89–100. [Google Scholar] [CrossRef]
- Brochu, A.B.; Sumrall, C.D. Phylogenetic Nomenclature and Paleontology. J. Paleontol. 2001, 75, 754–757. [Google Scholar] [CrossRef]
- Bryant, H.N. Explicitness, Stability, and Universality in the Phylogenetic Definition and Usage of Taxon Names: A Case Study of the Phylogenetic Taxonomy of the Carnivora (Mammalia). Syst. Biol. 1996, 45, 174–189. [Google Scholar] [CrossRef]
- Cantino, P.D.; Olmstead, R.G.; Wagstaff, S.J. A Comparison of Phylogenetic Nomenclature with the Current System: A Botanical Case Study. Syst. Biol. 1997, 46, 313–331. [Google Scholar] [CrossRef]
- Schulte, J.S.; Macey, J.R.; Larson, A.; Papenfuss, T.J. Molecular Tests of Phylogenetic Taxonomies: A General Procedure and Example Using Four Subfamilies of the Lizard Family Iguanidae. Mol. Phylogenetics Evol. 1998, 10, 367–376. [Google Scholar] [CrossRef]
- de Queiroz, K.; Gauthier, J. Phylogeny as a Central Principle in Taxonomy: Phylogenetic Definitions of Taxon Names. Syst. Biol. 1990, 39, 307–322. [Google Scholar] [CrossRef]
- de Queiroz, K.; Gauthier, J. Toward a phylogenetic system of biological nomenclature. Trends Ecol. Evol. 1994, 9, 27–31. [Google Scholar] [CrossRef]
- de Queiroz, K.; Cantino, P.D. Taxon Names, Not Taxa, Are Defined. Taxon 2001, 50, 821–826. [Google Scholar] [CrossRef]
- Vilstrup, J.T.; Seguin-Orlando, A.; Stiller, M.; Ginolhac, A.; Raghavan, M.; Nielsen, S.C.A.; Weinstock, J.; Froese, D.; Vasiliev, S.K.; Ovodov, N.D. Mitochondrial Phylogenomics of Modern and Ancient Equids. PLoS ONE 2013, 8, e55950. [Google Scholar]
- MacFadden, B.; Solounias, N.; Cerling, T.E. Ancient Diets, Ecology, and Extinction of 5-Million-Year-Old Horses from Florida. Science 1999, 283, 824–827. [Google Scholar] [CrossRef]
- Semprebon, G.M.; Rivals, F.; Solounias, N.; Hulbert, R.C. Paleodietary reconstruction of fossil horses from the Eocene through Pleistocene of North America. Palaeogeogr. Palaeoclim. Palaeoecol. 2016, 442, 110–127. [Google Scholar] [CrossRef]
- Miller, W.E. The Late Pliocene Las Tunas Local Fauna from Southernmost Baja California, Mexico. J. Paleontol. 1980, 54, 762–805. [Google Scholar]
- MacFadden, B.J.; Carranza-Castañeda, O. Cranium of Dinohippus mexicanus (Mammalia: Equidae) from the Early Pliocene (latest Hemphillian) of Central Mexico, and the origin of Equus. Bull. Fla. Mus. Nat. Hist. 2002, 43, 163–185. [Google Scholar]
- Carranza-Castañeda, O. Dinohippus mexicanus (Early—Late, Late, and Latest Hemphillian) and the transition to genus Equus, in Central Mexico faunas. Front. Earth Sci. 2019, 7, 89. [Google Scholar] [CrossRef]
- Lowenstein, J.M.; Rayder, O.A. Immunological systematics of the extinct quagga (Equidae). Experientia 1985, 41, 1192–1193. [Google Scholar] [CrossRef]
- George, M.; Rayder, O.A. Mitochondrial DNA evolution in the genus Equus. Mol. Biol. Evol. 1986, 3, 535–546. [Google Scholar]
- Oakefull, E.A.; Clegg, J.B. Phylogenetic Relationships within the Genus Equus and the Evolution of α and θ Globin Genes. J. Mol. Evol. 1998, 47, 772–783. [Google Scholar] [CrossRef]
- Kruger, K.; Gaillard, C.; Stranzinger, G.; Rieder, S. Phylogenetic analysis and species allocation of individual equids using microsatellite data. J. Anim. Breed. Genet. 2005, 122, 78–86. [Google Scholar] [CrossRef] [PubMed]
- Steiner, C.C.; Ryder, O.A. Molecular phylogeny and evolution of the Perissodactyla. Zool. J. Linn. Soc. 2011, 163, 1289–1303. [Google Scholar] [CrossRef]
- Orlando, L.; Mashkour, M.; Burke, A.; Douady, C.J.; Eisenmann, V.; Hanni, C. Geographic distribution of an extinct equid (Equus hydruntinus: Mammalia, Equidae) revealed by morphological and genetical analyses of fossils. Mol. Ecol. 2006, 15, 2083–2093. [Google Scholar] [CrossRef] [PubMed]
- Orlando, L.; Male, D.; Alberdi, M.T.; Prado, J.L.; Prieto, A.; Cooper, A.; Hanni, C. Ancient DNA Clarifies the Evolutionary History of American Late Pleistocene Equids. J. Mol. Evol. 2008, 66, 533–538. [Google Scholar] [CrossRef]
- Flint, J.; Ryder, O.A.; Clegg, J.B. Comparison of theα-globin gene cluster structure in Perissodactyla. J. Mol. Evol. 1990, 30, 36–42. [Google Scholar] [CrossRef]
- Wallner, B.; Brem, G.; Muller, M.; Achmann, R. Fixed nucleotide differences on the Y chromosome indicate clear divergence between Equus przewalskii and Equus caballus. Anim. Genet. 2003, 34, 453–456. [Google Scholar] [CrossRef]
- Kaminski, M. The biochemical evolution of the horse. Comp. Biochem. 1979, 63, 175–178. [Google Scholar] [CrossRef]
- Ishida, N.; Oyunsuren, T.; Mashima, S.; Mukoyama, H.; Naruya, S. Mitochondrial DNA sequences of various species of the genus Equus with special reference to the phylogenetic relationship between Przewalskii’s wild horse and domestic horse. J. Mol. Evol. 1995, 41, 180–188. [Google Scholar] [CrossRef]
- Oakenfull, E.A.; Lim, H.N.; Ryder, O.A. A survey of equid mitochondrial DNA: Implications for the evolution, genetic diversity and conservation of Equus. Conserv. Genet. 2000, 1, 341–355. [Google Scholar] [CrossRef]
- Cai, D.; Zhu, S.; Gong, M.; Zhang, N.; Wen, J.; Qiyao, L.; Sun, W.; Shao, X.; Guo, Y.; Cai, Y.; et al. Radiocarbon and genomic evidence for the survival of Equus Sussemionus until the late Holocene. eLife 2022, 11, e73346. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cirilli, O.; Machado, H.; Arroyo-Cabrales, J.; Barrón-Ortiz, C.I.; Davis, E.; Jass, C.N.; Jukar, A.M.; Landry, Z.; Marín-Leyva, A.H.; Pandolfi, L.; et al. Evolution of the Family Equidae, Subfamily Equinae, in North, Central and South America, Eurasia and Africa during the Plio-Pleistocene. Biology 2022, 11, 1258. https://doi.org/10.3390/biology11091258
Cirilli O, Machado H, Arroyo-Cabrales J, Barrón-Ortiz CI, Davis E, Jass CN, Jukar AM, Landry Z, Marín-Leyva AH, Pandolfi L, et al. Evolution of the Family Equidae, Subfamily Equinae, in North, Central and South America, Eurasia and Africa during the Plio-Pleistocene. Biology. 2022; 11(9):1258. https://doi.org/10.3390/biology11091258
Chicago/Turabian StyleCirilli, Omar, Helena Machado, Joaquin Arroyo-Cabrales, Christina I. Barrón-Ortiz, Edward Davis, Christopher N. Jass, Advait M. Jukar, Zoe Landry, Alejandro H. Marín-Leyva, Luca Pandolfi, and et al. 2022. "Evolution of the Family Equidae, Subfamily Equinae, in North, Central and South America, Eurasia and Africa during the Plio-Pleistocene" Biology 11, no. 9: 1258. https://doi.org/10.3390/biology11091258
APA StyleCirilli, O., Machado, H., Arroyo-Cabrales, J., Barrón-Ortiz, C. I., Davis, E., Jass, C. N., Jukar, A. M., Landry, Z., Marín-Leyva, A. H., Pandolfi, L., Pushkina, D., Rook, L., Saarinen, J., Scott, E., Semprebon, G., Strani, F., Villavicencio, N. A., Kaya, F., & Bernor, R. L. (2022). Evolution of the Family Equidae, Subfamily Equinae, in North, Central and South America, Eurasia and Africa during the Plio-Pleistocene. Biology, 11(9), 1258. https://doi.org/10.3390/biology11091258