
Four New Pale-Spored Species of Xylaria (Xylariaceae, Xylariales) with a Key to Worldwide Species on Fallen Fruits and Seeds

 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Collection and Morphological Study
2.2. Molecular Procedures and Phylogenetic Analyses

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Specimen No. | Origin | Host | GenBank Accession Number | Reference | ||
|---|---|---|---|---|---|---|---|
| ITS | RPB2 | β-Tubulin | |||||
| Amphirosellinia fushanensis | HAST 91111209 (HT) | China | dead twigs | GU339496 | GQ848339 | GQ495950 | [36,37] |
| A. nigrospora | HAST 91092308 (HT) | China | dead twigs | GU322457 | GQ848340 | GQ49595 | [36,37] |
| Astrocystis mirabilis | HAST 94070803 | China | bamboo culms | GU322448 | GQ844835 | GQ49594 | [36] |
| As. sublimbata | HAST 89032207 | China | bamboo culms | GU322447 | GQ844834 | GQ495940 | [36] |
| Kretzschmaria guyanensis | HAST 89062903 | China | bark | GU300079 | GQ844792 | GQ478214 | [36] |
| K. sandvicensis | JDR 113 | USA | wood | GU300076 | GQ844786 | GQ478211 | [36] |
| Nemania abortiva | BiSH 467 (HT) | USA | decayed wood | GU292816 | GQ844768 | GQ470219 | [36] |
| N. diffusa | HAST 91020401 | China | bark | GU292817 | GQ844769 | GQ470220 | [36] |
| Podosordaria mexicana | WSP 176 | Mexico | horse dung | GU324762 | GQ853039 | GQ844840 | [36] |
| P. muli | WSP 167 (HT) | Mexico | mule dung | GU324761 | GQ853038 | GQ844839 | [36] |
| Poronia pileiformis | WSP 88113001 (ET) | China | cow dung | GU324760 | GQ853037 | GQ502720 | [36] |
| Rosellinia buxi | JDR 99 | France | Buxus sempervivens | GU300070 | GQ844780 | GQ470228 | [36] |
| R. necatrix | HAST 89062904 | China | root | EF026117 | GQ844779 | EF025603 | [36] |
| Xylaria adscendens | HAST 570 | Guadeloupe | wood | GU300101 | GQ844817 | GQ487708 | [36] |
| X. aethiopica | YMJ 1136 | Ethiopia | pods of Millettia ferruginea | MH790445 | MH785222 | MH785221 | [11] |
| X. allantoidea | HAST 94042903 | China | trunk | GU324743 | GQ848356 | GQ502692 | [36] |
| X. amphithele | HAST 529 | Guadeloupe | dead leaves | GU300083 | GQ844796 | GQ478218 | [36] |
| X. apoda | HAST 90080804 | China | bark | GU322437 | GQ844823 | GQ495930 | [36] |
| X. arbuscula | HAST 89041211 | China | bark | GU300090 | GQ844805 | GQ478226 | [36] |
| X. atrosphaerica | HAST 91111214 | China | bark | GU322459 | GQ848342 | GQ495953 | [36] |
| X. berteri | HAST 90112623 | China | wood | GU324749 | GQ848362 | AY951763 | [36] |
| X. betulicola | FCATAS750 (HT) | China | leaves of Betula | MF774332 | N/A | N/A | [22] |
| X. brunneovinosa | HAST 720 (HT) | China | ground of bamboo plantation | EU179862 | GQ853023 | GQ502706 | [36,38] |
| X. carpophila | CBS 453.72 | Netherlands | - | MH860527 | N/A | N/A | [39] |
| X. cirrata | HAST 664 (ET) | China | ground of vegetable farm | EU179863 | GQ853024 | GQ502707 | [36,38] |
| X. cranioides | HAST 226 | China | wood | GU300075 | GQ844785 | GQ478210 | [36] |
| X. crinalis | FCATAS751 (HT) | China | wood | MF774330 | N/A | N/A | [22] |
| X. cubensis | JDR 860 | USA | wood | GU991523 | GQ848365 | GQ502700 | [36] |
| X. culleniae | JDR 189 | Thailand | pod | GU322442 | GQ844829 | GQ495935 | [36] |
| X. curta | HAST 92092022 | China | bark | GU322443 | GQ844830 | GQ495936 | [36] |
| X. digitata | HAST 919 | Ukraine | wood | GU322456 | GQ848338 | GQ495949 | [36] |
| X. enterogena | HAST 785 | French Guiana | wood | GU324736 | GQ848349 | GQ502685 | [36] |
| X. fabacearum | MFLU 16-1061 (HT) | Thailand | seed pods of Fabaceae | NR171104 | MT212202 | MT212220 | [8] |
| X. fabaceicola | MFLU 16-1072 (HT) | Thailand | seed pods of Fabaceae | NR171103 | MT212201 | MT212219 | [8] |
| X. feejeensis | HAST 92092013 | China | bark | GU322454 | GQ848336 | GQ495947 | [36] |
| X. ficicola | HMJAU 22818 | China | leaves and petioles of Ficus auriculata | MZ351258 | N/A | N/A | [40] |
| X. filiformis | GUM 1052 | Iran | herbaceous stem | KP218907 | N/A | N/A | [41] |
| X. fimbriata | HAST 491 | Martinique | termite nest | GU324753 | GQ853022 | GQ502705 | [36] |
| X. fissilis | HAST 367 | Martinique | bark | GU300073 | GQ844783 | GQ470231 | [36] |
| X. frustulosa | HAST 92092010 | China | bark | GU322451 | GQ844838 | GQ495944 | [36] |
| X. cf. glebulosa | HAST 431 | Martinique | fruit | GU322462 | GQ848345 | GQ495956 | [36] |
| X. globosa | HAST 775 | Guadeloupe | bark | GU324735 | GQ848348 | GQ502684 | [36] |
| X. grammica | HAST 479 | China | wood | GU300097 | GQ844813 | GQ487704 | [36] |
| X. griseosepiacea | HAST 641 (HT) | China | ground of mango orchard | EU179865 | GQ853031 | GQ502714 | [36,38] |
| X. guareae | PR71 | Puerto Rico | - | AY909009 | N/A | N/A | [42] |
| X. haemorrhoidalis | HAST 89041207 | China | bark | GU322464 | GQ848347 | GQ502683 | [36] |
| X. hedyosmicola | FCATAS857 | China | leaves of Hedyosmum orientale | MZ227023 | MZ683407 | MZ221183 | [40] |
| X. hypoxylon | HAST 95082001 | China | wood | GU300095 | GQ844811 | GQ487703 | [36] |
| X. intracolorata | HAST 90080402 | China | bark | GU324741 | GQ848354 | GQ502690 | [36] |
| X. intraflava | HAST 725 (HT) | China | ground of bamboo plantation | EU179866 | GQ853035 | GQ502718 | [36] |
| X. juruensis | HAST 92042501 | China | Arenga engleri | GU322439 | GQ844825 | GQ495932 | [36] |
| X. karyophthora | DRH059 | Guyana | seeds of Chlorocardium sp. | KY564220 | KY564216 | N/A | [12] |
| X. laevis | HAST 95072910 | China | bark | GU324747 | GQ848360 | GQ502696 | [36] |
| X. lindericola | FCATAS852 | China | leaves of Lindera robusta | MZ005635 | MZ031982 | MZ031978 | [40] |
| X. liquidambaris | HAST 93090701 | China | fruits of Liquidambar formosana | GU300094 | GQ844810 | GQ487702 | [36] |
| X. meliacearum | JDR 148 | Puerto Rico | petioles and infructescence of Guarea guidonia | GU300084 | GQ844797 | GQ478219 | [36] |
| X. microceras | HAST 414 | Guadeloupe | wood | GU300086 | GQ844799 | GQ478221 | [36] |
| X. montagnei | HAST 495 | Martinique | wood | GU322455 | GQ848337 | GQ495948 | [36] |
| X. multiplex | JDR 259 | USA | wood | GU300099 | GQ844815 | GQ487706 | [36] |
| X. muscula | HAST 520 | Guadeloupe | dead branch | GU300087 | GQ844800 | GQ478222 | [36] |
| X. nigripes | HAST 653 | China | ground of mango orchard | GU324755 | GQ853027 | GQ502710 | [36] |
| X. ochraceostroma | HAST 401 (HT) | China | ground of mango orchard | EU179869 | GQ853034 | GQ502717 | [36,38] |
| X. oligotoma | HAST 784 | French Guiana | wood | GU300092 | GQ844808 | GQ487700 | [36] |
| X. ophiopoda | HAST 93082805 | China | bark | GU322461 | GQ848344 | GQ495955 | [36] |
| X. oxyacanthae | YMJ 1184 | Germany | seeds of Carpinus betulus | MF773430 | MF773434 | MF773438 | [5,36] |
| X. oxyacanthae | YMJ 1320 | Germany | fruits of Cornus sanguinea | MF773431 | MF773435 | MF773439 | [5,36] |
| X. palmicola | PDD 604 | New Zealand | fruits of palm | GU322436 | GQ844822 | GQ495929 | [36] |
| X. papulis | HAST 89021903 | China | wood | GU300100 | GQ844816 | GQ487707 | [36] |
| X. phyllocharis | HAST 528 | Guadeloupe | dead leaves | GU322445 | GQ844832 | GQ495938 | [36] |
| X. plebeja | HAST 91122401 | China | trunk of Machilus zuihoensis | GU324740 | GQ848353 | GQ502689 | [36] |
| X. polymorpha | JDR 1012 | USA | wood | GU322460 | GQ848343 | GQ495954 | [36] |
| X. polysporicola | FCATAS848 | China | leaves of Polyspora hainanensis | MZ005592 | MZ031980 | MZ031976 | [40] |
| X. reevesiae | HAST 90071609 (HT) | China | fruits of Reevesia formosana | GU322435 | GQ844821 | GQ495928 | [36] |
| X. regalis | HAST 920 | India | log of Ficus racemosa | GU324745 | GQ848358 | GQ502694 | [36] |
| X. rogersii | FCATAS913 | China | fruits of Magnolia sp. | MZ648825 | MZ707119 | MZ695799 | This study |
| X. rogersii | FCATAS914 | China | fruits of Magnolia sp. | MZ648826 | MZ707120 | N/A | This study |
| X. rogersii | FCATAS915 (HT) | China | fruits of Magnolia sp. | MZ648827 | MZ707121 | MZ695800 | This study |
| X. schimicola | FCATAS896 (HT) | China | fruits of Schima noronhae | MZ648850 | MZ707114 | MZ695787 | This study |
| X. schimicola | FCATAS898 | China | fruits of Schima noronhae | MZ648851 | N/A | N/A | This study |
| X. schweinitzii | HAST 92092023 | China | bark | GU322463 | GQ848346 | GQ495957 | [36] |
| X. scruposa | HAST 497 | Martinique | wood | GU322458 | GQ848341 | GQ495952 | [36] |
| X. sicula | HAST 90071613 | China | fallen leaves | GU300081 | GQ844794 | GQ478216 | [36] |
| Xylaria sp. 6 | JDR 258 | USA | leaves of Tibouchina semidecandra | GU300082 | GQ844795 | GQ478217 | [36] |
| X. striata | HAST 304 | China | branch of Punica granatum | GU300089 | GQ844803 | GQ478224 | [36] |
| X. telfairii | HAST 90081901 | China | bark | GU324738 | GQ848351 | GQ502687 | [36] |
| X. theaceicola | FCATAS903 (HT) | China | fruits of Schima villosa | MZ648848 | MZ707115 | MZ695788 | This study |
| X. theaceicola | FCATAS904 | China | fruits of Schima villosa | MZ648849 | N/A | N/A | This study |
| X. tuberoides | HAST 475 | Martinique | wood | GU300074 | GQ844784 | GQ478209 | [36] |
| X. venustula | HAST 88113002 | China | bark | GU300091 | GQ844807 | GQ487699 | [36] |
| X. vivantii | HAST 519 (HT) | Martinique | fruits of Magnolia sp. | GU322438 | GQ844824 | GQ495931 | [36] |
| X. wallichii | FCATAS923 | China | fruits of Schima wallichii | MZ648861 | MZ707118 | MZ695793 | This study |
| X. wallichii | FCATAS924 | China | fruits of Schima wallichii | MZ648862 | N/A | MZ695794 | This study |
| X. wallichii | FCATAS911 (HT) | China | fruits of Schima wallichii | ON222810 | N/A | MZ695797 | This study |
| X. xanthinovelutina | HAST 553 | Martinique | fruit of Swietenia macrophylla | GU322441 | GQ844828 | GQ495934 | [36] |
3. Results
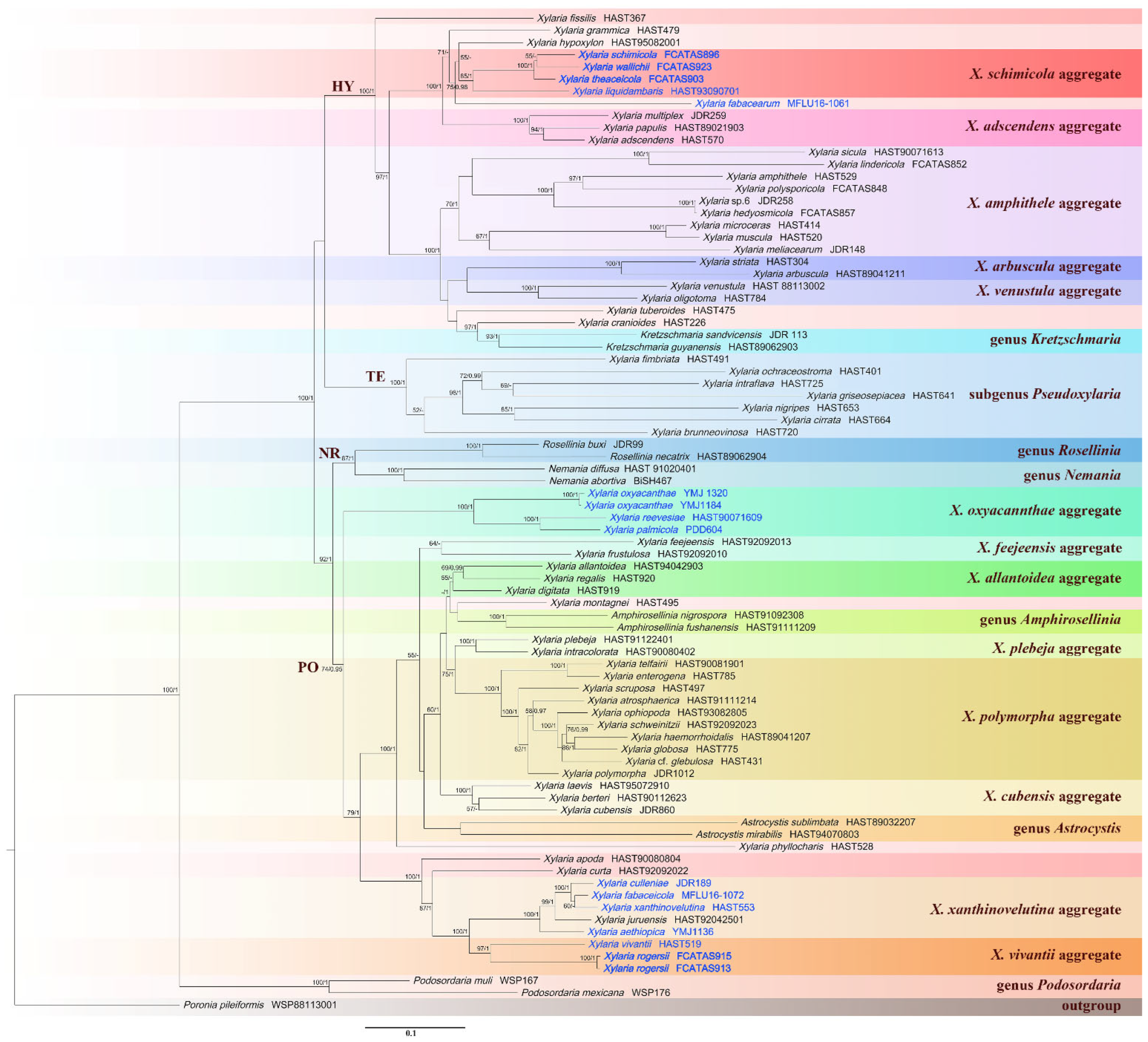
3.1. Molecular Phylogenetic Analysis
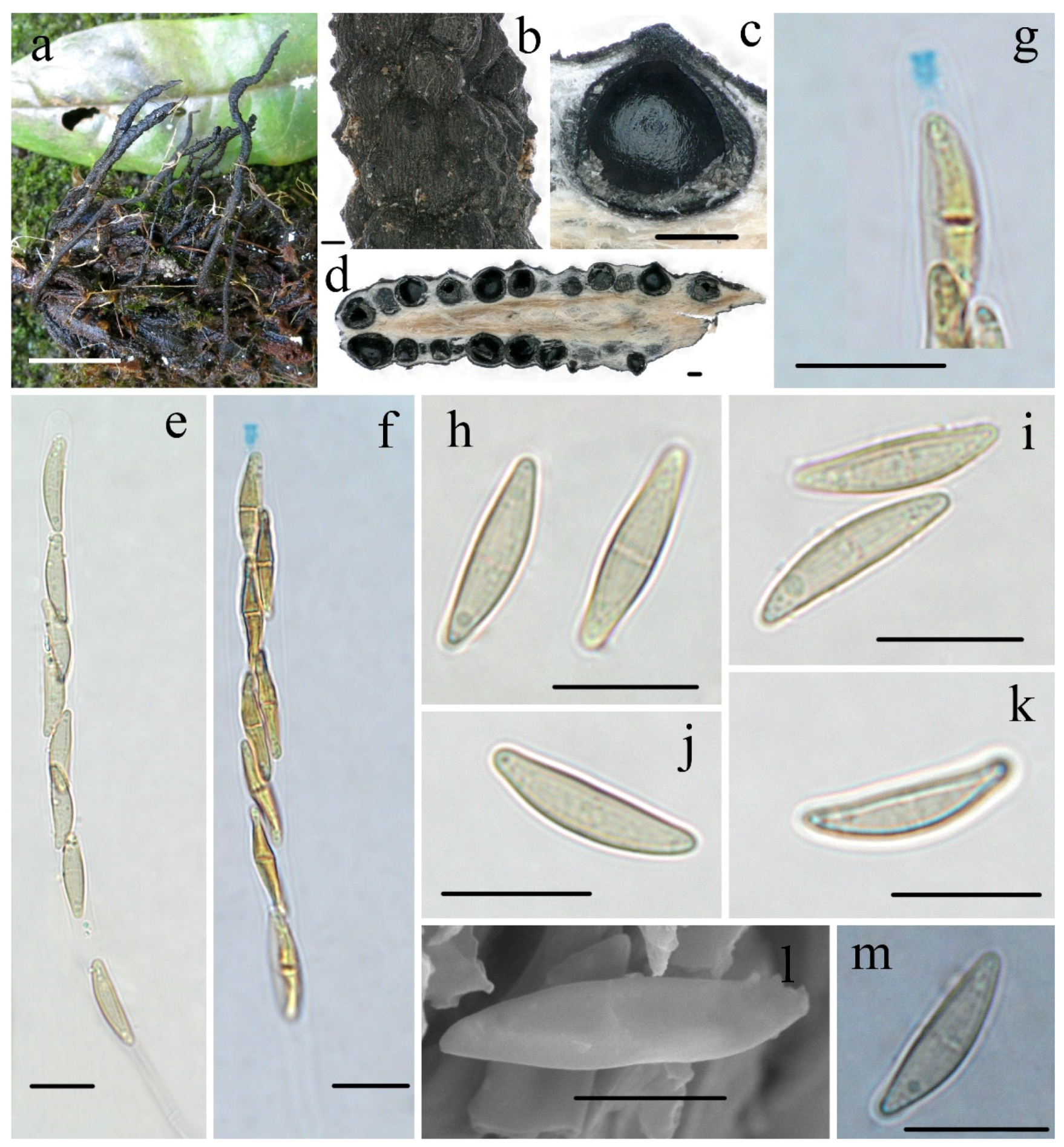
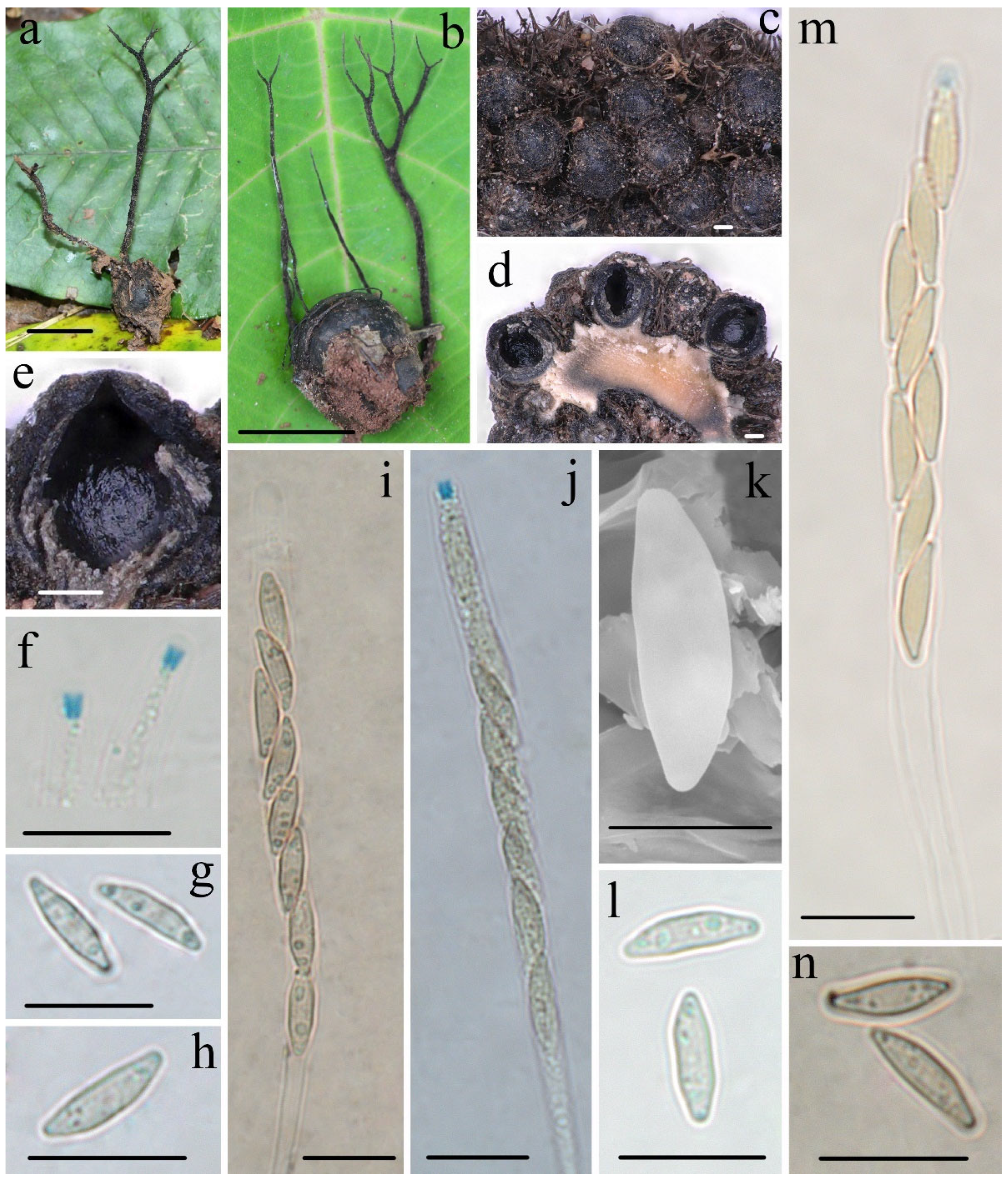
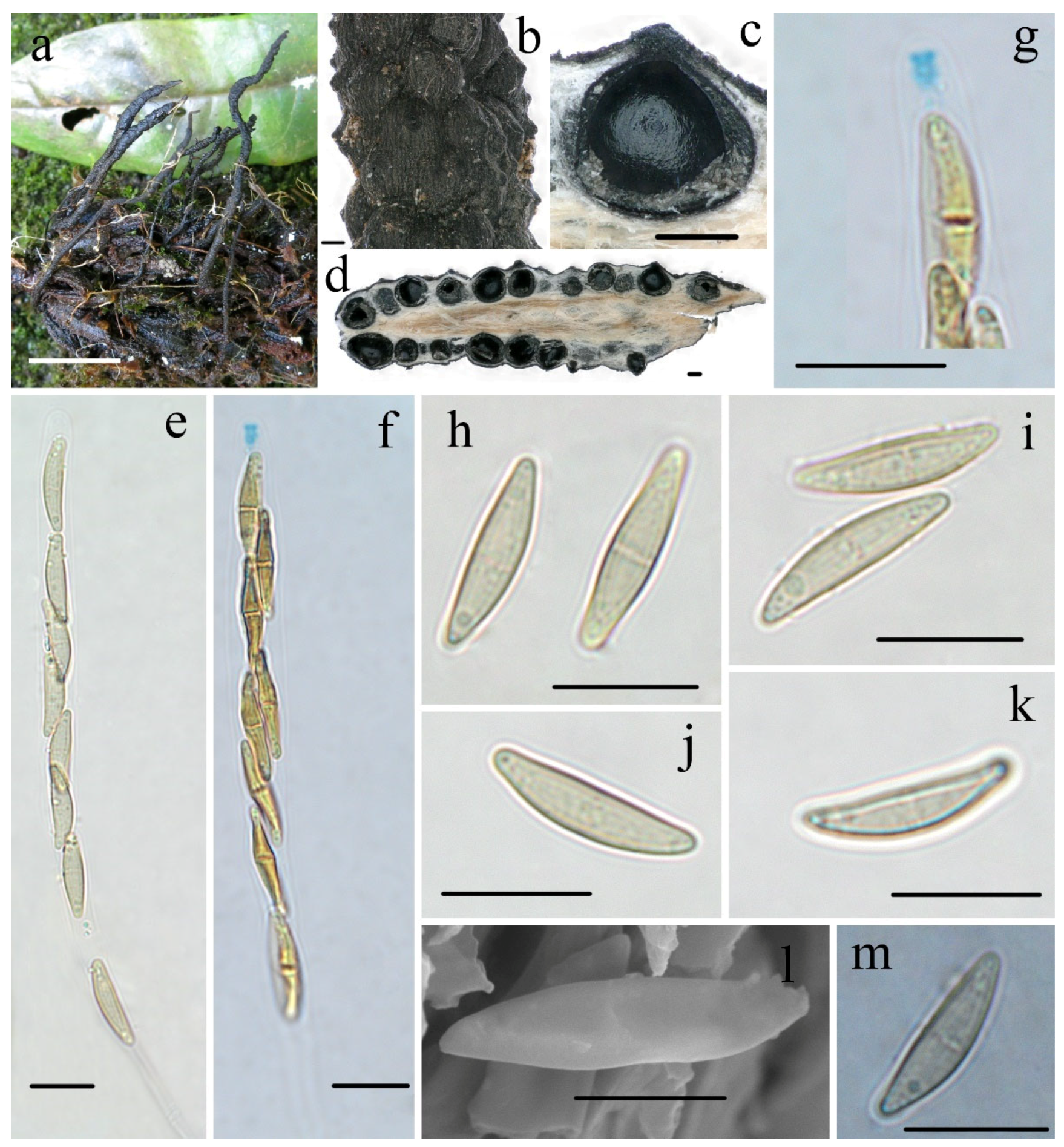
3.2. Taxonomy
4. Discussion and Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kirk, P.M.; Cannon, P.F.; Minter, D.W.; Stalpers, J.A. Dictionary of the Fungi, 10th ed.; CAB International: Wallingford, UK, 2008; p. 771. [Google Scholar]
- Index Fungorum. Available online: http://www.indexfungorum.org/names/names.asp (accessed on 1 March 2022).
- Dennis, R.W.G. Some Xylarias of tropical America. Kew Bull. 1956, 11, 401–444. [Google Scholar] [CrossRef]
- San Martín, F.; Rogers, J.D. A preliminary account of Xylaria of Mexico. Mycotaxon 1989, 34, 283–373. [Google Scholar]
- Ju, Y.M.; Rogers, J.D.; Hsieh, H.M. Xylaria species associated with fallen fruits and seeds. Mycologia 2018, 110, 726–749. [Google Scholar] [CrossRef]
- Rogers, J.D. Xylaria magnoliae sp. nov. and comments on several other fruit-inhabiting species. Can. J. Bot. 1979, 57, 941–945. [Google Scholar] [CrossRef]
- Læssøe, T.; Lodge, D.J. Three host-specific Xylaria species. Mycologia 1994, 86, 436–446. [Google Scholar] [CrossRef]
- Perera, R.H.; Hyde, K.D.; Maharachchikumbura, S.S.N.; Jones, E.B.G.; McKenzie, E.H.C.; Stadler, M.; Lee, H.B.; Samarakoon, M.C.; Ekanayaka, A.H.; Camporesi, E.; et al. Fungi on wild seeds and fruits. Mycosphere 2020, 11, 2108–2480. [Google Scholar] [CrossRef]
- Rogers, J.D.; San Martín, F.; Ju, Y.M. A reassessment of the Xylaria on Liquidambar fruits and two new taxa on Magnolia fruits. Sydowia 2002, 54, 91–97. [Google Scholar]
- Rogers, J.D.; Yeomans, R.; Adams, M.J. The relationship of Xylaria oxyacanthae to seeds of Crataegus monogyna. N. Amer. Fungi 2008, 3, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Fournier, J.; Ju, Y.M.; Hsieh, H.M.; Lindermann, U. Xylaria aethiopica sp. nov.—A new pod-inhabiting species of Xylaria (Xylariaceae) from Ethiopia. Ascomycete.org 2018, 10, 209–215. [Google Scholar]
- Husbands, D.R.; Urbina, H.; Lewis, S.M.; Aime, M.C. Xylaria karyophthora: A new seed-inhabiting fungus of Greenheart from Guyana. Mycologia 2018, 110, 434–447. [Google Scholar] [CrossRef]
- Teng, S.Q. Fungi of China; Science Press: Beijing, China, 1963; p. 808. [Google Scholar]
- Tai, F.L. Sylloge fungorum Sinicorum; Science Press: Beijing, China, 1979; p. 1527. [Google Scholar]
- Li, Y.L.; Li, H.J. A novel species of Xylaria. J. Nanjing Agric. Univ. 1994, 17, 145–147. [Google Scholar]
- Abe, Y.; Liu, Z. An annotated list of xylariaceous and diatrypaceous fungi collected from Mt. Fengyangshan and Mt. Baishanzu, Zhejiang Prov. in East China. Bul. Natl. Sci. Mus. Ser. B Bot. 1995, 21, 75–86. [Google Scholar]
- Xu, A.S. A new species of Xylaria. Mycosystema 1999, 18, 137–140. [Google Scholar]
- Zhu, Y.F.; Guo, L. Xylaria hainanensis sp. nov. (Xylariaceae) from China. Mycosystema 2011, 30, 526–528. [Google Scholar]
- Ma, H.X.; Vasilyeva, L.; Li, Y. A new species of Xylaria from China. Mycotaxon 2011, 116, 151–155. [Google Scholar] [CrossRef]
- Ma, H.X.; Vasilyeva, L.; Li, Y. The genus Xylaria (Xylariaceae) in the south of China—6. A new Xylaria species based on morphological and molecular characters. Phytotaxa 2013, 147, 48–54. [Google Scholar] [CrossRef]
- Ma, H.X.; Li, Y. Xylaria curta and X. partita (Xylariales) from Yunnan province. Austrian J. Mycol. 2017, 26, 99–105. [Google Scholar]
- Ma, H.X.; Li, Y. Xylaria crinalis and X. betulicola from China—Two new species with thread-like stromata. Sydowia 2018, 70, 37–49. [Google Scholar]
- Ma, H.X.; Qu, Z.; Peng, M.K.; Li, Y. Two penzigioid Xylaria species described from China based on morphological and molecular characters. Phytotaxa 2020, 436, 36–44. [Google Scholar] [CrossRef]
- Huang, G.; Guo, L.; Liu, N. Two new species of Xylaria and X. diminuta new to China. Mycotaxon 2014, 129, 149–152. [Google Scholar] [CrossRef]
- Rayner, R.W. A Mycological Color Chart; Cmi. & British Mycological Society Kew: London, UK, 1970. [Google Scholar]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Shinsky, J.J., White, T.J., Eds.; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar]
- Liu, Y.J.; Whelen, S.; Hall, B.D. Phylogenetic relationships among ascomycetes: Evidence from an RNA polymerse II subunit. Mol. Biol. Evol. 1999, 16, 1799–1808. [Google Scholar] [CrossRef] [PubMed]
- O’donnell, K.; Cigelnik, E. Two divergent intragenomic rDNA ITS2 types within a monophyletic lineage of the fungus Fusarium are nonorthologous. Mol. Phylogenet. Evol. 1997, 7, 103–116. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, H.M.; Ju, Y.M.; Rogers, J.D. Molecular phylogeny of Hypoxylon and related genera. Mycologia 2005, 97, 844–865. [Google Scholar] [CrossRef] [PubMed]
- Wangsawat, N.; Ju, Y.M.; Phosri, C.; Whalley, A.J.S.; Suwannasai, N. Twelve new taxa of Xylaria associated with termite nests and soil from Northeast Thailand. Biology 2021, 10, 575. [Google Scholar] [CrossRef] [PubMed]
- MAFFT V.7 Online Server. Available online: https://mafft.cbrc.jp/alignment/server/ (accessed on 12 March 2022).
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucl. Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Stamatakis, A. RAxML Version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef] [PubMed]
- Ronquist, F.; Teslenko, M.; Van Der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [Green Version]
- Rambaut, A. FigTree v1.4.2, a Graphical Viewer of Phylogenetic Trees. Available online: http://tree.bio.ed.ac.uk/software/figtree/ (accessed on 2 March 2022).
- Hsieh, H.M.; Lin, C.R.; Fang, M.J.; Rogers, J.D.; Fournier, J.; Lechat, C.; Ju, Y.M. Phylogenetic status of Xylaria subgenus Pseudoxylaria among taxa of the subfamily Xylarioideae (Xylariaceae) and phylogeny of the taxa involved in the subfamily. Mol. Phylogenet. Evol. 2010, 54, 957–969. [Google Scholar] [CrossRef] [PubMed]
- Ju, Y.M.; Rogers, J.D.; Hsieh, H.M. Amphirosellinia gen. nov. and a new species of Entoleuca. Mycologia 2004, 96, 1393–1402. [Google Scholar] [CrossRef] [PubMed]
- Ju, Y.M.; Hsieh, H.M. Xylaria species associated with nests of Odontotermes formosanus in Taiwan. Mycologia 2007, 99, 936–957. [Google Scholar] [CrossRef]
- Vu, D.; Groenewald, M.; De Vries, M.; Gehrmann, T.; Stielow, B.; Eberhardt, U.; Al-Hatmi, A.; Groenewald, J.Z.; Cardinali, G.; Houbraken, J.; et al. Large-scale generation and analysis of filamentous fungal DNA barcodes boosts coverage for kingdon fungi and reveals thresholds for fungal species and higher taxon delimitation. Stud. Mycol. 2019, 92, 135–154. [Google Scholar] [CrossRef] [PubMed]
- Pan, X.Y.; Song, Z.K.; Qu, Z.; Liu, T.D.; Ma, H.X. Three new Xylaria species (Xylariaceae, Xylariales) on fallen leaves from Hainan Tropical Rainforest National Park. Mycokeys 2022, 86, 47–63. [Google Scholar] [CrossRef] [PubMed]
- Hashemi, S.A.; Zare, R.; Khodaparast, S.A.; Elahinia, S.A. A new Xylaria species from Iran. Mycologia Iranica 2015, 2, 1–10. [Google Scholar]
- Peláez, F.; González, V.; Platas, G.; Sánchez-Ballesteros, J.; Rubio, V. Molecular phylogenetic studies within the Xylariaceae based on ribosomal DNA sequences. Fungal Divers. 2008, 31, 111–134. [Google Scholar]
- Dai, Y.C.; Yang, Z.L.; Cui, B.K.; Wu, G.; Yuan, H.S.; Zhou, L.W.; He, S.H.; Ge, Z.W.; Wu, F.; Wei, Y.L.; et al. Diversity and systematics of the important macrofungi in Chinese forests. Mycosystema 2021, 40, 770–805. [Google Scholar]
- Wang, K.; Chen, S.L.; Dai, Y.C.; Jia, Z.F.; Li, T.H.; Liu, T.Z.; Phurbu, D.; Mamut, R.; Sun, G.Y.; Bau, T.; et al. Overview of China’ s nomenclature novelties of fungi in the new century (2000–2020). Mycosystema 2021, 40, 822–833. [Google Scholar]
- Fan, N.W.; Chang, H.S.; Cheng, M.J.; Hsieh, S.Y.; Liu, T.W.; Yuan, G.F.; Chen, I.S. Secondary metabolites from the endophytic fungus Xylaria cubensis. Helv. Chim. Acta 2014, 97, 1689–1699. [Google Scholar] [CrossRef]
- Biasetto, C.R.; Somensi, A.; Abdalla, V.C.P.; Abreu, L.M.; Gualtieri, S.C.J.; Pfenning, L.H.; Bolzani, V.S.; Araujo, A.R. Phytotoxic constituents from endophytic fungus Xylaria cubensis associated with Eugenia brasiliensis. Quim. Nova 2019, 42, 485–488. [Google Scholar] [CrossRef]




| 1. Ascospores pale or subhyaline | 2 |
| 1. Ascospores brown to dark brown | 7 |
| 2. Stromata tomentose on the fertile part | 3 |
| 2. Stromata glabrous on the fertile part | 5 |
| 3. Stromata with half- to fully exposed perithecial mounds, frequently dichotomously branched; ascospores (8.5–)9–10.5(–11) × (4–)4.5–5.5(–6) µm | X. wallichii |
| 3. Stromata with inconspicuous perithecial mounds, unbranched in most cases | 4 |
| 4. Ascospores (12.5–)13.5–15(–16) × (2.5–)3–3.5(–4) µm | X. magnoliae var. magnoliae * |
| 4. Ascospores (7.5–)8–9(–10) × (2.5–)3–3.5(–4) µm | X. magnoliae var. microspore * |
| 5. Ascospores with a conspicuous straight germ slit | X. theaceicola |
| 5. Ascospores without a discernible germ slit | 6 |
| 6. Stromata associated with fruits of Magnolia (Magnoliaceae); ascospores (13.0–)13.8–15.0(–15.6) × (3.3–)3.6–4.0(–4.4) µm | X. rogersii |
| 6. Stromata associated with fruits of Schima noronhae (Theaceae); ascospores (9.5–)10.5–12.0(–13.0) × (1.6–)1.9–2.5(–3.0) µm | X. schimicola |
| 7. Stromata tomentose on the fertile part | 8 |
| 7. Stromata glabrous on the fertile part | 15 |
| 8. Ascospores lacking a hyaline sheath and appendages | 9 |
| 8. Ascospores surrounded by a hyaline sheath and bearing non-cellular appendages at ends | 10 |
| 9. Stromata associated with fruits of Apeiba (Tiliaceae); ascospores (9.5–)10–12(–13) × (3–) 3.5–4(–4.5) µm | X. apeibae * |
| 9. Stromata associated with fruits of Bauhinia cumingniana (Fabaceae); ascospores (8–)8.5–9.5(–10) × 3–3.5(–4) µm | X. luzonensis * |
| 10. Ascospores with an oblique germ slit | 11 |
| 10. Ascospores with a straight germ slit | 12 |
| 11. Stromata associated with fruits of Magnolia (Magnoliaceae); ascospores brown to dark-brown, ellipsoidal-inequilateral, (14.5–)15–16.5(–17.5) × 4.5–5.5(–6) µm | X. vivantii * |
| 11. Stromata associated with fruits of Elizabetha pod; ascospores brown, fusoid-inequilateral, (14–)14.5–16(–17) × (3.5–)4–4.5(–5) µm | X. rossmanae * |
| 12. Ascospores (11–)11.5–13.5(–14.5) × (4–)4.5–5(–5.5) µm | X. patrisiae * |
| 12. Ascospores mostly smaller than 11.5 µm | 13 |
| 13. Stromata associated with capsules of Cullenia excelsa (Malvaceae); ascospores (7.5–)8–9(–9.5) × (3.5–)4–4.5(–5) µm | X. culleniae * |
| 13. Stromata associated with other substrates | 14 |
| 14. Ascospores (9–)9.5–11(–12) × (3.5–) 4–4.5(–5) µm | X. xanthinovelutina * |
| 14. Ascospores 7.5–10 × 3.4–4.8 µm | X. fabaceicola ** |
| 15. Stromata capitate, subglobose, or obconical | 16 |
| 15. Stromata cylindrical to filiform | 18 |
| 16. Stromata associated with decaying leaves of Clusia (Clusiaceae); ascospores (11.6–)12.8–16.7(–18) × 8–15 µm | X. clusiae * |
| 16. Stromata associated with fruits of other substrates | 17 |
| 17. Stromata associated with fruits of Gluazuma; ascospores (15–)15.5–16.5(–17) × (5.5–)6–7(–7.5) µm | X. guazumae * |
| 17. Stromata associated with different substrates; ascospores (14.5–)15.5–18(–19) × (5–)5.5–6.5(–7) µm | X. heloidea * |
| 18. Ascospores surrounded by a hyaline sheath and bearing non-cellular appendages at ends | X. aethiopica# |
| 18. Ascospores lacking a hyaline sheath | 19 |
| 19. Stromata filiform | 20 |
| 19. Stromata cylindrical | 22 |
| 20. Stromata growing on leaves and fruit remains of Quercus polymorpha (Fagaceae); ascospores 12–14.5(–16) × 4–4.5(–5) µm | X. duranii * |
| 20. Stromata associated with fruits or seeds | 21 |
| 21. Stromata associated with dead angiospermous seeds; ascospores (11.5–)13–15(–15.5) × (4.5–)5–5.5(–6) µm | X. himalayensis * |
| 21. Stromata associated with Sloanea capsules; ascospores (9.5–)10–11.5(–12.5) × (3.5–)4–4.5(–5) µm | X. warburgii * |
| 22. Stromata associated with pericarps or pods | 23 |
| 22. Stromata associated with fruits or seeds | 24 |
| 23. Stromata associated with pericarps Fagus sylvatica (Fagaceae); ascospores (9.5–)11–12(–13) × (4–)4.5–5(–5.5) µm | X. carpophila * |
| 23. Stromata associated with decaying seed pods of Fabaceae; ascospores 9–11.2 × 3–4.3 µm | X. fabacearum ** |
| 24. Stromata lacking a striped outer layer | 25 |
| 24. Stromata overlain with a striped outer layer | 27 |
| 25. Stromata with perithecial mounds fully exposed; ascospores (9.5–)10.5–11.5(–12.5) × (5–)5.5–6.5(–7) µm | X. jaliscoensis * |
| 25. Stromata with perithecial mounds inconspicuous to protuberant | 26 |
| 26. Stromata associated with seeds of Chlorocardium rodiei (Lauraceae); ascospores (8–)8.5–9.5(–10.5) × 3.5–4(–4.5) µm | X. karyophthora## |
| 26. Stromata associated with fallen fruits of Beilschmiedia percoriacea (Lauraceae); ascospores (11–)12–14 × 4–5(–6) µm | X. beilschmiediae### |
| 27. Ascospores with a spiral germ slit, (13–)13.5–15(–16) × (4.5–)5–5.5(–6) µm; on fruits of Liquidambar (Altingiaceae) | X. liquidambaris * |
| 27. Ascospores with a straight germ slit | 28 |
| 28. Stromata associated with fallen fruits | 29 |
| 28. Stromata associated with seeds | 30 |
| 29. Stromata associated with fallen fruits of Euphorbia (Euphorbiaceae); ascospores (8–)8.5–9.5(–10) × (3.5–)4–5(–5.5) µm | X. euphorbiicola * |
| 29. Stromata associated with fallen fruits of Reevesia formosana (Sterculiaceae); ascospores (8.5–)9–10.5(–11) × (4–)4.5–5.5(–6) µm | X. reevesiae * |
| 30. Stromata associated with palm seeds | 31 |
| 30. Stromata associated with seeds of dicots | 32 |
| 31. Stromata associated with seeds of Euterpe globosa; ascospores (13.5–)14.5–16.5(–18.5) × (6–)6.5–7.5(–8.5) µm | X. palmicola * |
| 31. Stromata associated with palm seeds; ascospores (9.5–)10.5–12(–13.5) × (4–)4.5–5.5(–6) µm | X. rhizocola * |
| 32. Stromata associated with seeds of Psidium guajava (Myrtaceae); ascospores (8.5–)9–10.5(–12) × (4–)4.5–5(–5.5) µm | X. psidii * |
| 32. Stromata associated with seeds of various plants; ascospores (9.5–)10–11.5(–12) × (4–)4.5–5.5(–6) µm | X. oxyacanthae * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, H.; Song, Z.; Pan, X.; Qu, Z.; Yang, Z.; Li, Y.; Zhu, A. Four New Pale-Spored Species of Xylaria (Xylariaceae, Xylariales) with a Key to Worldwide Species on Fallen Fruits and Seeds. Biology 2022, 11, 885. https://doi.org/10.3390/biology11060885
Ma H, Song Z, Pan X, Qu Z, Yang Z, Li Y, Zhu A. Four New Pale-Spored Species of Xylaria (Xylariaceae, Xylariales) with a Key to Worldwide Species on Fallen Fruits and Seeds. Biology. 2022; 11(6):885. https://doi.org/10.3390/biology11060885
Chicago/Turabian StyleMa, Haixia, Zikun Song, Xiaoyan Pan, Zhi Qu, Zhanen Yang, Yu Li, and Anhong Zhu. 2022. "Four New Pale-Spored Species of Xylaria (Xylariaceae, Xylariales) with a Key to Worldwide Species on Fallen Fruits and Seeds" Biology 11, no. 6: 885. https://doi.org/10.3390/biology11060885
APA StyleMa, H., Song, Z., Pan, X., Qu, Z., Yang, Z., Li, Y., & Zhu, A. (2022). Four New Pale-Spored Species of Xylaria (Xylariaceae, Xylariales) with a Key to Worldwide Species on Fallen Fruits and Seeds. Biology, 11(6), 885. https://doi.org/10.3390/biology11060885