Proteolyzed Variant of IgG with Free C-Terminal Lysine as a Biomarker of Prostate Cancer

 ,
, 
 , , and
, , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Patients
2.2. Plasminogen Preparation
2.3. ELISA
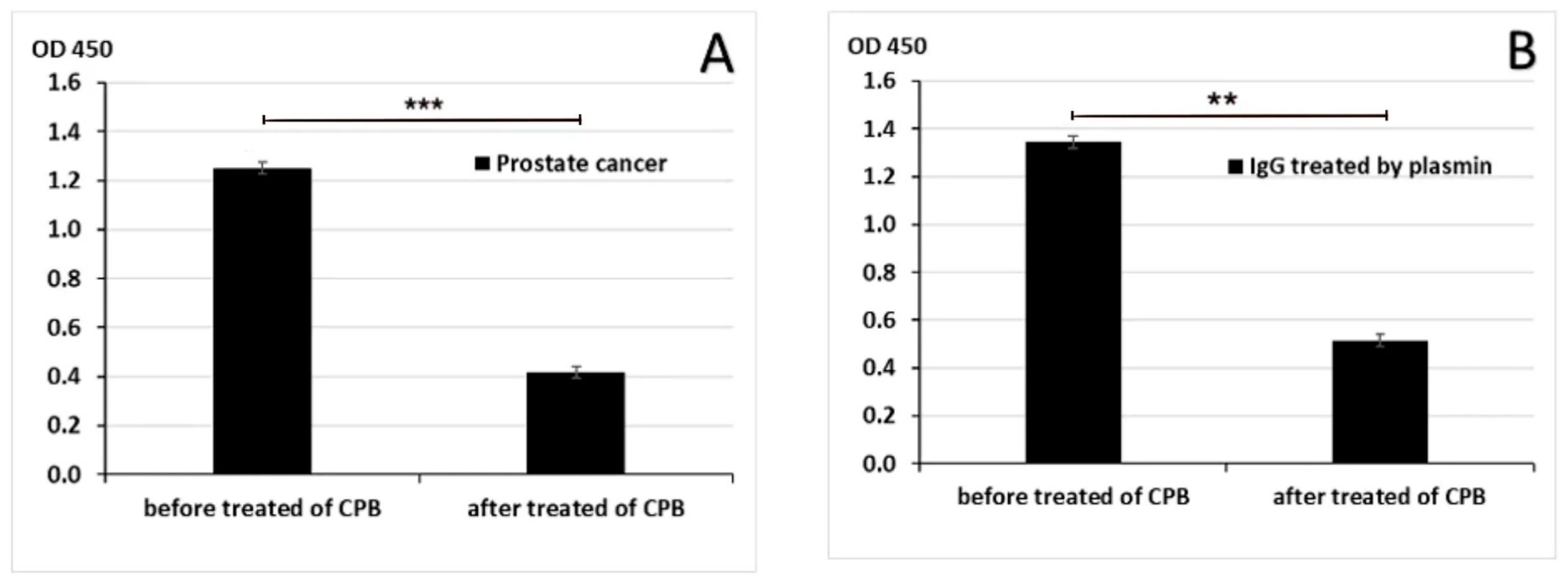
2.4. Elimination of C-Terminal Lysine Residues with Carboxypeptidase B (CPB)
3. Results
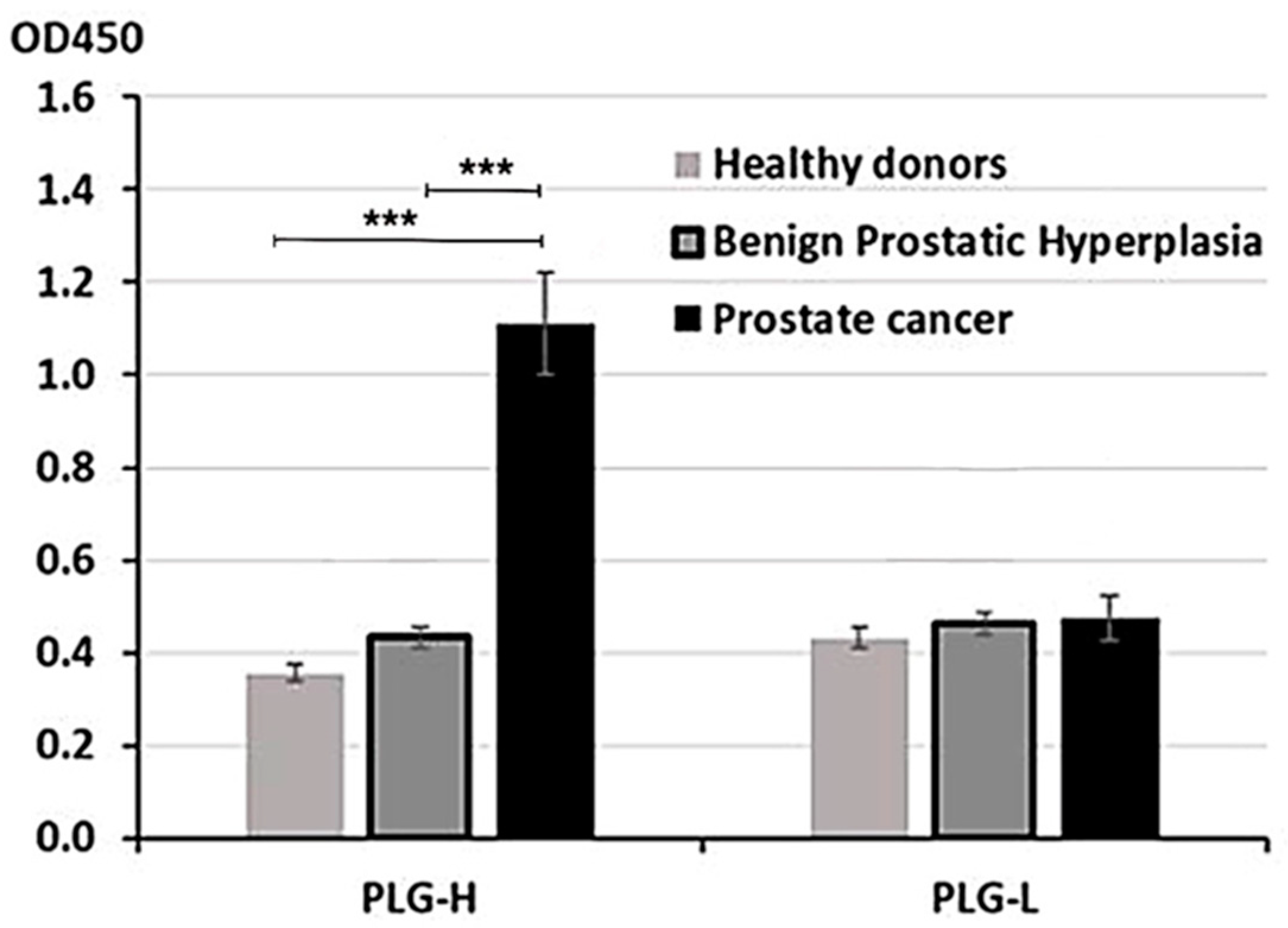
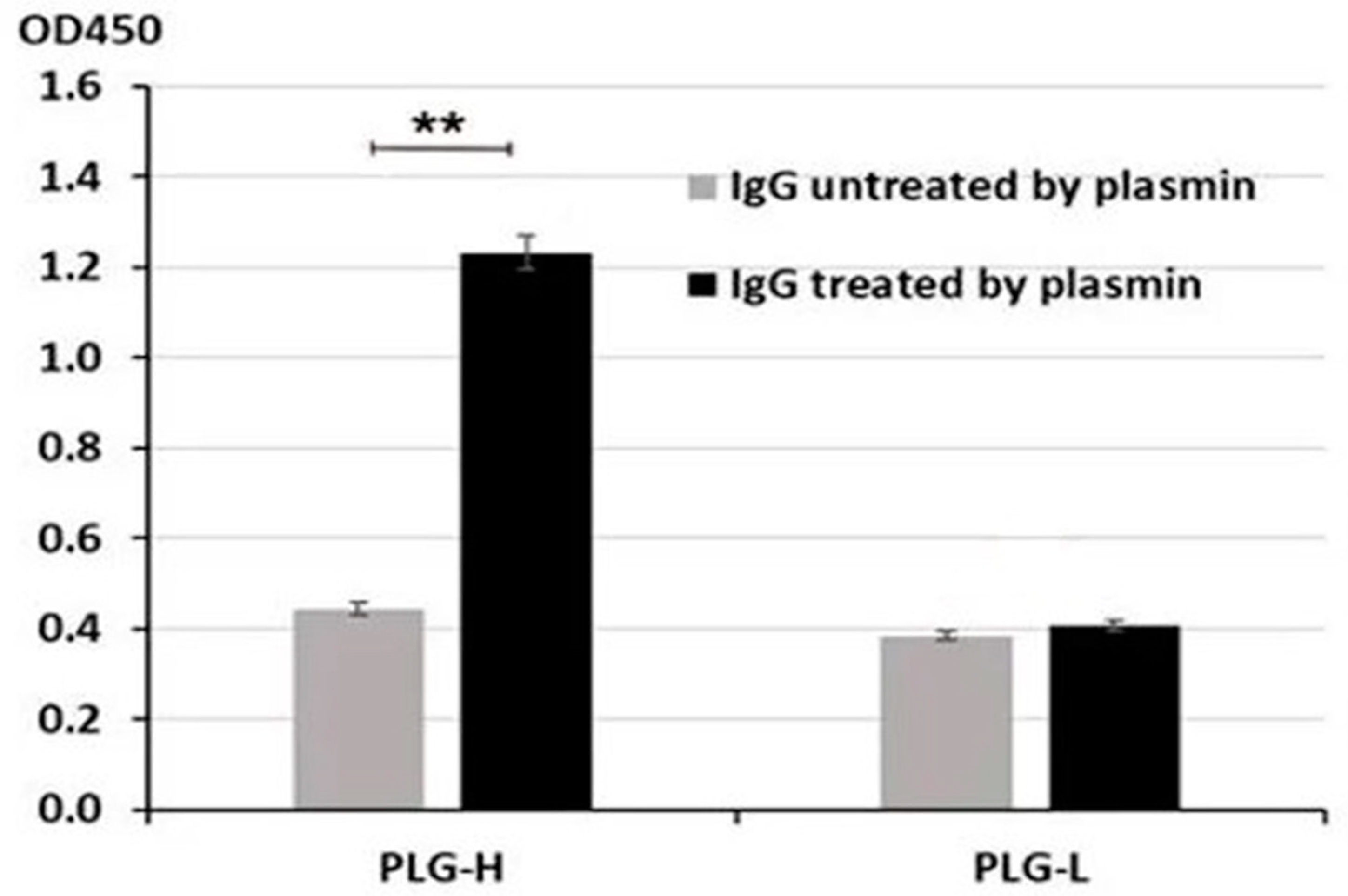
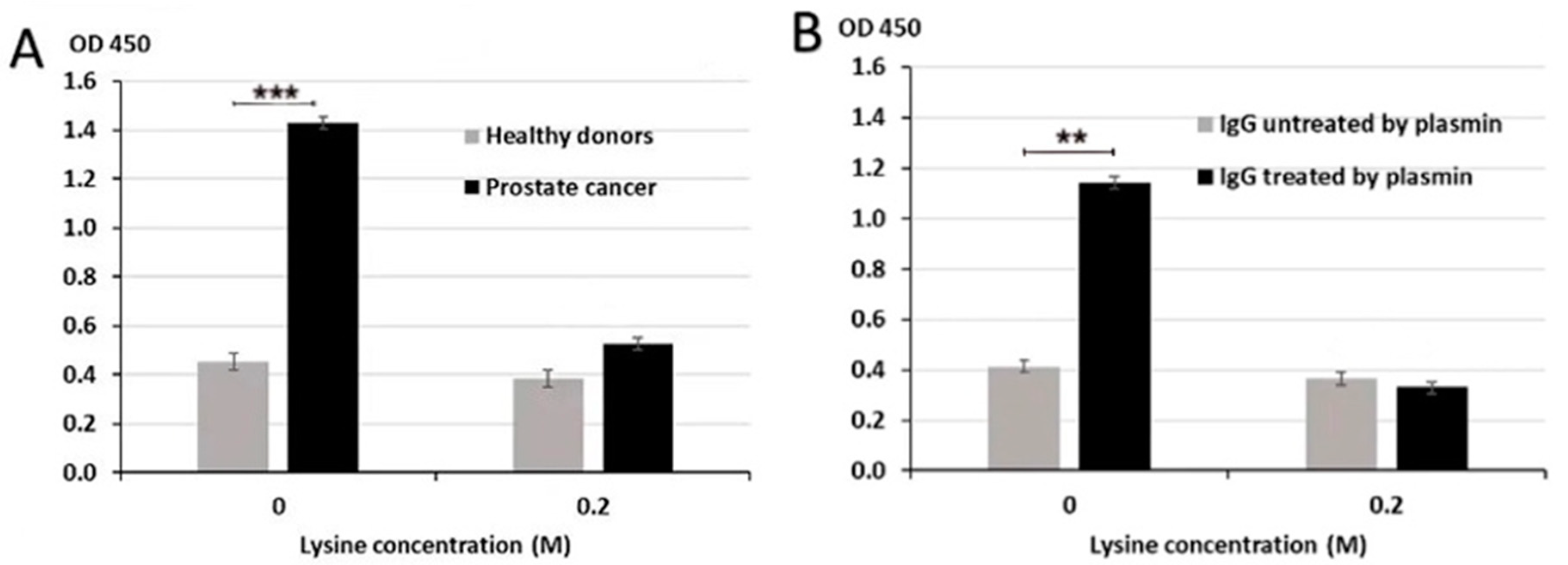
3.1. Blood IgG-Lys in PC, BPH, and Healthy
3.2. Receiver Operating Characteristic (ROC) Curve Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics. CA Cancer J. Clin. 2020, 70, 7–30. [Google Scholar] [CrossRef]
- Kirby, R.S.; Fitzpatrick, J.M.; Irani, J. Prostate cancer diagnosis in the new millennium: Strengths and weaknesses of prostate-specific antigen and the discovery and clinical evaluation of prostate cancer gene 3 (PCA3). BJU Int. 2009, 103, 441–445. [Google Scholar] [CrossRef]
- Wilbur, J. Prostate cancer screening: The continuing controversy. Am. Fam. Physician 2008, 78, 1377–1384. [Google Scholar] [PubMed]
- Schmid, M.; Trinh, Q.-D.; Graefen, M.; Fisch, M.; Chun, F.; Hansen, J. The role of biomarkers in the assessment of prostate cancer risk prior to prostate biopsy: Which markers matter and how should they be used? World J. Urol. 2014, 32, 871–880. [Google Scholar] [CrossRef]
- McNally, C.J.; Ruddock, M.W.; Moore, T.; McKenna, D.J. Biomarkers That Differentiate Benign Prostatic Hyperplasia from Prostate Cancer: A Literature Review. Cancer Manag. Res. 2020, 12, 5225–5241. [Google Scholar] [CrossRef] [PubMed]
- Grubb, R.L., III. Prostate Cancer: Update on Early Detection and New Biomarkers. Mo. Med. 2018, 115, 132–134. [Google Scholar]
- Otero, J.R.; Gomez, B.G.; Juanatey, F.C.; Touijer, K.A. Prostate cancer biomarkers: An update. Urol. Oncol. Semin. Orig. Investig. 2014, 32, 252–260. [Google Scholar] [CrossRef]
- Pepper, M.S. Role of the Matrix Metalloproteinase and Plasminogen Activator–Plasmin Systems in Angiogenesis. Arter. Thromb. Vasc. Biol. 2001, 21, 1104–1117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chana-Muñoz, A.; Jendroszek, A.; Sønnichsen, M.; Wang, T.; Ploug, M.; Jensen, J.K.; Andreasen, P.A.; Bendixen, C.; Panitz, F. Origin and diversification of the plasminogen activation system among chordates. BMC Evol. Biol. 2019, 19, 27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahmood, N.; Mihalcioiu, C.; Rabbani, S.A. Multifaceted Role of the Urokinase-Type Plasminogen Activator (uPA) and Its Receptor (uPAR): Diagnostic, Prognostic, and Therapeutic Applications. Front. Oncol. 2018, 8, 24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Behrendt, N.; Høyer-Hansen, G.; Johnsen, M.; Lund, L.; Ploug, M.; Rømer, J.; Danø, K. Plasminogen activation and cancer. Thromb. Haemost. 2005, 93, 676–681. [Google Scholar] [CrossRef]
- Parfyonova, Y.; Plekhanova, O.S.; Tkachuk, V.A. Plasminogen activators in vascular remodeling and angiogenesis. Biochemistry 2002, 67, 119–134. [Google Scholar] [CrossRef] [PubMed]
- De Visser, K.E.; Eichten, A.; Coussens, L.M. Paradoxical roles of the immune system during cancer development. Nat. Rev. Cancer 2006, 6, 24–37. [Google Scholar] [CrossRef]
- Brezski, R.J.; Jordan, R.E. Cleavage of IgGs by proteases associated with invasive diseases: An evasion tactic against host immunity? mAbs 2010, 2, 212–220. [Google Scholar] [CrossRef] [Green Version]
- Morgan, E.L.; Hugli, T.E.; Weigle, W.O. Isolation and identification of a biologically active peptide derived from the CH3 domain of human IgG1. Proc. Natl. Acad. Sci. USA 1982, 79, 5388–5391. [Google Scholar] [CrossRef] [Green Version]
- Harpel, P.C.; Sullivan, R.; Chang, T.S. Binding and activation of plasminogen on immobilized immunoglobulin G. Identifi-cation of the plasmin-derived Fab as the plasminogen-binding fragment. J. Biol. Chem. 1989, 264, 616–624. [Google Scholar] [CrossRef]
- Goufman, E.I.; Iakovlev, V.N.; Tikhonova, N.B.; Lokshin, A.E. Quantification of autoantibodies to plasminogen in plasma of patients with cancer. Cancer Biomark. 2015, 15, 281–287. [Google Scholar] [CrossRef]
- van Winden, A.W.J.; Broek, I.V.D.; Gast, M.-C.W.; Engwegen, J.Y.M.N.; Sparidans, R.W.; van Dulken, E.J.; Depla, A.C.T.M.; Cats, A.; Schellens, J.H.M.; Peeters, P.H.M.; et al. Serum Degradome Markers for the Detection of Breast Cancer. J. Proteome Res. 2010, 9, 3781–3788. [Google Scholar] [CrossRef] [PubMed]
- Grozdanić, M.; Vidmar, R.; Vizovišek, M.; Fonović, M. Degradomics in Biomarker Discovery. Proteom. Clin. Appl. 2019, 13, e1800138. [Google Scholar] [CrossRef]
- Deutsch, D.G.; Mertz, E.T. Plasminogen: Purification from Human Plasma by Affinity Chromatography. Science 1970, 170, 1095–1096. [Google Scholar] [CrossRef] [PubMed]
- Aisina, R.B.; Mukhametova, L.I.; Gershkovich, K.B.; Yakovlev, V.N.; Goufman, E.I.; Tikhonova, N.B. Effect of Specific Cleavage of Immunoglobulin G by Plasmin on the Binding and Activation of Plasminogen. Russ. J. Bioorganic Chem. 2018, 44, 210–216. [Google Scholar] [CrossRef]
- Goufman, E.I.; Yakovlev, V.N.; Tikhonova, N.B.; Aisina, R.B.; Yarygin, K.N.; Mukhametova, L.I.; Gershkovich, K.B.; Gulin, D.A. Autoantibodies to Plasminogen and Their Role in Tumor Diseases. Bull. Exp. Biol. Med. 2015, 158, 493–496. [Google Scholar] [CrossRef] [PubMed]
- DeClerck, Y.A.; Mercurio, A.M.; Stack, M.S.; Chapman, H.A.; Zutter, M.M.; Muschel, R.J.; Raz, A.; Matrisian, L.M.; Sloane, B.F.; Noel, A.; et al. Proteases, Extracellular Matrix, and Cancer: A Workshop of the Path B Study Section. Am. J. Pathol. 2004, 164, 1131–1139. [Google Scholar] [CrossRef]
- Capello, M.; Ferri-Borgogno, S.; Cappello, P.; Novelli, F. α-enolase: A promising therapeutic and diagnostic tumor target. FEBS J. 2011, 278, 1064–1074. [Google Scholar] [CrossRef] [Green Version]
- Ather, M.H.; Abbas, F.; Faruqui, N.; Israr, M.; Pervez, S. Correlation of three immunohistochemically detected markers of neuroendocrine differentiation with clinical predictors of disease progression in prostate cancer. BMC Urol. 2008, 8, 21–25. [Google Scholar] [CrossRef] [Green Version]
- Foss, C.A.; Mease, R.C.; Cho, S.Y.; Kim, H.J.; Pomper, M.G. GCPII Imaging and Cancer. Curr. Med. Chem. 2012, 19, 1346–1359. [Google Scholar] [CrossRef]
- Hupe, M.C.; Philippi, C.; Roth, D.; Kümpers, C.; Ribbat-Idel, J.; Becker, F.; Joerg, V.; Duensing, S.; Lubczyk, V.H.; Kirfel, J.; et al. Expression of Prostate-Specific Membrane Antigen (PSMA) on Biopsies Is an Independent Risk Stratifier of Prostate Cancer Patients at Time of Initial Diagnosis. Front. Oncol. 2018, 8, 623. [Google Scholar] [CrossRef]
- Stephan, C.; Ralla, B.; Jung, K. Prostate-specific antigen and other serum and urine markers in prostate cancer. Biochim. Biophys. Acta Bioenerg. 2014, 1846, 99–112. [Google Scholar] [CrossRef]
- Slatkoff, S.; Gamboa, S.; Zolotor, A.J.; Mounsey, A.L.; Jones, K. PURLs: PSA testing: When it’s useful, when it’s not. J. Fam. Pract. 2011, 60, 357–360. [Google Scholar] [PubMed]
- Löfman, O.; Lindahl, T.; Varenhorst, E. Age-specific reference values for serum prostate-specific antigen in a community-based population of healthy Swedish men. Scand. J. Clin. Lab. Investig. 1997, 57, 225–232. [Google Scholar] [CrossRef] [PubMed]
- Prcic, A.; Begic, E.; Hiros, M. Usefulness of Total PSA Value in Prostate Diseases Diagnosis. Acta Inform. Medica 2016, 24, 156–161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chapple, C. Medical treatment for benign prostatic hyperplasia. BMJ 1992, 304, 1198–1199. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cohort | Age, Yearsold (Median) | n | tPSA, min-max, ng/mL | Staging | |
|---|---|---|---|---|---|
| Gleason Score | Stage | ||||
| Healthy controls | 45–67 (62) | 30 | 0–1.5 | - | - |
| BPH | 51–65 (59) | 30 | 6–21 | - | - |
| PC | 53–67 (57) | 30 | 6–31.2 | 3 + 4, n = 17 4 + 3, n = 13 | T1, n = 6 T2a, n = 17 T3a, n = 7 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lokshin, A.; Mikhaleva, L.M.; Goufman, E.I.; Boltovskaya, M.N.; Tikhonova, N.B.; Stepanova, I.I.; Stepanov, A.A.; Potoldykova, N.V.; Vinarov, A.Z.; Stemmer, P.; et al. Proteolyzed Variant of IgG with Free C-Terminal Lysine as a Biomarker of Prostate Cancer. Biology 2021, 10, 817. https://doi.org/10.3390/biology10080817
Lokshin A, Mikhaleva LM, Goufman EI, Boltovskaya MN, Tikhonova NB, Stepanova II, Stepanov AA, Potoldykova NV, Vinarov AZ, Stemmer P, et al. Proteolyzed Variant of IgG with Free C-Terminal Lysine as a Biomarker of Prostate Cancer. Biology. 2021; 10(8):817. https://doi.org/10.3390/biology10080817
Chicago/Turabian StyleLokshin, Anna, Lyudmila M. Mikhaleva, Eugene I. Goufman, Marina N. Boltovskaya, Natalia B. Tikhonova, Irina I. Stepanova, Alexandr A. Stepanov, Natalia V. Potoldykova, Andrey Z. Vinarov, Paul Stemmer, and et al. 2021. "Proteolyzed Variant of IgG with Free C-Terminal Lysine as a Biomarker of Prostate Cancer" Biology 10, no. 8: 817. https://doi.org/10.3390/biology10080817
APA StyleLokshin, A., Mikhaleva, L. M., Goufman, E. I., Boltovskaya, M. N., Tikhonova, N. B., Stepanova, I. I., Stepanov, A. A., Potoldykova, N. V., Vinarov, A. Z., Stemmer, P., & Iakovlev, V. (2021). Proteolyzed Variant of IgG with Free C-Terminal Lysine as a Biomarker of Prostate Cancer. Biology, 10(8), 817. https://doi.org/10.3390/biology10080817