
Adaptation of an Invasive Pest to Novel Environments: Life History Traits of Drosophila suzukii in Coastal and Mainland Areas of Greece during Overwintering

 ,
, 



Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Rearing of Flies
2.2. Production of Summer and Winter Morphs
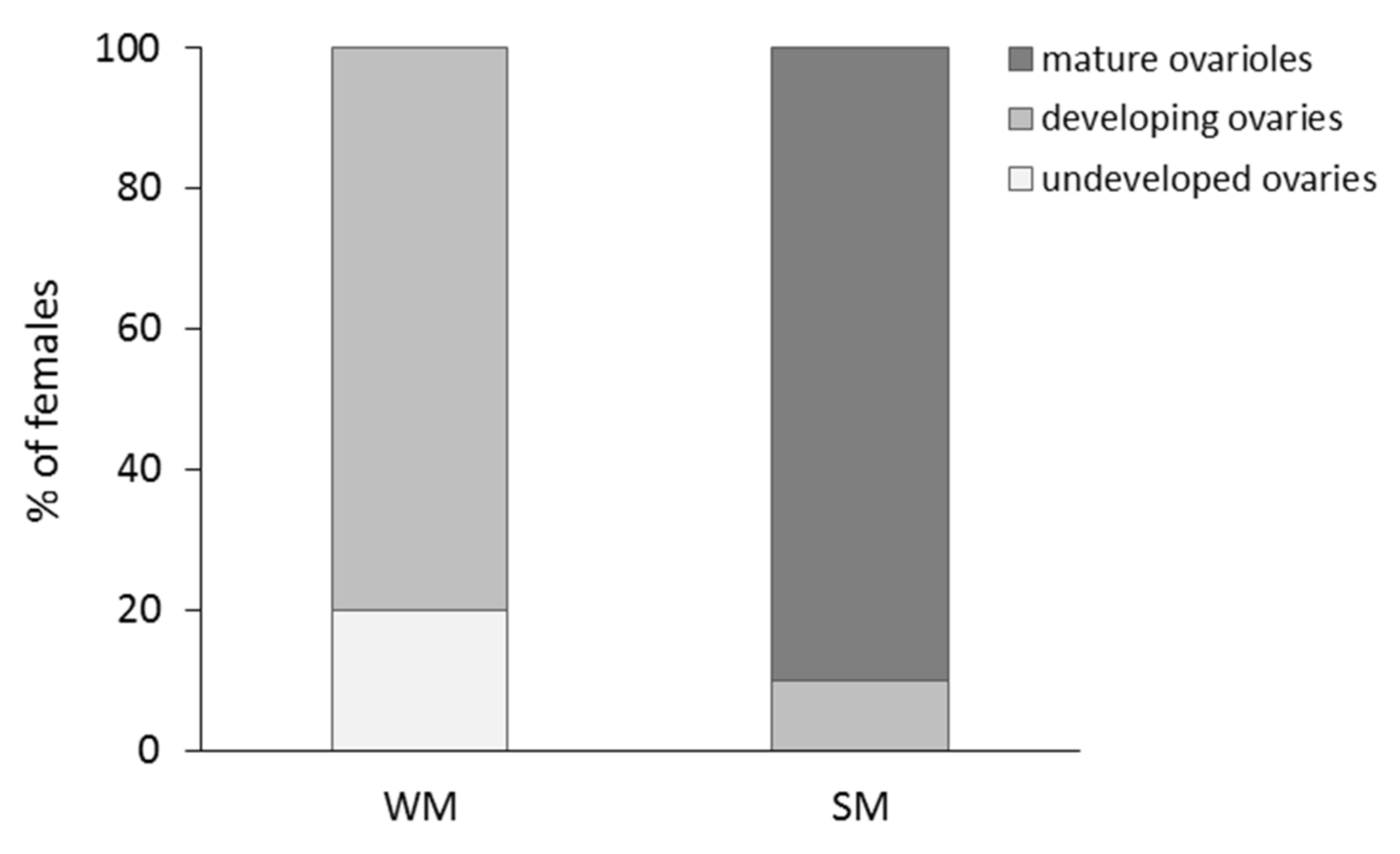
2.3. Mating and Assessment of Ovarian Development of SM and WM Females
2.4. Climatic Conditions Prevailing in the Mainland and the Coastal Area
2.5. Exposure of Flies to Field Conditions: Recording Survival Rates and Progeny Production during Overwintering
2.6. Statistical Analysis
3. Results
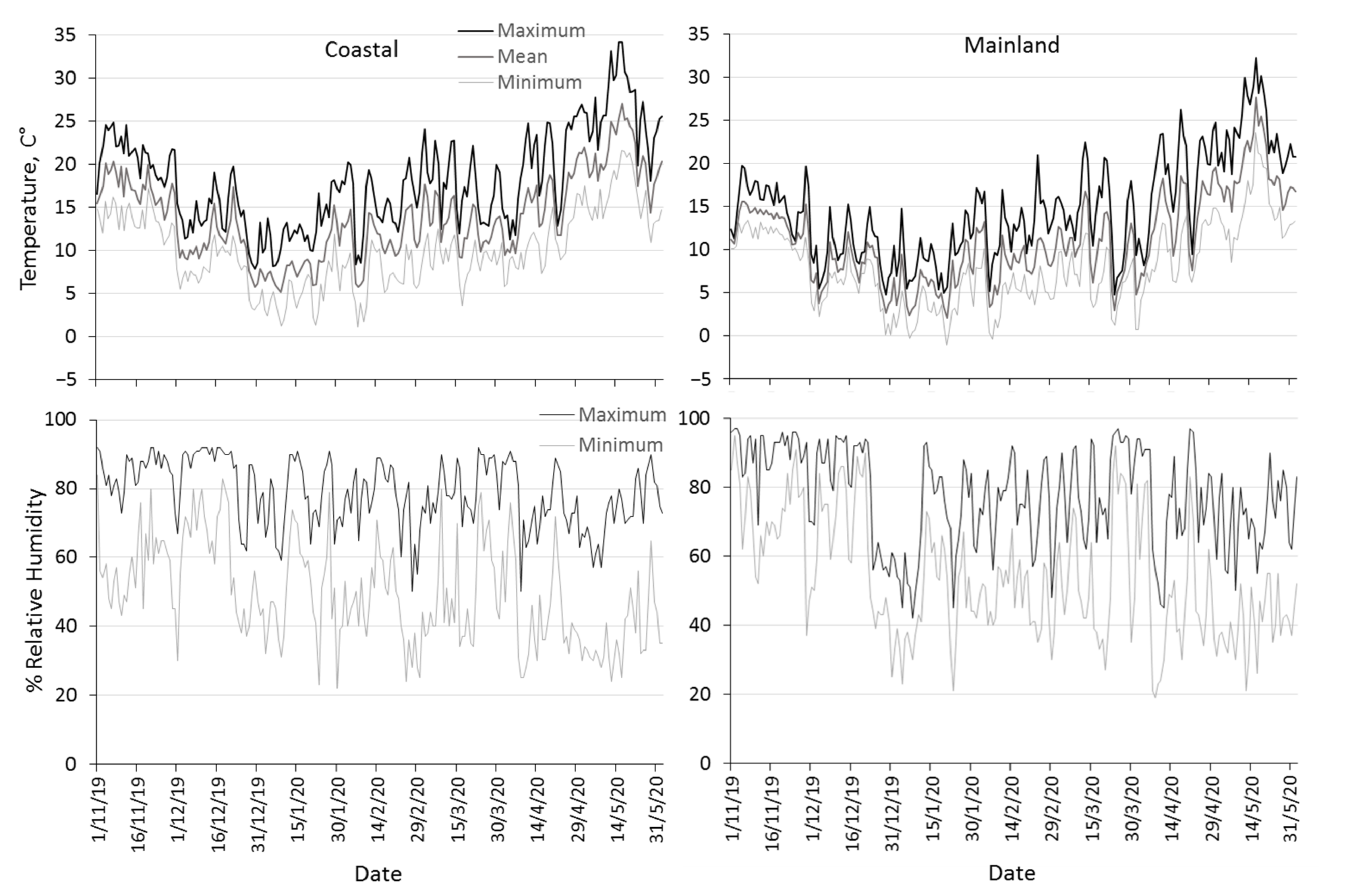
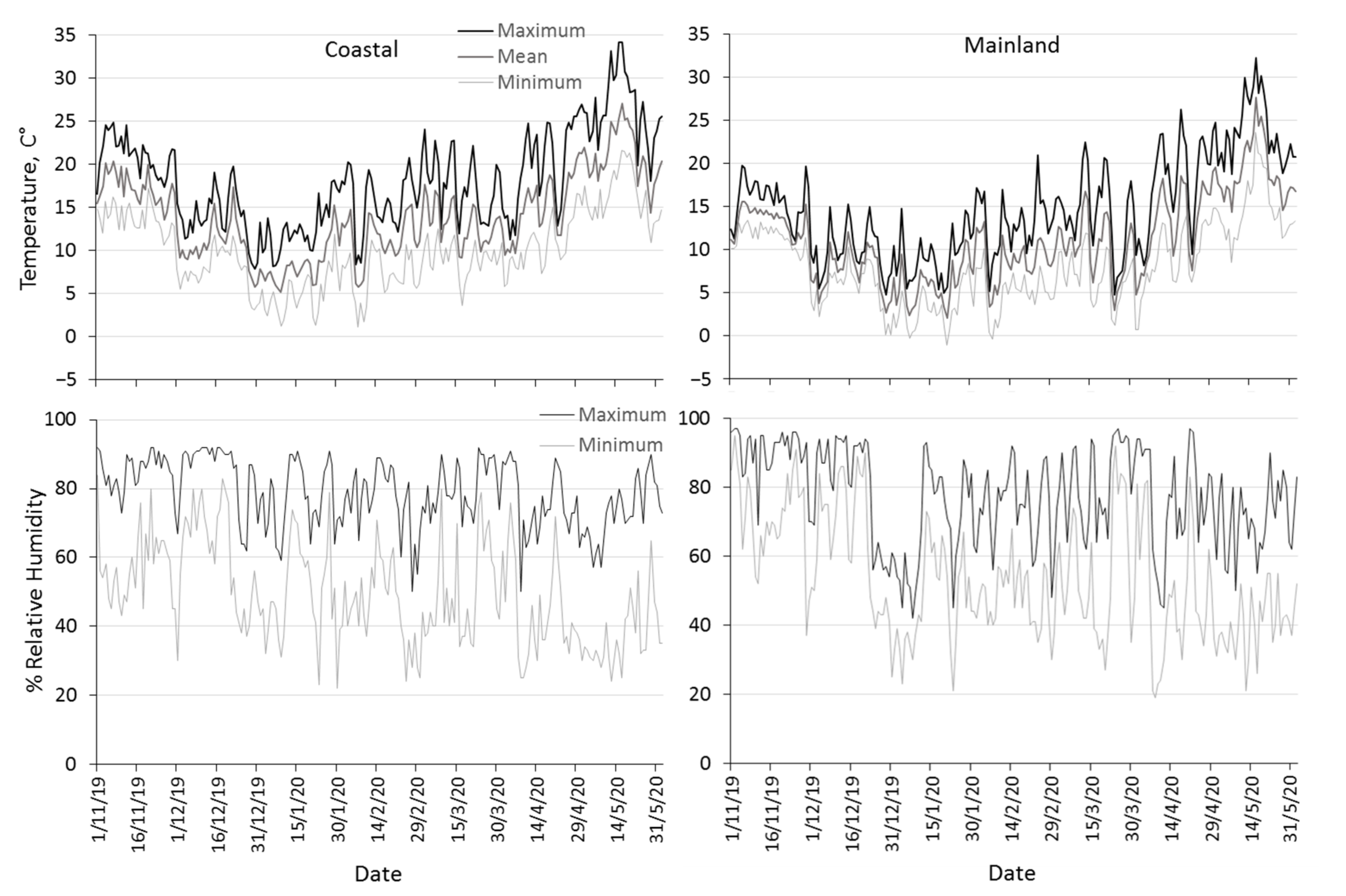
3.1. Climatic Conditions Prevailing in the Mainland and the Coastal Area
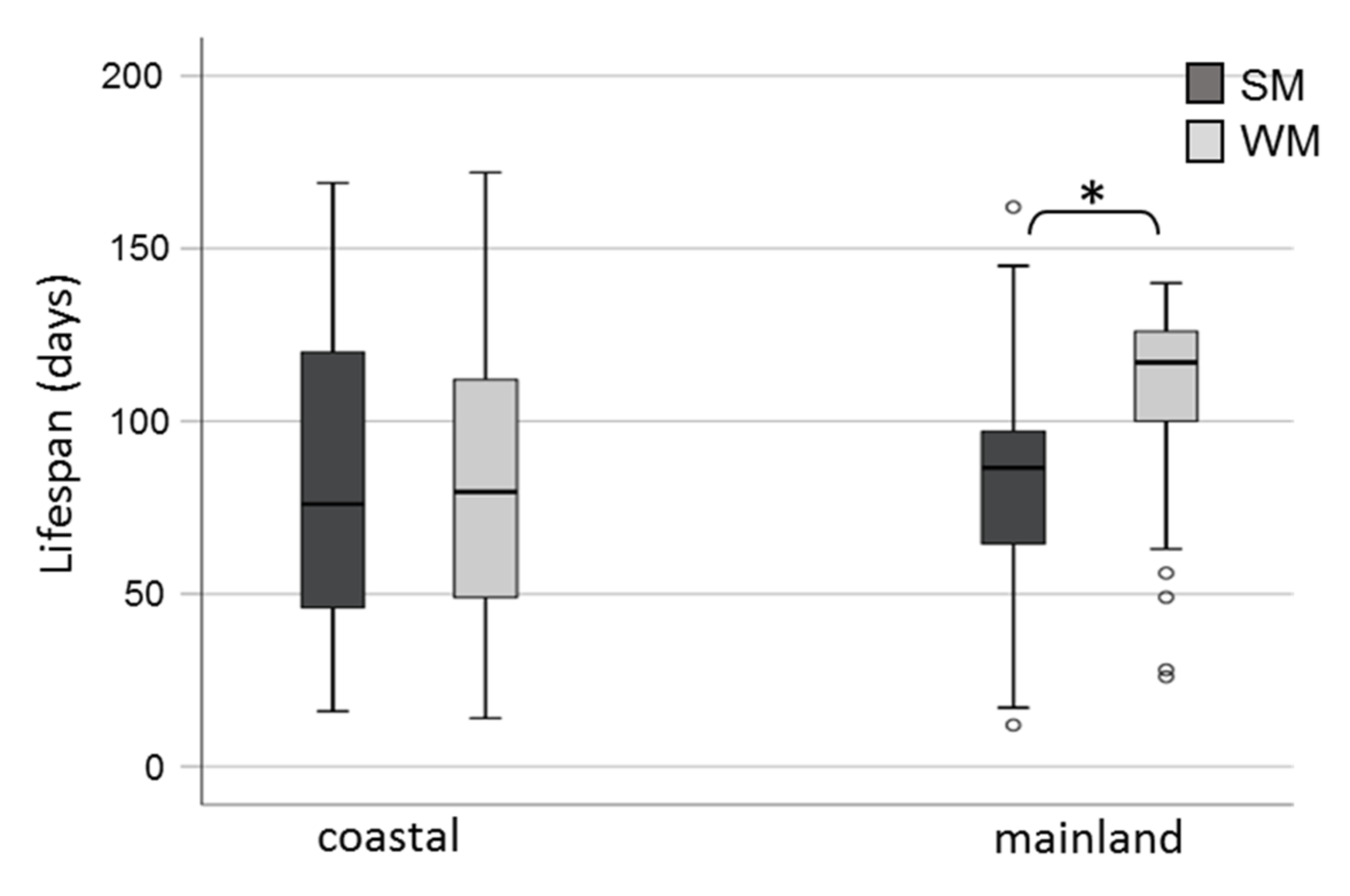
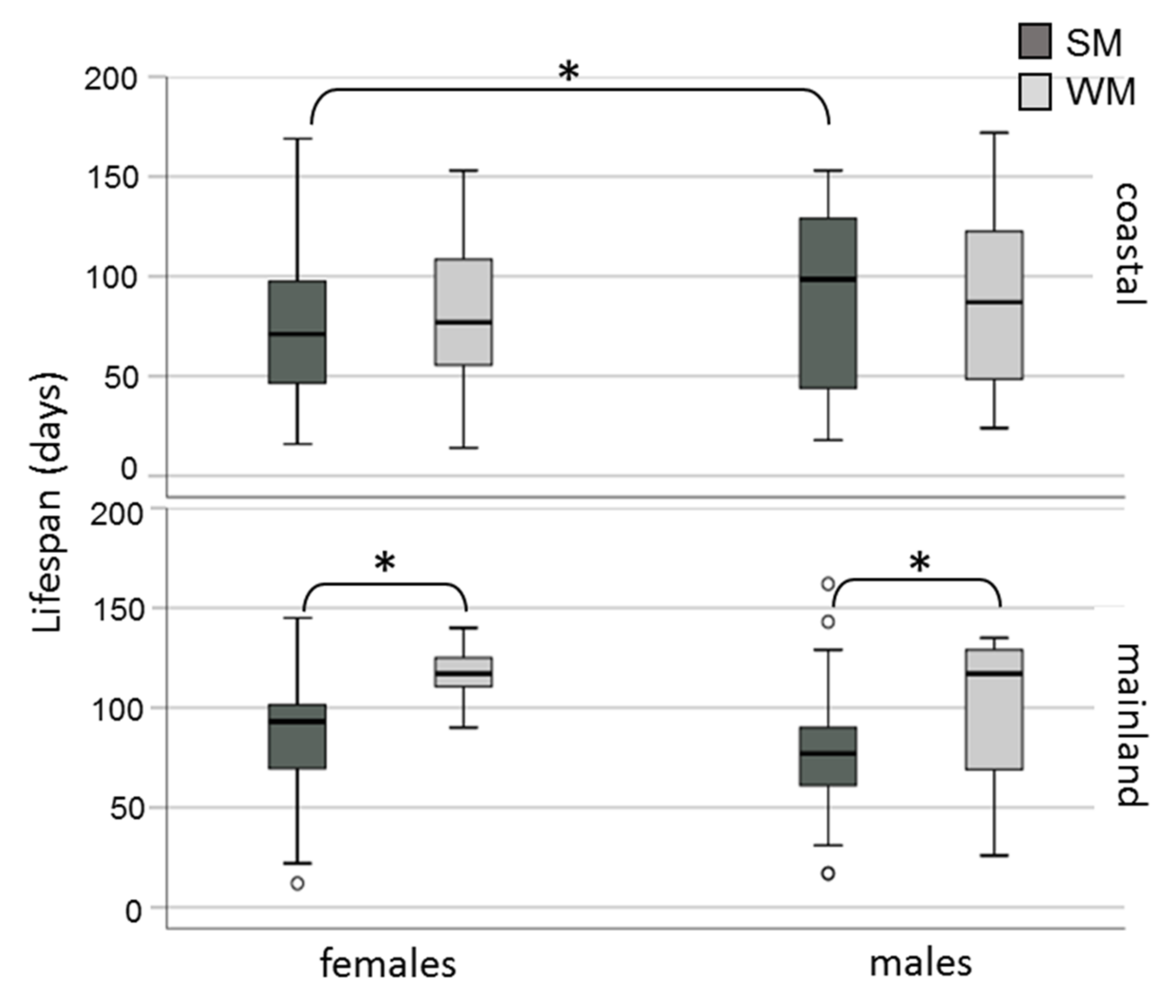
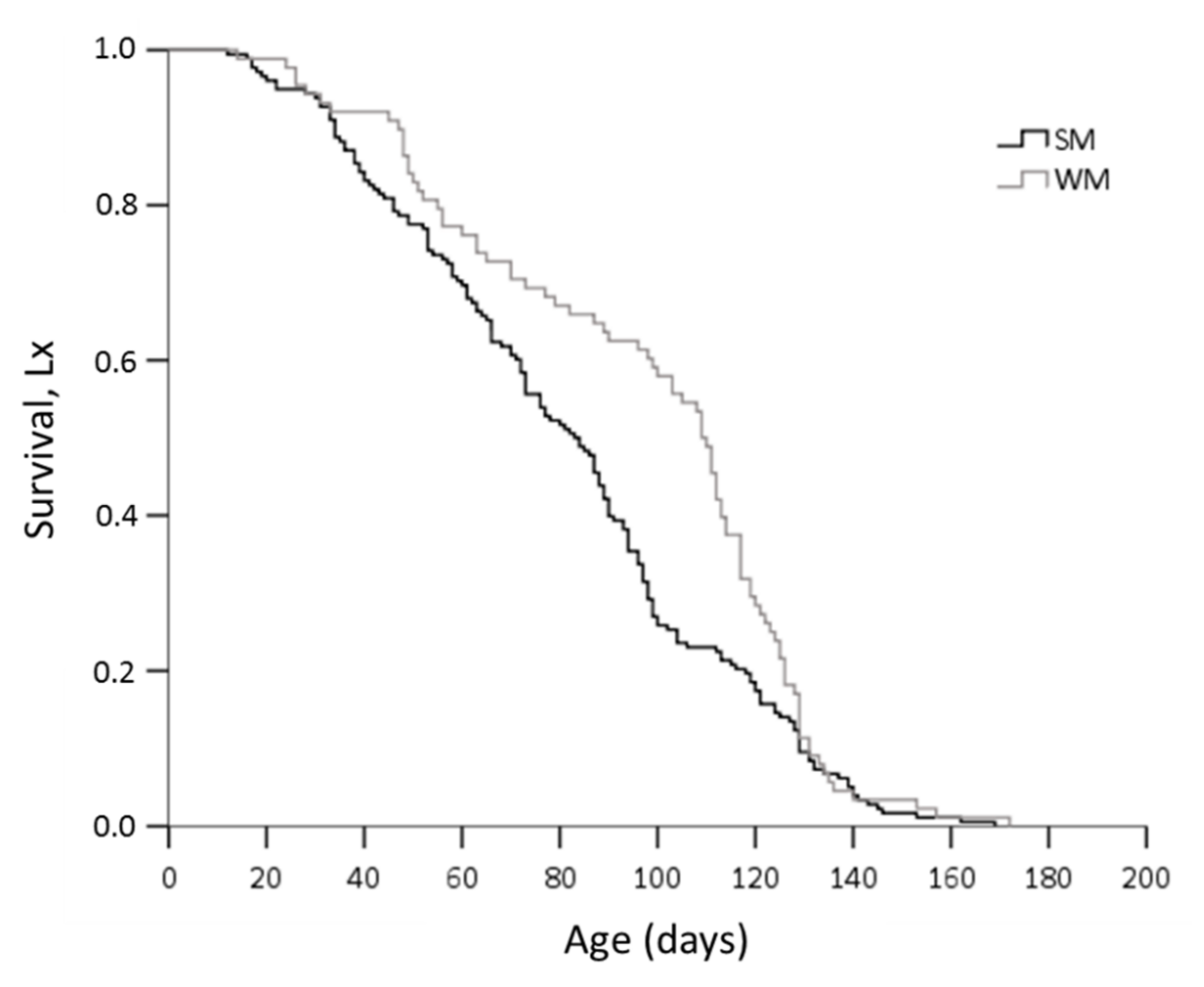
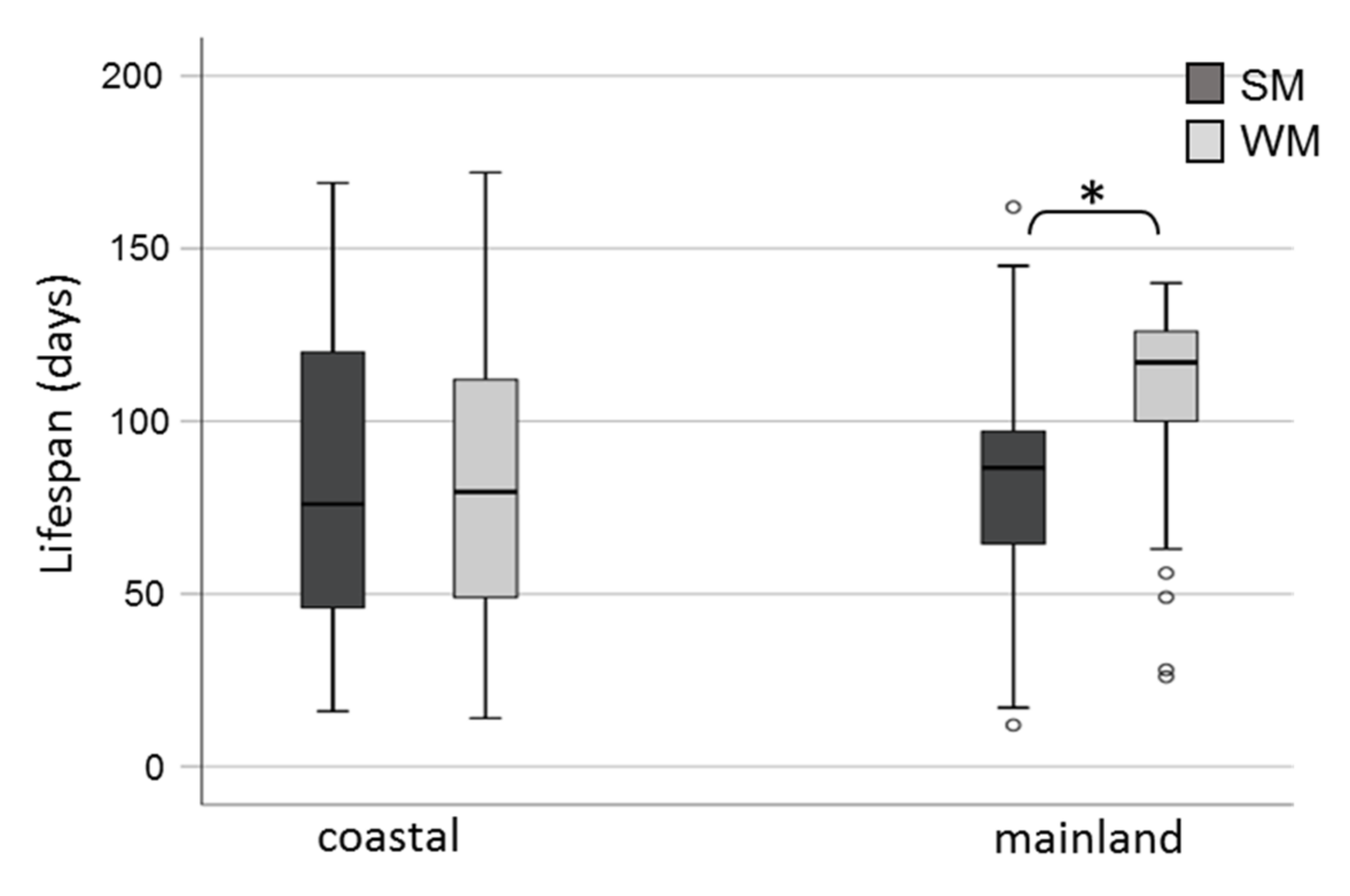
3.2. Survival Rates of Overwintering Adults
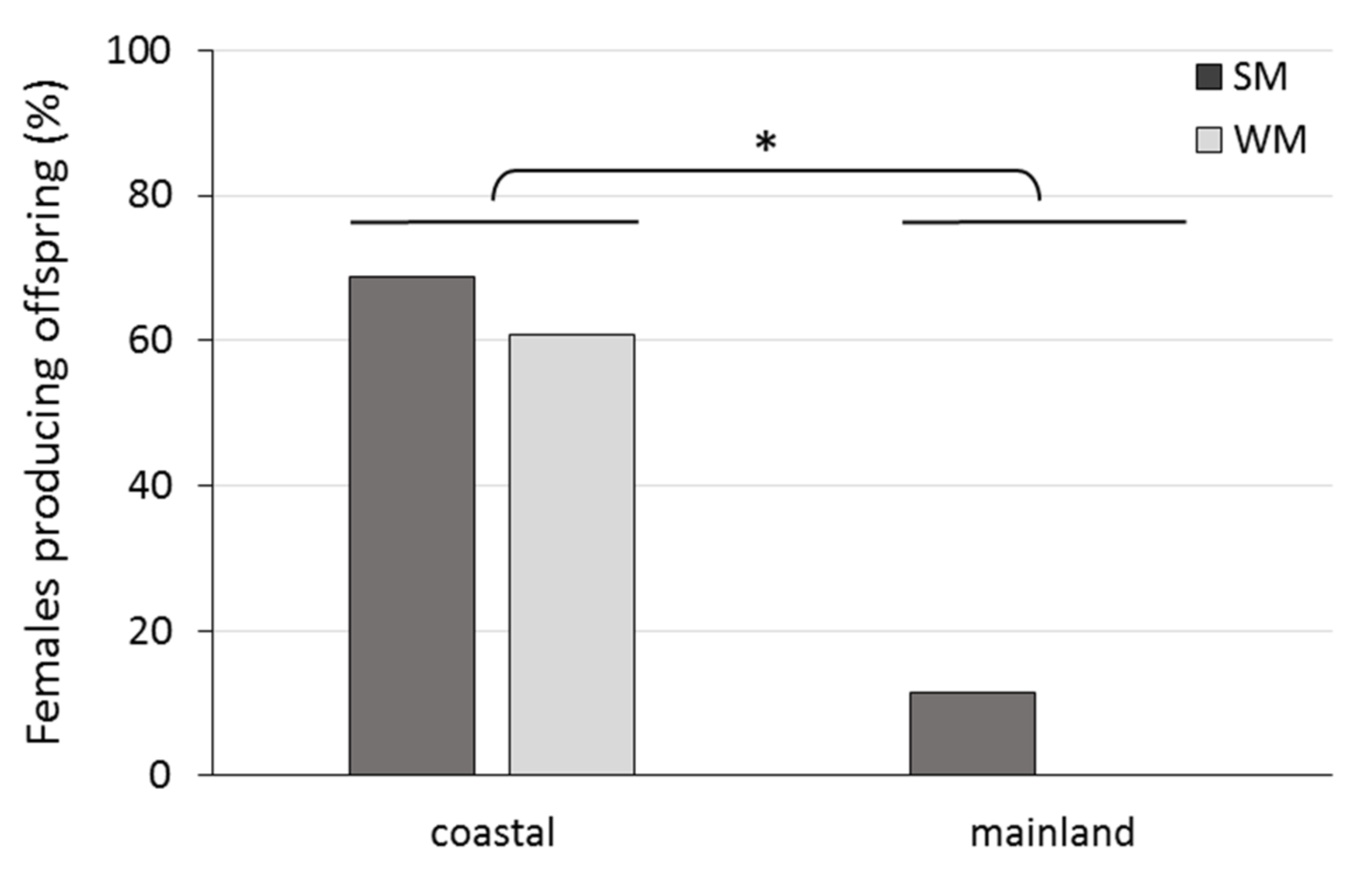
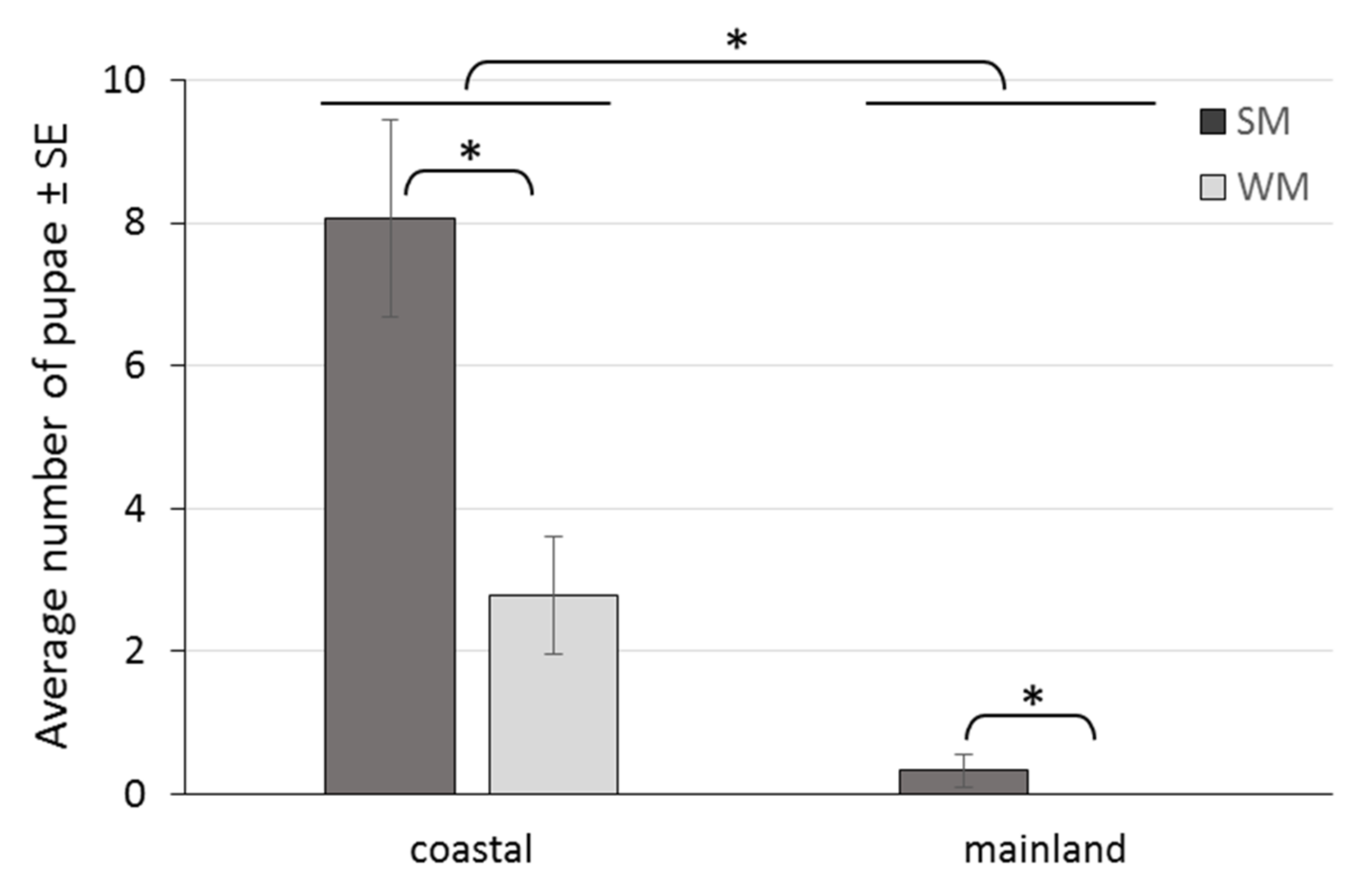
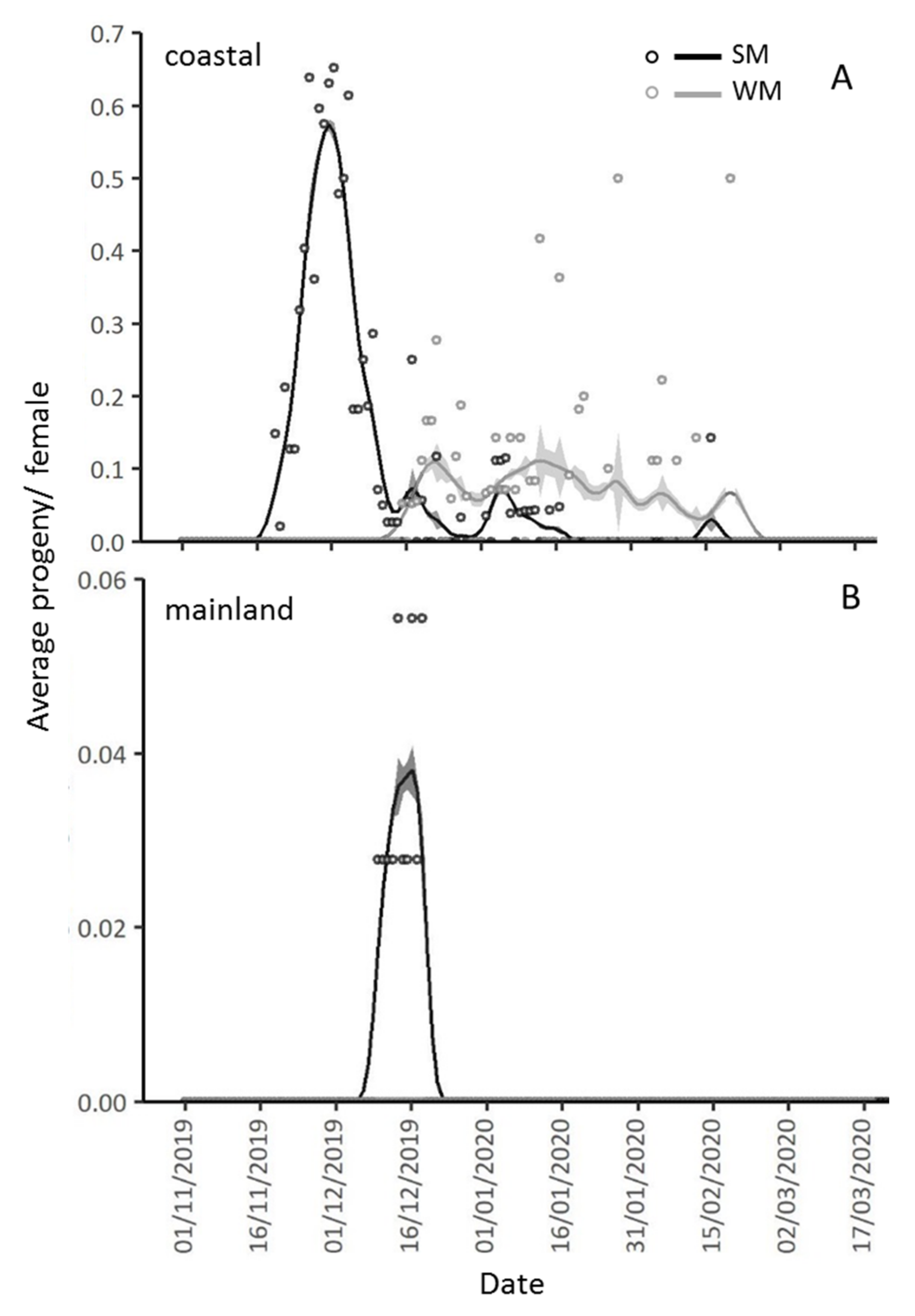
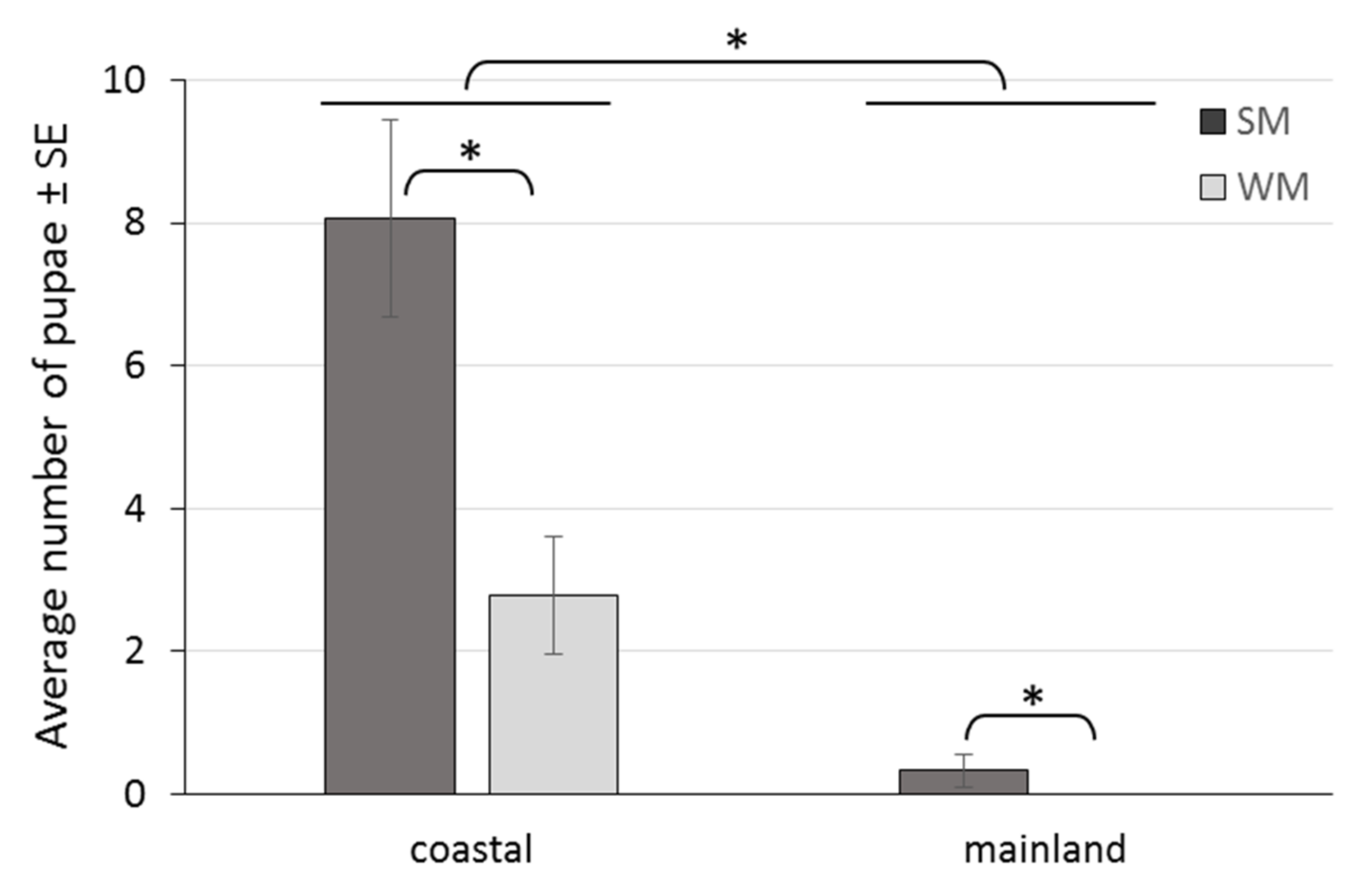
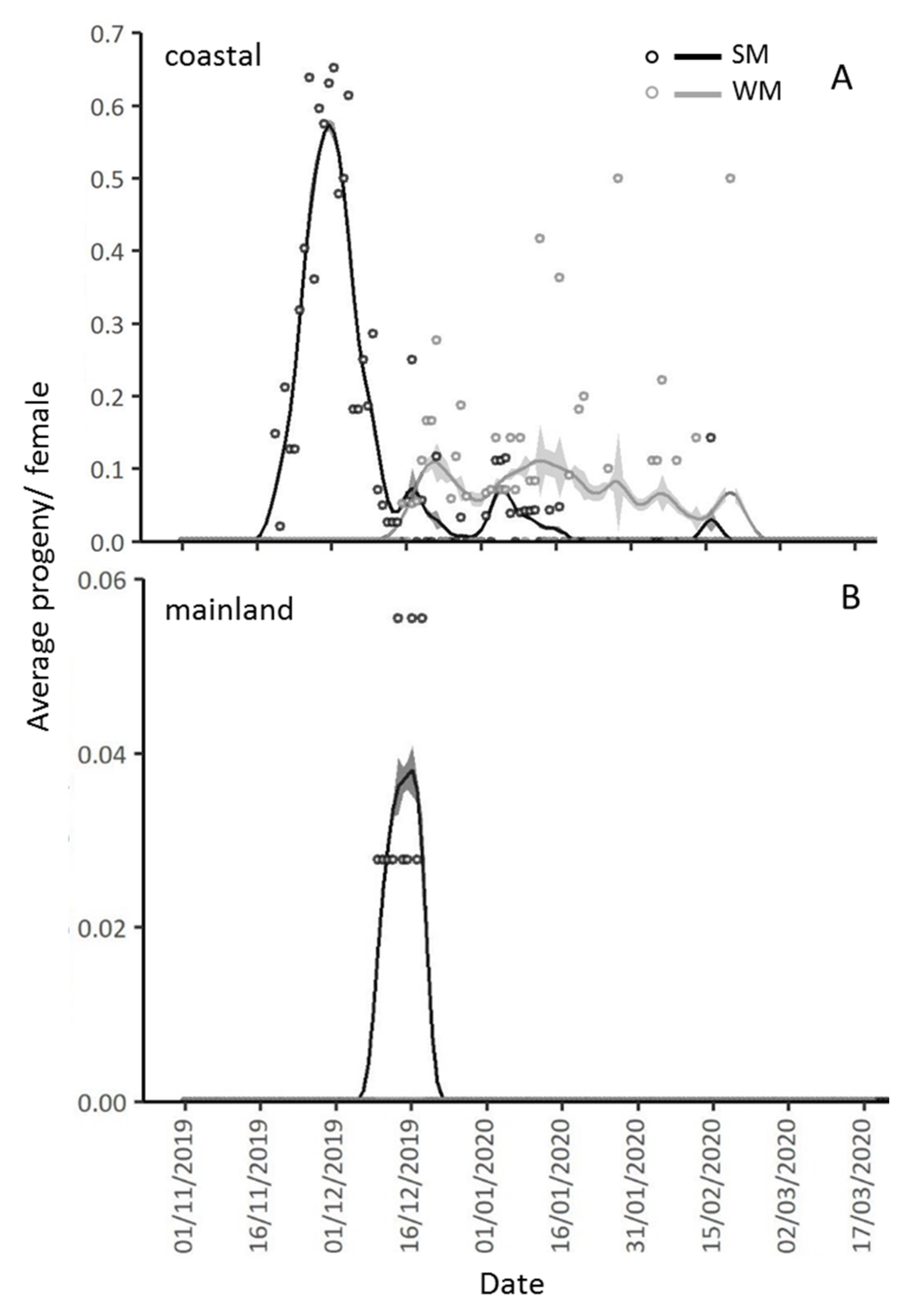
3.3. Reproductive Rates of Overwintering Females
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Burrack, H.J.; Fernandez, G.E.; Spivey, T.; Kraus, D.A. Variation in selection and utilization of host crops in the field and laboratory by Drosophila suzukii Matsumara (Diptera: Drosophilidae), an invasive frugivore. Pest Manag. Sci. 2013, 69, 1173–1180. [Google Scholar] [CrossRef]
- Cini, A.; Ioriatti, C.; Anfora, G. A review of the invasion of Drosophila suzukii in Europe and a draft research agenda for integrated pest management. Bull. Insectol. 2012, 65, 149–160. [Google Scholar]
- Lee, J.C.; Bruck, D.J.; Dreves, A.J.; Ioriatti, C.; Vogt, H.; Baufeld, P. In Focus: Spotted wing drosophila, Drosophila suzukii, across perspectives. Pest Manag. Sci. 2011, 67, 1349–1351. [Google Scholar] [CrossRef]
- Tochen, S.; Dalton, D.T.; Wiman, N.; Hamm, C.; Shearer, P.W.; Walton, V.M. Temperature-related development and population parameters for Drosophila suzukii (Diptera: Drosophilidae) on cherry and blueberry. Environ. Entomol. 2014, 43, 501–510. [Google Scholar] [CrossRef] [Green Version]
- Asplen, M.K.; Anfora, G.; Biondi, A.; Choi, D.S.; Chu, D.; Daane, K.M.; Gibert, P.; Gutierrez, A.P.; Hoelmer, K.A.; Hutchison, W.D.; et al. Invasion biology of spotted wing Drosophila (Drosophila suzukii): A global perspective and future priorities. J. Pest Sci. 2015, 88, 469–494. [Google Scholar] [CrossRef]
- Calabria, G.; Máca, J.; Bächli, G.; Serra, L.; Pascual, M. First records of the potential pest species Drosophila suzukii (Diptera: Drosophilidae) in Europe. J. Appl. Entomol. 2012, 136, 139–147. [Google Scholar] [CrossRef]
- Cini, A.; Anfora, G.; Escudero-Colomar, L.A.; Grassi, A.; Santosuosso, U.; Seljak, G.; Papini, A. Tracking the invasion of the alien fruit pest Drosophila suzukii in Europe. J. Pest Sci. 2014, 87, 559–566. [Google Scholar] [CrossRef]
- EPPO Global Database. Available online: https://gd.eppo.int/ (accessed on 1 May 2020).
- Papachristos, D.P.; Matakoulis, C.; Papadopoulos, N.T.; Lagkouranis, A.; Zarpas, K.D.; Milonas, P.G. First record of the harmful pest Drosophila suzukii (Diptera: Drosophilidae) in Greece. In Proceedings of the 15th Panhellenic Entomological Congress, Kavala, Greece, 22–25 October 2013; Andreadis, S.S., Pappas, M.L., Eds.; Hellenic Entomological Society: Kavala, Greece, 2013. [Google Scholar]
- Gibert, J.-M.; Peronnet, F.; Schlötterer, C. Phenotypic plasticity in Drosophila pigmentation caused by temperature sensitivity of a chromatin regulator network. PLoS Genet. 2007, 3, e30. [Google Scholar] [CrossRef] [PubMed]
- Kivelä, S.M.; Välimäki, P.; Gotthard, K. Seasonality maintains alternative life-history phenotypes. Evolution 2013, 67, 3145–3160. [Google Scholar] [CrossRef]
- Shearer, P.W.; West, J.D.; Walton, V.M.; Brown, P.H.; Svetec, N.; Chiu, J.C. Seasonal cues induce phenotypic plasticity of Drosophila suzukii to enhance winter survival. BMC Ecol. 2016, 16, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Enriquez, T.; Colinet, H. Basal tolerance to heat and cold exposure of the spotted wing drosophila, Drosophila suzukii. PeerJ 2017, 5, e3112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Panel, A.D.C.; Pen, I.; Pannebakker, B.A.; Helsen, H.H.M.; Wertheim, B. Seasonal morphotypes of Drosophila suzukii differ in key life-history traits during and after a prolonged period of cold exposure. Ecol. Evol. 2020, 10, 9085–9099. [Google Scholar] [CrossRef] [PubMed]
- Stephens, A.R.; Asplen, M.K.; Hutchison, W.D.; Venette, R.C. Cold Hardiness of Winter-Acclimated Drosophila suzukii (Diptera: Drosophilidae) Adults. Environ. Entomol. 2015, 44, 1619–1626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stockton, D.; Wallingford, A.; Loeb, G. Phenotypic Plasticity Promotes Overwintering Survival in A Globally Invasive Crop Pest, Drosophila suzukii. Insects 2018, 9, 105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wallingford, A.K.; Loeb, G.M. Developmental Acclimation of Drosophila suzukii (Diptera: Drosophilidae) and Its Effect on Diapause and Winter Stress Tolerance. Environ. Entomol. 2016, 45, 1081–1089. [Google Scholar] [CrossRef]
- Grassi, A.; Gottardello, A.; Dalton, D.T.; Tait, G.; Rendon, D.; Ioriatti, C.; Gibeaut, D.; Rossi Stacconi, M.V.; Walton, V.M. Seasonal reproductive biology of Drosophila suzukii (Diptera: Drosophilidae) in temperate climates. Environ. Entomol. 2018, 47, 166–174. [Google Scholar] [CrossRef]
- Zerulla, F.N.; Schmidt, S.; Streitberger, M.; Zebitz, C.P.W.; Zelger, R. On the overwintering ability of Drosophila suzukii in South Tyrol. J. Berry Res. 2015, 5, 41–48. [Google Scholar] [CrossRef] [Green Version]
- Panel, A.D.C.; Zeeman, L.; Van der Sluis, B.J.; Van Elk, P.; Pannebakker, B.A.; Wertheim, B.; Helsen, H.H. Overwintered Drosophila suzukii are the main source for infestations of the first fruit crops of the season. Insects 2018, 9, 145. [Google Scholar] [CrossRef] [Green Version]
- Papanastasiou, S.A.; Rodovitis, V.G.; Bataka, E.P.; Verykouki, E.; Papadopoulos, N.T. Population Dynamics of Drosophila suzukii in Coastal and Mainland Sweet Cherry Orchards of Greece. Insects 2020, 11, 621. [Google Scholar] [CrossRef]
- Jaramillo, S.L.; Mehlferber, E.; Moore, P.J. Life-history trade-offs under different larval diets in Drosophila suzukii (Diptera: Drosophilidae). Physiol. Entomol. 2015, 40, 2–9. [Google Scholar] [CrossRef]
- Emiljanowicz, L.M.; Ryan, G.D.; Langille, A.; Newman, J. Development, reproductive output and population growth of the fruit fly pest Drosophila suzukii (Diptera: Drosophilidae) on artificial diet. J. Econ. Entomol. 2014, 107, 1392–1398. [Google Scholar] [CrossRef] [Green Version]
- Kinjo, H.; Kunimi, Y.; Nakai, M. Effects of temperature on the reproduction and development of Drosophila suzukii (Diptera: Drosophilidae). Appl. Entomol. Zool. 2014, 49, 297–304. [Google Scholar] [CrossRef]
- Hamby, K.A.; Bellamy, D.E.; Chiu, J.C.; Lee, J.C.; Walton, V.M.; Wiman, N.G.; York, R.M.; Biondi, A. Biotic and abiotic factors impacting development, behavior, phenology, and reproductive biology of Drosophila suzukii. J. Pest Sci. 2016, 89, 605–619. [Google Scholar] [CrossRef]
- Tochen, S.; Woltz, J.M.; Dalton, D.T.; Lee, J.C.; Wiman, N.G.; Walton, V.M. Humidity affects populations of Drosophila suzukii (Diptera: Drosophilidae) in blueberry. J. Appl. Entomol. 2016, 140, 47–57. [Google Scholar] [CrossRef]
- Nylin, S.; Gotthard, K. Plasticity in Life-History Traits. Annu. Rev. Entomol. 1998, 43, 63–83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dalton, D.T.; Walton, V.M.; Shearer, P.W.; Walsh, D.B.; Caprile, J.; Isaacs, R. Laboratory survival of Drosophila suzukii under simulated winter conditions of the Pacific Northwest and seasonal field trapping in five primary regions of small and stone fruit production in the United States. Pest Manag. Sci. 2011, 67, 1368–1374. [Google Scholar] [CrossRef] [PubMed]
- Kimura, M.T. Cold and heat tolerance of drosophilid flies with reference to their latitudinal distributions. Oecologia 2004, 140, 442–449. [Google Scholar] [CrossRef] [PubMed]
- Ryan, G.D.; Emiljanowicz, L.; Wilkinson, F.; Kornya, M.; Newman, J.A. Thermal tolerances of the spotted-wing Drosophila Drosophila suzukii (Diptera: Drosophilidae). J. Econ. Entomol. 2016, 109, 746–752. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wallingford, A.K.; Lee, J.C.; Loeb, G.M. The influence of temperature and photoperiod on the reproductive diapause and cold tolerance of spotted-wing drosophila, Drosophila suzukii. Entomol. Exp. Appl. 2016, 159, 327–337. [Google Scholar] [CrossRef] [Green Version]
- Mitsui, H.; Beppu, K.; Kimura, M.T. Seasonal life cycles and resource uses of flower-and fruit-feeding drosophilid flies (Diptera: Drosophilidae) in central Japan. Entomol. Sci. 2010, 13, 60–67. [Google Scholar] [CrossRef]
- Toxopeus, J.; Jakobs, R.; Ferguson, L.V.; Gariepy, T.D.; Sinclair, B.J. Reproductive arrest and stress resistance in winter-acclimated Drosophila suzukii. J. Insect Physiol. 2016, 89, 37–51. [Google Scholar] [CrossRef] [Green Version]
- Rendon, D.; Buser, J.; Tait, G.; Lee, J.C.; Walton, V.M. Survival and fecundity parameters of two Drosophila suzukii (Diptera: Drosophilidae) morphs on variable diet under suboptimal temperatures. J. Insect Sci. 2018, 18, 8. [Google Scholar] [CrossRef] [Green Version]
- Kacar, G.; Wang, X.G.; Stewart, T.J.; Daane, K.M. Overwintering survival of Drosophila suzukii (Diptera: Drosophilidae) and the effect of food on adult survival in California’s San Joaquin Valley. Environ. Entomol. 2016, 45, 763–771. [Google Scholar] [CrossRef] [PubMed]
- Prais, S.J.; Winsten, C.B. Trend Estimators and Serial Correlation; Cowles Commission: Chicago, IL, USA, 1954; pp. 1–26. [Google Scholar]
- Halekoh, U.; Højsgaard, S.; Yan, J. The R package geepack for generalized estimating equations. J. Stat. Softw. 2006, 15, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Yan, J. Geepack: Yet another package for generalized estimating equations. R-News 2002, 2, 12–14. [Google Scholar]
- Yan, J.; Fine, J. Estimating equations for association structures. Stat. Med. 2004, 23, 859–874. [Google Scholar] [CrossRef]
- Mohr, F.X. Prais-Winsten Estimator for AR(1) Serial Correlation. R Package Version 1.1.1. 2019. Available online: https://CRAN.R-project.org/package=prais (accessed on 5 July 2021).
- Rendon, D.; Walton, V.; Tait, G.; Buser, J.; Lemos Souza, I.; Wallingford, A.; Loeb, G.; Lee, J. Interactions among morphotype, nutrition, and temperature impact fitness of an invasive fly. Ecol. Evol. 2019, 9, 2615–2628. [Google Scholar] [CrossRef] [Green Version]
- Kimura, M.; Awasaki, T.; Ohtsu, T.; Shimada, K. Seasonal changes in glycogen and trehalose content in relation to winter survival of four temperate species of Drosophila. J. Insect Physiol. 1992, 38, 871–875. [Google Scholar] [CrossRef]
- Ohtsu, T.; Kimura, M.; Hori, S. The influence of eclosion timing on winter survival and triacylglycerol accumulation in four temperate species of Drosophila. Physiol. Entomol. 1995, 20, 248–252. [Google Scholar] [CrossRef]
- Jakobs, R.; Ahmadi, B.; Houben, S.; Gariepy, T.D.; Sinclair, B.J. Cold tolerance of third-instar Drosophila suzukii larvae. J. Insect Physiol. 2017, 96, 45–52. [Google Scholar] [CrossRef] [Green Version]
- Teets, N.M.; Denlinger, D.L. Physiological mechanisms of seasonal and rapid cold-hardening in insects. Physiol. Entomol. 2013, 38, 105–116. [Google Scholar] [CrossRef]
- Stockton, D.; Wallingford, A.; Rendon, D.; Fanning, P.; Green, C.K.; Diepenbrock, L.; Ballman, E.; Walton, V.M.; Isaacs, R.; Leach, H.; et al. Interactions between Biotic and Abiotic Factors Affect Survival in Overwintering Drosophila suzukii (Diptera: Drosophilidae). Environ. Entomol. 2019, 48, 454–464. [Google Scholar] [CrossRef]
- Colinet, H.; Hoffmann, A.A. Comparing phenotypic effects and molecular correlates of developmental, gradual and rapid cold acclimation responses in Drosophila melanogaster. Funct. Ecol. 2012, 26, 84–93. [Google Scholar] [CrossRef]
- Chakir, M.; Chafik, A.; Moreteau, B.; Gibert, P.; David, J.R. Male sterility thermal thresholds in Drosophila: D. simulans appears more cold-adapted than its sibling D. melanogaster. Genetica 2002, 114, 195–205. [Google Scholar] [CrossRef] [PubMed]
- Boulétreau-merle, J.; Fouillet, P. How to overwinter and be a founder: Egg-retention phenotypes and mating status in Drosophila melanogaster. Evol. Ecol. 2002, 16, 309–332. [Google Scholar] [CrossRef]
- Rossi-Stacconi, M.V.; Kaur, R.; Mazzoni, V.; Ometto, L.; Grassi, A.; Gottardello, A.; Rota-Stabelli, O.; Anfora, G. Multiple lines of evidence for reproductive winter diapause in the invasive pest Drosophila suzukii: Useful clues for control strategies. J. Pest Sci. 2016, 89, 689–700. [Google Scholar] [CrossRef]
- Prowse, N.; Partridge, L. The effects of reproduction on longevity and fertility in male Drosophila melanogaster. J. Insect Physiol. 1997, 43, 501–512. [Google Scholar] [CrossRef]
- Papanastasiou, S.A.; Nakas, C.T.; Carey, J.R.; Papadopoulos, N.T. Condition-Dependent Effects of Mating on Longevity and Fecundity of Female Medflies: The Interplay between Nutrition and Age of Mating. PLoS ONE 2013, 8, e70181. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Location | Lat. (North) | Long. (East) | Treatment | Number |
|---|---|---|---|---|
| Coastal: Volos | 39°23′17.6′′ | 22°56′24.6′′ | SM females | 50 |
| WM females | 25 | |||
| SM males | 50 | |||
| WM males | 25 | |||
| Mainland: Naousa | 40°37′45.5′′ | 22°04′11.0′′ | SM females | 50 |
| WM females | 25 | |||
| SM males | 50 | |||
| WM males | 25 |
| Variable (Ref: Coastal Area) | B (SE) | p Value |
|---|---|---|
| Maximum Temperature | −3.76 (1.51) | 0.013 |
| Mean Temperature | −3.41 (1.61) | 0.035 |
| Minimum Temperature | −2.76 (1.52) | 0.069 |
| Maximum Relative Humidity | −1.30 (2.42) | 0.591 |
| Minimum Relative Humidity | 3.91 (3.03) | 0.198 |
| Factor | Number of Progeny | |
|---|---|---|
| Wald t-Test (df) | p Value | |
| Mean Temperature | 3.03 (1) | 0.082 |
| Area (ref: mainland) | 6.99 (1) | 0.008 |
| Phenotype (ref: WM) | 10.76 (1) | 0.001 |
| Days in the field | 4.78 (1) | 0.029 |
| Mean Temperature × Area | 4.09 (1) | 0.043 |
| Area × Phenotype | 2383.76 (1) | <0.001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Papanastasiou, S.A.; Rodovitis, V.G.; Verykouki, E.; Bataka, E.P.; Papadopoulos, N.T. Adaptation of an Invasive Pest to Novel Environments: Life History Traits of Drosophila suzukii in Coastal and Mainland Areas of Greece during Overwintering. Biology 2021, 10, 727. https://doi.org/10.3390/biology10080727
Papanastasiou SA, Rodovitis VG, Verykouki E, Bataka EP, Papadopoulos NT. Adaptation of an Invasive Pest to Novel Environments: Life History Traits of Drosophila suzukii in Coastal and Mainland Areas of Greece during Overwintering. Biology. 2021; 10(8):727. https://doi.org/10.3390/biology10080727
Chicago/Turabian StylePapanastasiou, Stella A., Vasilis G. Rodovitis, Eleni Verykouki, Evmorfia P. Bataka, and Nikos T. Papadopoulos. 2021. "Adaptation of an Invasive Pest to Novel Environments: Life History Traits of Drosophila suzukii in Coastal and Mainland Areas of Greece during Overwintering" Biology 10, no. 8: 727. https://doi.org/10.3390/biology10080727
APA StylePapanastasiou, S. A., Rodovitis, V. G., Verykouki, E., Bataka, E. P., & Papadopoulos, N. T. (2021). Adaptation of an Invasive Pest to Novel Environments: Life History Traits of Drosophila suzukii in Coastal and Mainland Areas of Greece during Overwintering. Biology, 10(8), 727. https://doi.org/10.3390/biology10080727