Abstract
Wild boars, widely distributed across natural, agricultural, and urban landscapes, represent an ideal sentinel species for monitoring the emergence and spread of antimicrobial resistance (AMR) at the human–wildlife–livestock interface within the One Health framework. This review summarizes current knowledge on the prevalence, diversity, AMR, and epidemiological significance of major enteric pathogens isolated from wild boars in the European Union, with particular attention to their potential role in AMR dissemination. Numerous studies have reported variable prevalence rates for Salmonella spp., Yersinia enterocolitica, Shiga toxin-producing Escherichia coli (STEC), and Campylobacter spp. High prevalence rates has been observed in fecal samples—35% for Salmonella, 27% for Y. enterocolitica and STEC, and 66% for Campylobacter—highlighting the role of wild boars as carriers and the associated risk of carcass contamination during slaughter. Tonsils represent a key niche for Y. enterocolitica, with prevalence reaching 35%. Several studies have identified resistance to antimicrobials classified by the World Health Organization as critically important or high priority for human medicine, including fluoroquinolone-resistant non-typhoidal Salmonella spp. and third-generation cephalosporin-resistant Y. enterocolitica, raising notable public health concerns. Despite increasing interest, most available studies remain descriptive and geographically limited, providing limited insight into AMR acquisition and transmission pathways in wild boars. New approaches—such as resistome analyses and epidemiological cut-off values—offer added value to distinguish wild-type from acquired-resistant strains and to better understand AMR dissemination dynamics. Integrating wildlife into One Health surveillance systems is essential to capture the full complexity of AMR spread.
1. Introduction
Wild boar (Sus scrofa) is one of the most widely distributed mammals worldwide, with populations increasing steadily over the past 30 years [1]. This growth is driven by its r-strategist reproductive traits [2,3] and marked ecological adaptability, as an opportunistic omnivore capable of adjusting its diet and thriving in diverse environments—from natural and agricultural areas to peri-urban and urban settings—while exploiting anthropogenic resources [4,5].
Other factors contributing to the increase in wild boar numbers include the scarcity of natural predators: humans represent the main cause of mortality, primarily through hunting and, to a lesser extent, traffic collisions [4]. Additionally, habitat fragmentation can influence wild boar movement and distribution, leading to increased mobility in search of resources, altered foraging behavior, and potentially greater human–wildlife conflicts [6,7].
As a consequence of population growth and geographic expansion, new challenges have emerged. The broad distribution of wild boars and their presence across diverse habitats highlight their major epidemiological role as hosts and reservoirs for several zoonotic agents such as Mycobacterium bovis [8], Brucella spp. [9], and pathogens capable of spill-over to domestic animals, such as African Swine fever virus [10]. There is a growing body of scientific literature on the role of wild boars as carriers of pathogens and indicator bacteria that may also harbor antimicrobial resistance determinants, as Salmonella spp., Shiga-toxins producing E. coli (STEC), Yersinia enterocolitica, Campylobacter spp., methicillin-resistant Staphylococcus aureus (MRSA), vancomycin-resistant enterococci, etc. [11,12,13,14,15,16].
Moreover, the synurbization—a specific form of synanthropization defined as the adaptation of animal populations to human-modified environments—of wild boars increases the likelihood of pathogen spillover events. Their frequent interactions with domestic animals, pets, and humans in peri-urban and urban environments facilitate the transmission of enteric pathogens and antimicrobial-resistant bacteria across ecological boundaries [13].
Antimicrobial resistance (AMR) is a major global challenge for both human and animal health, accelerated by human activities [17,18]. Many antimicrobials used in veterinary medicine and agriculture belong to the same classes as those used in human medicine [19], including substances deemed “critically important” by the World Health Organization [20]. Managing AMR therefore requires a coordinated One Health approach [21], which acknowledges the interconnectedness of human, animal, plant, and environmental health [22].
Recently, the One Health model has been further refined to include the pivotal role of the geographical proximity of ecosystems in the emergence and dissemination of health-related traits [23]. Within this framework, AMR represents the quintessential One Health issue [24]. Bacteria and their genes have a remarkable capacity to adapt and mutate rapidly, and to move within and between humans, animals, and the environment through horizontal gene transfer [25]. AMR (particularly in Enterobacterales at the fecal level) has been already widely observed in wild reptiles, birds, and mammals [26,27,28,29]. However, knowledge of the transmission dynamics within environmental and wildlife compartments remains limited, and the directionality of these mechanisms is complex to determine and mostly assumed [5]. In this context, wild boars represent the ideal model species for understanding the emergence, spread, and persistence of AMR at the human–wildlife–livestock interface.
The aim of this comprehensive review was to analyze current knowledge on the prevalence, diversity (in terms of identified species, serotypes, and biotypes), AMR, and epidemiological significance of major enteric pathogens (Salmonella, Y. enterocolitica, STEC, and Campylobacter) isolated from wild boars in the European Union. Novel approaches, such as resistome analysis and the use of epidemiological cut-off values, were also considered, as they help address existing gaps in the understanding of AMR acquisition mechanisms and transmission routes.
In addition, the review examined the role of the wild boar as a model of synurbization, highlighting how the increasing expansion of this species into peri-urban and urban environments enhances opportunities for interactions with humans, domestic animals, and anthropogenic sources of contamination. This aspect is particularly relevant when considering sentinel bacteria for AMR surveillance, such as Enterococcus spp., which can reflect environmental and human-associated antimicrobial pressure.
2. Materials and Methods
This comprehensive review focused on the four leading zoonotic agents in the European Union, as identified in the most recent EFSA and ECDC report [30]: Campylobacter spp., Salmonella spp., Shiga toxin-producing E. coli (STEC), and Y. enterocolitica.
A structured literature search was conducted using the SCOPUS database. The search strategy included combinations of keywords such as “wild boar”, “Sus scrofa”, “antimicrobial resistance”, “AMR”, “antibiotic resistance”, “Salmonella”, “Yersinia enterocolitica”, “STEC”, “Shiga toxin–producing Escherichia coli”, “Campylobacter”, “Enterobacterales”, “foodborne pathogens”, and “Europe”. Moreover, studies regarding Enterococcus spp. were also included as considered important sentinel bacteria for antibiotic resistance surveillance.
The review includes studies published over the past 15 years to provide a comprehensive overview of the prevalence and antimicrobial resistance of enteric pathogens isolated from wild boars. Although this extended time span allows the inclusion of a larger body of literature, it is acknowledged that diagnostic methods, antimicrobial susceptibility testing standards, and breakpoint interpretations have changed over the years, complicating the interpretation of temporal trends. The primary focus of the review remains on more recent studies (last 10 years), whereas older studies are included to provide historical context and to illustrate long-term patterns.
Titles and abstracts were screened to identify studies reporting occurrence, phenotypic and/or genotypic antimicrobial resistance in the target microorganisms isolated from wild boars.
Exclusion criteria included studies conducted outside the European Union.
3. Silent Carriers at the Human–Wildlife Interface: Enteric Pathogens in European Wild Boars
The scientific literature provides substantial evidence of the role of wild boars as reservoir of enteric pathogenic microorganisms, with particular emphasis on the four leading zoonotic agents in the European Union, as identified in the most recent EFSA and ECDC report [30]: Campylobacter spp., Salmonella spp., STEC and Y. enterocolitica.
Investigations into the carrier status of wild boar have been conducted primarily on fecal samples, although other matrices such as lymph nodes and tonsils are also commonly examined. In addition, to evaluate the risk of meat contamination during slaughter and subsequent processing, analyses frequently include carcass, meat and organs surface, such as the liver.
In the following sections, findings on prevalence, influencing factors, and diversity of Salmonella spp., Y. enterocolitica, Campylobacter spp., and STEC from studies conducted in various European Union countries are presented.
3.1. Prevalence and Diversity of Salmonella spp.
Salmonella spp. is a major cause of food-borne illness in the world, with an incidence of 18.0 cases per 100,000 inhabitants in the European Union [30] and an estimate of 1.35 million infections in the United States every year [31].
Domesticated animals are considered the main reservoir of non-typhoidal Salmonella, spp. but wild species can also act as asymptomatic carriers, and their role has gained increasing attention in recent years [32].
Carriage of Salmonella spp. in wild boars has been investigated in several studies across different European Union countries.
Table 1 summarizes the prevalence of Salmonella spp. detected in different sample types (feces, lymph nodes, carcass surface, skin, tonsils, organs, blood) collected from wild boars in various European Union countries. The sampling period is indicated. Prevalence is expressed as the number of positive samples out of the total number examined, with the corresponding percentage reported in brackets. Details regarding the detection method (culture, molecular, and/or immunoenzymatic assays) are also provided.
Reported prevalence in fecal samples shows wide variability, ranging from 0% to 35.6%. In lymph nodes, prevalence rates vary between 0% and 17.8% [11,13,33,34,35,36,37,38,39,40,41,42,43,44,45,46,47,48,49,50]. Comparisons between studies are not always straightforward, as testing procedures vary. Differences in analytical methods (culture vs. culture combined with PCR) and sample types (intestinal contents vs. rectal swabs) can markedly influence estimates of Salmonella spp. prevalence, with rectal swabs generally less sensitive [13,33,40,43]. Fecal shedding is also intermittent in carrier wild boars, particularly after the acute phase, reducing detection probability [51,52]. Additionally, the amount of feces analyzed (1–25 g) significantly affects diagnostic sensitivity, as shown in both wild boar studies and pig research [33,36,42,53,54,55].
The fecal–oral route is considered the most frequent pathway for Salmonella spp. infection, which is why mesenteric lymph nodes are commonly examined. However, inhalation of contaminated aerosols or dust and infection via the respiratory tract are alternative routes [56,57]. Consequently, tonsils and other oral cavity tissues, which drain to the sub-mandibular lymph nodes, should also be included in studies. Sannö et al. [40] reported a Salmonella spp. prevalence of 12% in wild boar sub-mandibular lymph nodes, slightly higher than the 10% found in mesenteric lymph nodes but lower than the 14.7% observed in tonsils. Likewise, Gil Molino et al. [43] found Salmonella spp. prevalence to be approximately three times higher in tonsils (18.7%) than in mandibular lymph nodes (5.1%), underlining the importance of including these samples when assessing Salmonella spp. circulation in wild boar.
Several interconnected factors influence Salmonella spp. prevalence in wild boar populations, including habitat, climate, population structure, and sampling season [42,58]. Age is particularly important: younger animals (<24 months) show higher prevalence due to immature immunity, unstable gut microbiota, and greater exposure [36,38,46]. Age also affects detection by sampling site, as piglets tend to carry Salmonella spp. in lymph nodes, whereas older pigs shed it in feces, enhancing transmission [58]. Gender and seasonality further contribute, with higher prevalence in socially grouped females [33] and during warmer months, when environmental conditions promote bacterial survival and fecal–oral spread [41,42].
In the studies examined, a high variability of Salmonella spp. serovars and/or subspecies has frequently been reported. For instance, Chiari et al. [36] identified 30 different serotypes classified in three subspecies—enterica, diarizonae and houtenae—and Zottola et al. [38] identified 15 different serovars of S. enterica subsp. enterica, along with more than 20 isolates belonging to other subspecies such as salamae, diarizonae, and houtenae. Similarly, eight different serovars were detected in a wild boar population inhabiting the Asinara Natural Park, a small island off the northern coast of Sardinia with a surface area of only 51.9 km2, further highlighting the remarkable diversity of serovars [46].
This pattern is commonly observed in wildlife in general, and in wild boars in particular, as a result of their multiple sources of exposure, including livestock farming and waste disposal [32,36]. In addition, the omnivorous feeding habits of wild boars contribute to this variability, since they may consume other mammals, birds, reptiles, and amphibians that act as Salmonella spp. carriers [43,59].
A point of concern is that some of the most frequently detected serovars in wild boars—such as Typhimurium, Coeln, Enteritidis, Thompson, and Newport [38,42,46]—are among the most commonly reported in human salmonellosis cases in Europe in 2023 [30]. Another notable finding is the detection of serovars shared with livestock in areas characterized by extensive cattle farming, suggesting a possible spillover between the two species [60]. Conversely, other authors [42], using Pulsed-Field Gel Electrophoresis (PFGE), compared isolates from wild boars with those recovered from domestic pigs in an area characterized by intensive farming, finding an overlap only at the serotype level and none at the PFGE level, thus indicating the effectiveness of the biosecurity measures implemented in this type of farming system.
Taken together, these findings highlight the relevance of wild boars as a key species for understanding Salmonella circulation at the human–animal–environment interface, underscoring the importance of adopting a One Health perspective in surveillance and risk assessment.
Figure 1 and Figure 2 show the number of Salmonella enterica subspecies and the number serotypes of Salmonella enterica subs. enterica identified, respectively.
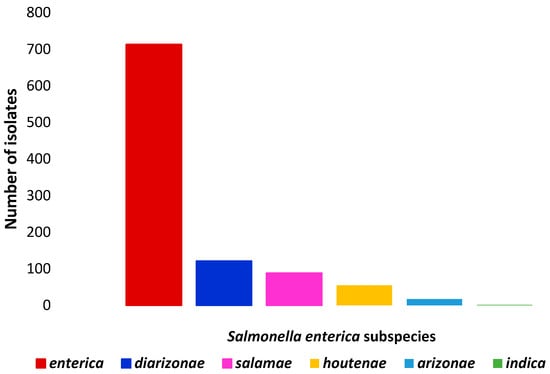
Figure 1.
Number of isolates of each Salmonella enterica subspecies identified in wild boars.
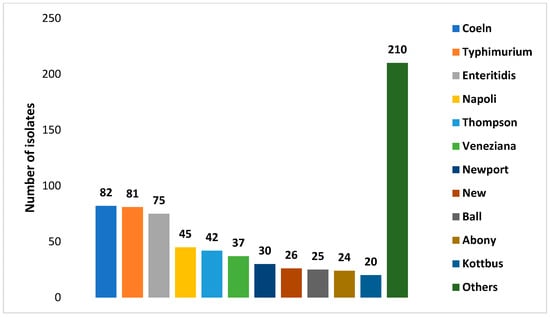
Figure 2.
Numbers of isolates of each Salmonella enterica subs. enterica serotypes identified in wild boars’ isolates.

Table 1.
Salmonella spp. prevalence expressed as positive/total (%) investigated in different sample types collected from wild boars in various countries of the European Union.
Table 1.
Salmonella spp. prevalence expressed as positive/total (%) investigated in different sample types collected from wild boars in various countries of the European Union.
| Country | Sampling Period | Source | Positive/Total (%) | Comments on Detection Method | Ref. |
|---|---|---|---|---|---|
| Portugal | 2005–2006 | Feces (rectum content) | 17/77 (22.1) | Culture | [34] |
| Switzerland | 2007–2008 | Feces (not specified) | 0/73 | RT–PCR | [33] |
| 0/73 | Culture (for RT-PCR positive samples) | ||||
| Tonsils | 19/153 (35.8) | RT-PCR | |||
| 8/153 (5.2) | Culture (for RT-PCR positive samples) | ||||
| Italy | 2007–2010 | Feces (intestinal content) | 326/1313 (24.8) | Culture | [36] |
| Spain | 2007–2011 | Feces (rectum content) | 66/214 (30.8) | Culture | [35] |
| Spain | 2009–2011 | Feces (rectum content) | 1/574 (0.17) | Culture | [37] |
| Carcass surface | 5/585 (0.8) | ||||
| Italy | 2010–2012 | Feces (colon content) | 54/499 (10.8) | Culture | [38] |
| Blood (serum) | 255/383 (66.5) | Enzyme-Linked Immunosorbent Assay | |||
| Spain | 2010–2015 | Feces (colon content) | 25/838 (2.9) | Culture | [43] |
| Lymph nodes (sub-mandibular) | 21/415 (5.1) | ||||
| Tonsils | 40/214 (18.7) | ||||
| Italy | 2012–2013 | Muscle (diaphragm or leg) | 7/194 (3.6) | Enzyme-Linked Fluorescent Assay followed by culture for positive samples | [61] |
| Portugal | 2013–2014 | Feces (rectum content) | 1/21 (4.7) | Culture | [39] |
| Serbia | 2013–2014 | Feces (rectum content) | 13/425 (3.1) | Culture | [50] |
| Carcass surface | 4/425 (0.9) | ||||
| Skin surface | 1/425 (0.2) | ||||
| Italy | 2013–2017 | Liver | 260/4335 (6) | PCR followed by culture for positive samples (results referred to cultural positivity) | [62] |
| Sweden | 2014–2016 | Feces (not specified) | 7/90 (7.8) | RT-PCR preceded by culture (results referred to PCR positivity) | [40] |
| Lymph nodes (mesenteric) | 9/90 (10) | ||||
| Lymph nodes (sub-mandibular) | 3/25 (12) | ||||
| Tonsils | 20/136 (14.7) | ||||
| Denmark | 2014–2016 | Feces (rectum content) | 0/115 | Culture | [44] |
| Spain | 2015–2016 | Feces (rectal swabs) | 4/130 (3.1) | Culture | [13] |
| Serbia | 2015 | Carcass surface Skin surface | 4/210 (1.9) 3/210 (1.4) | Culture | [63] |
| Germany | 2016 | Feces (not specified) | 13/552 (2.4) | Culture | [47] |
| Finland | 2016 | Blood (serum) | 69/181 (38) | Enzyme-Linked Immunosorbent Assay | [64] |
| Organs (kidney and spleen) | 6/130 (4.6) | RT-PCR followed by culture for positive samples (results referred to PCR positivity) | |||
| Italy | 2016–2017 | Feces (caecum content) | 4/57 (7) | Culture | [41] |
| Lymph nodes (mesenteric) | 2/57 (3.5) | ||||
| Carcass surface | 0/30 | ||||
| Italy | 2016–2019 | Feces (colon content) | 32/90 (35.6) | Culture | [46] |
| Lymph nodes (mesenteric) | 16/90 (17.8) | ||||
| Carcass surface | 1/90 (1.1) | ||||
| Italy | 2017–2018 | Feces (not specified) | 30/189 (15.9) | Culture | [42] |
| Lymph nodes (mesenteric) | 6/189 (3.2) | ||||
| Italy | 2017–2020 | Feces (not specified) | Not reported | Culture (previous study) | [48] |
| Lymph nodes (mesenteric) | |||||
| Carcass surface | |||||
| Italy | 2018–2019 | Carcass (excision method) | 3/120 (2.5) | Culture | [65] |
| Italy | 2018–2023 | Feces (rectum content) | 1/280 (0.3) | Culture | [49] |
| Carcass surface | 5/280 (1.8) | Enzyme-Linked Fluorescent Assay | |||
| Liver | 3/280 (1.1) | Enzyme-Linked Fluorescent Assay | |||
| Italy | 2018–2020 | Feces (rectal swabs) | 7/287 (2.4) | Culture | [45] |
| Spleen | 6/287 (2) | ||||
| Liver | 5/287 (1.7) | ||||
| Italy | 2019 | Tonsils | 0/36 | Culture | [66] |
| Carcass surface | 0/36 | ||||
| Meat (forearm area) | 1/36 (2.8) | ||||
| Italy | 2020 | Carcass surface | 5/64 (7.8) | Culture | [67] |
| Lymph nodes (mesenteric) | 5/64 (7.8) | ||||
| Italy | 2020–2022 | Feces (colon content) | 3/66 (4.5) | Culture | [11] |
| Lymph nodes (mesenteric) | 0/66 | ||||
| Carcass surface | 0/49 |
RT-PCR: Real-Time Polymerase Chain Reaction.
3.2. Unveiling Y. enterocolitica: Prevalence Trends and Bio-Serotypes
Between the microorganisms of the genus Yersinia, Y. enterocolitica is the species most frequently linked to human disease (yersiniosis). Y. enterocolitica is classified into six biotypes (BT) with varying pathogenicity: 1A (non-pathogenic), 1B (highly pathogenic), and 2–5 (weakly pathogenic). It is further subdivided into over 70 serotypes based on antigenic differences. The main bio-serotypes responsible for human yersiniosis are 1B/O:8, 2/O:9, 2/O:5,27, and 4/O:3 [68,69].
Yersiniosis is currently the fourth most common zoonosis in Europe, with a number of cases of 8738 in 2023 [30]. The pig is considered the main reservoir of Y. enterocolitica, from which strains belonging to pathogenic bio-serotypes for humans are most frequently isolated, while reports on the occurrence of Y. enterocolitica in wild boars are scarcer, and the epidemiological link with domestic pigs is still unknown [70].
As regards pigs, the tonsils represent the ecological niche of the microorganism [71,72] and this source is usually analyzed also in wild boars’ investigations.
Table 2 summarizes the prevalence of Y. enterocolitica detected in different sample types (feces, lymph nodes, carcass surface, meat, tonsils, organs, blood) collected from wild boars in various European Union countries. The sampling period is indicated. Prevalence is expressed as the number of positive samples out of the total number examined, with the corresponding percentage reported in brackets. Details regarding the detection method are also provided: culture (cold and/or warm enrichment) and/or molecular or immunoenzymatic assays.
Comparison between studies is not always straightforward. In addition to the factors already mentioned for Salmonella spp.—such as sample size-variability is also influenced by the different cultural methods employed. In fact, warm and/or cold cultures are often applied in parallel to exploit the uncommon ability of this enteric pathogen to replicate at refrigeration temperatures [69]. Cold enrichment remains more sensitive and reliable for detecting Y. enterocolitica in animal matrices, though it is slower, as demonstrated by different investigations carried out in wild boars [73,74,75]. Warm enrichment may be useful for rapid screening, but combining both methods increases overall detection probability and strain diversity [73,74,75].
As in domestic pigs, the tonsils appear to act as the main niche for the microorganism, with prevalence rates as high as 35% reported in Switzerland [33] and 17.9% in Germany [70]. In the latter study [70], 89.5% of isolates were identified as BT 1A, while the remaining belonged to BT 1B. Although BT 1B is generally considered highly pathogenic for humans, in this case the isolates lacked the virulence plasmid, raising questions about their pathogenic potential. In the same study [70], the highest prevalence was observed in animals aged 12–24 months, although the influence of age was not statistically evaluated. Multi Locus Sequence Typing (MLST) analysis revealed a high degree of heterogeneity among BT 1A isolates, despite samples being collected within a small hunting area during a single hunting season.
In fecal samples, prevalence ranges between 0% and 27% [11,33,67,75,76]. As in the tonsils, most isolates were identified as BT 1A, but other BTs, including serotypes 2/O:9 and 4/O:3—commonly associated with human yersiniosis and pigs—were also detected, albeit in low numbers. Syczylo et al. [74] in Poland identified BT 1B in isolates from rectal swabs samples. These isolates only harbored the ystB gene, which is associated with strains of low or uncertain pathogenic potential, but its presence suggests the capability to induce gastrointestinal symptoms, especially in susceptible subjects [77].
Regarding lymph nodes, prevalence rates of 5–6% were found in the mesenteric district [13,40]. Interestingly, Sanno et al. [40] reported a higher prevalence (12%) in submandibular lymph nodes, similar to that detected in tonsils (14%), presumably due to the anatomical proximity between these sites.
Several studies have reported seasonal variations, with higher prevalence rates observed in autumn and winter, coinciding with the wild boar hunting season in most European countries. This trend is likely related to stress factors such as low temperatures and food scarcity [13,78,79], but also to the already mentioned ability of Y. enterocolitica to multiplicate at low temperatures.
Interestingly, the presence of high-density ovine populations has also been identified as a factor influencing Y. enterocolitica prevalence in wild boars, as reported by Arrausi-Subiza et al. [78]. Sheep have been described as reservoirs of Y. enterocolitica [80,81], and sheep milk has also been identified as a source of potentially pathogenic strains [82], highlighting once again the relevance of a One Health perspective in understanding pathogen transmission dynamics at the wildlife–livestock–environment interface.
Table 2.
Y. enterocolitica prevalence expressed as positive/total (%) in different sample types collected from wild boars in various countries of the European Union.
Table 2.
Y. enterocolitica prevalence expressed as positive/total (%) in different sample types collected from wild boars in various countries of the European Union.
| Country | Sampling Period | Source | Pos/Tot (%) | Comments on Detection Method | Ref. |
|---|---|---|---|---|---|
| Switzerland | 2007–2008 | Feces (not specified) | 4/73 (5.5) | RT-PCR | [33] |
| 1/73 (1.3) | Culture (direct plating of RT-PCR positive samples) | ||||
| Tonsils | 26/73 (36) | Real-Time PCR | |||
| 6/73 (8) | Culture (direct plating of RT-PCR positive samples) | ||||
| Italy | 2008–2010 | Carcass surface | 3/251 (1.2) | Culture (cold enrichment) | [83] |
| Spain | 2009–2012 | Tonsils | 24/72 (33.3) | RT-PCR followed by cultural (direct plating of RT-PCR enrichment of RT-PCR positive samples) | [78] |
| Poland | 2012–2013 | Feces (rectal swabs) | 40/151 (26.5) | Culture (warm and cold enrichment) | [75] |
| Germany | 2012–2013 | Tonsils | 19/111 (17.1) | Culture (warm enrichment) | [70] |
| Italy | 2012–2013 | Muscle (diaphragm and leg) | 34/230 (14.8) | Culture (cold enrichment) | [61] |
| Czech Republic | 2013–2014 | Meat juice from diaphragm | 89/135 (81.9) | Enzyme-Linked Immunosorbent Assay (antibodies anti-Yersinia spp.) | [84] |
| Poland | 2013–2014 | Feces (rectal swabs) | 110/434 (25.3) | Culture (warm and cold enrichment) | [74] |
| Italy | 2013–2018 | Liver | 126/4890 (2.6) | Culture (warm enrichment) | [85] |
| Italy | 2014–2015 | Muscle (Longissimus dorsi) | 0/22 | Culture (cold enrichment) | [86] |
| Sweden | 2014–2016 | Feces | 4/90 (4.4) | RT-PCR | [40] |
| Lymph nodes (mesenteric) | 6/67 (6.7) | ||||
| Lymph nodes (sub-mandibular) | 3/25 (12) | ||||
| Tonsils | 19/136 (14) | ||||
| Italy | 2015–2018 | Feces (not specified) | 0/107 | RT-PCR followed by culture (warm enrichment) for positive samples | [76] |
| Finland | 2016 | Blood (serum) | 102/181 (56) | Enzyme-Linked Immunosorbent Assay (antibodies anti-Yersinia spp.) | [64] |
| Organs (kidney, spleen) | 22/130 (17) | RT-PCR followed by culture (warm enrichment) for positive samples | |||
| Italy | 2017 | Muscle (shoulder area) | 2/22 (9) | RT-PCR followed by culture (cold enrichment) for positive samples | [87] |
| Italy | 2017–2019 | Feces | 19/305 (6.2) | Culture (warm and cold enrichment) | [67] |
| Lymph nodes (mesenteric) | 10/305 (3.3) | ||||
| Italy | 2018–2020 | Feces (rectal swabs) | 54/287 (18.8) | Culture (cold enrichment) | [45] |
| Italy | 2019 | Carcass surface | 12/36 (33.3) | Culture (warm enrichment) | [66] |
| Tonsils | 9/36 (25) | ||||
| Meat (forearm area) | 10/36 (27) | ||||
| Italy | 2020 | Lymph nodes (mesenteric) | 0/64 | Culture (cold enrichment) | [79] |
| Carcass surface | 0/64 | ||||
| Italy | 2020–2022 | Feces (colon content) | 18/66 (27.3) | Culture (cold enrichment) | [13] |
| Lymph nodes (mesenteric) | 3/66 (4.5) | ||||
| Carcass surface | 3/49 (6.1) |
RT-PCR: Real-Time Polymerase Chain Reaction.
3.3. Campylobacter spp. Occurrence and Epidemiological Insights
Campylobacter spp. is recognized as a major global public health concern accounting for 58.9% of all the reported and confirmed cases of zoonotic diseases in 2023 [30] with the main causative species of campylobacteriosis being C. jejuni and C. coli, which are known to have their primary reservoir in avian species, particularly chickens [88], but have also been isolated from wild animals, along with other species as C. lari and C. lanienae.
Table 3 summarizes the prevalence of Campylobacter spp. detected in different sample types (feces, lymph nodes, carcass surface, tonsils, organs, meat, bile) collected from wild boars in various European Union countries. The sampling period is indicated. Prevalence is expressed as the number of positive samples out of the total number examined, with the corresponding percentage reported in brackets. When available, Campylobacter identified species were reported.
In feces samples Campylobacter spp. prevalence can be comprised in a range between 0 and 66% [13,33,35,41,89,90]. This huge variability can be due to the fact that most studies apply methods that favor the growth of thermotolerant species (as C. jejuni and C. coli), thus excluding the possibility to detect also non-thermotolerant campylobacters, such as C. lanianae, C. hyointestinalis and C. fetus and underestimating the real prevalence [14]. Particularly C. lanianae, recognized as potential causes of human illness, is predominantly isolated from wild boars, domestic pigs and feral swine with prevalence which can be as high as 70% in Spain [13,89] and 40.8% in Italy [14,90]. However, many authors have also isolated C. coli with high prevalence rates. This is the case, for example, of the investigation of Castillo-Contreras et al. [13] who made a comparison of wild boar carriage of Campylobacter spp. in three different zones of the metropolitan area of Barcelona, with three different degrees of urbanization. Although no clear relationship was found between urbanization and specific Campylobacter species, C. lanienae was more frequently isolated in the less urbanized area, suggesting a natural diet as a potential source of infection. Conversely, an anthropogenic source of C. coli infection (e.g., exposure to rubbish) could be hypothesized, as its prevalence was twice as high in more urbanized areas compared to the less urbanized one. This pattern implies that increasing urbanization may shift wild boar exposure towards human-associated sources of contamination (e.g., refuse, wastewater, food scraps), thereby influencing both the composition of their enteric microbiota and their potential to act as vectors of pathogens with relevance for public health.
Some authors investigated the effect of potential influencing factors on Campylobacter spp. in wild boars: sex, weight, age did not reveal a significant impact on both organs and carcasses. Campylobacter spp. prevalence on wild boar carcasses was higher, although not significant, with environmental temperature on the day of hunting above 15 °C [41]. In the study by Carbonero et al. [89], the presence of artificial waterholes was significantly associated with an increased prevalence of Campylobacter spp. in wild boars. This is likely due to the ability of certain Campylobacter species to survive in aquatic environments, with outbreaks in humans linked to contaminated water having also been reported [91,92].
According to the same authors, Campylobacter spp. occurrence was significantly higher during the winter months. This may be due to the fact that lower temperatures support the bacterium’s ability to survive in the environment [89]. Moreover, experimental studies have shown that Campylobacter spp. can persist in water for longer periods at low temperatures, which could explain the observed association between waterholes and its presence in wild boars [93].
Table 3.
Campylobacter spp. prevalence expressed as positive/total (%) in different sample types collected from wild boars in various countries of the European Union.
Table 3.
Campylobacter spp. prevalence expressed as positive/total (%) in different sample types collected from wild boars in various countries of the European Union.
| Country | Sampling Period | Source | Pos/Tot (%) | Campylobacter Species Identified (%) | Ref. |
|---|---|---|---|---|---|
| Germany | 2006–2007 | Muscle (various carcass area) | 3/127 (2.4) | C. coli (66.6), C. jejuni (33.3) | [94] |
| Spain | 2009–2011 | Feces (rectal content) | 188/287 (66) | C. spp. (one C. jejuni) | [37] |
| Spain | 2010–2011 | Feces (rectal content) | 10/41(24.4) | C. coli (21.1), other thermophilic species (79.9) | [35] |
| Spain | 2011–2012 | Feces (intestine or rectum) | 49/126 (38.9) | C. lanianae (69.4), C coli (16.3), C. jejuni (4.11), others (10.2) | [89] |
| Italy | 2012–2019 | Feces (not specified) Liver Muscle | Not reported | C. coli (91.7), C. jejuni (8.3) | [95] |
| Italy | 2016 | Feces (caecum content) | 29/56 (51.8) | Not investigated | [41] |
| Lymph nodes (mesenteric) | 0/56 | ||||
| Carcass surface | 5/30 (16.7) | ||||
| Spain | 2015–2016 | Feces (rectal swabs) | 79/130 (60.8) | C. lanienae (46.2), C. coli (16.2), C. hyointestinalis (0.8) | [13] |
| Italy | 2018–2019 | Feces (rectal swabs) | 78/183 (42.6) | C. coli (48), C. lanianae (42), C. jejuni (6), C. hyointestinalis (4) | [90] |
| Carcass surface | 10/55 (18.2) | ||||
| Liver | 9/187 (4.81) | ||||
| Bile | 3/152 (1.9) | ||||
| Italy | 2019 | Feces (rectal content) | 38/76 (50) | C. lanienae (40.8), C. hyointestinalis (14.5), C. coli (7.9), C. jejuni (1.3), C. fetus (1.3) | [14] |
| Italy | 2019 | Carcass surface | 4/36 (11.1) | C. coli (33), C. jejuni (33), other species (33) | [66] |
| Meat (leg) | 2/36 (5.5) | ||||
| Italy | 2019–2020 | Meat (leg) | 0/28 | [87] |
3.4. Prevalence and Diversity of STEC
The pathogenic E. coli pathotypes include enteroaggregative E. coli (EAEC), enteropathogenic E. coli (EPEC) and Shiga toxin-producing E. coli (STEC), and its subgroup of enterohemorrhagic E. coli (EHEC) [12]. STEC are of particular concern for public health and STEC infections were the third cause of zoonoses with over 10,217 cases in humans in 2023 [30]. Their pathogenicity is primarily linked to the production of Shiga toxins (Stx1, Stx2 and variants). In humans, STEC infections can result not only in acute gastroenteritis but also in severe systemic complications with the most serious outcome being the hemolytic uremic syndrome (HUS), a condition defined by hemolytic anemia, thrombocytopenia, and acute renal failure [96]. The best-known STEC serotype is O157:H7, which has been associated with numerous large-scale outbreaks worldwide. However, non-O157 serotypes such as O26, O103, O111, and O145 are recognized as significant causes of disease [97].
STEC transmission mainly occurs via food and water contaminated with fecal material [98]. While many domestic and wild animals act as asymptomatic carriers due to the absence of Shiga toxins receptors, wildlife—particularly ruminants such as deer—represents an important reservoir contributing to environmental contamination and the spread of infection [99].
Also, wild boars have been described as carriers of E. coli O157:H7 and other non-0157 STEC strains that are potential human pathogens.
Table 4 summarizes the prevalence of STEC detected in different sample types (feces, lymph nodes, tonsils, carcass surface, meat, and meat products) collected from wild boars in various European Union countries. The sampling period is indicated. Prevalence is expressed as the number of positive samples out of the total number examined, with the corresponding percentage reported in brackets. When available, STEC serogroups (reported as non-O157 and O157:H7) and other identified E. coli pathotypes are also indicated, with the corresponding percentages provided in brackets.
Prevalence in feces samples of non-O157 STEC can vary between 3 and >28% [12,37,47,100,101,102,103,104].
In some investigations, lower prevalence, or even no STEC detection, could be influenced by the fact that only O157:H7 presence is investigated [105,106]. Although most of the investigations mostly identified non-O157 isolates, some authors found serogroups associated with clinical case. Dias et al. [103] identified O27 serogroup, associated with hospitalization with neonatal HUS and bloody diarrhea [97]. In a study conducted in Italy [107], beside a quite high STEC prevalence (21.7%) in rectal swabs, also a 6.3% of EHEC was identified, harboring the virulence gene eae, responsible for the typical attaching and effacing lesions and associated with EPEC. In Spain, Mora et al. [100] identified 0.38% of O157:H7 isolates from wild boars and other serotypes associated with HUS and human outbreaks, as O126:H11 and O128:H2, as well as serotypes associated with diarrhea as O146.
Other pathotypes are often identified from wild boar samples as EPEC [101,102,108], EHEC [107], but also Extra-intestinal pathogenic E. coli as uropathogenic [108].
Table 4.
STEC prevalence expressed as positive/total (%) in different sample types collected from wild boars in various countries of the European Union. When possible, distinction between serotype O157:H7 and other serotypes (non-O157) has been reported.
Table 4.
STEC prevalence expressed as positive/total (%) in different sample types collected from wild boars in various countries of the European Union. When possible, distinction between serotype O157:H7 and other serotypes (non-O157) has been reported.
| Country | Sampling Period | Source | Pos/Tot (%) | STEC Serogroups (%) and Others Pathotypes Identified (%) | Ref. |
|---|---|---|---|---|---|
| Portugal | 2009–2010 | Feces (rectal swabs) | 22/262 (8.4) | non-O157 (8.4), O157:H7 (0.38) | [100] |
| Spain | 2009–2011 | Frozen meat | 1/36 (2.7) | non-O157 (100%) | [109] |
| Meat products | 2/21 (9.5) | ||||
| Spain | 2009–2011 | Feces (rectal content) | 4/117 (3.4) | Detection method only for O157:H7 | [106] |
| Sweden | 2010–2011 | Feces (not specified) | 0/88 | Detection method only for O157:H7 | [105] |
| Lymph nodes (mesenteric) | 0/56 | ||||
| Tonsils | 0/175 | ||||
| Spain | 2009–2011 | Feces (rectal content) Carcass surface | 11/301 (4) 12/310 (4) | non-O157 (100%) | [37] |
| Portugal | 2013–2014 | Feces (rectal content) | 1/21 (4.8) | non-O157 (100%) | [103] |
| Spain | 2013–2015 | Feces (not specified) | 3/90 (3.3) | non O157 (100%) E. coli EPEC (3.3) | [101] |
| Germany | 2016 | Feces (not specified) | 37/536 (6.9) | O157:H7 (8.3) non-O157 (91.7) | [47] |
| Poland | 2017–2018 | Feces (rectal swabs) | 64/152 (28.3) | non-O157 (92.1), O157 (7.9) E. coli EPEC (17.11) | [102] |
| Portugal | 2017–2019 | Feces (environment or rectal content) | 8/56 (14) | non-O157 (100%) | [12] |
| Italy | 2018–2019 | Feces (rectal swabs) | 13/200 (6.5) | Serogroups not reported EHEC (6.3), EAEC (5.7), aEPEC (3.4), and unspecific pathotypes also identified | [107] |
| Italy | 2019–2020 | Meat (forearm area) | 12/28 (42.8) | Serogroups not reported | [66] |
| Switzerland | 2021 | Meat (not specified) | 3/25 (12) | non O157 (100%) | [110] |
| Switzerland | 2022–2023 | Feces (colon content) | 13/59 (22) | non O157 (100%) | [104] |
STEC: Shiga toxin-producing E. coli; EPEC: enteropathogenic E. coli; EHEC: enterohemorrhagic; EAEC. enteroaggregative E. coli; aEPEC: atypical enteropathogenic E. coli.
3.5. Wild Boar as Carriers of Enteric Pathogens
Carriers were defined as animals testing positive in feces and/or lymph nodes and/or tonsils. Their relevance lies in the fact that infected wild boars may contaminate the environment through fecal dispersion, carcass or internal organs during dressing operations, or transmit pathogens directly to humans or to the environment during handling. This aspect is particularly critical when considered within a One Health perspective, as carriers may represent a bridge for pathogen dissemination across wildlife, domestic animals, humans, and environmental compartments.
Supplementary Figures S1–S4 illustrate the prevalence of carrier wild boars reported in different European Union countries. As previously noted, these values must be interpreted with caution due to differences in diagnostic methods, sampling strategies, and analytical approaches, which may influence the outcomes.
Overall, the prevalence of carrier wild boars testing positive in feces and/or lymph nodes and/or tonsils ranges from approximately 0 to 24.8% for Salmonella spp. [11,13,33,34,35,36,37,38,39,40,41,42,43,44,45,46,47,49,50,66,67], and from 8.2 to 33.3% for Y. enterocolitica [13,33,40,45,66,67,70,74,75,76,78,79]. For Campylobacter spp. and STEC, carrier prevalence has been assessed at the fecal level only and ranges from 24.4 to 66% [13,14,35,36,37,41,89,90,95] and 3.3 to 28.3%, respectively [12,37,47,100,101,102,103,104,105,106,107].
Several authors investigated more than one pathogen simultaneously, and in some cases, co-infections were reported. For example, Cilia et al. [45] detected both Salmonella enterica subs. houtenae and Y. enterocolitica in the feces of one wild boar. Sanno et al. [40] reported two animals positive for both Salmonella spp. and Y. enterocolitica in the tonsils, whereas Siddi et al. [11] identified two wild boars positive for both pathogens in feces or lymph nodes. Other studies screened for multiple pathogens (e.g., Salmonella, Campylobacter, STEC, Y. enterocolitica) but did not report co-infections [13,35,37,41].
The identification of carrier wild boars underscores their potential role in food safety hazards and AMR dissemination, reinforcing the need to consider this species as an integral component of One Health surveillance programs.
3.6. Focus on the Risk of Meat Contamination; Presence of Zoonotic Pathogens on Carcass Surface, Organs, and Meat
Several factors influence the microbiological quality of game meat, including the microorganisms carried by the animal, hunting conditions, and practices during butchering, handling, and storage [65]. Consequently, the safety of wild boar meat depends both on the health status of the hunted animal and on the hygiene conditions of slaughtering and processing environments. These aspects may lead to meat contamination and pose a risk of zoonotic infection to consumers [111]. For this reason, it is particularly important to assess the presence of enteric zoonotic pathogens on the carcass surface and, when relevant, in muscles and edible organs such as the liver. Nevertheless, few studies include these matrices in their investigations.
Salmonella spp. has been detected on carcasses of hunted wild boars with low prevalence rates, ranging between 0 and approximately 2.5%, when evaluated using swab or sponge methods according to ISO 17604 or comparable techniques [11,37,41,46,50,63,66,67], either after skinning [49] or through the excision method [65].
Y. enterocolitica has been detected with prevalence rates ranging from 0 up to over 30% on carcass surfaces, and although only biotype 1A is usually identified, these results highlight a potential risk for consumers [11,66,83]. When investigated at muscle level, Y. enterocolitica has been detected with variable prevalence: 0% in Longissimus dorsi [86], 9% in shoulder samples [87], 14.8% in diaphragm and leg [61], and 27% in the forearm area [66].
In northern Italy (Liguria), Modesto et al. [85] investigated Y. enterocolitica in 4890 wild boar liver samples, reporting a prevalence of 2.6%. Most isolates (92.9%) were identified as biotype 1A, 3% as biotype 1B, and <1% as biotype 2. Notably, among the 1B isolates, two belonged to serotype O:8, one of the most frequent in human yersiniosis, suggesting a possible anthropogenic origin.
Liver samples have also been investigated for the presence of other pathogens, such as Salmonella spp.—with reported prevalence rates ranging from 1.7% to 6% [45,49,62]—and Campylobacter [95], in which the most common species involved in human campylobacteriosis (C. coli and C. jejuni) were detected, although prevalence data were not reported in this case.
The isolation of potentially pathogenic Y. enterocolitica strains and other enteric pathogens from wild boar liver is particularly relevant, as this organ is traditionally consumed only lightly cooked or even used raw in the preparation of certain meat products in Italy [62].
Campylobacter spp. was detected in 16.7% of wild boar carcasses in northern Italy [41]. In that study, the authors observed a correlation between bacterial load—expressed as total viable count (TVC) and Enterobacteriaceae—and the presence of Campylobacter spp. on carcasses, with prevalence increasing from 16.7% to 23% and 25% at TVC levels of 3–4 and >4 log CFU/cm2, respectively. Other studies reported prevalence ranging between 2% and 18% [66,90,94], with C. coli, C. jejuni, and C. lanienae being the most frequently identified species.
As for STEC, prevalence rates of 12% in meat samples from the forearm area [87], 9.5% in meat products and 2.7% in frozen meat samples have been reported [109]. Some isolates, although non-O157, belonged to serotypes and carried virulence genes associated with human infections, thus representing a potential public health concern.
More rarely, the presence of enteric pathogens has been investigated in other organs (e.g., kidneys, spleen) or in different types of samples such as blood and meat juice extracted from diaphragm, with highly variable prevalence results, mostly depending on the analytical method applied–as real-time PCR, Enzyme-Linked Immunosorbent Assay (ELISA)-rather than culture-based methods [64,84].
Overall, these results highlight the need for continuous monitoring and for the implementation of Good Hygiene Practices (GHP) and Good Manufacturing Practices (GMP) throughout wildlife hunting, field dressing, and carcass processing. Such measures are essential to minimize contamination risks and ensure the safety of wild boar meat, which may otherwise serve as a potential vehicle for zoonotic pathogens.
Table 5 summarizes the prevalence ranges of Salmonella spp., Y. enterocolitica, STEC, and Campylobacter spp., expressed as minimum–maximum percentages, detected in different sample types collected from wild boars across various European Union countries. These ranges reflect the substantial heterogeneity among the included studies, which differ in sampling design, matrices, and analytical methodologies (cultural, molecular, or immunoenzymatic). As a result, the reported values should be interpreted as broad indicative intervals rather than directly comparable prevalence estimates.
Table 5.
Prevalence ranges of Salmonella spp., Y. enterocolitica, STEC, and Campylobacter spp., expressed as minimum–maximum percentages, detected in different sample types collected from wild boars across various European Union countries.
4. Antimicrobial Resistance Profiles of Enteric Pathogens Detected from Wild Boars in European Union
Despite a vast and growing body of literature on AMR in medical and veterinary contexts, research addressing the complex transmission dynamics of AMR in environmental and wildlife compartments remains scarce, although such investigations are essential to fully operationalize the One Health approach [112].
In theory, wildlife animals are not exposed to antibiotic treatments; however, their direct and indirect interactions with livestock, domestic animals, and human-influenced environments, combined with their ability to move freely across diverse habitats, increase their likelihood of encountering selective agents, commensal bacteria, and resistant microorganisms [5]. Such interactions are thought to facilitate adaptive processes in commensal bacteria and promote the horizontal transfer of resistance genes within wildlife bacterial communities [113].
Wild boars, in particular, are increasingly recognized as potential sources of resistant foodborne pathogens for humans, primarily through the handling and consumption of their meat [114,115].
From the analysis of the scientific literature, it emerges that the prevalence rates of antibiotic resistance, as well as the phenotypic and genotypic resistance profiles found in enteric pathogens isolated from wild boars in various European Union countries, show considerable variability depending on the geographical location. Several factors contribute to this heterogeneity. Differences in environmental contamination levels, livestock density, and farming practices strongly influence the exposure of wild boars to resistant bacteria and selective agents [5,47,113]. Moreover, variations in sampling strategies (e.g., target organs, sample size, and season of collection), bacterial species investigated, and laboratory methodologies—most commonly the Kirby–Bauer disk diffusion test or the broth microdilution method (MIC)—further affect the comparability of results across studies [5,113]. Finally, the ecological behavior and movement patterns of wild boar populations, which differ markedly across European regions, may also contribute to the spatial heterogeneity of AMR occurrence in this species [112].
A useful approach adopted in recent studies is the interpretation of phenotypic AMR profiles based on the epidemiological cut-off values (ECOFFs), which allow the distinction between wild-type (WT) and non-wild-type (NWT) strains with acquired resistance [116,117]. This provides essential information to facilitate comparison between studies, to understand AMR dissemination dynamics and to assess the potential threat it may pose to public health.
In the analyzed literature, strains are defined as multidrug-resistant (MDR) when they are resistant to at least one antimicrobial in three or more classes, while they are defined as extensively drug-resistant (XDR) when they are resistant to at least one antimicrobial in all but one or two of the tested classes [118].
Figure 3 report the number of resistant Salmonella spp., Y. enterocolitica, Campylobacter spp. and STEC detected from wild boars referred to antibiotic classes.
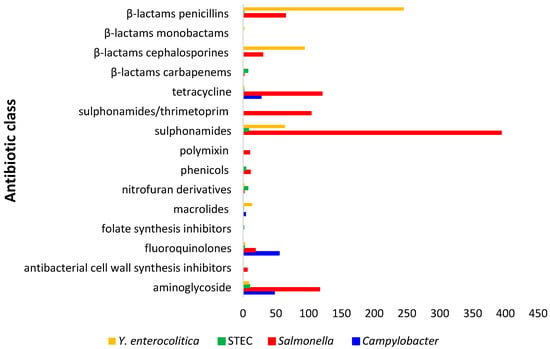
Figure 3.
Number of isolates of Salmonella spp., Y. enterocolitica, STEC and Campylobacter spp. resistant to different antibiotic classes.
In Figure 4, a heat map illustrating and comparing the antibiotic resistance profiles of Salmonella spp., Yersinia enterocolitica, STEC, and Campylobacter spp. isolated from wild boars is showed. The heat map displays the percentage of resistant isolates for each bacterial species across a panel of tested antibiotics. Colors range from yellow (low resistance) to dark red (high resistance), allowing visual identification of critical resistance patterns. Each row corresponds to an antibiotic, while each column represents a bacterial group, enabling side-by-side comparison of their resistance frequencies.
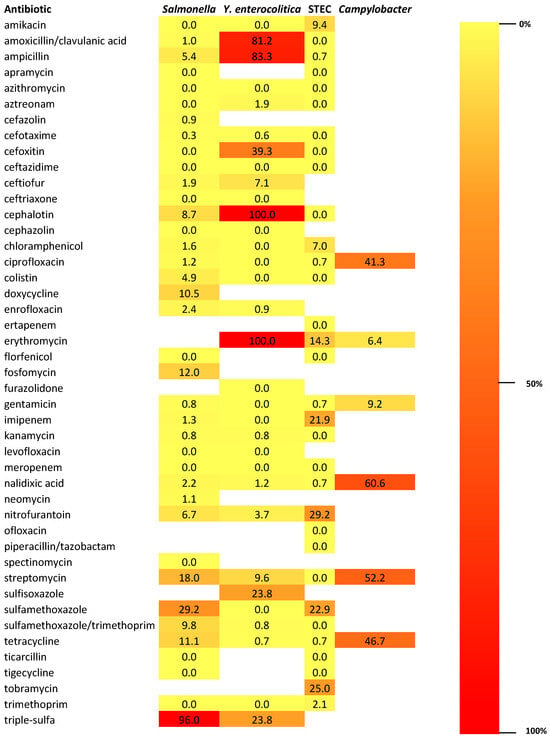
Figure 4.
Heat map illustrating and comparing the antibiotic resistance profiles of Salmonella spp., Yersinia enterocolitica, STEC, and Campylobacter spp. isolated from wild boars. The sidebar represents the color scale used in the heat map, showing how colors correspond to resistance levels—from 0% (light yellow) to 100% (dark red). Triple-sulfa: combination of sulfadiazine, sulfamerazine, and sulfamethazine.
It is well known that antibiotics used in veterinary medicine and agriculture often belong to the same classes as those employed in human medicine [119]. Among enteric pathogen isolated from wild boars, resistant strains have also been identified to antibiotic classes categorized as critically important, as Enterobacterales resistant to carbapenems and third-generation cephalosporins, and highly important, as non-typhoidal Salmonella spp. resistant to fluoroquinolones [62].
4.1. AMR Patterns in Salmonella Isolates
As regards Salmonella spp., high prevalence rates of resistance to sulfonamides have been reported. Resistance rates exceeding 85% for sulfamethoxazole were found in Italy [38] and in Spain [120]. Similarly, Razzuoli et al. [62] reported a resistance prevalence rate of 96% against triple-sulfa compounds. High resistance levels have also been observed for streptomycin and tetracycline with prevalence rates of 41.7% for both antibiotics in Italy [42] and of 46.2% and 25.3% in Spain [120] for streptomycin and tetracycline, respectively. Notably, even higher streptomycin resistance (61.1%) was detected by Cilia et al. [45].
AMR in Salmonella Isolates Against Critically Important and High Important Antibiotics
A particularly concerning finding is the detection of resistance to colistin in Italy and Spain [38,47,62]. This result is of particular concern because colistin represents one of the few remaining therapeutic options against multidrug-resistant Gram-negative bacteria, and the emergence of plasmid-mediated resistance genes (such as mcr variants) facilitates their rapid spread across bacterial species and ecosystems [121]. Piras et al. [46] identified Salmonella spp. strains resistant to fosfomycin, along with the presence of the corresponding resistance gene FosA7. Fosfomycin may represent a valuable therapeutic option for the treatment of cystitis caused by extended-spectrum β-lactamase (ESBL)-producing Enterobacteriaceae [122]. Notably, this resistance was detected in Salmonella spp. strains belonging to the serovars Agona and Derby, which were among the fifteen most frequently reported serovars in human salmonellosis cases in Europe in 2023 [30]. In the study by Zottola et al. [38], in addition to the high resistance rates reported, 54% of the isolates were classified as MDR. Alarmingly, resistance was also detected—albeit at low levels—against third-generation cephalosporins (ceftiofur and cefotaxime), which are classified by the World Health Organization (WHO) as critically important antimicrobials. Similar resistance trends were observed by Razzuoli et al. [62] and Gil-Molino et al. [120], who additionally observed resistance to fluoroquinolones, which belong to the WHO’s high-priority group for antimicrobial resistance.
4.2. AMR Patterns in Y. enterocolitica Isolates
As regards Y. enterocolitica, high rates of resistance to β-lactam antibiotics, such as ampicillin and first- and second-generation cephalosporins, due to β-lactamase production, are well documented in the literature and recognized by both the Clinical and Laboratory Standards Institute (CLSI) and EUCAST [123,124]. An interesting result emerged from the investigation by Modesto et al. [85], who observed an increase in resistance to sulfonamide compounds, particularly sulfisoxazole and triple-sulfa, between 2014 and 2017, reaching 35% of resistant strains for both drugs by the end of the observation period. Notably, the same study also reported resistance to ceftiofur in 7% of the isolates and a substantial increase in MDR strains from 9.5% to 40% over the same period. Furthermore, resistance to erythromycin has occasionally been reported not only in wild boars [125] but also in other wild species [76].
4.3. AMR Patterns in STEC Isolates
AMR is not often determined for STEC isolates, as antibiotic treatment of human infections caused by STEC can enhance the production of Shiga toxins, potentially worsening clinical outcomes [126,127]. When AMR is investigated in STEC isolates from wild animals, high levels of susceptibility are generally reported [76,109]. By applying epidemiological cut-off values (ECOFFs), Dias et al. [12] identified non-wild-type (NWT) phenotypes in STEC strains isolated from wild boar fecal samples for nitrofurantoin and sulfamethoxazole/trimethoprim (100%), imipenem (classified by WHO as critically important), and tobramycin (87.5%), as well as for amikacin (37.5%) and chloramphenicol (25%). Interestingly, full susceptibility had been reported for the same isolates using the disk diffusion method. The same study also found a high proportion (87.5%) of multidrug-resistant (MDR) strains. Similarly, a previous investigation conducted in Germany identified 4% of NWT STEC isolates from wild boars showing resistance to compounds belonging to seven different antimicrobial classes, including fluoroquinolones and β-lactams [47].
4.4. AMR Patterns in Campylobacter spp. Isolates
Few studies have investigated AMR in Campylobacter spp. isolates from wild boars. Carbonero et al. [89] in Spain examined several wild artiodactyl species, including wild boars and reported high levels of resistance to erythromycin (95.2%), followed by ciprofloxacin (62.5%), tetracycline (47.1%), and streptomycin (45%), but results were not differentiated between the animal species. Similar resistance trends were observed in a more recent study by Castillo-Contreras et al. [13], who detected high resistance rates to tetracycline and ciprofloxacin (both 66%) and to streptomycin (43%). Additionally, they reported very high resistance to nalidixic acid (95%), with none of the isolates being pansusceptible, and 67% of C. coli strains classified as MDR. The AMR profiles observed in C. coli, along with their detection in areas characterized by a high or medium degree of urbanization, suggest an anthropogenic origin for these resistant strains. Moreover, resistance to ciprofloxacin is particularly alarming, as it is one of the two antimicrobials considered critically important for the treatment of human campylobacteriosis [128].
5. Future Perspective in AMR: Studying the Fecal Resistome
Studies on antimicrobial resistance (AMR) in wild animals often face several limitations: (1) variability in methods used across studies; (2) relatively small sample sizes; (3) experimental designs driven by sampling convenience and short timeframes; (4) limited numbers of wild species investigated, with wild boars being the most frequently studied; and (5) predominantly descriptive approaches that provide only a snapshot of AMR occurrence. These limitations result in poor spatial and temporal coverage of AMR data and leave significant gaps in understanding the mechanisms of AMR acquisition and transmission routes [25,129].
A promising approach was proposed by Dias et al. [12], who examined the presence and diversity of antibiotic resistance genes (ARGs) in wild boar fecal microbiomes using metagenomic and culture-based methods (high-throughput qPCR and phenotypic testing of E. coli and Enterococcus spp.). In 56 samples from three Portuguese areas with different levels of human impact, they identified 62 ARGs associated with nine antibiotic classes—mainly tetracyclines and aminoglycosides—and 20 mobile genetic elements, including integrons. Regions with greater anthropogenic pressure exhibited higher ARG diversity and abundance, highlighting the influence of human activity.
Phenotypic resistance testing of E. coli and Enterococcus spp. isolates supported the molecular findings. Overall, 27% of E. coli and 83% of Enterococcus spp. were resistant to at least one antibiotic, primarily β-lactams, aminoglycosides, macrolides, and tetracyclines, with relatively few multidrug-resistant strains. When evaluated using epidemiological cut-off values (ECOFFs), 45% of E. coli and 38% of Enterococcus spp. were classified as non-wild-type (NWT), indicating acquired resistance beyond the natural susceptibility range, while the remaining strains were wild-type (WT), displaying only intrinsic resistance. The proportion of NWT strains was higher in areas with stronger human influence.
These results suggest that wild boars can serve as ecological sentinels for monitoring AMR in the environment. The study underscores the importance of including wildlife in national AMR surveillance programs and highlights the value of a One Health approach to safeguard both public and environmental health.
6. From Forest to City: The Wild Boar as an Example of Synurbization
The rise in wild boar populations and their expansion, driven by the previously mentioned factors, has been accompanied by synurbization—a specific form of synanthropization that refers to the adaptation of animal populations to human-modified environments [130,131]. This process fosters new interactions between wild boars and humans, often leading to conflicts, potential attacks on pets or people, traffic accidents, and increased zoonotic risks [132].
The Barcelona Metropolitan Area (MAB) represents an exemplary One Health setting, with one of the most thoroughly documented cases of wild boar synurbization. Population densities ranged from 5–15 individuals per km2 between 2004 and 2022 [133,134,135]. The area encompasses a gradient of urbanization: the city of Barcelona and the Universitat Autònoma de Barcelona (UAB) are predominantly urban, though the UAB retains more gardens, forests, and agricultural patches (60%) than the city (28%). In contrast, the nearby Serra de Collserola Natural Park (CNP) consists mainly of natural habitats, including scrubland, forest, and grassland, interspersed with recreational spaces and built-up areas [136]. This habitat mosaic provides a unique context to study how wild boars adapt to varying degrees of urbanization.
As previously mentioned, Castillo-Contreras et al. [13] examined the occurrence of zoonotic Campylobacter spp. in wild boar feces from the MAB, aiming to assess the genetic diversity of isolates, their possible link with anthropogenic sources, and the relationship between Campylobacter spp. carriage and the level of urbanization. Among the species identified, C. lanienae showed a prevalence of 46%, being more common in wild boars from less urbanized areas. This suggests that infection is likely associated with a diet based on natural resources rather than human-derived sources. Conversely, the higher prevalence of C. coli observed in the two most urbanized areas—approximately double that found in the less urbanized zone—points to a probable anthropogenic origin of C. coli infections. Similarly, Darwich et al. [137] reported E. coli strains carrying critical β-lactam resistance genes in the MAB. Wild boars foraging in urban and peri-urban areas were found to be more frequently exposed to AMR E. coli than those inhabiting natural environments.
Interesting findings have also emerged from studies on microorganisms other than enteric pathogens, such as Enterococcus spp. Navarro-Gonzalez et al. [114] compared two wild boar populations: one inhabiting Collserola Natural Park and its surroundings (classified as urban wild boars) and another from the National Game Reserve Ports de Tortosa i Beseit, a remote rural area. The authors reported a significantly higher prevalence of E. faecium resistant to tetracycline and showing high-level streptomycin resistance in urban wild boars compared with their rural counterparts. The meaning of these results lies in the fact that both streptomycin and tetracycline are among the most commonly used antibiotics in livestock production, indicating that urban wild boars may acquire resistant bacteria through contact with environments contaminated by agricultural runoff, manure, or other human-related sources. In an earlier study, the same authors [35] also detected, in an urban wild boar from Collserola Natural Park, a linezolid-resistant E. faecalis strain—strongly suggesting an anthropogenic origin, since linezolid is a fully synthetic antibiotic used exclusively in human medicine, and resistance remains extremely rare in isolates from food-producing animals [138,139].
These studies provide a valuable One Health model, illustrating how the interaction between wildlife, human activities, and the environment shapes pathogen circulation and antimicrobial resistance dynamics in synurbized wild boar populations.
7. Conclusions and Future Perspective
The growing evidence on enteric pathogens and antimicrobial resistance (AMR) in wild boars highlights their role as a sentinel species within the One Health framework. Current data reveal considerable variability in prevalence and resistance profiles across Europe, driven by ecological, anthropogenic, and methodological factors. While resistance to critically important antimicrobials remains relatively sporadic, its presence in wild boar isolates underscores the permeability of ecological boundaries between humans, livestock, and wildlife.
Despite the increasing recognition of wild boars as indicators of AMR spread, several knowledge gaps remain. Longitudinal studies are scarce, limiting our understanding of temporal trends and persistence of resistant strains in wildlife populations. Genomic and metagenomic analyses are still underutilized, hindering the identification of transmission pathways between wildlife, domestic animals, and humans. Furthermore, the impact of environmental contamination and the role of synurbic behavior in shaping AMR dynamics remain poorly quantified.
These findings call for the development of harmonized surveillance systems that integrate wildlife monitoring with existing AMR control programs for livestock and humans. Policy frameworks should promote standardized methodologies for sampling, detection, and reporting of AMR in wildlife. Additionally, cross-sectoral collaboration and data sharing are critical to inform risk assessment, management strategies, and public health interventions aimed at minimizing AMR dissemination across ecosystems.
Wild boars, given their expanding distribution and increasing interaction with human environments, represent not only a challenge for public health but also a unique opportunity to fill critical research gaps and guide evidence-based policy actions within a One Health approach.
Supplementary Materials
The following supporting information can be downloaded at https://www.mdpi.com/article/10.3390/antibiotics14121246/s1. Figure S1: Prevalence (%) of Salmonella spp. wild boars carriers at faeces and/or lymph nodes and/or tonsil level in different countries of the European Union.; Figure S2: Prevalence (%) of Y. enterocolitica wild boars carriers at faeces and/or lymph nodes and/or tonsil level in different countries of the European Union.; Figure S3: Prevalence (%) of Campylobacter spp. wild boars carriers at faeces level in different countries of the European Union.; Figure S4: Prevalence (%) of STEC wild boars carriers at faeces level in different countries of the European Union.
Author Contributions
Conceptualization, F.P.; methodology, F.P. and G.S.; validation, F.P., E.P.L.D.S. and C.S.; data curation, F.P. and G.S.; writing—original draft preparation, F.P. and G.S.; writing—review and editing, F.P. and G.S.; supervision, E.P.L.D.S. and C.S. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Massei, G.; Kindberg, J.; Licoppe, A.; Gačić, D.; Šprem, N.; Kamler, J.; Baubet, E.; Hohmann, U.; Monaco, A.; Ozoliņš, J. Wild boar populations up, numbers of hunters down? A review of trends and implications for Europe. Pest Manag. Sci. 2015, 71, 492–500. [Google Scholar] [CrossRef]
- Geisser, H.; Reyer, H.U. The influence of food and temperature on population density of wild boar Sus scrofa in the Thurgau (Switzerland). J. Zool. 2005, 267, 89–96. [Google Scholar] [CrossRef]
- Holland, E.P.; Burrow, J.F.; Dytham, C.; Aegerter, J.N. Modelling with uncertainty: Introducing a probabilistic framework to predict animal population dynamics. Ecol. Model. 2009, 220, 1203–1217. [Google Scholar] [CrossRef]
- Tack, J. Wild Boar (Sus scrofa) Populations in Europe: A Scientific Review of Population Trends and Implications for Management; European Landowners’ Organization: Brussels, Belgium, 2018; p. 56. [Google Scholar]
- Tinoco Torres, R.; Fernandes, J.; Carvalho, J.; Cunha, M.V.; Caetano, T.; Mendo, S.; Serrano, E.; Fonseca, C. Wild boar as a reservoir of antimicrobial resistance. Sci. Total Environ. 2020, 717, 135001. [Google Scholar] [CrossRef] [PubMed]
- Podgórski, T.; Borowik, T.; Lyjak, M.; Wozniakowski, G. Spatial epidemiology of African swine fever: Host, landscape and anthropogenic drivers of disease occurrence in wild boar. Prev. Vet. Med. 2020, 177, 104691. [Google Scholar] [CrossRef] [PubMed]
- Salazar, L.G.; Rose, N.; Hayes, B.; Hammami, P.; Baubet, E.; Desvaux, S.; Andraud, M. Effects of habitat fragmentation and hunting activities on African swine fever dynamics among wild boar populations. Prev. Vet. Med. 2022, 208, 105750. [Google Scholar] [CrossRef]
- Correa Lopes, B.; Roth, M.; Vidaletti, M.; Loiko, M.R.; da Silva Andrade, J.; Gisler Maciel, A.L.; Doyle, R.L.; Cavalheiro Bertagnolli, A.; Oliveira Rodrigues, R.; Driemeier, D.; et al. Investigation of Mycobacterium bovis and Metastrongylus sp. co-infection and its relationship to tuberculosis lesions’ occurrence in wild boars. Comp. Immunol. Microbiol. Infect. Dis. 2021, 77, 101674. [Google Scholar] [CrossRef]
- Martins Ruano, Z.; Mateus, T.L.; Vieira-Pinto, M. An insight into brucellosis in wild boar and domestic pigs in Europe: A systematic review. J. Infect. Public Health 2025, 18, 102691. [Google Scholar] [CrossRef]
- Sauter-Louis, C.; Conraths, F.J.; Probst, C.; Blohm, U.; Schulz, K.; Sehl, J.; Fischer, M.; Forth, J.H.; Zani, L.; Depner, K.; et al. African swine fever in wild boar in Europe—A review. Viruses 2022, 13, 1717. [Google Scholar] [CrossRef]
- Siddi, G.; Piras, F.; Meloni, M.P.; Gymoese, P.; Torpdahl, M.; Fredriksson-Ahomaa, M.; Migoni, M.; Cabras, D.; Cuccu, M.; De Santis, E.P.L.; et al. Hunted wild boars in Sardinia: Prevalence, antimicrobial resistance and genomic analysis of Salmonella and Yersinia enterocolitica. Foods 2024, 13, 65. [Google Scholar] [CrossRef]
- Dias, D.; Costa, S.; Fonseca, C.; Baraúna, R.; Caetano, T.; Mendo, S. Pathogenicity of Shiga toxin-producing Escherichia coli (STEC) from wildlife: Should we care? Sci. Total Environ. 2022, 812, 152324. [Google Scholar] [CrossRef]
- Castillo-Contreras, R.; Marín, M.; López-Olvera, J.R.; Ayats, T.; Fernández-Aguilar, X.; Lavín, S.; Mentaberre, G.; Cerdà-Cuéllar, M. Zoonotic Campylobacter spp. and Salmonella spp. carried by wild boars in a metropolitan area: Occurrence, antimicrobial susceptibility and public health relevance. Sci. Total Environ. 2022, 822, 153444. [Google Scholar] [CrossRef]
- Kerkhof, P.J.; Peruzy, M.F.; Murru, N.; Houf, K. Wild boars as reservoir for Campylobacter and Arcobacter. Vet. Microbiol. 2022, 270, 109462. [Google Scholar] [CrossRef]
- Tayh, G.; Srairi, S.; Selmi, R.; Chehida, F.B.; Mamlouk, A.; Daaloul-Jedidi, M.; Messadi, L. Risk for public health of multiresistant Shiga toxin-producing Escherichia coli (STEC) in wild boar (Sus scrofa) in Tunisia. Microb. Pathog. 2025, 201, 107366. [Google Scholar] [CrossRef]
- Seinige, D.; von Altrock, A.; Kehrenberg, C. Genetic diversity and antibiotic susceptibility of Staphylococcus aureus isolates from wild boars. Comp. Immunol. Microbiol. Infect. Dis. 2017, 54, 7–12. [Google Scholar] [CrossRef]
- Ahmed, S.K.; Hussein, S.; Qurbani, K.; Ibrahim, R.H.; Fareeq, A.; Mahmood, K.A.; Gamal, M.M. Antimicrobial resistance: Impacts, challenges, and future prospects. J. Med. Surg. Public Health 2024, 2, 100081. [Google Scholar] [CrossRef]
- World Organisation for Animal Health (WOAH). Tripartite AMR Project: Working Together to Fight Antimicrobial Resistance. 2023. Available online: https://rr-americas.woah.org/en/projects/ue-ram/ (accessed on 5 November 2025).
- EFSA (European Food Safety Authority); ECDC (European Centre for Disease Prevention and Control). The European Union summary report on antimicrobial resistance in zoonotic and indicator bacteria from humans, animals and food in 2022–2023. EFSA J. 2025, 23, e9237. [Google Scholar] [CrossRef] [PubMed]
- WHO. Critically Important Antimicrobials for Human Medicine: 6th Revision; World Health Organization: Geneva, Switzerland, 2019. [Google Scholar]
- Doyle, C.; Wall, K.; Fanning, S.; McMahon, B.J. Making sense of sentinels: Wildlife as the One Health bridge for environmental antimicrobial resistance surveillance. J. Appl. Microbiol. 2025, 136, lxaf017. [Google Scholar] [CrossRef] [PubMed]
- White, A.; Hughes, J.M. Critical importance of a One Health approach to antimicrobial resistance. EcoHealth 2019, 16, 404–409. [Google Scholar] [CrossRef]
- Baquero, F.; Coque, T.M.; Martinez, J.L.; Aracil-Gisbert, S.; Lanza, V.S. Gene transmission in the One Health microbiosphere and the channels of antimicrobial resistance. Front. Microbiol. 2019, 10, 2892. [Google Scholar] [CrossRef]
- Robinson, T.P.; Bu, D.P.; Carrique-Mas, J.; Fèvre, E.M.; Gilbert, M.; Grace, D.; Hay, S.I.; Jiwakanon, J.; Kakkar, M.; Kariuki, S.; et al. Antibiotic resistance is the quintessential One Health issue. Trans. R. Soc. Trop. Med. Hyg. 2016, 110, 377–380. [Google Scholar] [CrossRef]
- Vezeau, N.; Kahn, L. Current understanding and knowledge gaps regarding wildlife as reservoirs of antimicrobial resistance. Am. J. Vet. Res. 2024, 85, 1–9. [Google Scholar] [CrossRef]
- Bonnedahl, J.; Järhult, J.D. Antibiotic resistance in wild birds. Upsala J. Med. Sci. 2014, 119, 113–116. [Google Scholar] [CrossRef] [PubMed]
- Vittecoq, M.; Godreuil, S.; Prugnolle, F.; Durand, P.; Brazier, L.; Renaud, N.; Arnal, A.; Aberkane, S.; Jean-Pierre, H.; Gauthier-Clerc, M.; et al. Antimicrobial resistance in wildlife: An emerging issue. Trends Microbiol. 2016, 24, 448–459. [Google Scholar] [CrossRef]
- Unger, F.; Eisenberg, T.; Prenger-Berninghoff, E.; Leidner, U.; Semmler, T.; Ewers, C. Phenotypic and genomic characterization of ESBL- and AmpC-β-lactamase-producing Enterobacterales isolates from imported healthy reptiles. Antibiotics 2024, 13, 1230. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, R.; Abreu, R.; Serrano, I.; Such, R.; Garcia-Vila, E.; Quirós, S.; Cunha, E.; Tavares, L.; Oliveira, M. Resistant Escherichia coli isolated from wild mammals from two rescue and rehabilitation centers in Costa Rica: Characterization and public health relevance. Sci. Rep. 2024, 14, 8039. [Google Scholar] [CrossRef]
- EFSA (European Food Safety Authority); ECDC (European Centre for Disease Prevention and Control). The European Union One Health 2023 Zoonoses Report. EFSA J. 2024, 22, e9106. [Google Scholar] [CrossRef]
- Centers for Disease Control and Prevention (CDC). Salmonella and Food. U.S. Department of Health and Human Services, 2023. Available online: https://www.cdc.gov/salmonella/ (accessed on 5 November 2025).
- Paulsen, P.; Smulders, F.J.M.; Hilbert, F. Salmonella in meat from hunted game: A central European perspective. Food Res. Int. 2012, 45, 609–616. [Google Scholar] [CrossRef]
- Wacheck, S.; Fredriksson-Ahomaa, M.; König, M.; Stolle, A.; Stephan, R. Wild Boars as an important reservoir for foodborne pathogens. Foodborne Pathog. Dis. 2010, 7, 307–312. [Google Scholar] [CrossRef]
- Vieira-Pinto, M.; Morais, L.; Caleja, C.; Themudo, P.; Torres, C.; Igrejas, G.; Poeta, P.; Martins, C. Salmonella sp. in game (Sus scrofa and Oryctolagus cuniculus). Foodborne Pathog. Dis. 2011, 8, 739–740. [Google Scholar] [CrossRef]
- Navarro-Gonzalez, N.; Casas-Díaz, E.; Porrero, C.M.; Mateos, A.; Domínguez, L.; Lavín, S.; Serrano, E. Food-Borne zoonotic pathogens and antimicrobial resistance of indicator bacteria in urban wild boars in Barcelona, Spain. Vet. Microbiol. 2013, 167, 686–689. [Google Scholar] [CrossRef]
- Chiari, M.; Zanoni, M.; Tagliabue, S.; Lavazza, A.; Alborali, L.G. Salmonella serotypes in wild boars (Sus scrofa) hunted in northern Italy. Acta Vet. Scand. 2013, 55, 42. [Google Scholar] [CrossRef]
- Díaz-Sánchez, S.; Sánchez, S.; Herrera-León, S.; Porrero, C.; Blanco, J.; Dahbi, G.; Blanco, J.E.; Mora, A.; Mateo, R.; Hanning, I.; et al. Prevalence of shiga toxin-producing Escherichia coli, Salmonella spp. and Campylobacter spp. in large game animals intended for consumption: Relationship with management practices and livestock influence. Vet. Microbiol. 2013, 163, 274–281. [Google Scholar] [CrossRef]
- Zottola, T.; Montagnaro, S.; Magnapera, C.; Sasso, S.; De Martino, L.; Bragagnolo, A.; D’Amici, L.; Condoleo, R.; Pisanelli, G.; Iovane, G.; et al. Prevalence and antimicrobial susceptibility of Salmonella in European wild boar (Sus scrofa): Latium region, Italy. Comp. Immunol. Microbiol. Infect. Dis. 2013, 36, 161–168. [Google Scholar] [CrossRef]
- Dias, D.; Torres, R.T.; Kronvall, G.; Fonseca, C.; Mendo, S.; Caetano, T. Assessment of antibiotic resistance of Escherichia coli isolates and screening of Salmonella spp. in wild ungulates from Portugal. Res. Microbiol. 2015, 166, 584–593. [Google Scholar] [CrossRef]
- Sannö, A.; Rosendal, T.; Aspán, A.; Backhans, A.; Jacobson, M. Distribution of enteropathogenic Yersinia spp. and Salmonella spp. in the Swedish wild boar population, and assessment of risk factors that may affect their prevalence. Acta Vet. Scand. 2018, 60, 40. [Google Scholar] [CrossRef]
- Stella, S.; Tirloni, E.; Castelli, E.; Colombo, F.; Bernardi, C. Microbiological evaluation of carcasses of wild boar hunted in a hill area of northern Italy. J. Food Prot. 2018, 81, 1519–1525. [Google Scholar] [CrossRef]
- Bonardi, S.; Bolzoni, L.; Zanoni, R.G.; Morganti, M.; Corradi, M.; Gilioli, S.; Pongolini, S. Limited exchange of Salmonella among domestic pigs and wild boars in Italy. EcoHealth 2019, 16, 420–428. [Google Scholar] [CrossRef]
- Gil Molino, M.; García Sánchez, A.; Risco Pérez, D.; Gonçalves Blanco, P.; Quesada Molina, A.; Rey Pérez, J.; Martín Cano, F.E.; Cerrato Horrillo, R.; Hermoso-de-Mendoza Salcedo, J.; Fernández Llario, P. Prevalence of Salmonella spp. in tonsils, mandibular lymph nodes and faeces of wild boar from Spain and genetic relationship between isolates. Transbound. Emerg. Dis. 2019, 66, 1218–1226. [Google Scholar] [CrossRef]
- Petersen, H.H.; Takeuchi-Storm, N.; Enemark, H.L.; Nielsen, S.T.; Larsen, G.; Chriél, M. Surveillance of important bacterial and parasitic infections in Danish wild boars (Sus scrofa). Acta Vet. Scand. 2020, 62, 41. [Google Scholar] [CrossRef]
- Cilia, G.; Turchi, B.; Fratini, F.; Bilei, S.; Bossù, T.; De Marchis, M.L.; Cerri, D.; Pacini, M.I.; Bertelloni, F. Prevalence, virulence and antimicrobial susceptibility of Salmonella spp., Yersinia enterocolitica and Listeria monocytogenes in European wild boar (Sus. scrofa) hunted in Tuscany (central Italy). Pathogens 2021, 10, 93. [Google Scholar] [CrossRef]
- Piras, F.; Spanu, V.; Siddi, G.; Gymoese, P.; Spanu, C.; Cibin, V.; Schjørring, S.; De Santis, E.P.L.; Scarano, C. Whole-genome sequencing analysis of highly prevalent Salmonella serovars in wild boars from a national park in Sardinia. Food Control 2021, 130, 108247. [Google Scholar] [CrossRef]
- Plaza-Rodríguez, C.; Alt, K.; Grobbel, M.; Hammerl, J.A.; Irrgang, A.; Szabo, I.; Stingl, K.; Schuh, E.; Wiehle, L.; Pfefferkorn, B.; et al. Wildlife as sentinels of antimicrobial resistance in Germany? Front. Vet. Sci. 2021, 7, 627821. [Google Scholar] [CrossRef] [PubMed]
- Bolzoni, L.; Bonardi, S.; Tansini, C.; Scaltriti, E.; Menozzi, I.; Morganti, M.; Conter, M.; Pongolini, S. Different roles of wild boars and livestock in Salmonella transmission to humans in Italy. EcoHealth 2023, 20, 122–132. [Google Scholar] [CrossRef]
- Altissimi, C.; Primavilla, S.; Roila, R.; Gavaudan, S.; Morandi, B.; Di Lullo, S.; Coppini, M.; Baldinelli, C.; Cai, D.; Branciari, R.; et al. Salmonella in wild boar meat: Prevalence and risk assessment in central Italy (Umbria and Marche region). Foods 2024, 13, 1156. [Google Scholar] [CrossRef] [PubMed]
- Petrović, J.; Mirčeta, J.; Velhner, M.; Stojanov, I.; Ratajac, R.; Prodanov-Radulović, J. Salmonella in wild boars (Sus scrofa): Influence of hunting and dressing procedures on meat safety. Arch. Vet. Med. 2024, 17, 51–67. [Google Scholar] [CrossRef]
- Beloeil, P.A.; Chauvin, C.; Proux, K.; Rose, N.; Queguiner, S.; Eveno, E.; Houdayer, C.; Rose, V.; Fravalo, P.; Madec, F. Longitudinal serological responses to Salmonella enterica of growing pigs in a subclinically infected herd. Prev. Vet. Med. 2003, 60, 207–226. [Google Scholar] [CrossRef]
- Ivanek, R.; Österberg, J.; Gautam, R.; Sternberg Lewerin, S. Salmonella fecal shedding and immune responses are dose- and serotype-dependent in pigs. PLoS ONE 2012, 7, e34660. [Google Scholar] [CrossRef]
- Berends, B.R.; van Knapen, F.; Snijders, J.M.A.; Mossel, D.A.A. Identification and quantification of risk factors in animal management and transport regarding Salmonella spp. in pigs. Int. J. Food Microbiol. 1996, 44, 207–217. [Google Scholar] [CrossRef]
- Kranker, S.; Dahl, J.; Wingstrand, A. Bacteriological and serological examination and risk factor analysis of Salmonella occurrence in sow herds, including risk factors for high Salmonella seroprevalence in receiver finishing herds. Berl. Munch. Tierarztl. Wochenschr. 2001, 114, 350–352. [Google Scholar]
- EFSA BIOHAZ Panel. Scientific opinion on a quantitative microbiological risk assessment of Salmonella in slaughter and breeder pigs. EFSA J. 2010, 8, 1547. [Google Scholar] [CrossRef]
- Ainslie-Garcia, M.H.; Farzan, A.; Newman, J.E.; Friendship, R.M.; Lillie, B.N. Salmonella fecal shedding in pigs from birth to market and its association with the presence of Salmonella in palatine tonsils and submandibular lymph nodes at slaughter. Can. J. Vet. Res. 2018, 82, 249–255. [Google Scholar] [PubMed]
- Soliani, L.; Rugna, G.; Prosperi, A.; Chiapponi, C.; Luppi, A. Salmonella infection in pigs: Disease, prevalence, and a link between swine and human health. Pathogens 2023, 12, 1267. [Google Scholar] [CrossRef] [PubMed]
- Ward, M.P.; Cowled, B.; Galea, F.; Garner, M.G.; Laffan, S.W.; Marsh, I.; Negus, K.; Sarre, S.D.; Woolnough, A.P. Salmonella infection in a remote, isolated wild pig population. Vet. Microbiol. 2013, 162, 921–929. [Google Scholar] [CrossRef]
- Gaffuri, A.; Holmes, J.P. Salmonella infections. In Infectious Diseases of Wild Mammals and Birds in Europe; Gavier-Widén, D., Duff, J.P., Meredith, A., Eds.; Wiley-Blackwell: Oxford, UK, 2012; pp. 398–407. [Google Scholar] [CrossRef]
- Navarro-Gonzalez, N.; Mentaberre, G.; Porrero, C.M.; Serrano, E.; Mateos, A.; López-Martín, J.M.; Lavín, S.; Domínguez, L. Effect of cattle on Salmonella carriage, diversity and antimicrobial resistance in free-ranging wild boar (Sus scrofa) in northeastern Spain. PLoS ONE 2012, 7, e51614. [Google Scholar] [CrossRef]
- Flores Rodas, E.M.; Bogdanova, T.; Bossù, T.; Pecchi, S.; Tomassetti, F.; De Santis, P.; Tolli, R.; Condoleo, R.; Greco, S.; Brozzi, A.; et al. Microbiological assessment of freshly-shot wild boars meat in Lazio Region, Viterbo Territory: A preliminary study. Ital. J. Food Saf. 2014, 3, 1711. [Google Scholar] [CrossRef]
- Razzuoli, E.; Listorti, V.; Martini, I.; Migone, L.; Decastelli, L.; Mignone, W.; Berio, E.; Battistini, R.; Ercolini, C.; Serracca, L.; et al. Prevalence and antimicrobial resistances of Salmonella spp. isolated from wild boars in Liguria Region, Italy. Pathogens 2021, 10, 568. [Google Scholar] [CrossRef]
- Mirceta, J.; Petrovic, J.; Malesevic, M.; Blagojevic, B.; Antic, D. Assessment of microbial carcass contamination of hunted wild boars. Eur. J. Wildl. Res. 2017, 63, 37–44. [Google Scholar] [CrossRef]
- Fredriksson-Ahomaa, M.; London, L.; Skrzypczak, T.; Kantala, T.; Laamanen, I.; Biström, M.; Maunula, L.; Gadd, T. Foodborne zoonoses common in hunted wild boars. EcoHealth 2020, 17, 512–522. [Google Scholar] [CrossRef]
- Ranucci, D.; Roila, R.; Onofri, A.; Cambiotti, F.; Primavilla, S.; Miraglia, D.; Andoni, E.; Di Cerbo, A.; Branciari, R. Improving hunted wild boar carcass hygiene: Roles of different factors involved in the harvest phase. Foods 2021, 10, 1548. [Google Scholar] [CrossRef]
- Peruzy, M.F.; Murru, N.; Smaldone, G.; Proroga, Y.T.R.; Cristiano, D.; Fioretti, A.; Anastasio, A. Hygiene evaluation and microbiological hazards of hunted wild boar carcasses. Food Control 2022, 135, 108782. [Google Scholar] [CrossRef]
- Bonardi, S.; Tansini, C.; Cacchioli, A.; Soliani, L.; Poli, L.; Lamperti, L.; Gilioli, S. Enterobacteriaceae and Salmonella contamination of wild boar (Sus scrofa) carcasses: Comparison between different sampling strategies. Eur. J. Wildl. Res. 2021, 67, 88. [Google Scholar] [CrossRef] [PubMed]
- Tennant, S.M.; Grant, T.H.; Robins-Browne, R.M. Pathogenicity of Yersinia enterocolitica biotype 1A. FEMS Microbiol. Lett. 2003, 38, 127–137. [Google Scholar] [CrossRef]
- Fàbrega, A.; Vila, J. Yersinia enterocolitica: Pathogenesis, virulence and antimicrobial resistance. Enferm. Infecc. Microbiol. Clin. 2012, 30, 24–32. [Google Scholar] [CrossRef]
- Von Altrock, A.; Seinige, D.; Kehrenberg, C. Yersinia enterocolitica isolates from wild boars hunted in Lower Saxony, Germany. Appl. Environ. Microbiol. 2015, 81, 4835–4840. [Google Scholar] [CrossRef]
- Fredriksson-Ahomaa, M.; Gerhardt, M.; Stolle, A. High bacterial contamination of pig tonsils at slaughter. Meat Sci. 2009, 83, 334–336. [Google Scholar] [CrossRef]
- Fois, F.; Piras, F.; Torpdahl, M.; Mazza, R.; Ladu, D.; Consolati, S.G.; Spanu, C.; Scarano, C.; De Santis, E.P.L. Prevalence, bioserotyping and antibiotic resistance of pathogenic Yersinia enterocolitica detected in pigs at slaughter in Sardinia. Int. J. Food Microbiol. 2018, 283, 1–6. [Google Scholar] [CrossRef]
- Arrausi-Subiza, M.; Ibabe, J.C.; Atxaerandio, R.; Juste, R.A.; Barral, M. Evaluation of different enrichment methods for pathogenic Yersinia species detection by real-time PCR. BMC Vet. Res. 2014, 10, 192. [Google Scholar] [CrossRef]
- Syczyło, K.; Platt-Samoraj, A.; Bancerz-Kisiel, A.; Szczerba-Turek, A.; Pajdak-Czaus, J.; Łabuć, S.; Szweda, W. The prevalence of Yersinia enterocolitica in game animals in Poland. PLoS ONE 2018, 13, e0195136. [Google Scholar] [CrossRef]
- Bancerz-Kisiel, A.; Platt-Samoraj, A.; Szczerba-Turek, A.; Syczyło, K.; Szweda, W. The first pathogenic Yersinia enterocolitica bioserotype 4/O:3 strain isolated from a hunted wild boar (Sus scrofa) in Poland. Epidemiol. Infect. 2015, 143, 2758–2765. [Google Scholar] [CrossRef]
- Carella, E.; Romano, A.; Domenis, L.; Robetto, S.; Spedicato, R.; Guidetti, C.; Orusa, R. Characterisation of Yersinia enterocolitica strains isolated from wildlife in the northwestern Italian Alps. J. Vet. Res. 2022, 66, 141. [Google Scholar] [CrossRef]
- Delibato, E.; Ventola, E.; Lovari, S.; Farneti, S.; Finazzi, G.; Owczarek, S.; Stefano, B. Molecular characterization of Yersinia enterocolitica strains to evaluate virulence-associated genes. Ann. Ist. Super. Sanità 2023, 59, 280–285. [Google Scholar] [CrossRef]
- Arrausi-Subiza, M.; Gerrikagoitia, X.; Alvarez, V.; Ibabe, J.C.; Barral, M. Prevalence of Yersinia enterocolitica and Yersinia pseudotuberculosis in wild boars in the Basque Country, northern Spain. Acta Vet. Scand. 2016, 58, 4. [Google Scholar] [CrossRef] [PubMed]
- Bonardi, S.; Brémont, S.; Vismarra, A.; Poli, I.; Diegoli, G.; Bolzoni, L.; Corradi, M.; Gilioli, S.; Le Guern, A.S. Is Yersinia bercovieri surpassing Yersinia enterocolitica in wild boars (Sus scrofa)? EcoHealth 2020, 17, 388–392. [Google Scholar] [CrossRef]
- Slee, K.J.; Skilbeck, N.W. Epidemiology of Yersinia pseudotuberculosis and Y. enterocolitica infections in sheep in Australia. J. Clin. Microbiol. 1992, 30, 712–715. [Google Scholar] [CrossRef]
- Nikolaou, K.; Hensel, A.; Bartling, C.; Tomaso, H.; Arnold, T.; Rösler, U.; Ganter, M.; Petry, T.; Neubauer, H. Prevalence of anti-Yersinia outer protein antibodies in goats in Lower Saxony. J. Vet. Med. B Infect. Dis. Vet. Public Health 2005, 52, 17–24. [Google Scholar] [CrossRef]
- Piras, F.; Spanu, C.; Sanna, R.; Siddi, G.; Mocci, A.M.; Demontis, M.; Meloni, M.P.; Spanu, V.; De Santis, E.P.L.; Scarano, C. Detection, virulence genes and antimicrobial resistance of Yersinia enterocolitica in sheep and goat raw milk. Int. Dairy J. 2021, 117, 105011. [Google Scholar] [CrossRef]
- Avagnina, A.; Nucera, D.; Grassi, M.A.; Ferroglio, E.; Dalmasso, A.; Civera, T. The microbiological conditions of carcasses from large game animals in Italy. Meat Sci. 2012, 91, 266–271. [Google Scholar] [CrossRef]
- Lorencová, A.; Babák, V.; Lamká, J. Serological prevalence of enteropathogenic Yersinia spp. in pigs and wild boars from different production systems in the Moravian region, Czech Republic. Foodborne Pathog. Dis. 2016, 13, 275–279. [Google Scholar] [CrossRef]
- Modesto, P.; De Ciucis, C.G.; Vencia, W.; Pugliano, M.C.; Mignone, W.; Berio, E.; Masotti, C.; Ercolini, C.; Serracca, L.; Andreoli, T.; et al. Evidence of antimicrobial resistance and presence of pathogenicity genes in Yersinia enterocolitica isolate from wild boars. Pathogens 2021, 10, 398. [Google Scholar] [CrossRef] [PubMed]
- Russo, C.; Balloni, S.; Altomonte, I.; Martini, M.; Nuvoloni, R.; Cecchi, F.; Pedonese, F.; Salari, F.; Santana Da Silva, A.M.; Torracca, B.; et al. Fatty acid and microbiological profile of the meat (Longissimus dorsi muscle) of wild boar (Sus scrofa scrofa) hunted in Tuscany. Ital. J. Anim. Sci. 2017, 16, 1–8. [Google Scholar] [CrossRef]
- Peruzy, M.F.; Cristiano, D.; Delibato, E.; D’Alessio, N.; Proroga, Y.T.R.; Capozza, R.L.; Rippa, A.; Murru, N. Presence of enteric bacterial pathogens in meat samples of wild boar hunted in Campania Region, Southern Italy. Ital. J. Food Saf. 2022, 11, 9967. [Google Scholar] [CrossRef]
- Li, Z.; Cai, H.; Xu, B.; Dong, Q.; Jia, K.; Lin, Z.; Wang, X.; Liu, Y.; Qin, X. Prevalence, antibiotic resistance, resistance and virulence determinants of Campylobacter jejuni in China: A systematic review and meta-analysis. One Health 2025, 20, 100990. [Google Scholar] [CrossRef]
- Carbonero, A.; Paniagua, J.; Torralbo, A.; Arenas-Montes, A.; Borge, C.; García-Bocanegra, I. Campylobacter infection in wild artiodactyl species from southern Spain: Occurrence, risk factors and antimicrobial susceptibility. Comp. Immunol. Microbiol. Infect. Dis. 2014, 37, 115–121. [Google Scholar] [CrossRef]
- Ziomek, M.; Gondek, M.; Torracca, B.; Marotta, F.; Garofolo, G.; Wieczorek, K.; Michalak, K.; Fratini, F.; Pedonese, F. Occurrence of Campylobacter in faeces, livers and carcasses of wild boars hunted in Tuscany (Italy) and evaluation of MALDI-TOF MS for the identification of Campylobacter species. Foods 2023, 12, 778. [Google Scholar] [CrossRef]
- Abe, T.; Haga, S.; Yokoyama, K.; Watanabe, N. An outbreak of Campylobacter jejuni subsp. jejuni infection via tap water. Jpn. J. Infect. Dis. 2008, 61, 327. [Google Scholar] [CrossRef] [PubMed]
- Richardson, G.; Thomas, D.R.; Smith, R.M.; Nehaul, L.; Ribeiro, C.D.; Brown, A.G.; Salmon, R.L. A community outbreak of Campylobacter jejuni infection from a chlorinated public water supply. Epidemiol. Infect. 2007, 135, 1151–1158. [Google Scholar] [CrossRef]
- Gonzalez, M.; Hanninen, M.L. Effect of temperature and antimicrobial resistance on survival of Campylobacter jejuni in well water: Application of the Weibull model. J. Appl. Microbiol. 2012, 113, 284–293. [Google Scholar] [CrossRef]
- Atanassova, V.; Apelt, J.; Reich, F.; Klein, G. Microbiological quality of freshly shot game in Germany. Meat Sci. 2008, 78, 414–419. [Google Scholar] [CrossRef]
- Marotta, F.; Di Marcantonio, L.; Janowicz, A.; Pedonese, F.; Di Donato, G.; Ardelean, A.; Nuvoloni, R.; Di Giannatale, E.; Garofolo, G. Genotyping and antibiotic resistance traits in Campylobacter jejuni and coli from pigs and wild boars in Italy. Front. Cell. Infect. Microbiol. 2020, 10, 592512. [Google Scholar] [CrossRef] [PubMed]
- Karmali, M.A. Infection by verocytotoxin-producing Escherichia coli. Clin. Microbiol. Rev. 1989, 2, 15–38. [Google Scholar] [CrossRef]
- Koutsoumanis, K.; Allende, A.; Alvarez-Ordóñez, A.; Bover-Cid, S.; Chemaly, M.; Davies, R.; De Cesare, A.; Herman, L.; Hilbert, F.; Lindqvist, R.; et al. Pathogenicity assessment of Shiga toxin-producing Escherichia coli (STEC) and the public health risk posed by contamination of food with STEC. EFSA J. 2020, 18, e05967. [Google Scholar] [CrossRef]
- Frank, E.; Bonke, R.; Drees, N.; Heurich, M.; Märtlbauer, E.; Gareis, M. Shiga toxin-producing Escherichia coli (STEC) shedding in a wild roe deer population. Vet. Microbiol. 2019, 239, 108479. [Google Scholar] [CrossRef] [PubMed]
- Amézquita-López, B.A.; Soto-Beltrán, M.; Lee, B.G.; Yambao, J.C.; Quiñones, B. Isolation, genotyping and antimicrobial resistance of Shiga toxin-producing Escherichia coli. J. Microbiol. Immunol. Infect. 2018, 51, 425–434. [Google Scholar] [CrossRef] [PubMed]
- Mora, A.; López, C.; Dhabi, G.; López-Beceiro, A.M.; Fidalgo, L.E.; Díaz, E.A.; Martinez-carrasco, C.; Mamani, R.; Herrera, A.; Blanco, J.E.; et al. Seropathotypes, phylogroups, Stx subtypes, and intimin types of wildlife-carried, Shiga toxin-producing Escherichia coli strains with the same characteristics as human-pathogenic isolates. Appl. Environ. Microbiol. 2012, 78, 2578–2585. [Google Scholar] [CrossRef] [PubMed]
- Alonso, C.A.; Mora, A.; Díaz, D.; Blanco, M.; González-Barrio, D.; Ruiz-Fons, F.; Simon, C.; Blanco, J.; Torres, C. Occurrence and characterization of stx and/or eae-positive Escherichia coli isolated from wildlife, including a typical EPEC strain from a wild boar. Vet. Microbiol. 2017, 207, 69–73. [Google Scholar] [CrossRef]
- Szczerba-Turek, A.; Socha, P.; Bancerz-Kisiel, A.; Platt-Samoraj, A.; Lipczynska-Ilczuk, K.; Siemionek, J.; Kończyk, K.; Terech-Majewska, E.; Szweda, W. Pathogenic potential to humans of Shiga toxin-producing Escherichia coli isolated from wild boars in Poland. Int. J. Food Microbiol. 2019, 300, 8–13. [Google Scholar] [CrossRef]
- Dias, D.; Caetano, T.; Torres, R.T.; Fonseca, C.; Mendo, S. Shiga toxin-producing Escherichia coli in wild ungulates. Sci. Total Environ. 2019, 651, 203–209. [Google Scholar] [CrossRef]
- Nüesch-Inderbinen, M.; Barmettler, K.; Stevens, M.J.A.; Cernela, N. Shiga toxin-producing Escherichia coli isolated from hunted wild boar (Sus scrofa) in Switzerland. Schweiz. Arch. Tierheilkd 2024, 166, 131–140. [Google Scholar]
- Sannö, A.; Aspán, A.; Hestvik, G.; Jacobson, M. Presence of Salmonella spp., Yersinia enterocolitica, Yersinia pseudotuberculosis and Escherichia coli O157:H7 in wild boars. Epidemiol. Infect. 2014, 142, 2542–2547. [Google Scholar] [CrossRef]
- Navarro-Gonzalez, N.; Porrero, M.C.; Mentaberre, G.; Serrano, E.; Mateos, A.; Cabal, A.; Domínguez, L.; Lavín, S. Escherichia coli O157:H7 in wild boars (Sus scrofa) and Iberian ibex (Capra pyrenaica) sharing pastures with free-ranging livestock in a natural environment in Spain. Vet. Q. 2015, 35, 102–106. [Google Scholar] [CrossRef]
- Bertelloni, F.; Cilia, G.; Bogi, S.; Ebani, V.V.; Turini, L.; Nuvoloni, R.; Cerri, D.; Fratini, F.; Turchi, B. Pathotypes and antimicrobial susceptibility of Escherichia coli isolated from wild boar (Sus. scrofa) in Tuscany. Animals 2020, 10, 744. [Google Scholar] [CrossRef]
- Siddi, G.; Piras, F.; Gymoese, P.; Torpdahl, M.; Meloni, M.P.; Cuccu, M.; Migoni, M.; Cabras, D.; Fredriksson-Ahomaa, M.; De Santis, E.P.; et al. Pathogenic profile and antimicrobial resistance of Escherichia coli, Escherichia marmotae and Escherichia ruysiae detected from hunted wild boars in Sardinia (Italy). Int. J. Food Microbiol. 2024, 421, 110790. [Google Scholar] [CrossRef]
- Díaz-Sánchez, S.; Sánchez, S.; Sánchez, M.; Herrera-León, S.; Hanning, I.; Vidal, D. Detection and characterization of Shiga toxin-producing Escherichia coli in game meat and ready-to-eat meat products. Int. J. Food Microbiol. 2012, 160, 179–182. [Google Scholar] [CrossRef]
- Nüesch-Inderbinen, M.; Treier, A.; Stevens, M.J.; Stephan, R. Whole genome sequence-based characterisation of Shiga toxin-producing Escherichia coli isolated from game meat originating from several European countries. Sci. Rep. 2023, 13, 3247. [Google Scholar] [CrossRef] [PubMed]
- Gill, C.O. Microbiological conditions of meats from large game animals and birds. Meat Sci. 2007, 77, 149–160. [Google Scholar] [CrossRef] [PubMed]
- Vezeau, N.; Kahn, L.H. Spread and mitigation of antimicrobial resistance at the wildlife-urban and wildlife-livestock interfaces. J. Am. Vet. Med. Assoc. 2024, 262, 741–747. [Google Scholar] [CrossRef] [PubMed]
- Navarro-Gonzalez, N.; Castillo-Contreras, R.; Casas-Díaz, E.; Morellet, N.; Porrero, M.C.; Molina-Vacas, G.; Torres, R.T.; Fonseca, C.; Mentaberre, G.; Domínguez, L.; et al. Carriage of antibiotic-resistant bacteria in urban versus rural wild boars. Eur. J. Wildl. Res. 2018, 64, 1–9. [Google Scholar] [CrossRef]
- Dolejská, M.; Literák, I. Wildlife is overlooked in the epidemiology of medically important antibiotic-resistant bacteria. Antimicrob. Agents Chemother. 2019, 63, e01167-19. [Google Scholar] [CrossRef]
- Altissimi, C.; Noé-Nordberg, C.; Ranucci, D.; Paulsen, P. Presence of foodborne bacteria in wild boar and wild boar meat—A literature survey for the period 2012–2022. Foods 2023, 12, 1689. [Google Scholar] [CrossRef]
- Weber, R.E.; Fleige, C.; Layer, F.; Neumann, B.; Kresken, M.; Werner, G. Determination of a tentative epidemiological cut-off value (ECOFF) for dalbavancin and Enterococcus faecium. Antibiotics 2021, 10, 915. [Google Scholar] [CrossRef]
- Akwongo, C.J.; Borrelli, L.; Houf, K.; Fioretti, A.; Peruzy, M.F.; Murru, N. Antimicrobial resistance in wild game mammals: A glimpse into the contamination of wild habitats in a systematic review and meta-analysis. BMC Vet. Res. 2025, 21, 14. [Google Scholar] [CrossRef]
- Magiorakos, A.P.; Srinivasan, A.; Carey, R.B.; Carmeli, Y.; Falagas, M.E.; Giske, C.G.; Harbarth, S.; Hindler, J.F.; Kahlmeter, G.; Olsson-Liljequist, B.; et al. Multidrug-resistant, extensively drug-resistant and pandrug-resistant bacteria: An international expert proposal for interim standard definitions for acquired resistance. Clin. Microbiol. Infect. 2012, 18, 268–281. [Google Scholar] [CrossRef]
- European Centre for Disease Prevention and Control (ECDC); European Food Safety Authority (EFSA); European Medicines Agency (EMA). ECDC/EFSA/EMA second joint report on the integrated analysis of antimicrobial consumption (AMC) and antimicrobial resistance (AMR) in bacteria from humans and food-producing animals. EFSA J. 2017, 15, 4872. [Google Scholar] [CrossRef]
- Gil-Molino, M.; Gonçalves, P.; Risco, D.; Martín-Cano, F.E.; García, A.; Rey, J.; Quesada, A. Dissemination of antimicrobial-resistant isolates of Salmonella spp. in wild boars and its relationship with management practices. Transbound. Emerg. Dis. 2022, 69, e1488–e1502. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.-Y.; Wang, Y.; Walsh, T.R.; Yi, L.-X.; Zhang, R.; Spencer, J.; Shen, J. Emergence of plasmid-mediated colistin resistance mechanism MCR-1 in animals and human beings in China: A microbiological and molecular biological study. Lancet Infect. Dis. 2016, 16, 161–168. [Google Scholar] [CrossRef] [PubMed]
- Falagas, M.E.; Kastoris, A.C.; Kapaskelis, A.M.; Karageorgopoulos, D.E. Fosfomycin for the treatment of multidrug-resistant, including extended-spectrum beta-lactamase producing, Enterobacteriaceae infections: A systematic review. Lancet Infect. Dis. 2010, 10, 43–50. [Google Scholar] [CrossRef]
- CLSI (Clinical and Laboratory Standards Institute). M100 Performance Standards for Antimicrobial Susceptibility Testing A CLSI Supplement for Global Application, 28th ed.; CLSI supplementM100; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2018. [Google Scholar]
- EUCAST. Breakpoint Tables for Interpretation of MICs and Zone Diameters 2020. Version 10.0. 2020. Available online: http://www.eucast.org/fileadmin/src/media/PDFs/EUCAST_files/Breakpoint_tables/v_10.0_Breakpoint_Tables.pdf (accessed on 10 November 2025).
- Fredriksson-Ahomaa, M.; Wacheck, S.; Bonke, R.; Stephan, R. Different enteropathogenic Yersinia strains found in wild boars and domestic pigs. Foodborne Pathog. Dis. 2011, 8, 733–737. [Google Scholar] [CrossRef] [PubMed]
- Begum, J.; Mir, N.A.; Dev, K.; Khan, I.A. Dynamics of antibiotic resistance with special reference to Shiga toxin-producing Escherichia coli infections. J. Appl. Microbiol. 2018, 125, 1228–1237. [Google Scholar] [CrossRef]
- Mühlen, S.; Dersch, P. Treatment strategies for infections with Shiga toxin-producing Escherichia coli. Front. Cell. Infect. Microbiol. 2020, 10, 169. [Google Scholar] [CrossRef]
- European Food Safety Authority (EFSA); European Centre for Disease Prevention and Control (ECDC). The European Union summary report on antimicrobial resistance in zoonotic and indicator bacteria from humans, animals and food in 2017/2018. EFSA J. 2020, 18, 6007. [Google Scholar] [CrossRef]
- Benavides, J.A.; Salgado-Caxito, M.; Torres, C.; Godreuil, S. Public health implications of antimicrobial resistance in wildlife at the One Health interface. Med. Sci. Forum 2024, 25, 1. [Google Scholar] [CrossRef]
- Luniak, M. Synurbization: Adaptation of animal wildlife to urban development. In Proceedings of the 4th International Symposium on Urban Wildlife Conservation, Tucson, AZ, USA, 1–5 May 1999; Shaw, W.S., Harris, L.K., VanDruff, L., Eds.; University of Arizona: Tucson, AZ, USA, 2004; pp. 50–55. [Google Scholar]
- Ruiz-Ponsell, L.; Monastiri, A.; Lopez-Roig, M.; Sauleda, S.; Bes, M.; Mentaberre, G.; Escobar-González, M.; Costafreda, M.I.; López-Olvera, J.R.; Serra-Cobo, J. Endemic maintenance of human-related hepatitis E virus strains in synurbic wild boars, Barcelona Metropolitan Area, Spain. Sci. Total Environ. 2024, 955, 176871. [Google Scholar] [CrossRef]
- Conejero, C.; González-Crespo, C.; Fatjó, J.; Castillo-Contreras, R.; Serrano, E.; Lavín, S.; Mentaberre, G.; López-Olvera, J.R. Between conflict and reciprocal habituation: Human–wild boar coexistence in urban areas. Sci. Total Environ. 2024, 936, 173258. [Google Scholar] [CrossRef]
- Castillo-Contreras, R.; Carvalho, J.; Serrano, E.; Mentaberre, G.; Fernández-Aguilar, X.; Colom, A.; González-Crespo, C.; Lavín, S.; López-Olvera, J.R. Urban wild boars prefer fragmented areas with food resources near natural corridors. Sci. Total Environ. 2018, 615, 282–288. [Google Scholar] [CrossRef]
- Castillo-Contreras, R.; Mentaberre, G.; Fernández-Aguilar, X.; Conejero, C.; Colom-Cadena, A.; Ráez-Bravo, A.; González-Crespo, C.; Espunyes, J.; Lavín, S.; López-Olvera, J.R. Wild boar in the city: Phenotypic responses to urbanisation. Sci. Total Environ. 2021, 773, 145593. [Google Scholar] [CrossRef]
- Conejero, C.; Castillo-Contreras, R.; González-Crespo, C.; Serrano, E.; Mentaberre, G.; Lavín, S.; López-Olvera, J.R. Past experiences drive citizen perception of wild boar in urban areas. Mamm. Biol. 2019, 96, 68–72. [Google Scholar] [CrossRef]
- Castillo-Contreras, R.; Magen, L.; Birtles, R.; Varela-Castro, L.; Hall, J.L.; Conejero, C.; Aguilar, X.F.; Colom-Cadena, A.; Lavín, S.; Mentaberre, G.; et al. Ticks on wild boar in the metropolitan area of Barcelona (Spain) are infected with spotted fever group rickettsiae. Transbound. Emerg. Dis. 2021, 68, 3223–3231. [Google Scholar] [CrossRef]
- Darwich, L.; Seminati, C.; López-Olvera, J.R.; Vidal, A.; Aguirre, L.; Cerdá, M.; Garcias, B.; Valldeperes, M.; Castillo-Contreras, R.; Migura-Garcia, L.; et al. Detection of beta-lactam-resistant Escherichia coli and toxigenic Clostridioides difficile strains in wild boars foraging in an anthropization gradient. Animals 2021, 11, 1585. [Google Scholar] [CrossRef] [PubMed]
- De Jong, A.; Thomas, V.; Simjee, S.; Godinho, K.; Schiessl, B.; Klein, U.; Butty, P.; Vallé, M.; Marion, H.; Shryock, T.R. Pan-European monitoring of susceptibility to human-use antimicrobial agents in enteric bacteria isolated from healthy food-producing animals. J. Antimicrob. Chemother. 2012, 67, 638–665. [Google Scholar] [CrossRef] [PubMed]
- De Jong, A.; Simjee, S.; El Garch, F.; Moyaert, H.; Rose, M.; Youala, M.; Dry, M. Antimicrobial susceptibility of enterococci recovered from healthy cattle, pigs and chickens in nine EU countries (EASSA Study) to critically important antibiotics. Vet. Microbiol. 2018, 216, 168–175. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).