
Effects of Acute High-Temperature Stress on Physical Responses of Yellowfin Tuna (Thunnus albacares)



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Experiment Design and Sample Collection
2.2. Enzyme Activity Measurement
2.3. Calculations and Statistical Analysis
3. Results
3.1. Changes in Serum Indexes of Juvenile Yellowfin Tuna under an Acute Temperature Rise
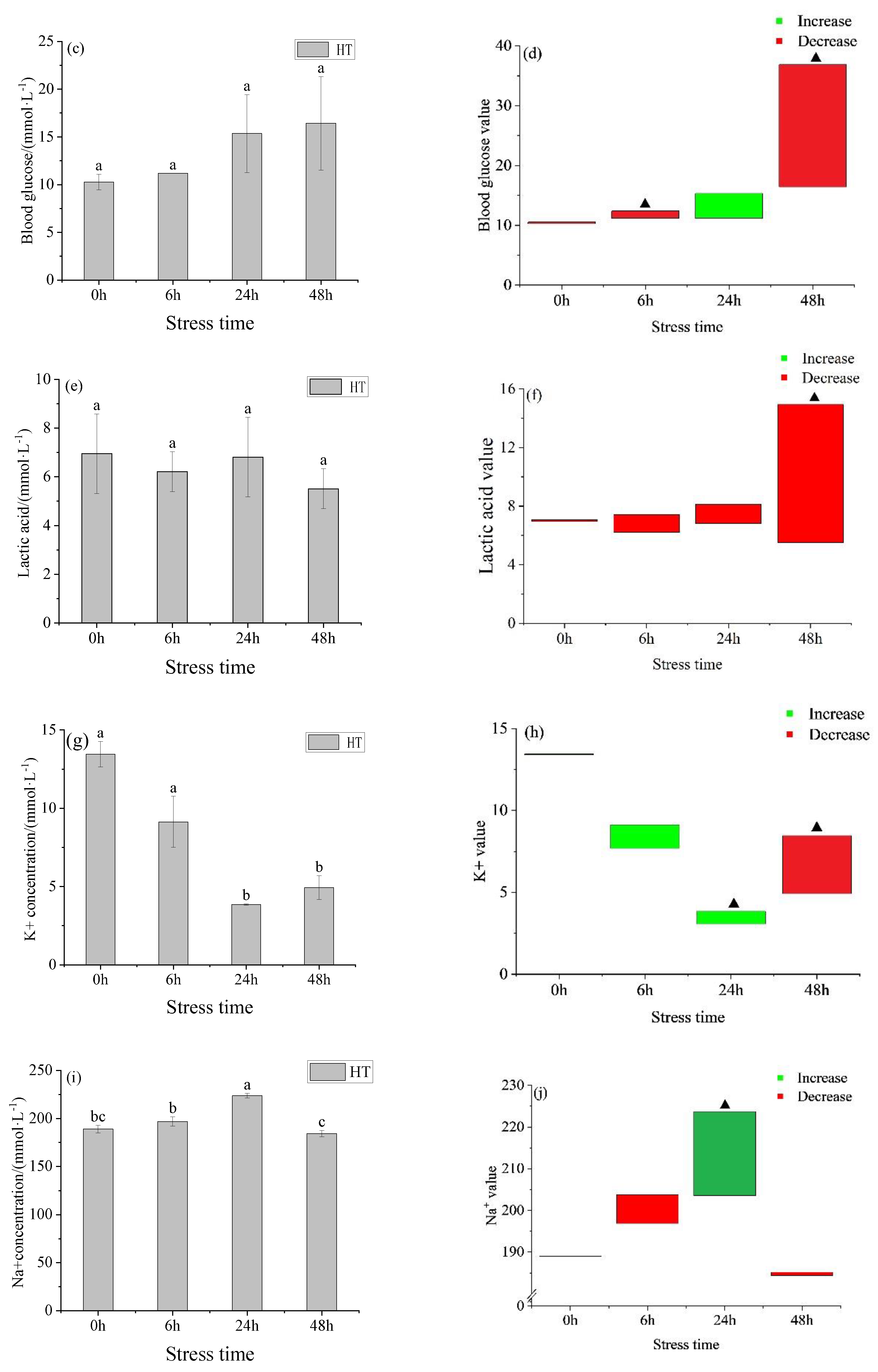
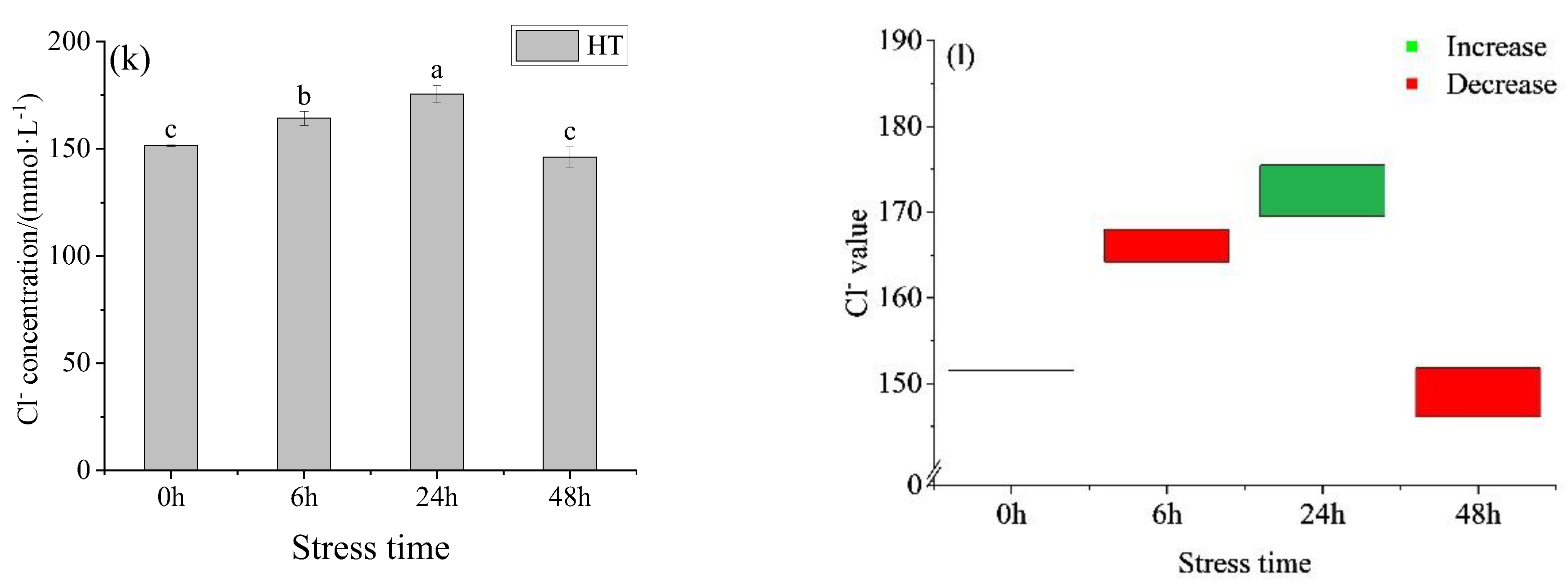
3.1.1. Changes in Ion Concentration, Osmotic Pressure, Blood Glucose, and Lactic Acid in the Serum of Juvenile Yellowfin Tuna under an Acute Temperature Rise
3.1.2. Changes in Serum Cortisol in Juvenile Yellowfin Tuna under an Acute Temperature Rise
3.1.3. Changes of Metabolic Indexes in Serum of Juvenile Yellowfin Tuna under an Acute Temperature Rise
3.1.4. Changes of Immune Indexes in Serum of Juvenile Yellowfin Tuna under an Acute Temperature Rise
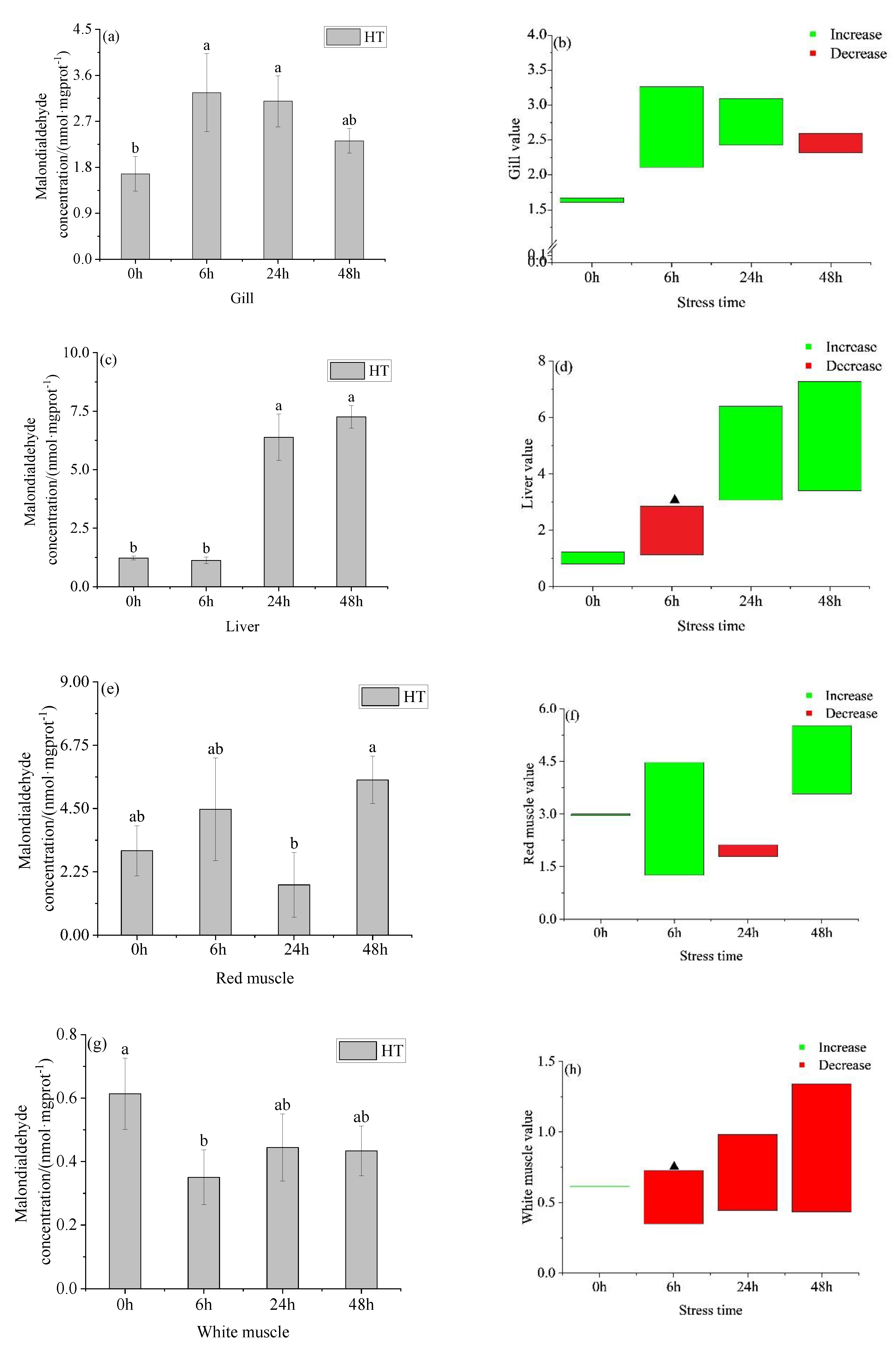
3.2. Changes in Oxidative Stress Parameters in Organs of Juvenile Yellowfin Tuna under an Acute Temperature Rise
3.3. Changes in Immune Indexes in the Liver of Juvenile Yellowfin Tuna under an Acute Temperature Rise
4. Discussion
4.1. Changes in Serum Indexes of Juvenile Yellowfin Tuna under an Acute Temperature Rise
4.1.1. Changes in Ion Concentration, Osmotic Pressure, Blood Glucose, Lactic Acid, and Cortisol in Serum of Juvenile Yellowfin Tuna under an Acute Temperature Rise
4.1.2. Changes of Metabolic Indexes in Serum of Juvenile Yellowfin Tuna under an Acute Temperature Rise
4.2. Changes in Oxidative Stress Parameters and Immune Indexes of Juvenile Yellowfin Tuna under an Acute Temperature Rise
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Joy, S.; Alikunju, A.P. Oxidative stress and antioxidant defense responses of Etroplus suratensis to acute temperature fluctuations. J. Therm. Biol. 2017, 70, 20–26. [Google Scholar] [CrossRef] [PubMed]
- Leggatt, R.A.; Brauner, C.J. Effects of acclimation and incubation temperature on the glutathione antioxidant system in killifish and RTH-149 cells. Comp. Biochem. Physiol. Mol. Integr. Physiol. 2007, 146, 317–326. [Google Scholar] [CrossRef] [PubMed]
- Maltar-Strmecki, N.; Ljubic-Beer, B. Effect of the gamma radiation on histamine production, lipid peroxidation and antioxidant parameters during storage at two different temperatures in sardine (Sardina pilchardus). Food Control 2013, 34, 132–137. [Google Scholar] [CrossRef]
- Li, Z.H.; Li, P. Regulation of glutathione-dependent antioxidant defense system of grass carp Ctenopharyngodon idellaunder the combined stress of mercury and temperature. Environ. Sci. Pollut. Res. 2020, 28, 1689–1696. [Google Scholar] [CrossRef]
- Clotfelter, E.D.; Lapidus, S.J.H. The effects of temperature and dissolved oxygen on antioxidant defences and oxidative damage in the fathead minnow Pimephales promelas. J. Fish Biol. 2013, 82, 1086–1092. [Google Scholar] [CrossRef]
- Wen, X.; Chu, P. Combined effects of low temperature and salinity on the immune response, antioxidant capacity and lipid metabolism in the pufferfish (Takifugu fasciatus). Aquaculture 2020, 531, 735866. [Google Scholar] [CrossRef]
- Pereira, L.A.L.; Amanajas, R.D. Health of the Amazonian fish tambaqui (Colossoma macropomum): Effects of prolonged photoperiod and high temperature. Aquaculture 2021, 541, 736836. [Google Scholar] [CrossRef]
- Ainsworth, A.J.; Dexiang, C. Effect of temperature on the immune system of channel catfish (Ictalurus punctatus)—I. Leucocyte distribution and phagocyte function in the anterior kidney at 10 degrees C. Comparative biochemistry and physiology. Comp. Physiol. 1991, 100, 907–912. [Google Scholar]
- Zapata, A.G.; Varas, A. Seasonal variations in the immune system of lower vertebrates. Immunol. Today 1992, 13, 142–147. [Google Scholar] [CrossRef]
- Boveris, A.; Chance, B. The mitochondrial generation of hydrogen peroxide. General properties and effect of hyperbaric oxygen. Biochem. J. 1973, 134, 707–716. [Google Scholar] [CrossRef]
- Rubbo, H.; Radi, R. Nitric oxide regulation of superoxide and peroxynitrite-dependent lipid peroxidation. Formation of novel nitrogen-containing oxidized lipid derivatives. J. Biol. Chem. 1994, 269, 26066–26075. [Google Scholar] [CrossRef] [PubMed]
- Volkoff, H.; Ronnestad, I. Effects of temperature on feeding and digestive processes in fish. Temperature 2020, 7, 307–320. [Google Scholar] [CrossRef] [PubMed]
- Davis, K.B. Temperature affects physiological stress responses to acute confinement in sunshine bass (Morone chrysops x Morone saxatilis). Comp. Biochem. Physiol. Mol. Integr. Physiol. 2004, 139, 433–440. [Google Scholar] [CrossRef] [PubMed]
- Mommsen, T.P.; Vijayan, M.M. Cortisol in teleosts: Dynamics, mechanisms of action, and metabolic regulation. Rev. Fish Biol. Fish. 1999, 9, 211–268. [Google Scholar] [CrossRef]
- Iversen, M.; Finstad, B. Stress responses in Atlantic salmon (Salmo salar L.) smolts during commercial well boat transports, and effects on survival after transfer to sea. Aquaculture 2005, 243, 373–382. [Google Scholar] [CrossRef]
- Liu, H.Y.; Fu, Z.Y. Effect of Transport Density on Greater Amberjack (Seriola dumerili) Stress, Metabolism, Antioxidant Capacity and Immunity. Front. Mar. Sci. 2020, 9, 931816. [Google Scholar] [CrossRef]
- Heink, A.E.; Parrish, A.N. Oxidative stress among SOD-1 genotypes in rainbow trout (Oncorhynchus mykiss). Aquat. Toxicol. 2014, 144, 75–82. [Google Scholar] [CrossRef]
- Garcia, D.; Lima, D. Decreased malondialdehyde levels in fish (Astyanax altiparanae) exposed to diesel: Evidence of metabolism by aldehyde dehydrogenase in the liver and excretion in water. Ecotoxicol. Environ. Saf. 2020, 190, 110107. [Google Scholar] [CrossRef]
- Ale, A.; Bacchetta, C. Low temperature stress in a cultured fish (Piaractus mesopotamicus) fed with Pyropia columbina red seaweed-supplemented diet. Fish Physiol. Biochem. 2021, 47, 829–839. [Google Scholar] [CrossRef]
- Kim, J.H.; Park, H.J. Changes in Hematological Parameters and Heat Shock Proteins in Juvenile Sablefish Depending on Water Temperature Stress. J. Aquat. Anim. Health 2019, 31, 147–153. [Google Scholar] [CrossRef]
- Nakamura, Y.N.; Ando, M. Changes of proximate and fatty acid compositions of the dorsal and ventral ordinary muscles of the full-cycle cultured Pacific bluefin tuna Thunnus orientalis with the growth. Food Chem. 2007, 103, 234–241. [Google Scholar] [CrossRef]
- Pang, J.H.; Cheng, Q.Q. The sequence and organization of complete mitochondrial genome of the yellowfin tuna, Thunnus albacares (Bonnaterre, 1788). Mitochondrial DNA Part A 2016, 27, 3111–3112. [Google Scholar] [CrossRef] [PubMed]
- Jeyasekaran, G.; Arunkumar, G. Authentication of commercially important tuna species landed in Tuticorin coast of Tamil Nadu, India by SE-AFLP method. Indian J. Fish. 2018, 64, 254–259. [Google Scholar] [CrossRef][Green Version]
- Nguyen, K.Q.; Phan, H.T. Length-length, Length-weight, and Weight-weight Relationships of Yellowfin (Thunnus Albacares) and Bigeye (Thunnus Obesus) Tuna Collected from the Commercial Handlines Fisheries in the South China Sea. Thalass. Int. J. Mar. Sci. 2022, 1–7. [Google Scholar] [CrossRef]
- Shen, H.H. Study on Management System of Tuna Fishery Resources. Ph.D. Thesis, Shanghai Ocean University, Shanghai, China, 2019. [Google Scholar]
- Benetti, D.D.; Partridge, G.J. Overview on status and technological advances in tuna aquaculture around the world. In Advances in Tuna Aquaculture; Benetti, D.D., Partridge, G.J., Buentello, A., Eds.; Academic Press: Cambridge, MA, USA, 2016; pp. 1–19. [Google Scholar]
- FAO. Thunnus albacares (Bonnaterre, 1788). 2022. Available online: https://www.fao.org/fishery/en/aqspecies/2497/en (accessed on 2 November 2022).
- WCPFC. WCPO Yellowfin Tuna (Thunnus albacares) Stock Status and Management Advice. Available online: https://www.wcpfc.int/doc/02/yellowfin-tuna (accessed on 17 February 2021).
- Zhang, P.; Yang, L. Current situation and prospect of tuna and squid resources exploitation in South China Sea. South. Fish. 2010, 6, 68–74. [Google Scholar]
- Liu, H.Y.; Fu, Z.Y. The Complete Mitochondrial Genome of Pennella sp. Parasitizing Thunnus albacares. Front. Cell. Infect. Microbiol. 2022, 12, 945152. [Google Scholar] [CrossRef] [PubMed]
- Schaefer, K.M.; Fuller, D.W. Movements, behavior, and habitat utilization of yellowfin tuna (Thunnus albacares) in the northeastern Pacific Ocean, ascertained through archival tag data. Mar. Biol. 2007, 152, 503–525. [Google Scholar] [CrossRef]
- Aoki, Y.; Aoki, A. Physiological and behavioural thermoregulation of juvenile yellowfin tuna Thunnus albacares in subtropical waters. Mar. Biol. 2020, 167, 1–14. [Google Scholar] [CrossRef]
- Kim, Y.S.; Delgado, D.I. Effect of temperature and salinity on hatching and larval survival of yellowfin tuna Thunnus albacares. Fish. Sci. 2015, 81, 891–897. [Google Scholar] [CrossRef]
- Woo, K.J.; Jeong, S.-H. Monitoring of Pathogens in Cultured Fish of Korea for the Summer Period from 2000 to 2006. J. Fish Pathol. 2006, 19, 207–214. [Google Scholar]
- Nordlie, F.G. Influence of environmental temperature on plasma ionic and osmotic concentrations in Mugil cephalus Lin. Comp. Biochem. Physiol. Comp. Physiol. 1976, 55, 379–381. [Google Scholar] [CrossRef] [PubMed]
- Rasio, E.A.; Bendayan, M.; Goresky, C.A. Effect of temperature change on the permeability of eel rete capillaries. Circ. Res. 1992, 70, 272–284. [Google Scholar] [CrossRef] [PubMed]
- Shahjahan, M.; Khatun, M.S. Nuclear and Cellular Abnormalities of Erythrocytes in Response to Thermal Stress in Common Carp Cyprinus carpio. Front. Physiol. 2020, 11, 543. [Google Scholar] [CrossRef]
- Costas, B.; Aragao, C. Different environmental temperatures affect amino acid metabolism in the eurytherm teleost Senegalese sole (Solea senegalensis Kaup, 1858) as indicated by changes in plasma metabolites. Amino Acids 2012, 43, 327–335. [Google Scholar] [CrossRef] [PubMed]
- Fan, X.P.; Qin, X.M. Effects of temperature on metabolism function and muscle quality of grouper during process of keeping alive with water. Trans. Chin. Soc. Agric. Eng. 2018, 24, 241–248. [Google Scholar]
- Arends, R.J.; van der Gaag, R. Differential expression of two pro-opiomelanocortin mRNAs during temperature stress in common carp (Cyprinus carpio L.). J. Endocrinol. 1998, 159, 85–91. [Google Scholar] [CrossRef] [PubMed]
- Metz, J.R.; van den Burg, E.H. Regulation of branchial Na+/K+-ATPase in common carp Cyptinus carpio L. acclimated to different temperatures. J. Exp. Biol. 2003, 206, 2273–2280. [Google Scholar] [CrossRef]
- Choi, C.Y.; Min, B.H. Molecular cloning of PEPCK and stress response of black porgy (Acanthopagrus schlegeli) to increased temperature in freshwater and seawater. Gen. Comp. Endocrinol. 2007, 152, 47–53. [Google Scholar] [CrossRef]
- Roychowdhury, P.; Aftabuddin, M. Thermal stress altered growth performance and metabolism and induced anaemia and liver disorder in Labeo rohita. Aquac. Res. 2020, 51, 1406–1414. [Google Scholar] [CrossRef]
- Hoseinifar, S.H.; Roosta, Z. The effects of Lactobacillus acidophilus as feed supplement on skin mucosal immune parameters, intestinal microbiota, stress resistance and growth performance of black swordtail (Xiphophorus helleri). Fish Shellfish Immunol. 2015, 42, 533–538. [Google Scholar] [CrossRef]
- Silva, M.J.D.; da Costa, F.F.B. Biological responses of Neotropical freshwater fish Lophiosilurus alexandri exposed to ammonia and nitrite. Sci. Total Environ. 2018, 616, 1566–1575. [Google Scholar] [CrossRef] [PubMed]
- Li, S.W.; Liu, Y.J. Physiological responses to heat stress in the liver of rainbow trout (Oncorhynchus mykiss) revealed by UPLC-QTOF-MS metabolomics and biochemical assays. Ecotoxicol. Environ. Saf. 2020, 242, 113949. [Google Scholar] [CrossRef] [PubMed]
- Cheng, C.H.; Guo, Z.X. Effect of dietary astaxanthin on the growth performance, non-specific immunity, and antioxidant capacity of pufferfish (Takifugu obscurus) under high temperature stress. Fish Physiol. Biochem. 2018, 44, 209–218. [Google Scholar] [CrossRef] [PubMed]
- Lushchak, V.I.; Bagnyukova, T.V. Temperature increase results in oxidative stress in goldfish tissues. 2. Antioxidant and associated enzymes. Comp. Biochem. Physiol. Toxicol. Pharmacol. 2006, 143, 36–41. [Google Scholar] [CrossRef]
- Dawood, M.A.O.; Alkafafy, M. The antioxidant responses of gills, intestines and livers and blood immunity of common carp (Cyprinus carpio) exposed to salinity and temperature stressors. Fish Physiol. Biochem. 2022, 48, 397–408. [Google Scholar] [CrossRef] [PubMed]
- Duran, A.; Talas, Z.S. Biochemical changes and sensory assessment on tissues of carp (Cyprinus carpio, Linnaeus 1758) during sale conditions. Fish Physiol. Biochem. 2009, 35, 709–714. [Google Scholar] [CrossRef]
- Phrompanya, P.; Panase, P. Histopathology and oxidative stress responses of Nile tilapia Oreochromis niloticus exposed to temperature shocks. Fish. Sci. 2021, 87, 491–502. [Google Scholar] [CrossRef]
- Wang, B.; Ma, G.X. Effects of Different Temperatures on the Antibacterial, Immune and Growth Performance of Crucian Carp Epidermal Mucus. Fishes 2022, 6, 66. [Google Scholar] [CrossRef]
- Huang, J.F.; Xu, Q.Y. Effects of temperature and dietary protein on gene expression of Hsp70 and Wap65 and immunity of juvenile mirror carp (Cyprinus carpio). Aquac. Res. 2015, 46, 2776–2788. [Google Scholar] [CrossRef]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, H.; Fu, Z.; Yu, G.; Ma, Z.; Zong, H. Effects of Acute High-Temperature Stress on Physical Responses of Yellowfin Tuna (Thunnus albacares). J. Mar. Sci. Eng. 2022, 10, 1857. https://doi.org/10.3390/jmse10121857
Liu H, Fu Z, Yu G, Ma Z, Zong H. Effects of Acute High-Temperature Stress on Physical Responses of Yellowfin Tuna (Thunnus albacares). Journal of Marine Science and Engineering. 2022; 10(12):1857. https://doi.org/10.3390/jmse10121857
Chicago/Turabian StyleLiu, Hongyan, Zhengyi Fu, Gang Yu, Zhenhua Ma, and Humin Zong. 2022. "Effects of Acute High-Temperature Stress on Physical Responses of Yellowfin Tuna (Thunnus albacares)" Journal of Marine Science and Engineering 10, no. 12: 1857. https://doi.org/10.3390/jmse10121857
APA StyleLiu, H., Fu, Z., Yu, G., Ma, Z., & Zong, H. (2022). Effects of Acute High-Temperature Stress on Physical Responses of Yellowfin Tuna (Thunnus albacares). Journal of Marine Science and Engineering, 10(12), 1857. https://doi.org/10.3390/jmse10121857